Molecular and Biochemical Mechanisms of Scutellum Color Variation in Bactrocera dorsalis Adults (Diptera: Tephritidae)

 ,
,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Adult Trapping
2.2. Sample Preparation
2.3. Genome Sequencing
2.4. SNP Detection
2.5. Principal Component and Selection Sweep Analysis
2.6. Gene Annotation and Enrichment Analysis
2.7. Determination of β-Carotene, Cytochrome P450 and Melanin
2.8. Data Analyses
3. Results
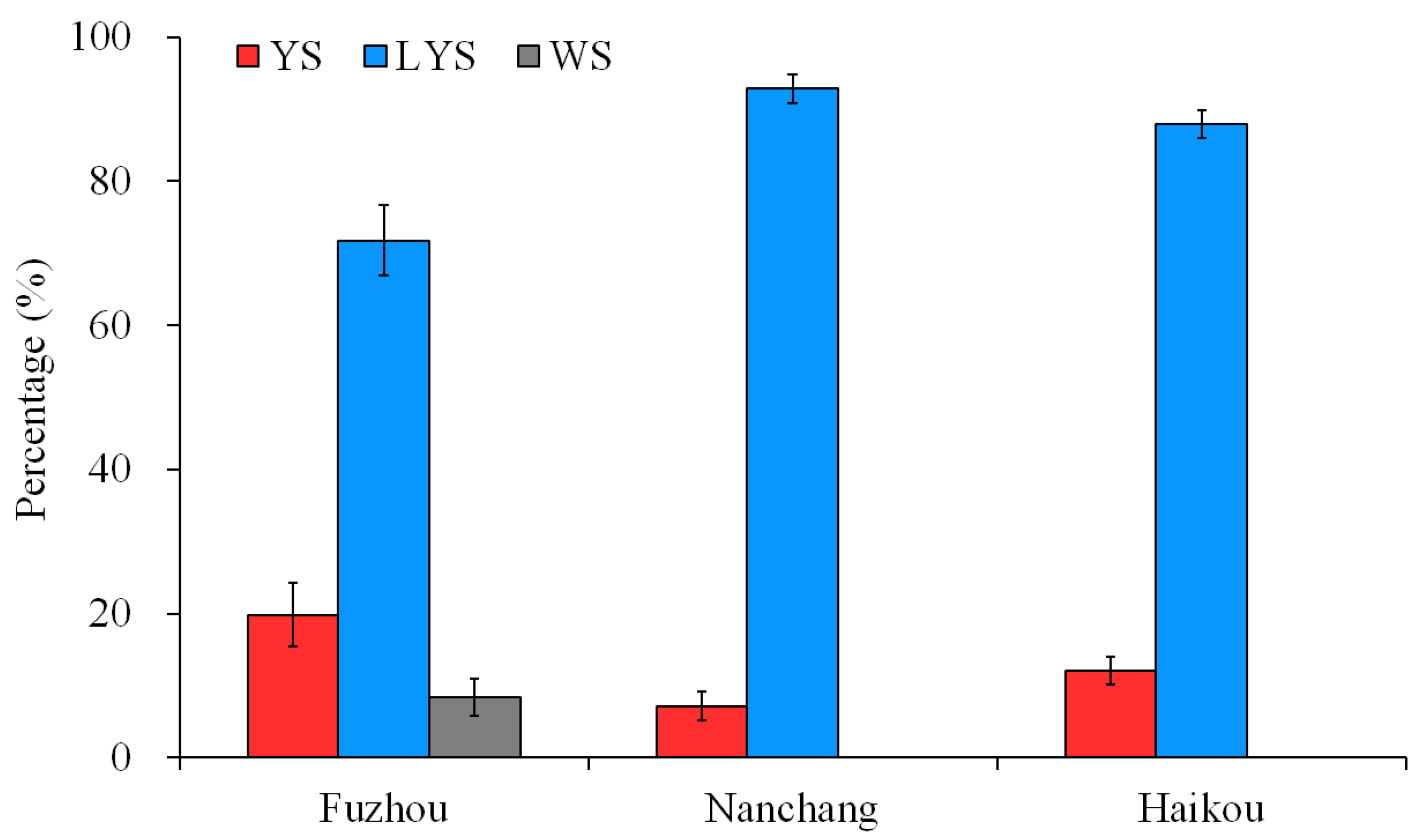
3.1. Color Variation of B. dorsalis Scutella
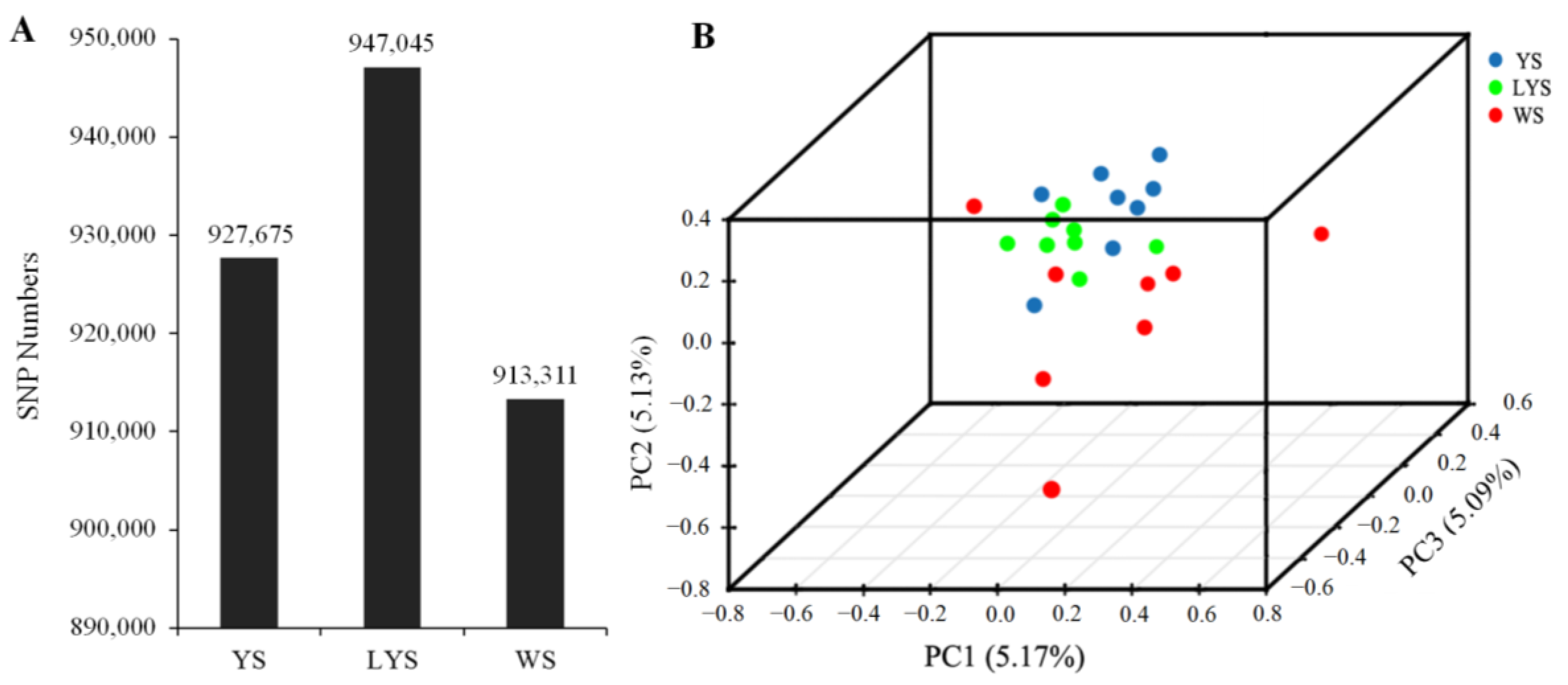
3.2. Genome Data of Three B. dorsalis Races
3.3. SNP Annotation of Genome Data
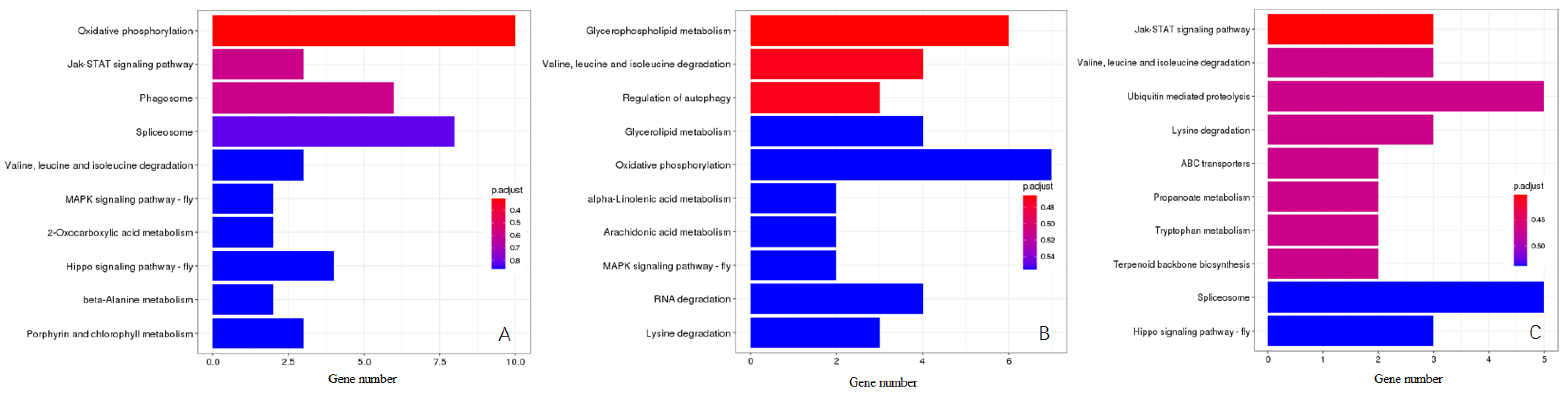
3.4. KEGG Pathway Enrichment Analysis
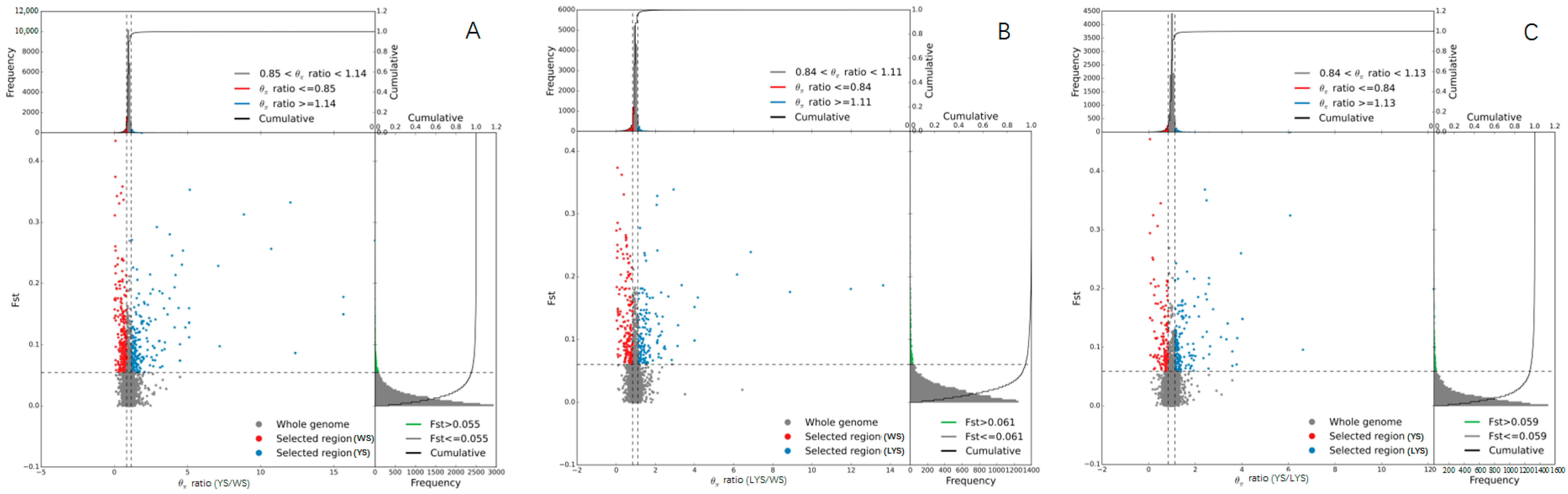
3.5. Differential Genes Between Each Two B. dorsalis Races
3.6. Genome-Wide Association Analysis Between Each Two B. dorsalis Races
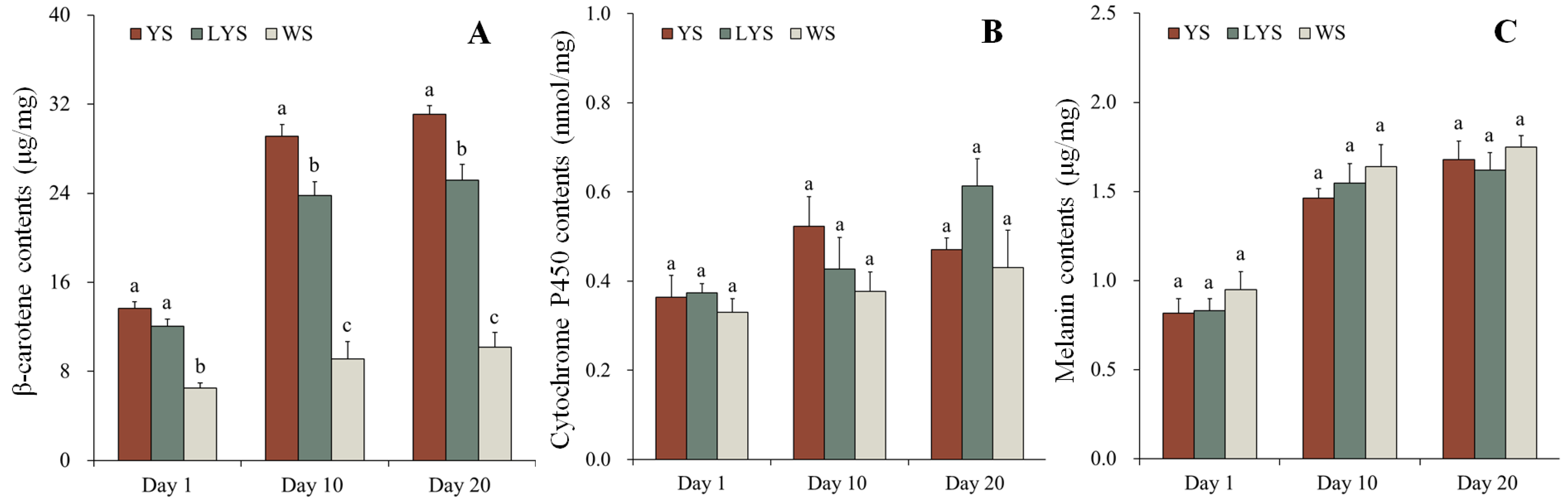
3.7. Pigment Contents
3.7.1. β-Carotene
3.7.2. Cytochrome P450
3.7.3. Melanin
4. Discussion
4.1. Ecological Differences Among the Three B. dorsalis Races
4.2. Genetic Relationship Among the Three B. dorsalis Races
4.3. Molecular Correlates of Scutellum Color Variation
4.4. Physiological Correlates of Scutellum Color Variation
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Nishida, T.; Vargas, R.I. Intraspecific variation in the wild male population, and its potential value in the mass rearing program, of the Oriental fruit fly, Bactrocera dorsalis (Hendel) (Diptera: Tephritidae). Proc. Hawaii. Entomol. Soc. 1992, 31, 83–90. [Google Scholar]
- Huo, K.K.; Zheng, Z.M. Variation of body color in Episyrphus balteatus. Entomol. Knowl. 2003, 40, 529–534. [Google Scholar]
- Bezzerides, A.L.; Mc Graw, K.J.; Parker, R.S.; Husseini, J. Elytra color as a signal of chemical defense in the Asian ladybird beetle Harmonia axyridis. Behav. Ecol. Sociobiol. 2007, 61, 1401–1408. [Google Scholar] [CrossRef]
- Gautier, M.; Yamaguchi, J.; Foucaud, J.; Loiseau, A.; Prud’homme, B. The genomic basis of color pattern polymorphism in the Harlequin ladybird. Curr. Biol. 2018, 28, 3296–3302. [Google Scholar] [CrossRef]
- Huo, K.K.; Zhang, H.J. Vriation of body color in Eristalis tenax. Sichuan J. Zool. 2005, 24, 451–454. [Google Scholar]
- Lozier, J.D.; Jackson, J.M.; Dillon, M.E.; Strange, J.P. Population genomics of divergence among extreme and intermediate color forms in a polymorphic insect. Ecol. Evol. 2016, 6, 1075–1091. [Google Scholar] [CrossRef]
