
Foraging Behavior of Two Pollen Wasp Species of the Genus Celonites Latreille, 1802 (Hymenoptera: Vespidae: Masarinae), from the Altai Mountains


Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Field Investigations
2.2. Morphological Investigations
2.3. DNA Barcoding
3. Results
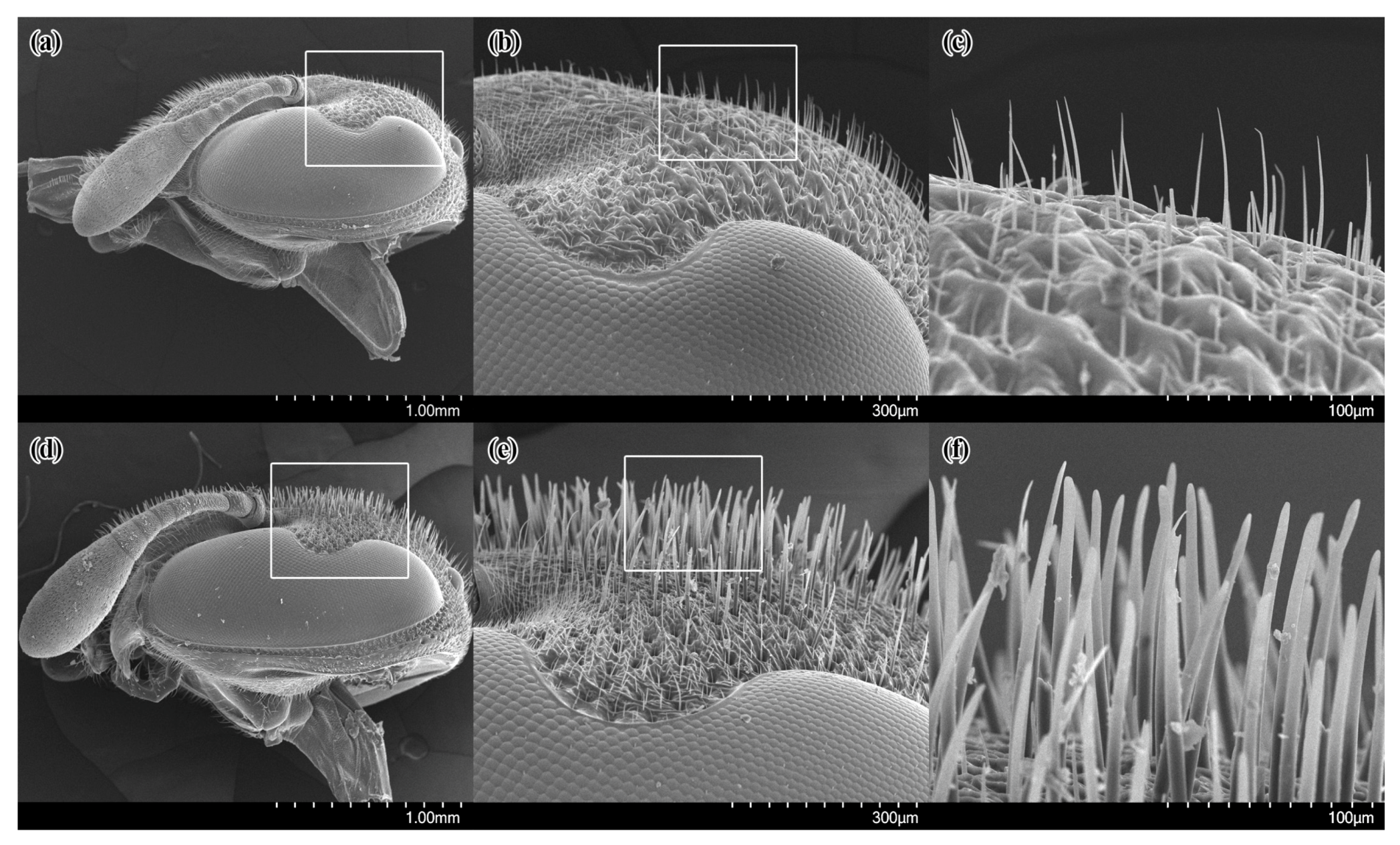
3.1. Pollen-Collecting Apparatus of the Wasps
3.2. Flower-Visiting Records
3.3. Behavior of the Wasps on Flowers
3.3.1. Asteraceae
3.3.2. Brassicaceae
3.3.3. Caryophyllaceae
3.3.4. Convolvulaceae
3.3.5. Fabaceae
3.3.6. Lamiaceae
3.3.7. Rosaceae
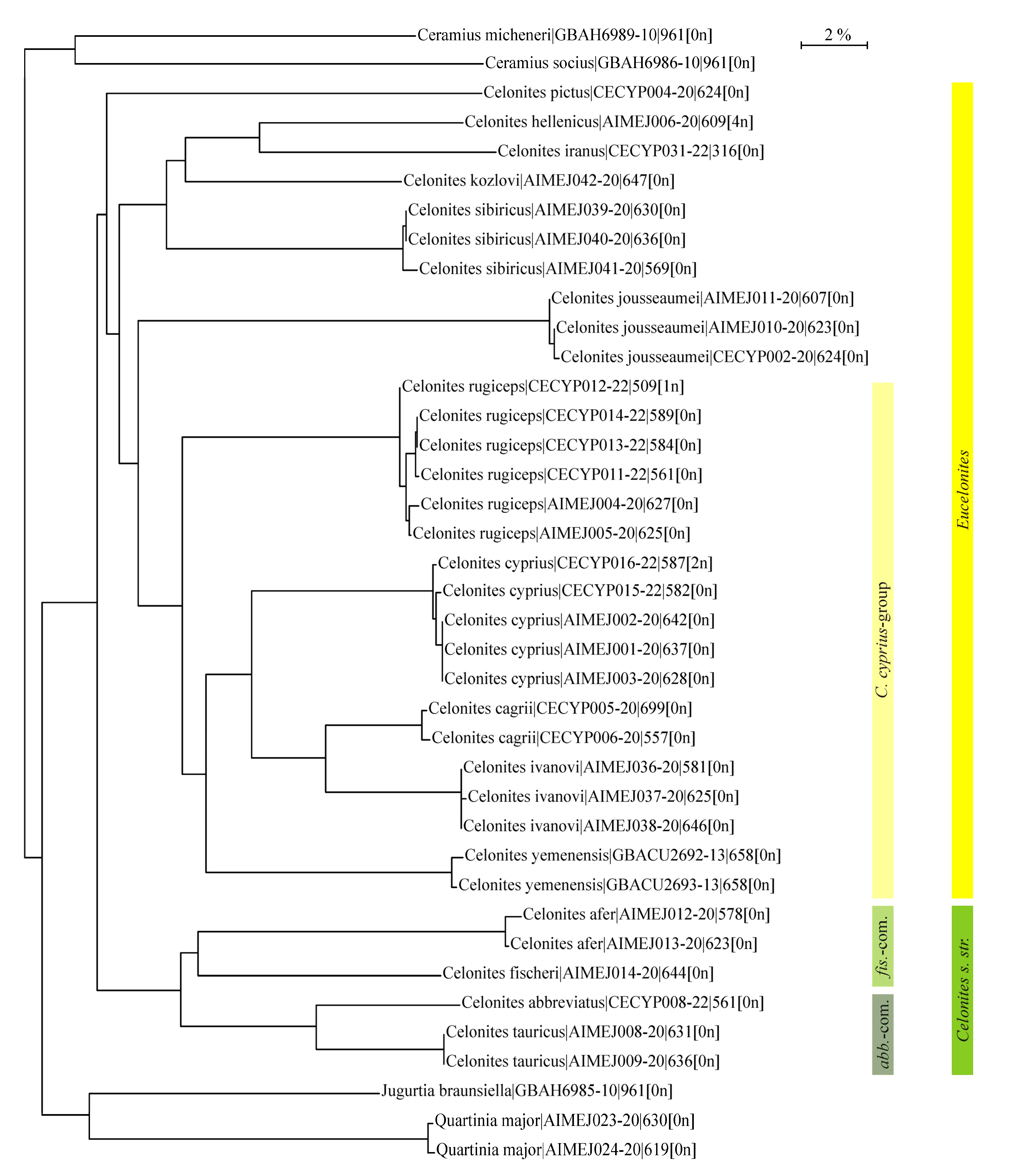
3.4. DNA Barcoding Results: Taxon Tree
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
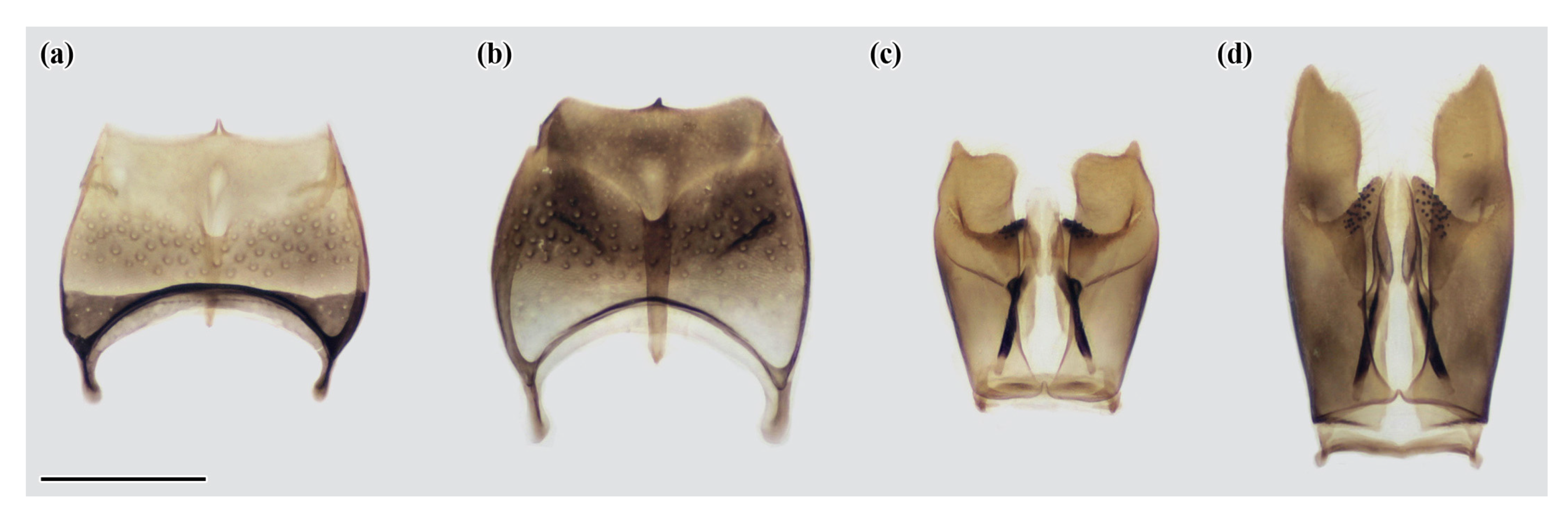
Appendix A. Re-Description of Celonites kozlovi Kostylev, 1935


Appendix B. Collected Material
References
