
Do Patches of Flowering Plants Enhance Insect Pollinators in Apple Orchards?





Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Orchards and Design
2.2. Establishment of Flower Patches
2.3. Landscape Elements around the Experimental Orchards
2.4. Flowering and Insect Presence Measurements
2.5. Statistical Analysis
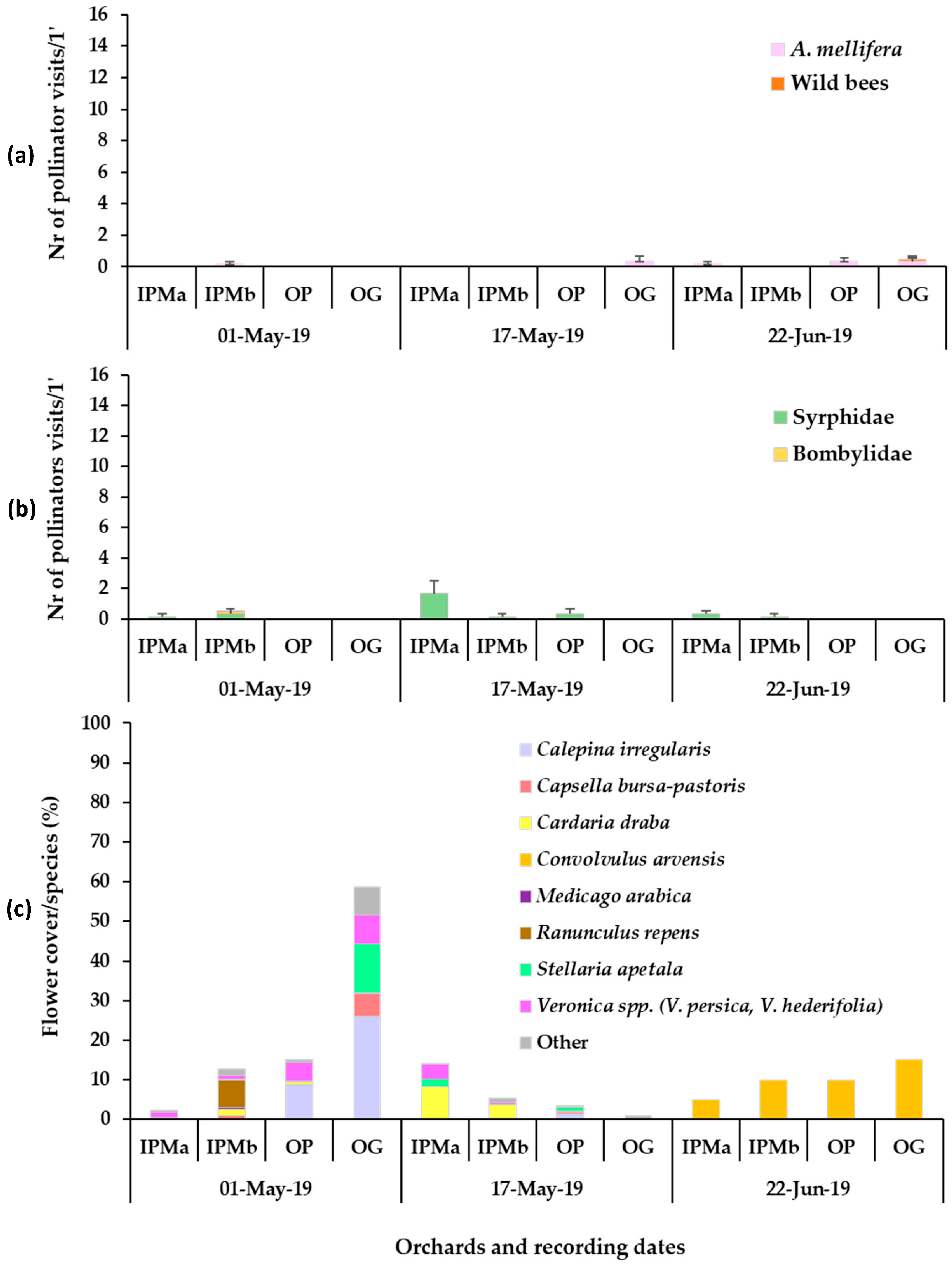
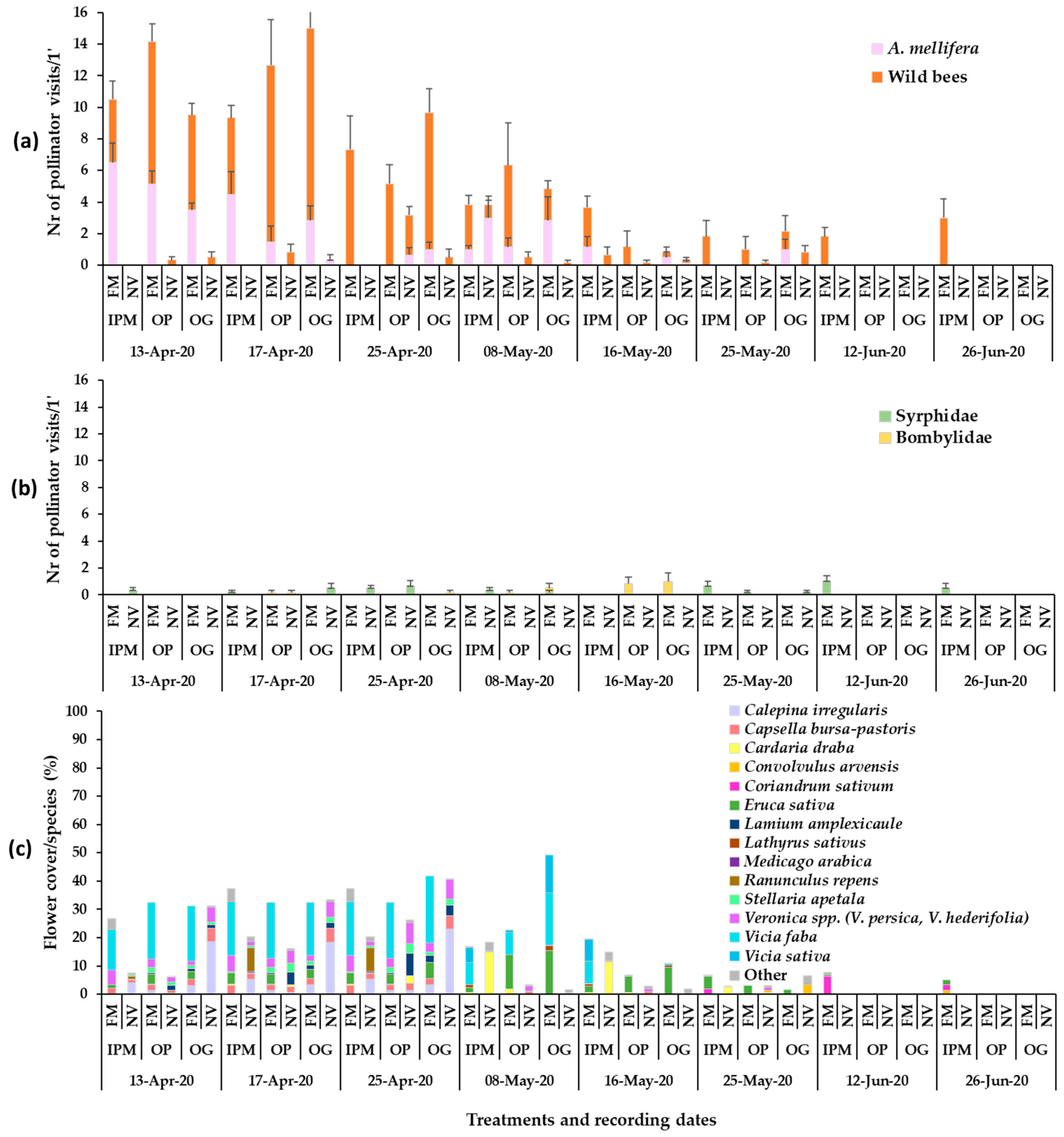