- Dai, X.Y.; Wang, Y.; Liu, Y.; Wang, R.J.; Su, L.; Yin, Z.J.; Zhao, S.; Chen, H.; Zheng, L.; Dong, X.L.; et al. Molecular correlates of diapause in Aphidoletes aphidimyza. Insects 2024, 15, 299. [Google Scholar] [CrossRef]
- Lozier, J.D.; Strange, J.P.; Koch, J.B. Landscape heterogeneity predicts gene flow in a widespread polymorphic bumble bee, Bombus bifarius (Hymenoptera: Apidae). Conserv. Genet. 2013, 14, 1099–1110. [Google Scholar] [CrossRef]
- Ando, T.; Matsuda, T.; Goto, K.; Hara, K.; Ito, A.; Hirata, J.; Yamaguchi, K. Repeated inversions within a pannier intron drive diversification of intraspecific colour patterns of ladybird beetles. Nat. Commun. 2018, 9, 3843. [Google Scholar] [CrossRef]
- Wu, N.R.; Wei, Y.T.; Guan, M. Transcriptome analysis of Brassica napus with different flower colors. Mol. Plant Breed. 2018, 16, 7234–7240. [Google Scholar] [CrossRef]
- Meng, Y.; Bai, Y.; Chen, D.; Ma, T.; Si, W.; Yuan, Y.; Chen, L.; Zhou, Y. Integration of transcriptome and metabolome reveals key regulatory mechanisms affecting sepal color variation in Aquilegia oxysepala. Sci. Hortic. 2024, 334, 113334. [Google Scholar] [CrossRef]
- Zhong, X.; Yang, M.; Zhang, X.; Fan, Y.; Wang, X.; Xiang, C. Comparative analysis of transcriptome and metabolome explores the underlying mechanism of pod color variation in pea (Pisum sativum L.). J. Plant Biochem. Biot. 2024, 33, 144–156. [Google Scholar] [CrossRef]
- Chen, Y.; Gong, Q.; Lai, J.; Song, M.; Long, Z. Transcriptome analysis identifies candidate genes associated with skin color variation in Triplophysa siluroides. Comp. Biochem. Phys. D 2020, 35, 100682. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.Y.; Wu, G.Q.; Wang, J.; Liu, C.L.; Qiu, L.H. Next-generation sequencing-based transcriptome analysis of Helicoverpa armigera Larvae immune-primed with Photorhabdus luminescens TT01. PLoS ONE 2017, 8, e80146. [Google Scholar] [CrossRef]
- Nijhout, H.F. Ommochrome pigmentation of the linea and rosa seasonal forms of Precis coenia (Lepidoptera: Nymphalidae). Arch. Insect Biochem. 1997, 36, 215–222. [Google Scholar] [CrossRef]
- Cromartie, R.I.T. Insect pigments. Annu. Rev. Entomol. 2002, 4, 59–76. [Google Scholar] [CrossRef]
- Sun, M.X.; Yin, H.W.; Wang, J.X.; Ling, A.P. Progress in research on insect pigments. Chin. J. Appl. Entomol. 2020, 57, 298–309. [Google Scholar]
- Blois, M.S. The melanins: Their synthesis and structure. In Photochemical and Photobiological Reviews; Smith, K.C., Ed.; Plenum Press: New York, NY, USA, 1978; pp. 115–134. [Google Scholar]