- Rahmani, Z.; Rakhshani, E.; Carpenter, J.M. Updated checklist of Vespidae (Hymenoptera: Vespoidea) in Iran. J. Insect Biodivers. Syst. 2020, 6, 27–86. [Google Scholar] [CrossRef]
- Gess, S.K. The Pollen Wasps. Ecology and Natural History of the Masarinae; Harvard University Press: Cambridge, MA, USA; London, UK, 1996; pp. pp. i–x + 1–340. [Google Scholar] [CrossRef]
- Gess, S.K.; Gess, F.W. Pollen Wasps and Flowers in Southern Africa. SANBI Biodiversity Series 18; South African National Biodiversity Institute: Pretoria, South Africa, 2010; pp. 1–147. [Google Scholar]
- Carpenter, J.M.; Garcete-Barrett, B.R.; Hermes, M.G. Catalog of the Neotropical Masarinae (Hymenoptera, Vespidae). Rev. Bras. Entomol. 2006, 50, 335–340. [Google Scholar] [CrossRef][Green Version]
- Richards, O.W. A Revisional Study of the Masarid Wasps (Hymenoptera, Vespoidea); British Museum of Natural History: London, UK, 1962. [Google Scholar]
- Mauss, V.; Kuba, K.; Krenn, H.W. Evolution of the multifunctional mouthparts of adult Vespidae. In Insect Mouthparts. Zoological Monographs 5; Krenn, H.W., Ed.; Springer: Cham, Switzerland, 2019; pp. 443–478. [Google Scholar] [CrossRef]
- Carpenter, J.M. The phylogenetic relationship and natural classification of the Vespoidea (Hymenoptera). Syst. Entomol. 1982, 7, 11–38. [Google Scholar] [CrossRef]
- Carpenter, J.M. The phylogenetic system of the Gayellini (Hymenoptera: Vespidae; Masarinae). Psyche 1988, 95, 211–241. [Google Scholar] [CrossRef]
- Mauss, V. Evolution verschiedener Lebensformtypen innerhalb basaler Teilgruppen der Faltenwespen (Hymenoptera, Vespidae). Denisia 2007, 20, 701–722. [Google Scholar]
- Piekarski, P.K.; Carpenter, J.M.; Lemmon, A.R.; Lemmon, E.M.; Sharanowski, B.J. Phylogenomic evidence overturns current conceptions of social evolution in wasps (Vespidae). Mol. Biol. Evol. 2018, 35, 2097–2109. [Google Scholar] [CrossRef]
- Gess, S.K.; Gess, F.W. Flower visiting by masarid wasps in southern Africa (Hymenoptera: Vespoidea: Masaridae). Ann. Cape Prov. Mus. Nat. Hist. 1989, 18, 95–134. [Google Scholar]
- Gess, S.K.; Gess, F.W.; Gess, R.W. Update on the flower associations of southern African Masarinae with notes on the nesting of Masarina strucki Gess and Celonites gariepensis Gess (Hymenoptera: Vespidae: Masarinae). J. Hymenopt. Res. 1997, 6, 75–91. [Google Scholar]
- Gess, S.K.; Gess, F.W. Distributions of flower associations of pollen wasps (Vespidae: Masarinae) in southern Africa. J. Arid Environ. 2004, 57, 17–44. [Google Scholar] [CrossRef]
- Carpenter, J.M. Checklist of species of the subfamily Masarinae (Hymenoptera: Vespidae). Amer. Mus. Novit. 2001, 3325, 1–39. [Google Scholar] [CrossRef]
- Fateryga, A.V.; Fadeev, K.I. The identity of Celonites montanus Mocsáry, 1906 (Hymenoptera: Vespidae: Masarinae) and its first record from Kazakhstan. Zootaxa 2023, submitted.
