
The Extraordinary Diversity of Merodon avidus Complex (Diptera: Syrphidae)—Adding New Areas, New Species and a New Molecular Marker
 , , and
, , and
Abstract
Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Molecular Analyses
2.2. Morphological Study
2.3. Type Material
2.4. Geometric Morphometric Analysis
3. Results
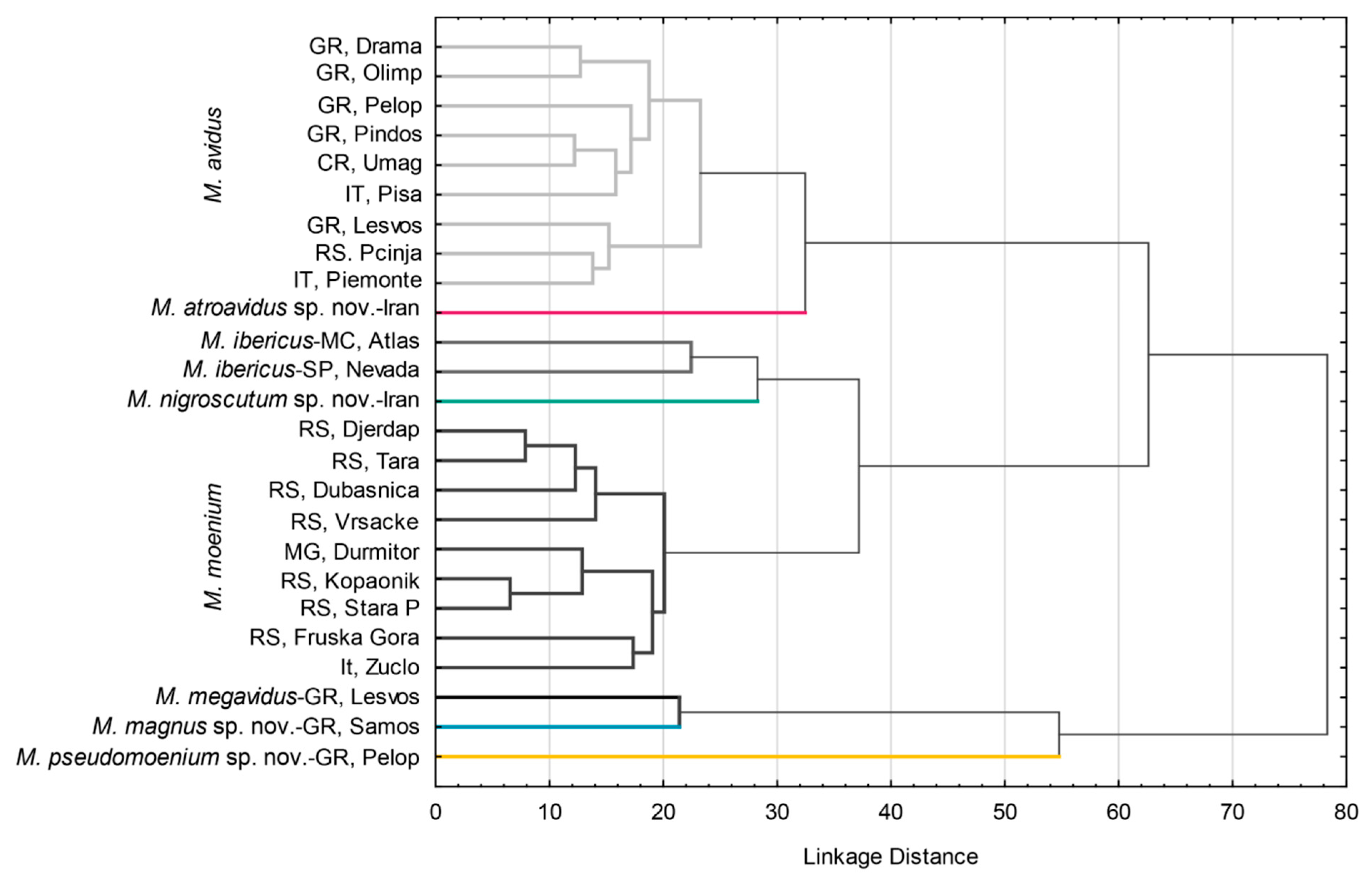
3.1. Molecular Analyses
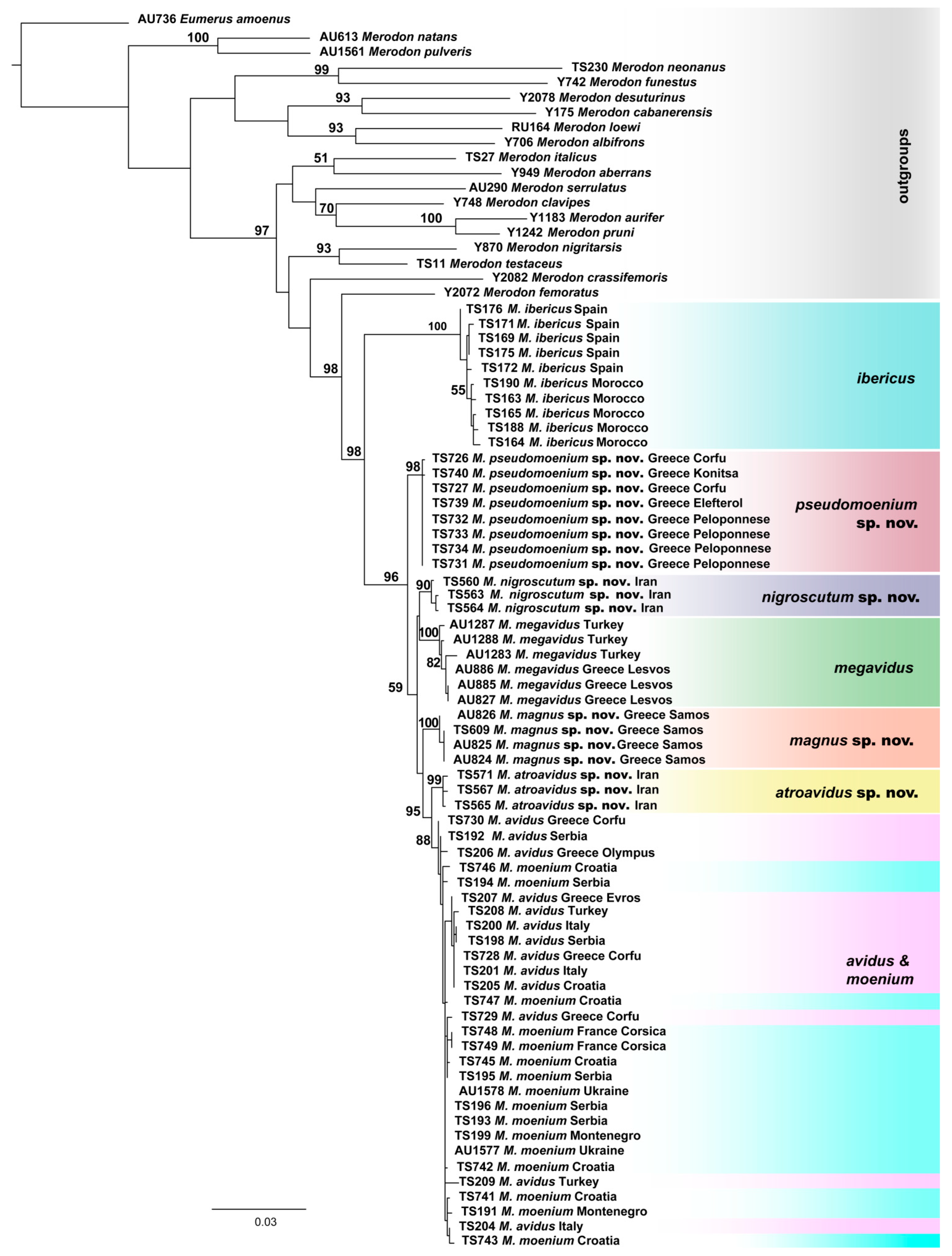
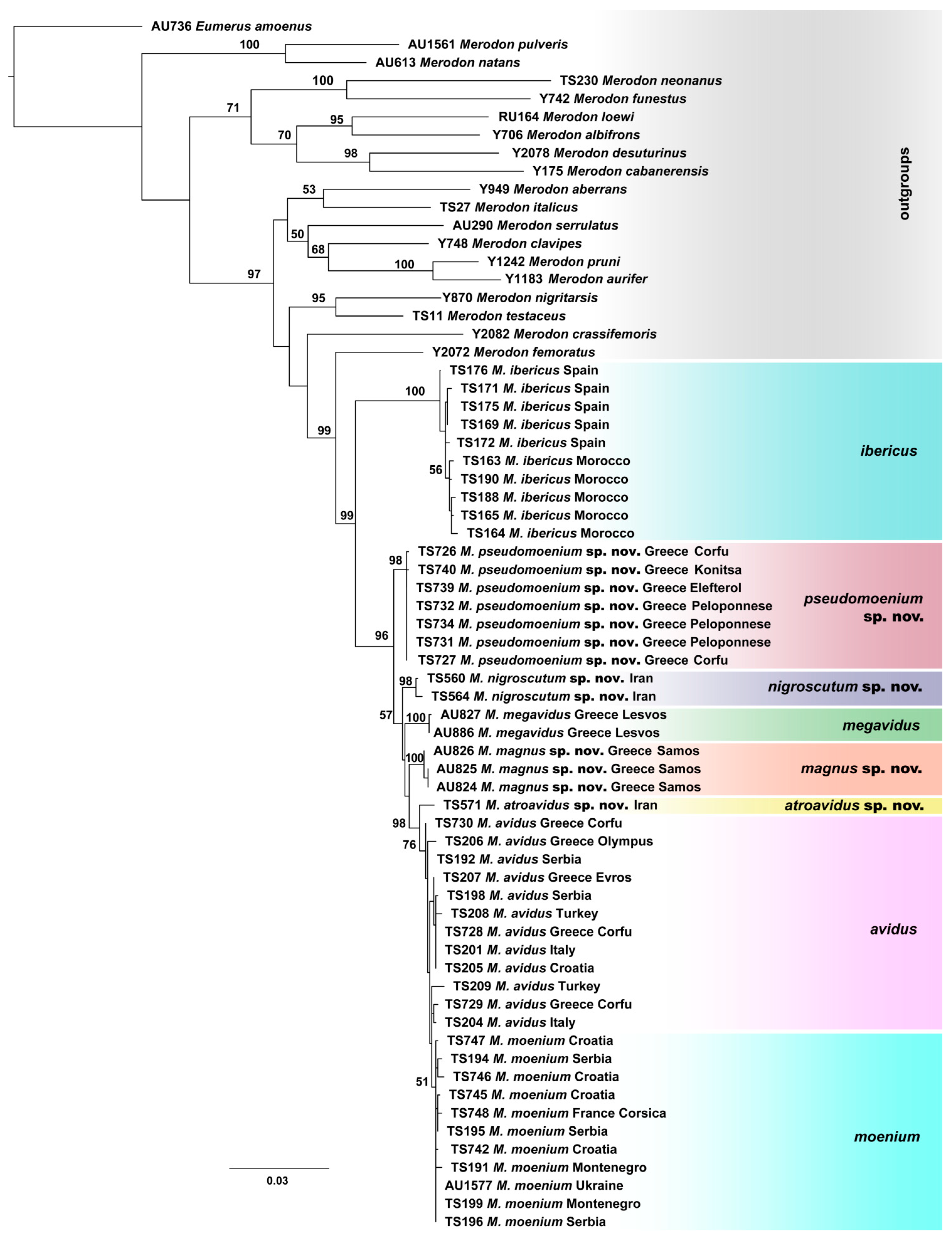
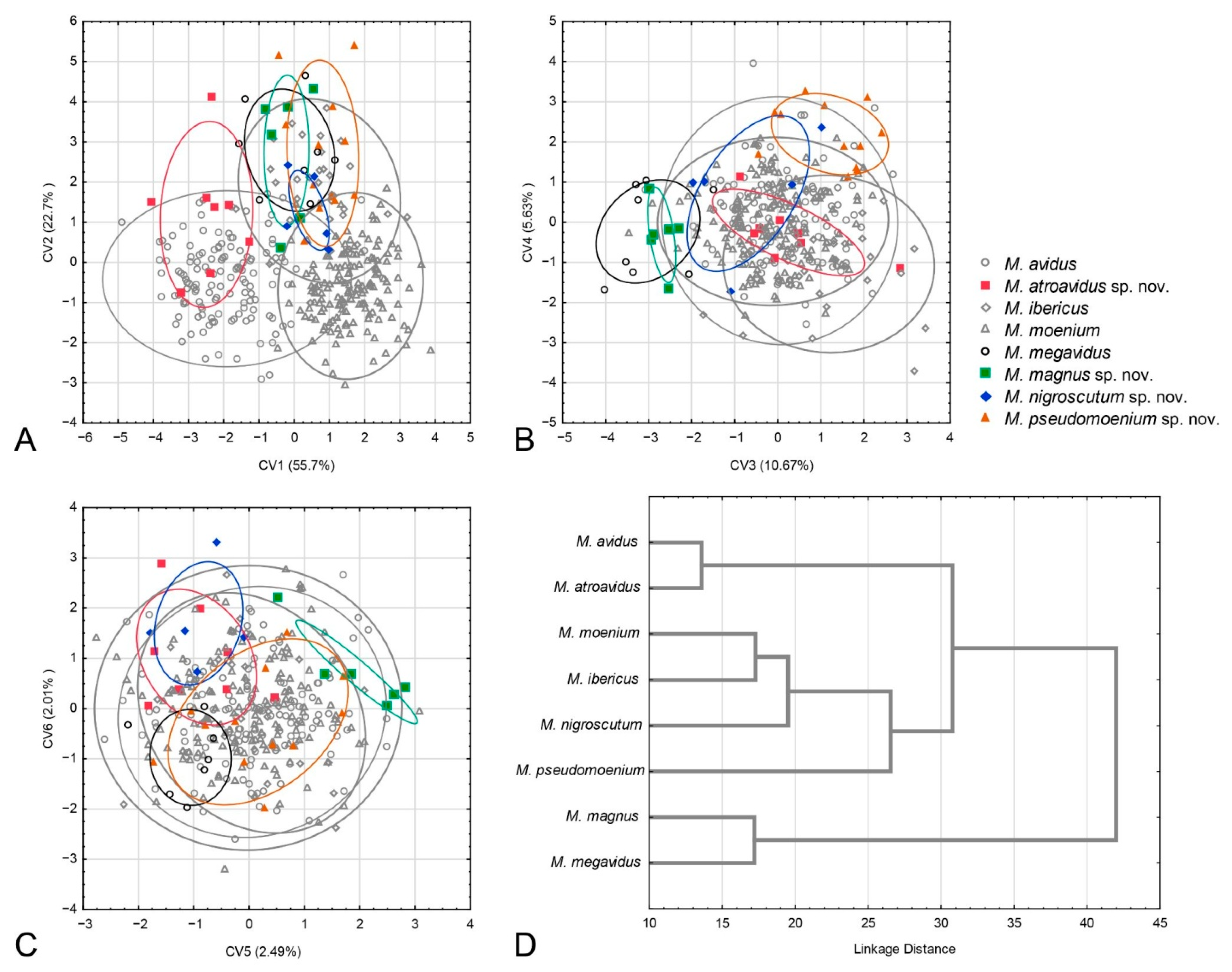
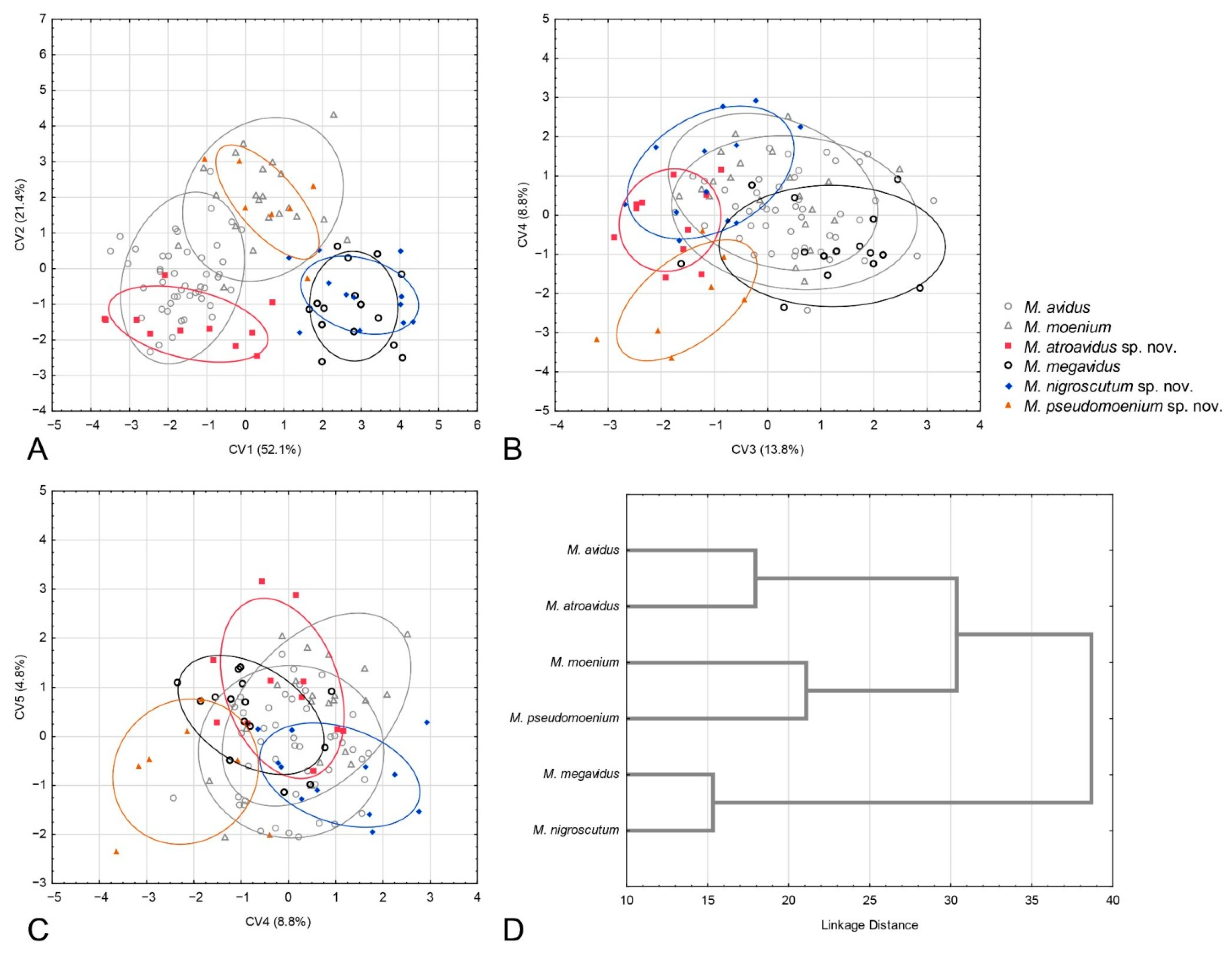
3.2. Geometric Morphometric Analysis
3.2.1. Sexual Dimorphism