- Tseng, M.; Bevanda, C.; Bhatti, S.S.; Black, E.N.; Chang, E.; Chiang, J.; Dhaliwal, H.; Dimitriou, A.; Gong, S.Y.; Halbe, E. Effects of temperature on monarch caterpillar pigment variation in nature. Insect conservation and diversity. Insect Conserv. Diver. 2023, 16, 164–171. [Google Scholar] [CrossRef]
- Puanmanee, K.; Wongpiyasatid, A.; Sutantawong, M.; Hormchan, P. Gamma irradiation effect on guava fruit fly, Bactrocera correcta (bezzi) (Diptera: Tephritidae). Kasetsart J. (Nat. Sci.) 2010, 44, 830–836. [Google Scholar]
- Ushakova, N.A.; Dontsov, A.E.; Sakina, N.L.; Karagodin, V.P.; Bastrakov, A.I. Preparation and characterization of Alphitobius diaperinus melanin. Appl. Biochem. Micro. 2018, 54, 425–431. [Google Scholar] [CrossRef]
- Feyereisen, R. Insect P450 enzymes. Annu. Rev. Entomol. 1999, 44, 507–533. [Google Scholar] [CrossRef] [PubMed]
- Rewitz, K.F.; Connor, M.B.O.; Gilbert, L.I. Molecular evolution of the insect Halloween family of cytochrome P450s: Phylogeny, gene organization and functional conservation. Insect Biochem. Molec. 2007, 37, 741–753. [Google Scholar] [CrossRef] [PubMed]
- Xu, C.J.; Zhang, S.L. Carotenoid biosynthesis and its regulation in plants. Plant Physiol. Comm. 2000, 36, 64–70. [Google Scholar]
- Davis, A.K. Intraspecific variation in wing colour is related to larval energy reserves in monarch butterflies (Danaus plexippus). Physiol. Eetomol. 2014, 39, 247–253. [Google Scholar] [CrossRef]
- Willadsen, P.C. Aculeate hymenopterans as aposematic and mimetic models. Front. Ecol. Evol. 2022, 10, 827319. [Google Scholar] [CrossRef]
- Yumnam, T.; Banerjee, B.; Kodandaramaiah, U. Pupal colour plasticity in the butterfly Catopsilia pomona (Lepidoptera: Pieridae). Biol. J. Linn. Soc. 2021, 134, 331–341. [Google Scholar] [CrossRef]
- Davis, A.K.; Cope, N.; Smith, A.; Solensky, M.J. Behaviour: Wing color predicts future mating success in male monarch butterflies. Ann. Entomol. Soc. Am. 2007, 100, 339–344. [Google Scholar] [CrossRef]
- Mello, N.D.; Sanchez, L.G.F.; Gawryszewski, F.M. Spatio-temporal colour variation of arthropods and their environment. Evol. Ecol. 2021, 36, 117–133. [Google Scholar] [CrossRef]
- Ellers, J.; Boggs, C.L. The evolution of wing color: Male mate choice opposes adaptive wing color divergence in colias butterflies. Evolution 2010, 57, 1100–1106. [Google Scholar] [CrossRef]