- Mauss, V.; Müller, A. First contribution to the bionomics of the pollen wasp Celonites fischeri Spinola, 1838 (Hymenoptera, Vespidae, Masarinae) in Cyprus. J. Hymenopt. Res. 2014, 39, 119–153. [Google Scholar] [CrossRef]
- Mauss, V.; Fateryga, A.V.; Yildirim, E.; Carpenter, J.M. Contribution to the taxonomy, bionomics and distribution of the Palaearctic Celonites cyprius-group (Hymenoptera, Vespidae, Masarinae) with the description of two new species from the North Caucasus and East Anatolia. J. Hymenopt. Res. 2022, 89, 109–155. [Google Scholar] [CrossRef]
- Fateryga, A.V.; Mauss, V.; Fateryga, V.V. New distributional records of Celonites tauricus (Hymenoptera, Vespidae, Masarinae) and new data on its behaviour at flowers. J. Hymenopt. Res. 2022, 92, 241–256. [Google Scholar] [CrossRef]
- Mauss, V.; Praz, C.J.; Müller, A.; Prosi, R.; Rosa, P. Description of the nest of the pollen wasp Celonites jousseaumei Du Buysson, 1906 (Hymenoptera, Vespidae, Masarinae) with a new host association of the cuckoo wasp Spintharina innesi (Du Buysson, 1894) (Hymenoptera, Chrysididae). J. Hymenopt. Res. 2022, 93, 139–149. [Google Scholar] [CrossRef]
- Kostylev, G. Materialien zur Kenntnis der Masariden-Fauna der Paläarktis. Arch. Muz. Zool. Univ. Moscou 1935, 2, 85–116, (In Russian and German). [Google Scholar]
- Gusenleitner, J. Über Vespoidea (Hymenoptera) aus der Mongolei und der Sovietunion. Linz. Biol. Beitr. 1991, 23, 631–641. [Google Scholar]
- Gusenleitner, J. Eine neue Celonites-Art aus Sibirien (Hymenoptera: Vespidae, Masarinae). Linz. Biol. Beitr. 2007, 39, 133–135. [Google Scholar]
- Fateryga, A.V. New records of Celonites kozlovi Kostylev, 1935 and C. sibiricus Gusenleitner, 2007 (Hymenoptera: Vespidae: Masarinae), with observations on their behavior at flowers. Far East. Entomol. 2020, 405, 20–32. [Google Scholar] [CrossRef]
- Müller, A.; Kuhlmann, M. Pollen hosts of western palaearctic bees of the genus Colletes (Hymenoptera: Colletidae): The Asteraceae paradox. Biol. J. Linn. Soc. 2008, 95, 719–733. [Google Scholar] [CrossRef]
- Faegri, K.; van der Pijl, L. The Principles of Pollination Ecology, 3rd ed.; Pergamon Press: Oxford, UK; New York, NY, USA; Toronto, ON, Canada; Sidney, Australia; Paris, France; Frankfurt, Germany, 1979. [Google Scholar]
- Müller, A. Convergent evolution of morphological specializations in Central European bee and honey wasp species as an adaptation to the uptake of pollen from nototribic flowers (Hymenoptera, Apoidea and Masaridae). Biol. J. Linn. Soc. 1996, 57, 235–252. [Google Scholar] [CrossRef]
- Seregin, A.P. Moscow University Herbarium (MW). In Occurrence Dataset; Seregin, A.P., Ed.; Version 1.58; Lomonosov Moscow State University: Moscow, Russia, 2023. [Google Scholar] [CrossRef]
- Krasnoborov, I.M.; Artemov, I.A. Key to Plants of the Altai Republic; Krasnoborov, I.M., Artemov, I.A., Eds.; Siberian Branch of the Russian Academy of Sciences Press: Novosibirsk, Russia, 2012. (In Russian) [Google Scholar]
- Plantarium. Plants and Lichens of Russia and Neighboring Countries: Open Online Galleries and Plant Identification Guide. Available online: http://www.plantarium.ru/ (accessed on 14 February 2023).
- POWO. Plants of the World Online; Royal Botanic Gardens: Kew, UK, 2021; Available online: http://www.plantsoftheworldonline.org/ (accessed on 14 February 2023).