3.2.2. Males
3.2.3. Females
3.2.4. Population Analysis
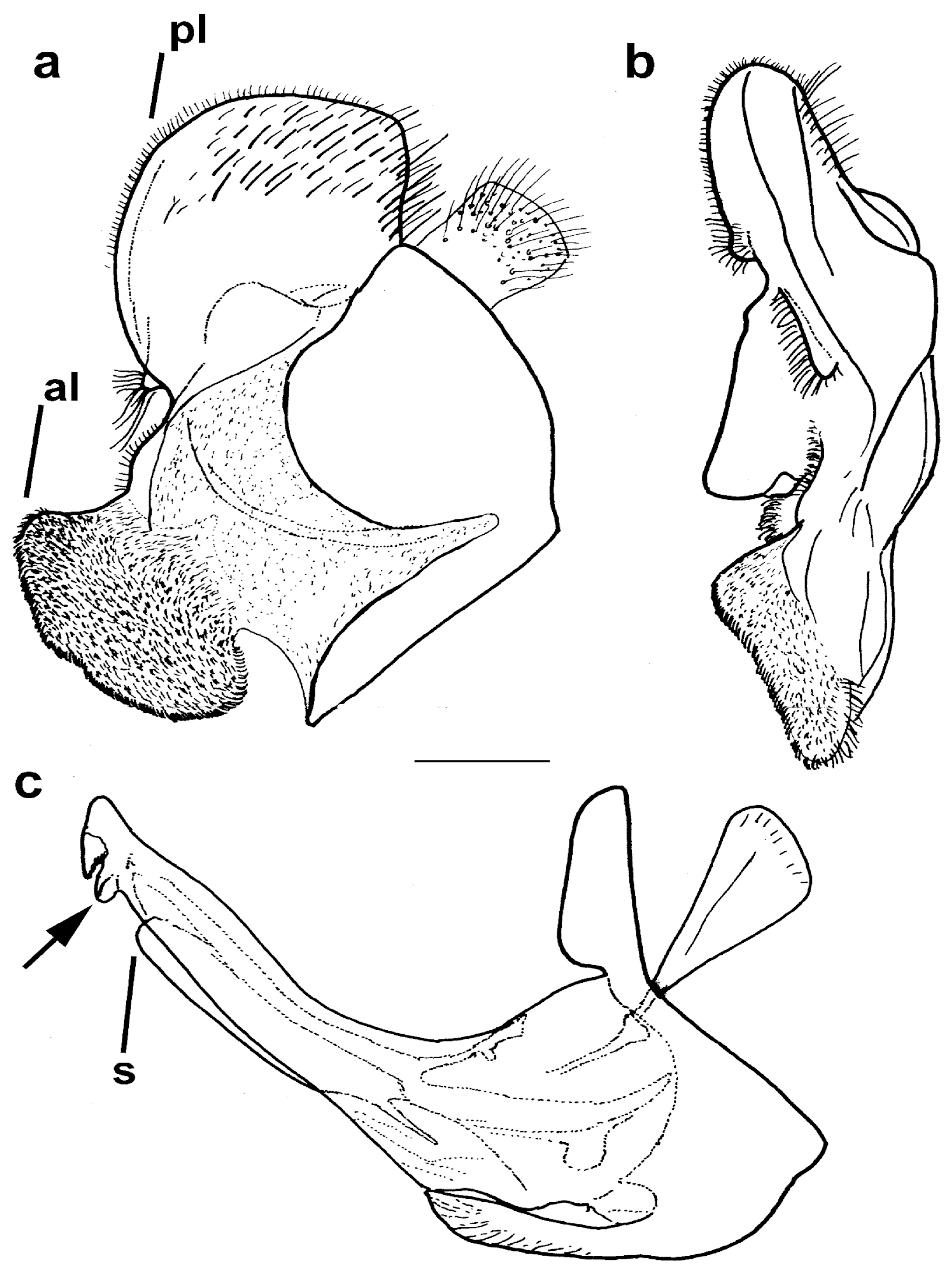
3.3. Morphology and Taxonomy
3.3.1. Merodon avidus Group
3.3.2. Merodon avidus Complex
3.4. Description of New Species


- Key for typical morpho-forms of species from Merodon avidus complex
- 1.