- Li, X.; Yang, H.; Wang, T.; Wang, J.; Wei, H.Y. Life history and adult dynamics of Bactrocera dorsalis in the citrus orchard of Nanchang, a subtropical area from China: Implications for a control timeline. ScienceAsia 2019, 45, 212–220. [Google Scholar] [CrossRef]
- Drew, R.A.I.; Hancock, D.L. The Bactrocera dorsalis complex of fruit flies (Diptera: Tephritidae: Dacinae) in Asia. B. Entomol. Res. 1994, 2, 1–68. [Google Scholar] [CrossRef]
- Aketarawong, N.; Bonizzoni, M.; Thanaphum, S.; Gomulski, L.M.; Gasperi, G.; Malacrida, A.R.; Gugliemino, C.R. Inferences on the population structure and colonization process of the invasive oriental fruit fly, Bactrocera dorsalis (Hendel). Mol. Ecol. 2010, 16, 3522–3532. [Google Scholar] [CrossRef] [PubMed]
- Drew, R.A.I.; Raghu, S. The fruit fly fauna (Diptera: Tephritidae: Dacinae) of the rainforest habitat of the Western Ghats, India. Raffles B. Zool. 2002, 50, 327–352. [Google Scholar] [CrossRef]
- Wan, X.W.; Liu, Y.H.; Zhang, B. Invasion history of the oriental fruit fly, Bactrocera dorsalis, in the Pacific-Asia region: Two main invasion routes. PLoS ONE 2012, 7, e36176. [Google Scholar] [CrossRef]
- Drew, R.A.I.; Raghu, S.; Halcoop, P. Bridging the morphological and biological species concepts: Studies on the Bactrocera dorsalis (Hendel) complex (Diptera: Tephritidae: Dacinae) in South-east Asia. Biol. J. Linn. Soc. 2008, 93, 217–226. [Google Scholar] [CrossRef]
- Drew, R.A.I.; Romig, M.C. Tropical Fruit Flies of South-East Asia: (Tephritidae: Dacinae); C.A.B. International: Wallingford, UK, 2013; p. 856. [Google Scholar]
- Schutze, M.K.; Aketarawong, N.; Amornsak, W.; Armstrong, K.F.; Augustinos, A.A.; Barr, N.; Bo, W.; Bourtzis, K.; Boykin, L.M.; Cáceres, C.; et al. Synonymization of key pest species within the Bactrocera dorsalis species complex (Diptera: 3 Tephritidae): Taxonomic changes based on 20 years of integrative morphological, molecular, cytogenetic, behavioral, and chemoecological data. Syst. Entomol. 2014, 40, 456–471. [Google Scholar] [CrossRef]
- Schutze, M.K.; Jessup, A.; Clarke, A.R. Wing shape as a potential discriminator of morphologically similar pest taxa within the Bactrocera dorsalis species complex (Diptera: Tephritidae). Bull. Entomol. Res. 2012, 102, 103–111. [Google Scholar] [CrossRef]
- Leblanc, L.; Hossain, M.A.; Khan, S.A.; San Jose, M.; Rubinoff, D. A Preliminary Survey of the Fruit Flies (Diptera: Tephritidae: Dacinae) of Bangladesh. Proc. Hawaii. Entomol. Soc. 2013, 45, 51–58. Available online: http://hdl.handle.net/10125/31004 (accessed on 15 July 2024).