- Kirschner, J.; Štěpánek, J. Dandelions in Central Asia: A revision of Taraxacum section Stenoloba. Preslia 2011, 83, 491–512. [Google Scholar]
- Inouye, D.W. The terminology of flower larceny. Ecology 1980, 61, 1251–1253. [Google Scholar] [CrossRef]
- Mauss, V. Description of Celonites andreasmuelleri sp. n. (Hymenoptera, Vespidae, Masarinae) from the Middle East with a key to the Palaearctic species of the C. abbreviatus-complex of the subgenus Celonites s. str. J. Hymenopt. Res. 2013, 31, 79–95. [Google Scholar] [CrossRef][Green Version]
- Schremmer, F. Der bisher unbekannte Pollensammelapparat der Honigwespe Celonites abbreviatus Vill. (Vespidae, Masarinae). Z. Morphol. Ökol. Tiere 1959, 48, 424–438. [Google Scholar] [CrossRef]
- Mauss, V. Observations on flower associations and mating behaviour of the pollen wasp species Celonites abbreviatus (Villers, 1789) in Greece (Hymenoptera: Vespidae, Masarinae). J. Hymenopt. Res. 2006, 15, 266–269. [Google Scholar]
- Mauss, V.; Fateryga, A.V.; Prosi, R. Taxonomy, distribution and bionomics of Celonites tauricus Kostylev, 1935, stat. n. (Hymenoptera, Vespidae, Masarinae). J. Hymenopt. Res. 2016, 48, 33–66. [Google Scholar] [CrossRef]
- Popov, V.B. Oligotrophism of species of the genus Quartinia Grib. (Hymenoptera, Vespoidea). Zool. Zhurnal 1948, 27, 317–328. (In Russian) [Google Scholar]
- Haeseler, V. Zum Necktarraub solitärer Faltenwespen (Hymenoptera: Vespoidea: Eumenidae). Entomol. Gen. 1980, 6, 49–55. [Google Scholar] [CrossRef]
- Haeseler, V. Ancistrocerus oviventris (Wesmael 1836), eine weitere Nektar raubende solitäre Faltenwespe (Hymenoptera: Vespoidea: Eumenidae). Faun.-Ökol. Mitt. 1997, 7, 259–266. [Google Scholar]
- Fateryga, A.V.; Podunay, Y.A. Nesting and biology of Alastor mocsaryi (Hymenoptera, Vespidae: Eumeninae). Entomol. Rev. 2018, 98, 1006–1016. [Google Scholar] [CrossRef]
- Fateryga, A.V.; Popovich, A.V.; Podunay, Y.A.; Fateryga, V.V. First data on the bionomics of Leptochilus (Euleptochilus) limbiferus (Morawitz, 1867) (Hymenoptera: Vespidae: Eumeninae), with taxonomic notes and new records. Zootaxa 2020, 4851, 289–304. [Google Scholar] [CrossRef] [PubMed]
- Fateryga, A.V. Two new Nearctic genera in the tribe Odynerini s. str. revealed on the bionomics and morphology, with a comment on the cocoons of the eumenine wasps (Hymenoptera: Vespidae: Eumeninae). Far East. Entomol. 2021, 427, 1–19. [Google Scholar] [CrossRef]
- Müller, A.; Westrich, P. Morphological specialisation for primary nectar robbing in a pollen specialist mining bee (Hymenoptera, Andrenidae). J. Hymenopt. Res. 2023, 95, 215–230. [Google Scholar] [CrossRef]
- Mauss, V.; Prosi, R. Identity and distribution of Celonites hermon Gusenleitner, 2002 (Hymenoptera, Vespidae, Masarinae) from the Middle East with a description of the hitherto unknown male. J. Hymenopt. Res. 2018, 66, 55–70. [Google Scholar] [CrossRef]
- Praz, C.J.; Müller, A.; Dorn, S. Specialized bees fail to develop on non-host pollen: Do plants chemically protect their pollen? Ecology 2008, 89, 795–804. [Google Scholar] [CrossRef] [PubMed]
- Sedivy, C.; Müller, A.; Dorn, S. Closely related pollen generalist bees differ in their ability to develop on the same pollen diet: Evidence for physiological adaptations to digest pollen. Funct. Ecol. 2011, 25, 718–725. [Google Scholar] [CrossRef]