- Pile on metafemur very short (as in Figure 20c,d) ……….…………….…………...........2
- -
- Pile on metafemur longer (as in Figure 20a,b) ………….……………….…………...........3
- 2.
- -
- 3.
- Terga in male black, in female only tergum 2 with reddish triangular lateral maculae (Figure 14c and Figure 15b); most of the posterior half of scutum and scutellum usually with black pilosity (Figure 16b); distribution: Caspian region of Iran and Azerbaijan (Figure 18) …………………………………………………………… ……Merodon nigroscutum sp. nov.
- -
- 4.
- -
- 5.
- -
- 6.
- -

3.5. Variability
3.5.1. Merodon atroavidus sp. nov.
3.5.2. Merodon avidus
3.5.3. Merodon ibericus
3.5.4. Merodon megavidus
3.5.5. Merodon moenium
3.5.6. Merodon pseudomoenium sp. nov.
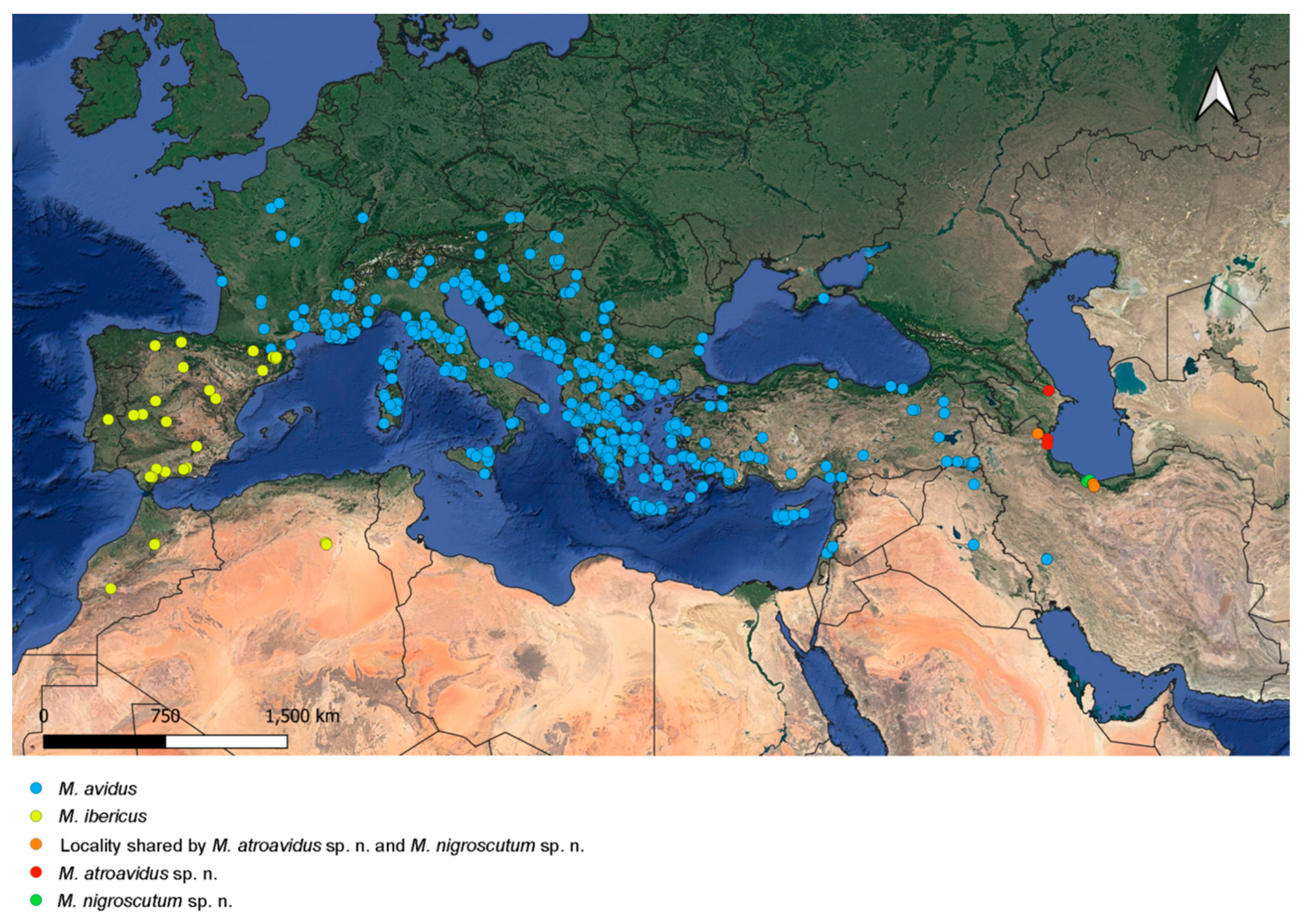
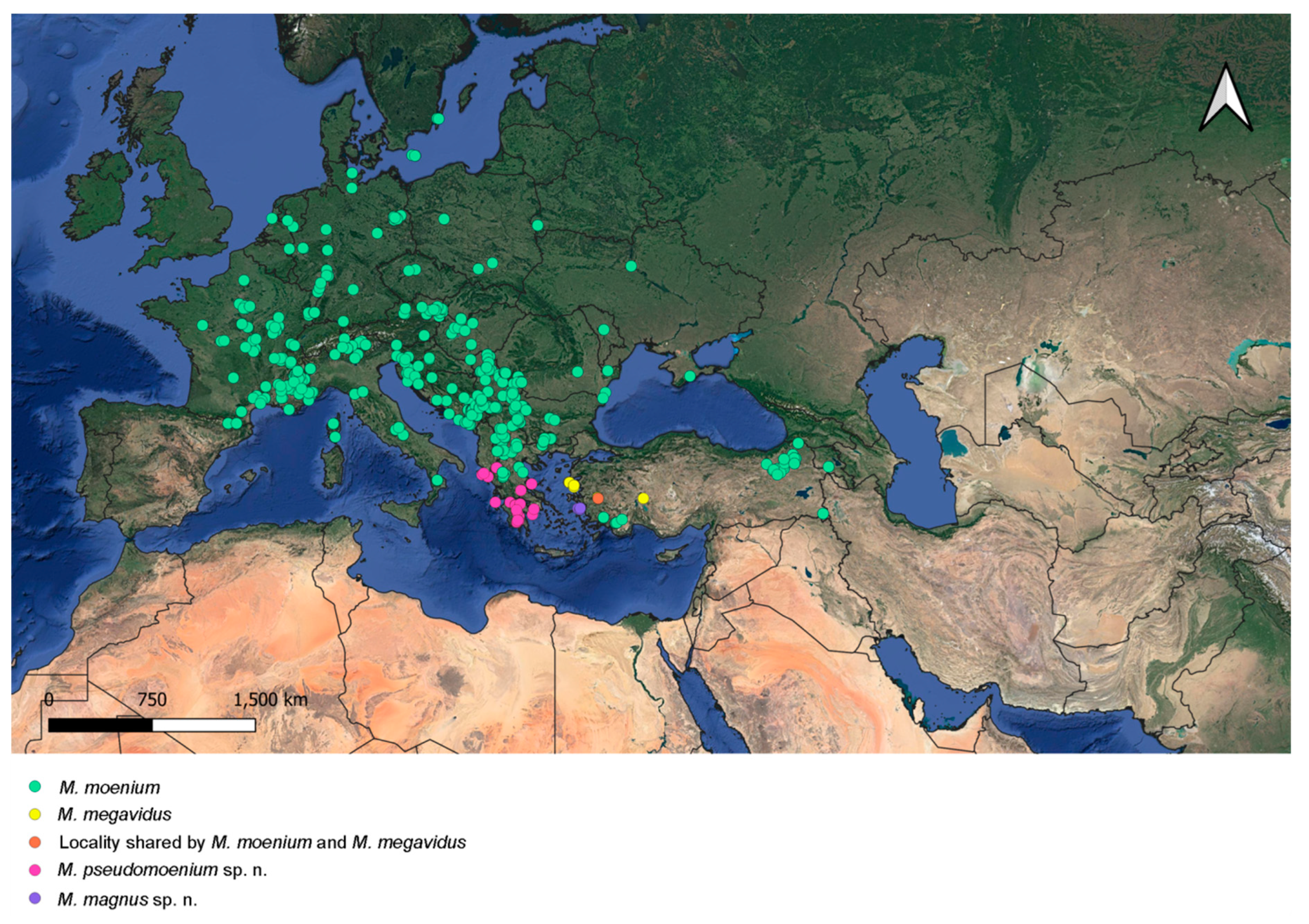
3.6. Distribution
3.7. Biological Notes
4. Discussion
4.1. Molecular Evidence
4.2. Morphological Characters
4.3. Geometric Morphometrics
4.4. Distribution
4.5. Integrative Taxonomy
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vujić, A.; Radenković, S.; Likov, L.; Veselić, S. Taxonomic complexity in the genus Merodon Meigen, 1803 (Diptera, Syrphidae). ZooKeys 2021, 1031, 85–124. [Google Scholar] [CrossRef]
- Vujić, A.; Tot, T.; Andrić, A.; Ačanski, J.; Zorić, L.Š.; Pérez-Bañón, C.; Aracil, A.; Veselić, S.; Arok, M.; Mengual Sanchis, X.; et al. Review of the Merodon natans group with description of a new species, a key to the adults of known species of the natans lineage and first descriptions of some preimaginal stages. Arthropod Syst. Phylogeny 2021, 79, 343–378. [Google Scholar] [CrossRef]
- Speight, M.C.D. Species Accounts of European Syrphidae, 2020. Syrph the Net, the Database of European Syrphidae (Diptera); Syrph the Net Publications: Dublin, Ireland, 2020; ISSN 1393-4546. [Google Scholar]
- Vujić, A.; Radenković, S.; Ståhls, G.; Ačanski, J.; Stefanović, A.; Veselić, S.; Andrić, A.; Hayat, R. Systematics and taxonomy of the ruficornis group of genus Merodon (Diptera: Syrphidae). Syst. Entomol. 2012, 37, 578–602. [Google Scholar] [CrossRef]
- Vujić, A.; Radenković, S.; Kočiš Tubić, N.; Likov, L.; Popov, G.; Rojo, S.; Miličić, M. Integrative taxonomy of the Merodon aberrans (Diptera, Syrphidae) species group: Distribution patterns and description of three new species. Contrib. Zool. 2022, 92, 51–96. [Google Scholar] [CrossRef]
- Ricarte, A.; Marcos-García, M.Á.; Rotheray, G.E. The early stages and life histories of three Eumerus and two Merodon species (Diptera: Syrphidae) from the Mediterranean region. Entomol. Fenn. 2008, 19, 129–141. [Google Scholar] [CrossRef]
- Marcos-García, M.A.; Vujić, A.; Mengual, X. Revision of Iberian species of the genus Merodon (Diptera: Syrphidae). Eur. J. Entomol. 2007, 104, 531–572. [Google Scholar] [CrossRef]
- Vujić, A.; Pérez-Bañón, C.; Radenković, S.; Ståhls, G.; Rojo, S.; Petanidou, T.; Šimić, S. Two new species of the genus Merodon Meigen 1803 (Diptera: Syrphidae) from the island of Lesvos (Greece), the eastern Mediterranean. Ann. Soc. Entomol. 2007, 43, 319–326. [Google Scholar] [CrossRef]
- Vujić, A.; Marcos-García, M.Á.; Sarıbıyık, S.; Ricarte, A. New data on the Merodon Meigen, 1803 fauna (Diptera: Syrphidae) of Turkey, including description of a new species and changes in the nomenclatural status of several taxa. Ann. Soc. Entomol. 2011, 47, 78–88. [Google Scholar] [CrossRef]
- Vujić, A.; Radenković, S.; Likov, L.; Trifunov, S.; Nikolić, T. Three new species of the Merodon nigritarsis group (Diptera: Syrphidae) from the Middle East. Zootaxa 2013, 3640, 442–464. [Google Scholar] [CrossRef] [PubMed]
- Vujić, A.; Radenković, S.; Ačanski, J.; Grković, A.; Taylor, M.; Şenol, S.G.; Hayat, R. Revision of the species of the Merodon nanus group (Diptera: Syrphidae) including three new spe cies. Zootaxa 2015, 4006, 439–462. [Google Scholar] [CrossRef]
- Vujić, A.; Radenković, S.; Likov, L.; Andrić, A.; Gilasian, E.; Barkalov, A. Two new enigmatic species of the genus Merodon Meigen (Diptera: Syrphidae) from the north-eastern Middle East. Zootaxa 2019, 4555, 187–208. [Google Scholar] [CrossRef] [PubMed]
- Vujić, A.; Radenković, S.; Likov, L.; Andrić, A.; Janković, M.; Ačanski, J.; Popov, G.; de Courcy Williams, M.; Šašić Zorić, L.; Djan, M. Conflict and congruence between morphological and molecular data: Revision of the Merodon constans group (Diptera: Syrphidae). Invertebr. Syst. 2020, 34, 406–448. [Google Scholar] [CrossRef]