- Tokushima, I.; Orankanok, W.; Tan, K.H.; Ono, H.; Nishida, R. Accumulation of phenylpropanoid and sesquiterpenoid volatiles in male rectal pheromonal glands of the guava fruit fly, Bactrocera correcta. J. Chem. Ecol. 2010, 36, 1327. [Google Scholar] [CrossRef]
- Clarke, A.R.; Armstrong, K.F.; Carmichael, A.E.; Milne, J.R.; Raghu, S.; Roderich, G.K.; Yeates, D.K. Invaisve phytophagous pests arising through a recent tropical evolutionary radiation: The Bactrocera dorsalis complex of fruit flies. Annu. Rev. Entomol. 2005, 50, 293–319. [Google Scholar] [CrossRef]
- Li, X.Z.; Wang, G.L.; Wang, C.L.; Li, W.J.; Lu, T.; Ge, Y.G.; Xu, C.K.; Zhong, X.; Wang, J.G.; Yang, H.Y. Long-term monitoring of Bactrocera and Zeugodacus spp. (Diptera: Tephritidae) in China and evaluation of different control methods for Bactrocera dorsalis (Hendel). Crop Prot. 2024, 182, 106708. [Google Scholar] [CrossRef]
- Lin, M.G.; Zhang, Y.N.; Wang, X.J.; Zeng, L. Trapping effect of methyl eugenol and protein bait on Bactrocera dorsalis (Hendel) in Hainan Carambola Orchard. N. Hortic. 2013, 16, 131–134. [Google Scholar]
- Simão, F.A.; Waterhouse, R.M.; Ioannidis, P.; Kriventseva, E.V.; Zdobnov, E.M. BUSCO: Assessing genome assembly and annotation completeness with single-copy orthologs. Bioinformatics 2015, 31, 3210–3212. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Andongma, A.A.; Dong, Y.; Chen, Z.; Xu, P.; Ren, X.; Krosch, M.N.; Clarke, A.R.; Niu, C. Rh6 gene modulates the visual mechanism of host utilization in fruit fly Bactrocera minax. Pest Manag. Sci. 2019, 75, 1621–1629. [Google Scholar] [CrossRef]
- Mao, X.; Cai, T.; Olyarchuk, J.G.; Wei, L. Automated genome annotation and pathway identification using the KEGG Orthology (KO) as a controlled vocabulary. Bioinformatics 2005, 21, 3787–3793. [Google Scholar] [CrossRef]
- Cingolani, P.; Platts, A.; Wang, L.L.; Coon, M.; Nguyen, T.; Wang, L.; Land, S.J.; Lu, X.Y.; Ruden, D.M. A program for annotating and predicting the effects of single nucleotide polymorphisms, SnpEff: SNPs in the genome of Drosophila melanogaster strain w1118; iso-2; iso-3. Fly 2012, 6, 80–92. [Google Scholar] [CrossRef]
- Li, T.; Liu, M.; Liu, Z.H.; Chen, A.L.; Liao, P.F.; Yang, H.; Dong, Z.P. Correlation between chlorfenapyr tolerance and contents of cytochrome P450 and b5 in silkworm Bombyx mori. Sci. Seric. 2017, 43, 0436–0441. [Google Scholar]
- Drew, R.A.I.; Tsuruta, K.; White, I.M. A new species of pest fruit fly (Diptera: Tephritidae: Dacinae) from Sri Lanka and Africa. Afr. Entomol. 2005, 13, 149–154. [Google Scholar] [CrossRef]
- Leblanc, L.; Jose, M.S.; Barr, N.; Rubinoff, D. A phylogenetic assessment of the polyphyletic nature and intraspecific color polymorphism in the Bactrocera dorsalis complex (Diptera, Tephritidae). ZooKeys 2015, 540, 339–367. [Google Scholar] [CrossRef]
- Boykin, L.M.; Schutze, M.K.; Krosch, M.N.; Chomič, A.; Chapman, T.A.; Englezou, A.; Armstrong, K.F.; Clarke, A.R.; Hailstones, D.; Cameron, S.L. Multi-gene phylogenetic analysis of southeast Asian pest members of the Bactrocera dorsalis species complex (Diptera: Tephritidae) does not support current taxonomy. J. Appl. Entomol. 2013, 138, 235–253. [Google Scholar] [CrossRef]
- Colgan, T.J.; Carolan, J.C.; Bridgett, S.J.; Sumner, S.; Blaxter, M.L.; Brown, M.J.F. Polyphenism in social insects: Insights from a transcriptome-wide analysis of gene expression in the life stages of the key pollinator, Bombus terrestris. BMC Genom. 2011, 12, 623–642. [Google Scholar] [CrossRef] [PubMed]
- Han, P.; Wang, X.; Niu, C.Y.; Dong, Y.C.; Zhu, J.Q.; Nicolas, N. Population dynamics, phenology, and overwintering of Bactrocera dorsalis (Diptera: Tephritidae) in Hubei Province, China. J. Pest Sci. 2011, 84, 289–295. [Google Scholar] [CrossRef]
- Satarkar, V.R.; Krishnamurthy, S.V.; Faleiro, J.R.; Verghese, A. Spatial distribution of major Bactrocera fruit flies attracted to methyl eugenol in different ecological zones of Goa, India. Int. J. Trop. Insect Sci. 2009, 29, 195–201. [Google Scholar] [CrossRef]
- Drew, R.A.I.; Hancock, D.L. Phylogeny of the Tribe Dacini (Dacinae) based on morphological, distributional, and biological data. In Fruit Flies (Tephritidae): Phylogeny and Evolution of Behavior; Aluja, M., Norrbom, A.L., Eds.; CRC Press: Boca Raton, FL, USA, 2000; pp. 491–504. [Google Scholar] [CrossRef]