- Müller, A. Host-plant specialization in Western Palearctic anthidiine bees (Hymenoptera: Apoidea: Megachilidae). Ecol. Monogr. 1996, 66, 235–257. [Google Scholar] [CrossRef]
- Mauss, V.; Müller, A.; Prosi, R. Flower associations and nesting of the pollen wasp Quartinia major Kohl, 1898 (Hymenoptera, Vespidae, Masarinae) in Morocco. J. Hymenopt. Res. 2018, 62, 15–31. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Locality Name | Latitude | Longitude | Date | Duration of Observations (hours) |
|---|---|---|---|---|
| Tydtuyaryk River valley | 50°04′25″ N | 88°25′12″ E | 12.VI.2022 | 1 |
| 18.VI.2022 | 2 | |||
| 21.VI.2022 | 2 | |||
| 22.VI.2022 | 2 | |||
| 23.VI.2022 | 1 | |||
| 26.VI.2022 | 0.25 | |||
| Mars Natural Landmark | 50°03′50″ N | 88°18′45″ E | 17.VI.2022 | 0.75 |
| 23.VI.2022 | 1.5 | |||
| 25.VI.2022 | 0.5 | |||
| 3 km to the north-east of Kokorya | 49°56′31″ N | 89°02′45″ E | 24.VI.2022 | 0.5 |
| 5 km to the north-east of Kokorya | 49°57′00″ N | 89°04′19″ E | 24.VI.2022 | 0.5 |
| 26.VI.2022 | 1 | |||
| Total | 13 |
| Plant Taxon | Σ Sightings of Flower-Visiting Individuals | |||
|---|---|---|---|---|
| C. kozlovi | C. sibiricus | |||
| ♀ | ♂ | ♀ | ♂ | |
| Asteraceae | ||||
| Crepidiastrum tenuifolium (Willd.) Sennikov | 2 | |||
| Klasea marginata (Tausch) Kitag. | 41 | 1 | ||
| Pseudopodospermum pubescens (DC.) Zaika, Sukhor. & N. Kilian | 26 | |||
| Taraxacum sp. | 10 | 2 | 2 | |
| Brassicaceae | ||||
| Sisymbrium polymorphum (Murray) Roth | 7 | 3 | 6 | |
| Caryophyllaceae | ||||
| Dianthus chinensis L. | 2 | |||
| Gypsophila patrinii Ser. | 2 | 1 | ||
| Convolvulaceae | ||||
| Convolvulus ammannii Desr. | 1 | |||
| Fabaceae | ||||
| Caragana bungei Ledeb. | 6 | |||
| Lamiaceae | ||||
| Dracocephalum peregrinum L. | 66 | 24 | 9 | |
| Ziziphora clinopodioides Lam. | 18 | 1 | ||
| Rosaceae | ||||
| Potentilla sericea L. | 1 | |||
| Sibbaldianthe bifurca (L.) Kurtto & T. Erikss. | 5 | |||
| Other plant taxa * | ||||
| Total | 184 | 1 | 33 | 18 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fateryga, A.V.; Mauss, V.; Proshchalykin, M.Y. Foraging Behavior of Two Pollen Wasp Species of the Genus Celonites Latreille, 1802 (Hymenoptera: Vespidae: Masarinae), from the Altai Mountains. Insects 2023, 14, 408. https://doi.org/10.3390/insects14050408
Fateryga AV, Mauss V, Proshchalykin MY. Foraging Behavior of Two Pollen Wasp Species of the Genus Celonites Latreille, 1802 (Hymenoptera: Vespidae: Masarinae), from the Altai Mountains. Insects. 2023; 14(5):408. https://doi.org/10.3390/insects14050408
Chicago/Turabian StyleFateryga, Alexander V., Volker Mauss, and Maxim Yu. Proshchalykin. 2023. "Foraging Behavior of Two Pollen Wasp Species of the Genus Celonites Latreille, 1802 (Hymenoptera: Vespidae: Masarinae), from the Altai Mountains" Insects 14, no. 5: 408. https://doi.org/10.3390/insects14050408
APA StyleFateryga, A. V., Mauss, V., & Proshchalykin, M. Y. (2023). Foraging Behavior of Two Pollen Wasp Species of the Genus Celonites Latreille, 1802 (Hymenoptera: Vespidae: Masarinae), from the Altai Mountains. Insects, 14(5), 408. https://doi.org/10.3390/insects14050408