- Vujić, A.; Likov, L.; Radenković, S.; Kočiš Tubić, N.; Djan, M.; Šebić, A.; Pérez-Bañón, C.; Barkalov, A.; Hayat, R.; Rojo, S.; et al. Revision of the Merodon serrulatus group (Diptera, Syrphidae). ZooKeys 2020, 909, 79–158. [Google Scholar] [CrossRef] [PubMed]
- Vujić, A.; Speight, M.C.D.; de Courcy Williams, M.; Rojo, S.; Ståhls, G.; Radenković, S.; Likov, L.; Miličić, M.; Pérez-Bañón, C.; Falk, S.; et al. Atlas of the Hoverflies of Greece; Brill: Leiden, The Netherlands, 2020. [Google Scholar] [CrossRef]
- Vujić, A.; Likov, L.; Popov, S.; Radenković, S.; Hauser, M. Revision of the Merodon aurifer group (Diptera: Syrphidae) with new synonyms of M. testaceus Sack, 1913. J. Asia Pac. Entomol. 2021, 24, 1301–1312. [Google Scholar] [CrossRef]
- Vujić, A.; Radenković, S.; Likov, L.; Gorše, I.; Djan, M.; Markov Ristić, Z.; Barkalov, A. Three new species of the Merodon ruficornis group (Diptera: Syrphidae) discovered at the edge of its range. Zootoxa 2022, 5182, 301–347. [Google Scholar] [CrossRef]
- Ståhls, G.; Vujić, A.; Pérez-Bañón, C.; Radenković, S.; Rojo, S.; Petanidou, T. COI barcodes for identification of Merodon hoverflies (Diptera, Syrphidae) of Lesvos Island, Greece. Mol. Ecol. Resour. 2009, 9, 1431–1438. [Google Scholar] [CrossRef]
- Ståhls, G.; Vujić, A.; Petanidou, T.; Cardoso, P.; Radenković, S.; Ačanski, J.; Pérez-Bañón, C.; Santos, R. Phylogeographic patterns of Merodon hoverflies in the Eastern Mediterranean region: Revealing connections and barriers. Ecol. Evol. 2016, 6, 2226–2245. [Google Scholar] [CrossRef] [PubMed]
- Radenković, S.; Vujić, A.; Ståhls, G.; Pérez-Bañon, C.; Rojo, S.; Petanidou, T.; Šimić, S. Three new cryptic species of the genus Merodon Meigen (Diptera: Syrphidae) from the island of Lesvos (Greece). Zootaxa 2011, 2735, 35–56. [Google Scholar] [CrossRef]
- Radenković, S.; Vujić, A.; Obreht Vidaković, D.; Djan, M.; Milić, D.; Veselić, S.; Ståhls, G.; Petanidou, T. Sky island diversification in the Merodon rufus group (Diptera, Syrphidae)—Recent vicariance in south-east Europe. Org. Divers. Evol. 2020, 20, 345–368. [Google Scholar] [CrossRef]
- Ačanski, J.; Vujić, A.; Djan, M.; Obreht-Vidaković, D.; Ståhls, G.; Radenković, S. Defining species boundaries in the Merodon avidus complex (Diptera, Syrphidae) using integrative taxonomy, with the description of a new species. Eur. J. Taxon. 2016, 237, 1–25. [Google Scholar] [CrossRef]
- Kaloveloni, A.; Tscheulin, T.; Vujić, A.; Radenković, S.; Petanidou, T. Winners and losers of climate change for the genus Merodon (Diptera: Syrphidae) across the Balkan Peninsula. Ecol. Modell. 2015, 313, 201–211. [Google Scholar] [CrossRef]
- Ačanski, J.; Miličić, M.; Likov, L.; Milić, D.; Radenković, S.; Vujić, A. Environmental niche divergence of species from Merodon ruficornis group (Diptera: Syrphidae). Arch. Biol. Sci. 2017, 69, 247–259. [Google Scholar] [CrossRef]
- Likov, L.; Vujić, A.; Kočiš Tubić, N.; Djan, M.; Veličković, N.; Rojo, S.; Pérez-Bañón, C.; Veselić, S.; Barkalov, A.; Rüstem, H.; et al. Systematic position and composition of Merodon nigritarsis and M. avidus groups (Diptera, Syrphidae) with a description of four new hoverflies species. Contrib. Zool. 2020, 89, 74–125. [Google Scholar] [CrossRef]
- Milankov, V.; Vujić, A.; Ludoški, J. Genetic divergence among cryptic taxa of Merodon avidus (Rossi, 1790) (Diptera: Syrphidae). Int. J. Dipt. Res. 2001, 12, 15–24. [Google Scholar]
- Milankov, V.; Ludoški, J.; Ståhls, G.; Stamenković, J.; Vujić, A. High molecular and phenotypic diversity in the Merodon avidus complex (Diptera, Syrphidae): Cryptic speciation in a diverse insect taxon. Zool. J. Linn. Soc. 2009, 155, 819–833. [Google Scholar] [CrossRef]
- Popović, D.; Ačanski, J.; Djan, M.; Obreht, D.; Vujić, A.; Radenković, S. Sibling species delimitation and nomenclature of the Merodon avidus complex (Diptera: Syrphidae). Eur. J. Entomol. 2015, 112, 790–809. [Google Scholar] [CrossRef]
- Hurkmans, W. A monograph of Merodon (Diptera: Syrphidae). Part 1. Tijdschr. Entomol. 1993, 136, 147–234. [Google Scholar]
- Vujić, A.; Radenković, S.; Likov, L. Revision of the Palaearctic species of the Merodon desuturinus group (Diptera, Syrphidae). ZooKeys 2018, 771, 105–138. [Google Scholar] [CrossRef]