- Wan, X.W.; Nardi, F.; Zhang, B.; Liu, Y.H. The oriental fruit fly, Bactrocera dorsalis, in China: Origin and gradual inland range expansion associated with population growth. PLoS ONE 2011, 6, e25238. [Google Scholar] [CrossRef] [PubMed]
- Gokulanathan, A.; Mo, H.H.; Park, Y. Insights on reproduction-related genes in the striped fruit fly, Zeugodacus scutellata (Hendel) (Diptera: Tephritidae). Arch. Insect Biochem. Physiol. 2024, 115, e22064. [Google Scholar] [CrossRef]
- Li, W.J.; Song, Y.J.; Han, H.L.; Xu, H.Q.; Wei, D.; Smagghe, G.; Wang, J.J. Genome-wide analysis of long non-coding RNAs in adult tissues of the melon fly, Zeugodacus cucurbitae (Coquillett). BMC Genom. 2020, 21, 600. [Google Scholar] [CrossRef]
- Grubbs, N.; Haas, S.; Beeman, R.W.; Lorenzen, M.D. The ABCsof eye color in Tribolium castaneum: Orthologs of the Drosophilawhite, scarlet and brown genes. Genetics 2015, 199, 749–759. [Google Scholar] [CrossRef]
- Yi, C.; Zheng, C.; Zeng, L.; Xu, Y. High genetic diversity in the offshore island populations of the tephritid fruit fly Bactrocera dorsalis. BMC Ecol. 2016, 16, 46. [Google Scholar] [CrossRef]
- Grasso, F.; Coppola, M.; Carbone, F.; Baldoni, L.; Alagna, F.; Perrotta, G.; Pérez-Pulido, A.J.; Garonna, A.; Facella, P.; Daddiego, L.; et al. The transcriptional response to the olive fruit fly (Bactrocera oleae) reveals extended differences between tolerant and susceptible olive (Olea europaea L.) varieties. PLoS ONE 2017, 12, e0183050. [Google Scholar] [CrossRef]
- Morooka, S.; Tanaka, Y.; Tojo, S. Genetic variation in developmental times among four pure lines exhibiting specific wing form and body color in the brown planthopper, Nilaparvata lugens (Hemiptera: Auchenorrhyncha). Appl. Entomol. Zool. 2011, 46, 229–237. [Google Scholar] [CrossRef]
- Riccardo, P.; Kapan, D.D.; Counterman, B.A.; Karla, M.; Lindstrom, D.P.; Reed, R.D.; Frederik, N.H.; Tomas, H.; Owen, M.M.W.; Casper, B. Multi-allelic major effect genes interact with minor effect QTLs to control adaptive color pattern variation in Heliconius erato. PLoS ONE 2013, 8, e57033. [Google Scholar] [CrossRef]
- Wilson, D.F.; Vinogradov, S.A. Mitochondrial cytochrome c oxidase: Mechanism of action and role in regulating oxidative phosphorylation. J. Appl. Physiol. 2014, 117, 1431–1439. [Google Scholar] [CrossRef] [PubMed]
- Bankar, T.N.; Dar, M.A.; Pandit, R.S. Diversity of pigments in insects, their synthesis and economic value for various industries. Res. Ecol. 2021, 3, 10–17. [Google Scholar] [CrossRef]
- Josien, M.L.; Fuson, N.; Lebas, J.M.; Gregory, T.M. An infrared spectroscopic study of the carbonyl stretching frequency in a group of ortho and para quinones. J. Chem. Phys. 1953, 21, 331–340. [Google Scholar] [CrossRef]
- Roth, L.M.; Stay, B. The occurrence of para-quinones in some arthropods, with emphasis on the quinone-secreting tracheal glands of Diploptera punctata (Blattaria). J. Insect Physiol. 1958, 1, 305–318. [Google Scholar] [CrossRef]
- Parkash, J.; Robblee, J.H.; Agnew, J.; Gibbs, E.; Collings, P.; Pasternack, R.F.; de Paula, J.C. Depolarized resonance light scattering by porphyrin and chlorophyll a aggregates. Biophys. J. 1998, 74, 2089–2099. [Google Scholar] [CrossRef]
- Sobotka, R.; Duhring, U.; Komenda, J.; Peter, E.; Gardian, Z.; Tichy, M.; Grimm, B.; Willde, A. Importance of the cyanobacterial gun4 protein for chlorophyll metabolism and assembly of photosynthetic complexes. J. Biol. Chem. 2008, 283, 25794–25802. [Google Scholar] [CrossRef]
- Weiss, M.J. Neuronal connections and the function of the corpora pedunculata in the brain of the American cockroach, Periplaneta americana (L.). J. Morphol. 1974, 142, 21–69. [Google Scholar] [CrossRef]
- Lanzrein, B.; Wilhelm, R.; Riechsteiner, R. Differential degradation of racemic and 10r-juvenile hormone iii by cockroach (Nauphoeta cinerea) haemolymph and the use of lipophorin for long-term culturing of Corpora allata. J. Insect Physiol. 1993, 39, 53–63. [Google Scholar] [CrossRef]
- Ashour, M.; Wink, M.; Gershenzon, J. Biochemistry of terpenoids: Monoterpenes, sesquiterpenes and diterpenes. In Biochemistry of Plant Secondary Metabolism; Wink, M., Ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2010. [Google Scholar] [CrossRef]
- Briscoe, A.D.; Chittka, L. The evolution of colour vision in insects. Annu. Rev. Entomol. 2001, 46, 471–510. [Google Scholar] [CrossRef]
- Shamim, G.; Ranjan, S.K.; Pandey, D.M.; Ranganathan, R. Biochemistry and biosynthesis of insect pigments. Eur. J. Entomol. 2014, 111, 149–164. [Google Scholar] [CrossRef]
- Badejo, O.; Skaldina, O.; Gilev, A.; Sorvari, J. Benefits of insect colours: A review from social insect studies. Oecologia 2020, 194, 27–40. [Google Scholar] [CrossRef] [PubMed]
- Feltwell, J.; Rothschild, M. Carotenoids in thirty-eight species of Lepidoptera. J. Zool. 1974, 174, 441–465. [Google Scholar] [CrossRef]
- Rothschild, M.; Mummery, R. Carotenoids and bile pigments in danaid and swallowtail butterflies. Biol. J. Linn. Soc. 1985, 24, 1–14. [Google Scholar] [CrossRef]
- Wittkopp, P.J.; Beldade, P. Development and evolution of insect pigmentation: Genetic mechanisms and the potential consequences of pleiotrophy. Rev. Semin. Cell Dev. Biol. 2009, 20, 65–71. [Google Scholar] [CrossRef]
- French, V. Pattern formation in colour of butterfly wings. Curr. Opin. Genet. Dev. 1997, 7, 524–529. [Google Scholar] [CrossRef]
- Warren, J.T.; Petryk, A.; Marqués, G.; Parvy, J.P.; Shinoda, T.; Itoyama, K.; Kobayashi, J.; Jarcho, M.; Li, Y.; O’Connor, M.B. Phantom encodes the 25-hydroxylase of Drosophila melanogaster and Bombyx mori: A P450 enzyme critical in ecdysone biosynthesis. Insect Biochem. Mol. Biol. 2004, 34, 991–1010. [Google Scholar] [CrossRef]
- Daborn, P.J.; Yen, J.L.; Bogwitz, M.R.; Le Goff, G.; Feil, E.; Jeffers, S.; Tijet, N.; Perry, T.; Heckel, D.; Batterham, P.; et al. A single p450 allele associated with insecticide resistance in Drosophila. Science 2002, 297, 2253–2256. [Google Scholar] [CrossRef]
- Li, X.; Schuler, M.A.; Berenbaum, M.R. Jasmonate and salicylate induce expression of herbivore cytochrome P450 genes. Nature 2002, 419, 712–715. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Raw Read | Clean Read | Q20 (%) | Q30 (%) | GC (%) | Mapping Rate (%) | Sequencing Depth | Coverage (%) |
|---|---|---|---|---|---|---|---|---|
| YS1 | 16,175,193 | 16,114,275 | 98.01 | 93.77 | 37.60 | 82.96 | 10.92 | 91.32 |
| YS2 | 13,674,576 | 13,624,567 | 98.05 | 93.85 | 37.57 | 83.49 | 9.86 | 90.61 |
| YS3 | 12,807,459 | 12,753,577 | 97.88 | 93.50 | 37.84 | 82.96 | 9.75 | 89.58 |
| YS4 | 14,123,606 | 14,066,036 | 97.88 | 93.46 | 37.43 | 83.89 | 10.05 | 91.01 |
| YS5 | 13,479,907 | 13,422,541 | 97.98 | 93.73 | 37.56 | 82.62 | 9.85 | 90.69 |
| YS6 | 13,698,693 | 13,646,031 | 97.91 | 93.53 | 37.58 | 83.32 | 9.97 | 90.41 |
| YS7 | 15,930,348 | 15,869,990 | 98.02 | 93.81 | 37.58 | 83.05 | 10.66 | 91.33 |
| YS8 | 15,557,059 | 15,495,381 | 98.01 | 93.76 | 37.96 | 82.84 | 10.68 | 90.73 |
| LYS1 | 14,689,206 | 14,633,672 | 97.99 | 93.70 | 37.74 | 83.52 | 10.30 | 91.18 |
| LYS3 | 15,045,434 | 14,990,061 | 97.99 | 93.76 | 37.70 | 83.79 | 10.50 | 91.02 |
| LYS4 | 14,472,519 | 14,422,056 | 97.99 | 93.76 | 37.56 | 83.64 | 10.36 | 90.61 |
| LYS5 | 14,444,804 | 14,390,911 | 98.04 | 93.85 | 37.79 | 83.75 | 10.28 | 90.56 |
| LYS6 | 15,877,274 | 15,817,763 | 98.02 | 93.70 | 38.24 | 83.63 | 11.11 | 89.86 |
| LYS7 | 13,789,486 | 13,735,167 | 97.87 | 93.43 | 37.65 | 83.23 | 10.08 | 90.36 |
| LYS8 | 14,336,392 | 14,273,582 | 98.09 | 93.97 | 37.91 | 83.96 | 9.20 | 91.54 |
| WS1 | 15,320,793 | 15,258,254 | 97.77 | 93.17 | 37.06 | 82.41 | 10.34 | 91.02 |
| WS2 | 14,961,581 | 14,896,256 | 97.73 | 92.96 | 37.06 | 81.71 | 9.52 | 90.33 |
| WS3 | 15,680,005 | 15,620,251 | 97.80 | 93.17 | 37.06 | 83.44 | 10.68 | 91.42 |
| WS5 | 12,445,398 | 12,394,219 | 97.82 | 93.28 | 37.26 | 83.52 | 9.62 | 90.04 |
| WS6 | 14,995,683 | 14,944,984 | 98.09 | 93.91 | 37.89 | 83.81 | 10.57 | 90.75 |
| WS7 | 12,230,812 | 12,182,772 | 97.93 | 93.61 | 37.33 | 83.99 | 9.61 | 89.69 |
| WS8 | 11,640,577 | 11,588,437 | 97.68 | 93.02 | 37.35 | 83.13 | 9.22 | 90.16 |
| First Level Classification | Secondary Classification | Ko Classification | Ko Pathway | Paired Samples | Gene Numbers |
|---|---|---|---|---|---|
| Environmental information processing | Signal transduction | ko04630 | Jak-STAT signaling pathway | YS vs. WS | 3 |
| LYS vs. WS | - | ||||
| YS vs. LYS | 3 | ||||