- Šašić, L.; Ačanski, J.; Vujić, A.; Ståhls, G.; Radenković, S.; Milić, D.; Obreht-Vidaković, D.; Djan, M. Molecular and morphological inference of three cryptic species within the Merodon aureus species group (Diptera: Syrphidae). PLoS ONE 2016, 11, e0160001. [Google Scholar] [CrossRef]
- Veselić, S.; Vujić, A.; Radenković, S. Three new Eastern-Mediterranean endemic species of the Merodon aureus group (Diptera: Syrphi dae). Zootaxa 2017, 4254, 401–434. [Google Scholar] [CrossRef]
- Radenković, S.; Šašić Zorić, L.; Djan, M.; Obreht Vidaković, D.; Ačanski, J.; Ståhls, G.; Veličković, N.; Markov, Z.; Petanidou, T.; Kočiš Tubić, N.; et al. Cryptic speciation in the Merodon luteomaculatus complex (Diptera: Syrphidae) from the eastern Mediterranean. J. Zool. Syst. Evol. Res. 2018, 56, 170–191. [Google Scholar] [CrossRef]
- Kočiš Tubić, N.; Ståhls, G.; Ačanski, J.; Djan, M.; Obreht Vidaković, D.; Hayat, R.; Khaghaninia, S.; Vujić, A.; Radenković, S. An integrative approach in the assessment of species delimitation and structure of the Merodon nanus species group (Diptera: Syrphidae). Org. Divers. Evol. 2018, 18, 479–497. [Google Scholar] [CrossRef]
- Djan, M.; Ståhls, G.; Veličković, N.; Ačanski, J.; Vidaković, D.O.; Rojo, S.; Pérez-Bañón, C.; Radenković, S.; Vujić, A. The Merodon planifacies subgroup (Diptera, Syrphidae): Congruence of molecular and morphometric evidences reveal new taxa in Drakensberg mountains valleys (Republic of South Africa). Zool. Anz. 2020, 287, 105–120. [Google Scholar] [CrossRef]
- Aracil, A.; Ačanski, J.; Pérez-Bañón, C.; Šikoparija, B.; Miličić, M.; Campoy, A.; Radenković, S.; Vujić, A.; Radišić, P.; Rojo, S. Characterization of preimaginal developmental stages of two cryptic South African species of the Merodon planifacies complex (Diptera: Syrphidae: Eristalinae: Merodontini), with differentiation through morphometry analysis. Arthropod Struct. Dev. 2022, 70, 101–187. [Google Scholar] [CrossRef]
- Vujić, A.; Radenković, S.; Barkalov, A.; Kočiš Tubić, N.; Likov, L.; Tot, T.; Popov, G.; Prokhorov, A.; Gilasian, E.; Anjum, S.; et al. Taxonomic revision of the Merodon tarsatus species group (Diptera, Syrphidae). Arthropod Syst. Phylogeny 2023, 81, 201–256. [Google Scholar] [CrossRef]
- Birdsall, K.; Zimmerman, E.; Teeter, K.; Gibson, G. Genetic variation for the positioning of wing veins in Drosophila melanogaster. Evol. Dev. 2000, 2, 16–24. [Google Scholar] [CrossRef]
- Vujić, A.; Ståhls, G.; Ačanski, J.; Bartsch, H.; Bygebjerg, R.; Stefanović, A. Systematics of Pipizini and taxonomy of European Pipiza Fallén: Molecular and morphological evidence (Diptera, Syrphidae). Zool. Scr. 2013, 42, 288–305. [Google Scholar] [CrossRef]
- Nedeljković, Z.; Ačanski, J.; Vujić, A.; Obreht, D.; Ðan, M.; Ståhls, G.; Radenković, S. Taxonomy of Chrysotoxum festivum Linnaeus, 1758 (Diptera: Syrphidae)—An integrative approach. Zool. J. Linn. Soc. 2013, 169, 84–102. [Google Scholar] [CrossRef]
- Nedeljković, Z.; Ačanski, J.; Đan, M.; Obreht-Vidaković, D.; Ricarte, A.; Vujić, A. An integrated approach to delimiting species borders in the genus Chrysotoxum Meigen, 1803 (Diptera: Syrphidae), with description of two new species. Contrib. Zool. 2015, 84, 285–304. [Google Scholar] [CrossRef]
- Chroni, A.; Grković, A.; Ačanski, J.; Vujić, A.; Radenković, S.; Veličković, N.; Djan, M.; Petanidou, T. Disentangling a cryptic species complex and defining new species within the Eumerus minotaurus group (Diptera: Syrphidae), based on integrative taxonomy and Aegean palaeogeography. Contrib. Zool. 2018, 87, 197–225. [Google Scholar] [CrossRef]
- Nei, M. Estimation of average heterozigosity and genetic distance from a small number of individuals. Genetics 1978, 89, 583–590. [Google Scholar] [CrossRef]
- Milić, D.; Radenković, S.; Radišić, D.; Andrić, A.; Nikolić, T.; Vujić, A. Stability and changes in the distribution of Pipiza hoverflies (Diptera, Syrphidae) in Europe under projected future climate conditions. PLoS ONE 2019, 14, e0221934. [Google Scholar] [CrossRef]
- Milić, D.; Radenković, S.; Ačanski, J.; Vujić, A. The importance of hidden diversity in insect conservation—A case study in hoverflies (the Merodon atratus complex, Syrphidae, Diptera). J. Insect Conserv. 2019, 23, 29–44. [Google Scholar] [CrossRef]
- Vujić, A.; Šašić Zorić, L.; Ačanski, J.; Likov, L.; Radenković, S.; Đan, M.; Milić, D.; Šebić, A.; Ranković, M.; Khaghaninia, S. Hide-and-seek with hoverflies: Merodon aureus—A species, a complex or a subgroup? Zool. J. Linn. Soc. 2020, 190, 974–1001. [Google Scholar] [CrossRef]
- Chen, H.; Rangasamy, M.; Tan, S.Y.; Wang, H.; Siegfried, B.D. Evaluation of five methods for total DNA extraction from western corn rootworm beetles. PLoS ONE 2010, 5, e11963. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Simon, C.; Frati, F.; Beckenbach, A.; Crespi, B.; Liu, H.; Flook, P. Evolution, weighting, and phylogenetic utility of mitochondrial gene sequences and a compilation of conserved polymerase chain reaction primers. Ann. Entomol. Soc. Am. 1994, 87, 651–701. [Google Scholar] [CrossRef]
- Belshaw, R.; Lopez-Vaamonde, C.; Degerli, N.; Quicke, D.L. Paraphyletic taxa and taxonomic chaining: Evaluating the classification of braconine wasps (Hymenoptera: Braconidae) using 28S D2-3 rDNA sequences and morphological characters. Biol. J. Linn. Soc. 2001, 73, 411–424. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/ NT. Nucleic Acids Symp. 1999, 41, 95–98. [Google Scholar]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Akaike, H. A new look at the statistical model identification. IEEE Trans. Autom. Control 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the Gateway Computing Environments Workshop (GCE), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarization in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A. FigTree: A Graphical Viewer of Phylogenetic Trees, Ver. 1.4.4. 2018. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 10 June 2023).
- Stamatakis, A. RAxML ver. 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, F.; Oliver, J.L.; Marin, A.; Medina, J.R. The general stochastic model of nucleotide substitution. J. Theor. Biol. 1990, 142, 485–501. [Google Scholar] [CrossRef] [PubMed]
- Goloboff, P.A. NONA Computer Program. Version 2.0. Tucuman, Argentina. 1999. Available online: www.softpedia.com/get/Scienece-CAD/NONA.shtml (accessed on 8 May 2023).
- Nixon, K.C. WinClada Ver. 1.00.08. Published by the Author. Ithaca. 2002. Available online: https://www.softwaresea.com/Windows/download-WinClada-10543895.htm (accessed on 8 May 2023).
- QGIS Development Team. QGIS Geographic Information System. Open Source Geospatial Foundation Project. 2021. Available online: http://qgis.osgeo.org (accessed on 17 May 2023).
- Hadley, A. CombineZ, Ver. 5. Published by the Author. 2006. Available online: http://www.hadleyweb.pwp.blueyonder.co.uk/CZ5/combinez5.htm (accessed on 17 May 2023).
- Thompson, F.C. A key to the genera of the flower flies (Diptera: Syrphidae) of the Neo tropical Region including descriptions of new genera and species and a glossary of taxo nomic terms. Contrib. Entomol. Int. 1999, 3, 321–378. [Google Scholar]
- Bookstein, F.L. Morphometrics Tools for Landmark Data; Cambridge University Press: New York, NY, USA, 1991. [Google Scholar]
- Rohlf, F.J. tpsDig Program: Digitize Coordinates of Landmarks and Capture Outlines, Ver. 2.31; Department of Ecology and Evolution, State University of New York, Stony Brook: New York, NY, USA, 2017; Available online: https://www.sbmorphometrics.org/soft-dataacq.html (accessed on 14 May 2023).
- Klingenberg, C.P. MorphoJ: An integrated software package for geometric morphometrics. Mol. Ecol. Resour. 2011, 11, 353–357. [Google Scholar] [CrossRef]
- Rohlf, F.J.; Slice, D.E. Extensions of the Procrustes method for the optimal superimposition of landmarks. Syst. Zool. 1990, 39, 40–59. [Google Scholar] [CrossRef]
- Zelditch, M.L.; Swiderski, D.L.; Sheets, H.D.; Fink, W.L. Geometric Morphometrics for Biologists: A Primer, 2nd ed.; Elsevier Academic Press: Amsterdam, The Netherlands, 2004. [Google Scholar]
- Baylac, M.; Frieß, M. Fourier Descriptors, Procrustes Superimposition, and Data Dimensionality: An Example of Cranial Shape Analysis in Modern Human Populations. In Modern Morphometrics in Physical Anthropology; Springer: Boston, MA, USA, 2005; pp. 145–165. [Google Scholar]
- TIBCO Software Inc. Statistica. Data Analysis Software System. 2018. Available online: https://www.tibco.com/ (accessed on 26 May 2023).