| ko04013 | MAPK signaling pathway–fly | YS vs. WS | 2 | ||
| LYS vs. WS | 2 | ||||
| YS vs. LYS | - | ||||
| ko04391 | Hippo signaling pathway–fly | YS vs. WS | 4 | ||
| LYS vs. WS | - | ||||
| YS vs. LYS | 3 | ||||
| Metabolism | Amino acid metabolism | ko00280 | Val, Leu and Ile degradation | YS vs. WS | 3 |
| LYS vs. WS | - | ||||
| YS vs. LYS | 3 | ||||
| Other amino acid metabolism | ko00410 | Beta-Alanime metabolism | YS vs. WS | 2 | |
| LYS vs. WS | - | ||||
| YS vs. LYS | 1 | ||||
| Genetic information processing | Translation | ko03040 | Spliceosome | YS vs. WS | 8 |
| LYS vs. WS | 7 | ||||
| YS vs. LYS | 5 |
| Gene Name | Gene ID | Chromosome | Position in Chromosome | Ko Pathway | Scutellum Color |
|---|---|---|---|---|---|
| blw | 105229678 | 3 | NC_064305.1 (87342785–87345945) | Oxidative phosphorylation | YS vs. WS LYS vs. WS |
| LOC105229962 | 105229962 | 3 | NC_064305.1 (79775155–79777217) | Oxidative phosphorylation | YS vs. WS LYS vs. WS |
| sev | 105230340 | 4 | NC_064306.1 (93256–105304) | MAPK signaling pathway | LYS vs. WS |
| LOC105231837 | 105231837 | 2 | NC_064304.1 (32268170–32288201) | Jak-STAT signaling pathway | YS vs. WS YS vs. LYS |
| LOC105231839 | 105231839 | 2 | NC_064304.1 (32236821–32264955) | Jak-STAT signaling pathway | YS vs. WS YS vs. LYS |
| LOC105231827 | 105231827 | 2 | NC_064304.1 (32354094–32363461) | Hippo signaling pathway | YS vs. WS YS vs. LYS |
| Aldh7A1 | 105232622 | 5 | NC_064307.1 (4433597–4436786) | Valine, Leucine and Isoleucine degradation; Lysine degradation | YS vs. WS LYS vs. WS YS vs. LYS |
| LOC105232613 | 105232613 | 2 | NC_064304.1 (402079–435348) | Glycerophospholipid metabolism | LYS vs. WS |
| LOC105232615 | 105232615 | 2 | NC_064304.1 (467110–483000) | Porphyrin and chlorophyll metabolism | YS vs. LYS |
| Prp19 | 105229955 | 3 | NC_064305.1 (79711816–79716434) | Ubiquitin-mediated proteolysis | YS vs. LYS |
| Fancl | 105231847 | 2 | NC_064304.1 (32106230–32107938) | Ubiquitin-mediated proteolysis | YS vs. LYS |
| Ube4B | 105222370 | 1 | NC_064303.1 (107841509–107851668) | Ubiquitin-mediated proteolysis | YS vs. LYS |
| Nedd4 | 105226493 | 5 | NC_064307.1 (4497638–4502273) | Ubiquitin-mediated proteolysis | YS vs. LYS |
| Herc4 | 105226491 | 5 | NC_064307.1 (4508221–4513959) | Ubiquitin-mediated proteolysis | YS vs. LYS |
| Mdr50 | 105223845 | 3 | NC_064305.1 (8667810–8682111) | ABC Transporters | YS vs. LYS |
| Mdr65 | 105226497 | 5 | NC_064307.1 (4482003–4493223) | ABC Transporters | YS vs. LYS |
| Acat2 | 105232621 | 5 | NC_064307.1 (4436978–4438999) | Terpenoid backbone biosynthesis | YS vs. LYS |
| LOC105223844 | 105223844 | 3 | NC_064305.1 (8687891–8694379) | Terpenoid backbone biosynthesis | YS vs. LYS |
| Dhdds | 105227096 | Unplaced Scaffold | NW_026038081.1 (117795–118974) | Terpenoid backbone biosynthesis | YS vs. LYS |
| LOC105228052 | 105228052 | 3 | NC_064305.1 (905942–908404) | Biosynthesis of secondary metabolites | YS vs. WS LYS vs. WS YS vs. LYS |
| LOC105229681 | 105229681 | 3 | NC_064305.1 (87358196–87359090) | Metabolism of xenobiotics by cytochrome P450 | YS vs. WS LYS vs. WS YS vs. LYS |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, G.; Li, W.; Wu, J.; Xu, Y.; Xu, Z.; Xie, Q.; Ge, Y.; Yang, H.; Li, X. Molecular and Biochemical Mechanisms of Scutellum Color Variation in Bactrocera dorsalis Adults (Diptera: Tephritidae). Insects 2025, 16, 76. https://doi.org/10.3390/insects16010076
Wang G, Li W, Wu J, Xu Y, Xu Z, Xie Q, Ge Y, Yang H, Li X. Molecular and Biochemical Mechanisms of Scutellum Color Variation in Bactrocera dorsalis Adults (Diptera: Tephritidae). Insects. 2025; 16(1):76. https://doi.org/10.3390/insects16010076
Chicago/Turabian StyleWang, Guangli, Weijun Li, Jiazhan Wu, Ye Xu, Zhaohuan Xu, Qingxiu Xie, Yugui Ge, Haiyan Yang, and Xiaozhen Li. 2025. "Molecular and Biochemical Mechanisms of Scutellum Color Variation in Bactrocera dorsalis Adults (Diptera: Tephritidae)" Insects 16, no. 1: 76. https://doi.org/10.3390/insects16010076
APA StyleWang, G., Li, W., Wu, J., Xu, Y., Xu, Z., Xie, Q., Ge, Y., Yang, H., & Li, X. (2025). Molecular and Biochemical Mechanisms of Scutellum Color Variation in Bactrocera dorsalis Adults (Diptera: Tephritidae). Insects, 16(1), 76. https://doi.org/10.3390/insects16010076