- Marcos-García, M.Á.; Vujić, A.; Ricarte, A.; Ståhls, G. Towards an integrated taxonomy of the Merodon equestris species complex (Diptera: Syrphidae) including description of a new species, with additional data on Iberian Merodon. Can. Entomol. 2011, 143, 332–348. [Google Scholar] [CrossRef]
- Patwardhan, A.; Ray, S.; Roy, A. Molecular markers in phylogenetic studies—A review. J. Phylogenetics Evol. Biol. 2014, 2, 131. [Google Scholar] [CrossRef]
- Moraes, E.M.; Manfrin, M.H.; Laus, A.C.; Rosada, R.S.; Bomfin, S.C.; Sene, F.M. Wing shape heritability and morphological divergence of the sibling species Drosophila mercatorum and Drosophila paranaensis. Heredity 2004, 92, 466–473. [Google Scholar] [CrossRef] [PubMed]
- Mezey, J.G.; Houle, D. The dimensionality of genetic variation for wing shape in Drosophila melanogaster. Evolution 2005, 59, 1027–1038. [Google Scholar] [CrossRef]
- Dworkin, I.; Gibson, G. Epidermal growth factor receptor and transforming growth factor-b signaling contributes to variation for wing shape in Drosophila melanogaster. Genetics 2006, 173, 1417–1431. [Google Scholar] [CrossRef] [PubMed]
- Yeaman, S.; Chen, Y.; Whitlock, M.C. No effect of environmental heterogeneity on the maintenance of genetic variation in wing shape in Drosophila melanogaster. Evolution 2010, 64, 3398–3408. [Google Scholar] [CrossRef]
- Šasić, L.; Ačanski, J.; Djan, M.; Kočiš-Tubić, N.; Veličković, N.; Radenković, S.; Vujić, A. Integrative taxonomy of Merodon caerulescens complex (Diptera: Syrphidae) evidence of cryptic speciation. Zb. Matice Srp. Prir. Nauk. 2018, 135, 103–118. [Google Scholar] [CrossRef]
- Ricarte, A.; Souba-Dols, G.J.; Hauser, M.; Marcos-García, M.Á. A review of the early stages and host plants of the genera Eumerus and Merodon (Diptera: Syrphidae), with new data on four species. PLoS ONE 2017, 12, e0189852. [Google Scholar] [CrossRef]
- Preradović, J.; Andrić, A.; Radenković, S.; Šašić Zorić, L.; Pérez-Baňón, C.; Campoy, A.; Vujić, A. Pupal stages of three species of the phytophagous genus Merodon Meigen (Diptera: Syrphidae). Zootaxa 2018, 4420, 229–242. [Google Scholar] [CrossRef]
- Mittermeier, R.A.; Myers, N.; Thomsen, J.B.; Da Fonseca, G.A.; Olivieri, S. Biodiversity hotspots and major tropical wilderness areas: Approaches to setting conservation priorities. Conserv. Biol. 1998, 12, 516–520. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Da Fonseca, G.A.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef]
- Didukh, Y.P.; Litvinskaya, S.A.; Novosad, V.V. Botanico-geographical regionalization of the Crimea-Novorossiysk Province. Bot. Zhurn. 1990, 75, 494–507. [Google Scholar]
- Cordova, C.E. Holocene Mediterranization of the Southern Crimean vegetation: Paleoecological records, regional climate change, and possible non-climatic influences. In The Black Sea Flood Question: Changes in Coastline, Climate, and Human Settlement; Yanko-Hombach, V., Gilbert, A.S., Panin, N., Dolukhanov, P.M., Eds.; Springer: Berlin, Germany, 2007; pp. 319–344. [Google Scholar]
- Cordova, C.E. The Mediterraneanization of Crimea. Physical and cultural processes in landscape transformation. J. Mediterr. Geogr. 2016, 126, 25–36. [Google Scholar] [CrossRef]
- Vyed’, I.P. Klimatichyeskiy Atlas Kryma [Climate Atlas of Crimea]; Tavriya-Plyus: Odesa, Ukraine, 2000; p. 120. [Google Scholar]
- Prokhorov, A.V.; Popov, G.V.; Zaika, M.I. New Records of Hoverflies (Diptera, Syrphidae) from Ukraine. II. Brachyopini and Merodontini. Vestn. Zool. 2018, 52, 125–136. [Google Scholar] [CrossRef]
- Ricarte, A.; Nedeljković, Z.; Rotheray, G.E.; Lyszkowski, R.M.; Hancock, E.G.; Watt, K.; Hewitt, S.M.; Horsfield, D.; Wilkinson, G. Syrphidae (Diptera) from the Greek island of Lesvos, with description of two new species. Zootaxa 2012, 3175, 1–23. [Google Scholar] [CrossRef]
- Ačanski, J.; Vujić, A.; Zorić, L.Š.; Radenković, S.; Djan, M.; Ristić, Z.M.; Ståhls, G. Merodon chalybeus subgroup: An additional piece of the M. aureus group (Diptera, Syrphidae) puzzle. Ann. Zool. Fenn. 2022, 59, 79–109. [Google Scholar] [CrossRef]
- Zorić, L.Š.; Ačanski, J.; Vujić, A.; Ståhls, G.; Djan, M.; Radenković, S. Resolving the taxonomy of the Merodon dobrogensis species subgroup (Diptera: Syrphidae), with the description of a new species. Can. Entomol. 2020, 152, 36–59. [Google Scholar] [CrossRef]
- Hewitt, G.M. Post-glacial re-colonization of European biota. Biol. J. Linn. Soc. 1999, 68, 87–112. [Google Scholar] [CrossRef]
- Ahmadzadeh, F.; Flecks, M.; Rödder, D.; Böhme, W.; Ilgaz, Ç.; Harris, D.J.; Carretero, M.A. Multiple dispersal out of Anatolia: Biogeography and evolution of oriental green lizards. Biol. J. Linn. Soc. 2013, 110, 398–408. [Google Scholar] [CrossRef]
- Ashrafzadeh, M.R.; Kaboli, M.; Naghavi, M.R. Mitochondrial DNA analysis of Iranian brown bears (Ursus arctos) reveals new phylogeographic lineage. Mamm. Biol. 2016, 81, 1–9. [Google Scholar] [CrossRef]
- Dufresnes, C.; Litvinchuk, S.N.; Leuenberger, J.; Ghali, K.; Zinenko, O.; Stöck, M.; Perrin, N. Evolutionary melting pots: A biodiversity hotspot shaped by ring diversifications around the Black Sea in the Eastern tree frog (Hyla orientalis). Mol. Ecol. 2016, 25, 4285–4300. [Google Scholar] [CrossRef] [PubMed]
- Naderi, G.; Kaboli, M.; Koren, T.; Karami, M.; Zupan, S.; Rezaei, H.R.; Krystufek, B. Mitochondrial evidence uncovers a refugium for the fat dormouse (Glis glis Linnaeus, 1766) in Hyrcanian forests of northern Iran. Mamm. Biol. 2014, 79, 202–207. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Character | Summer | Spring |
|---|---|---|
| colour of basoflagellomere | reddish-yellow | dark brown |
| colour of male metatibia | yellow | brown medially |
| colour of terga | mostly reddish-yellow (except terga 1–2 medially) | mostly dark (except terga 2–3 laterally) |
| body pilosity | reddish | grey-yellow |
| pile on scutum | all yellow (except wing bases) | posterior half black pilose (except posterior margin) |
| pollinosity on tergum 2 | distinct | indistinct |
| Character | Low Altitudes | High Altitudes |
|---|---|---|
| size | large (13–16 mm) | medium (10–13 mm) |
| metatibia | reddish-yellow | yellow with dark medial ring |
| body pilosity | orange to reddish-yellow | pale yellow |
| black adpressed pile at basitarsus of metaleg dorsally | only a few, limited to the basal half (Figure 9e) | on most of the surface (Figure 9f) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vujić, A.; Kočiš Tubić, N.; Radenković, S.; Ačanski, J.; Likov, L.; Arok, M.; Gorše, I.; Djan, M. The Extraordinary Diversity of Merodon avidus Complex (Diptera: Syrphidae)—Adding New Areas, New Species and a New Molecular Marker. Insects 2024, 15, 105. https://doi.org/10.3390/insects15020105
Vujić A, Kočiš Tubić N, Radenković S, Ačanski J, Likov L, Arok M, Gorše I, Djan M. The Extraordinary Diversity of Merodon avidus Complex (Diptera: Syrphidae)—Adding New Areas, New Species and a New Molecular Marker. Insects. 2024; 15(2):105. https://doi.org/10.3390/insects15020105
Chicago/Turabian StyleVujić, Ante, Nataša Kočiš Tubić, Snežana Radenković, Jelena Ačanski, Laura Likov, Maja Arok, Iva Gorše, and Mihajla Djan. 2024. "The Extraordinary Diversity of Merodon avidus Complex (Diptera: Syrphidae)—Adding New Areas, New Species and a New Molecular Marker" Insects 15, no. 2: 105. https://doi.org/10.3390/insects15020105
APA StyleVujić, A., Kočiš Tubić, N., Radenković, S., Ačanski, J., Likov, L., Arok, M., Gorše, I., & Djan, M. (2024). The Extraordinary Diversity of Merodon avidus Complex (Diptera: Syrphidae)—Adding New Areas, New Species and a New Molecular Marker. Insects, 15(2), 105. https://doi.org/10.3390/insects15020105