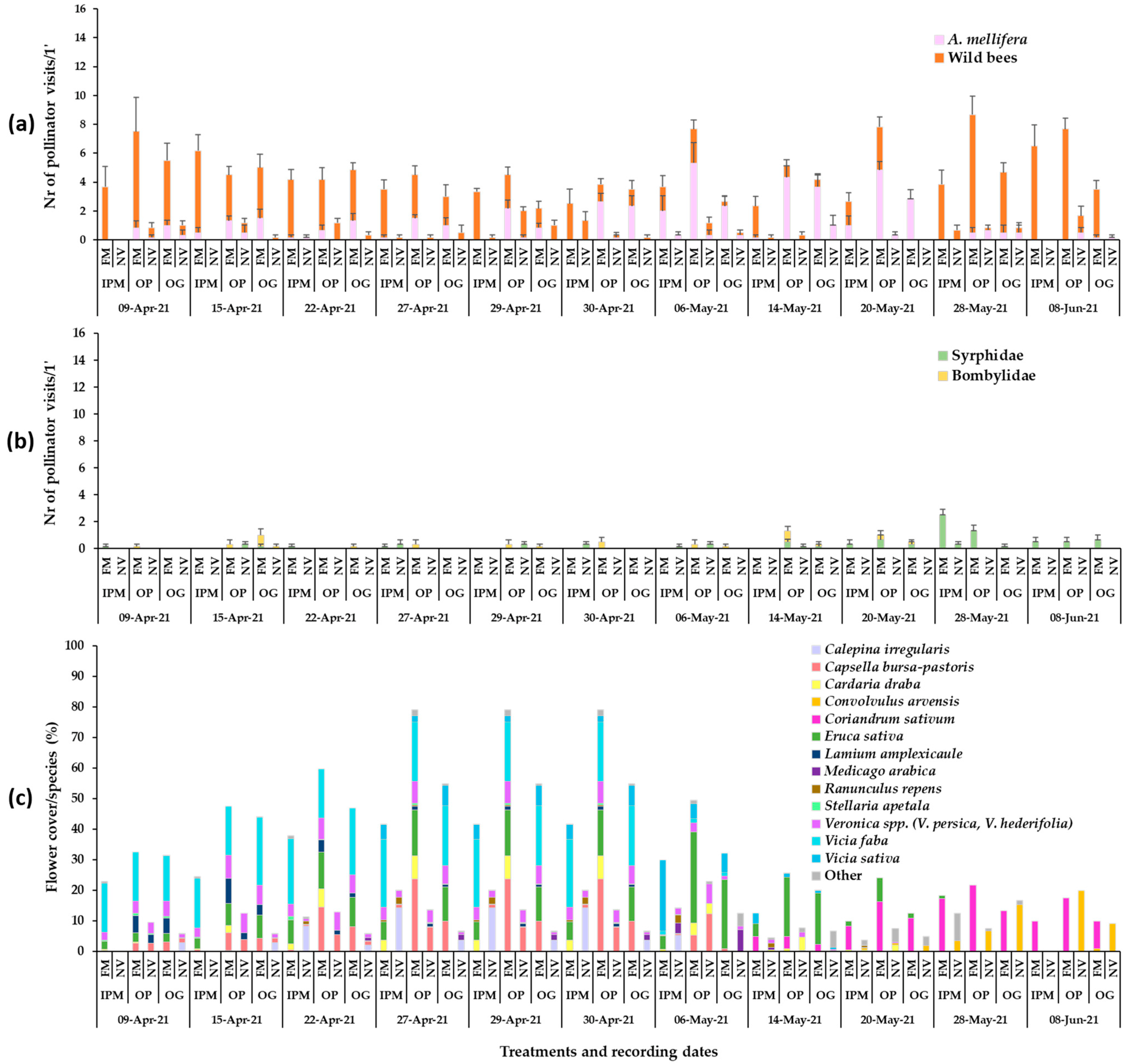
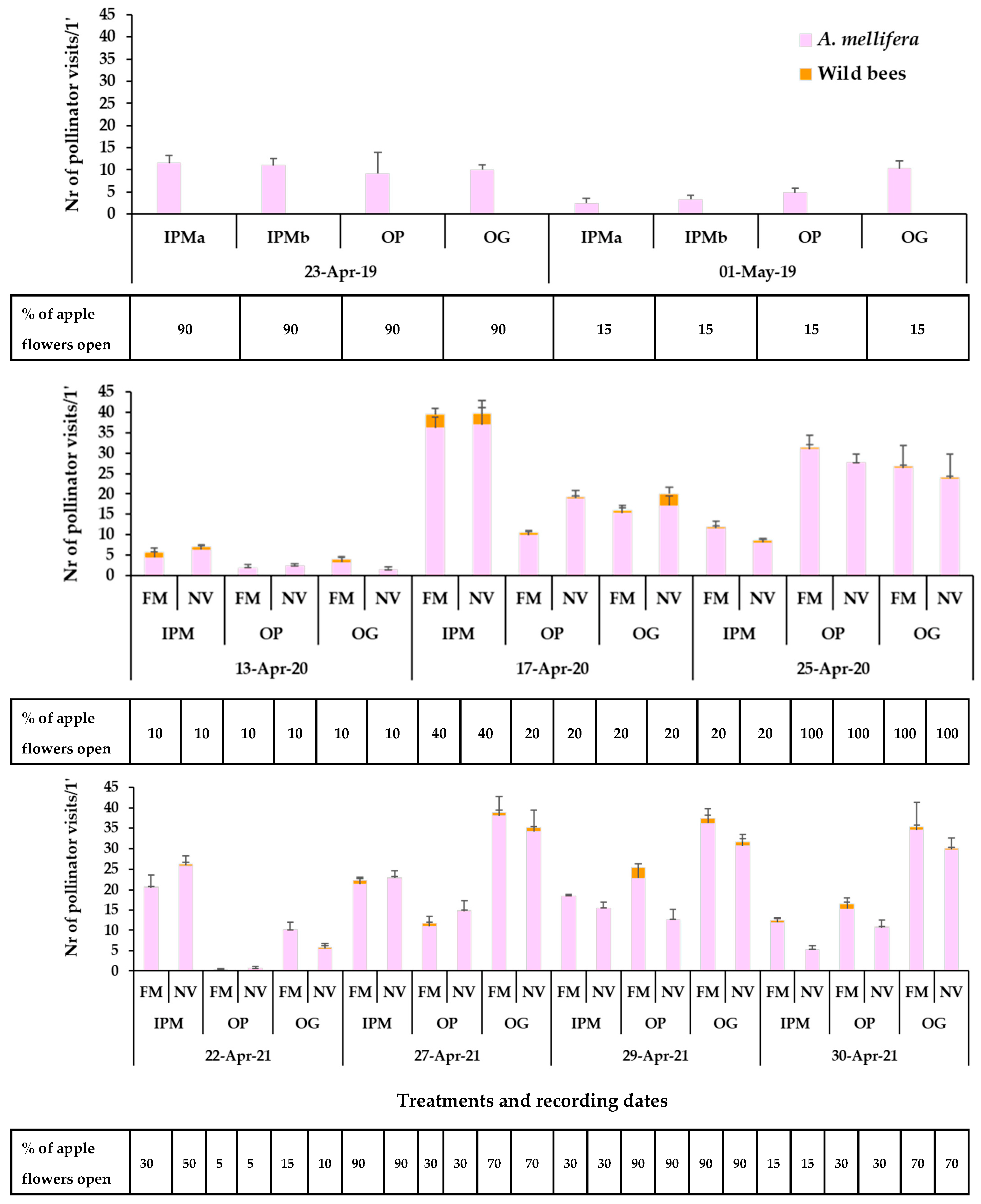
3. Results
3.1. Plant Mixture Establishment and Flowering
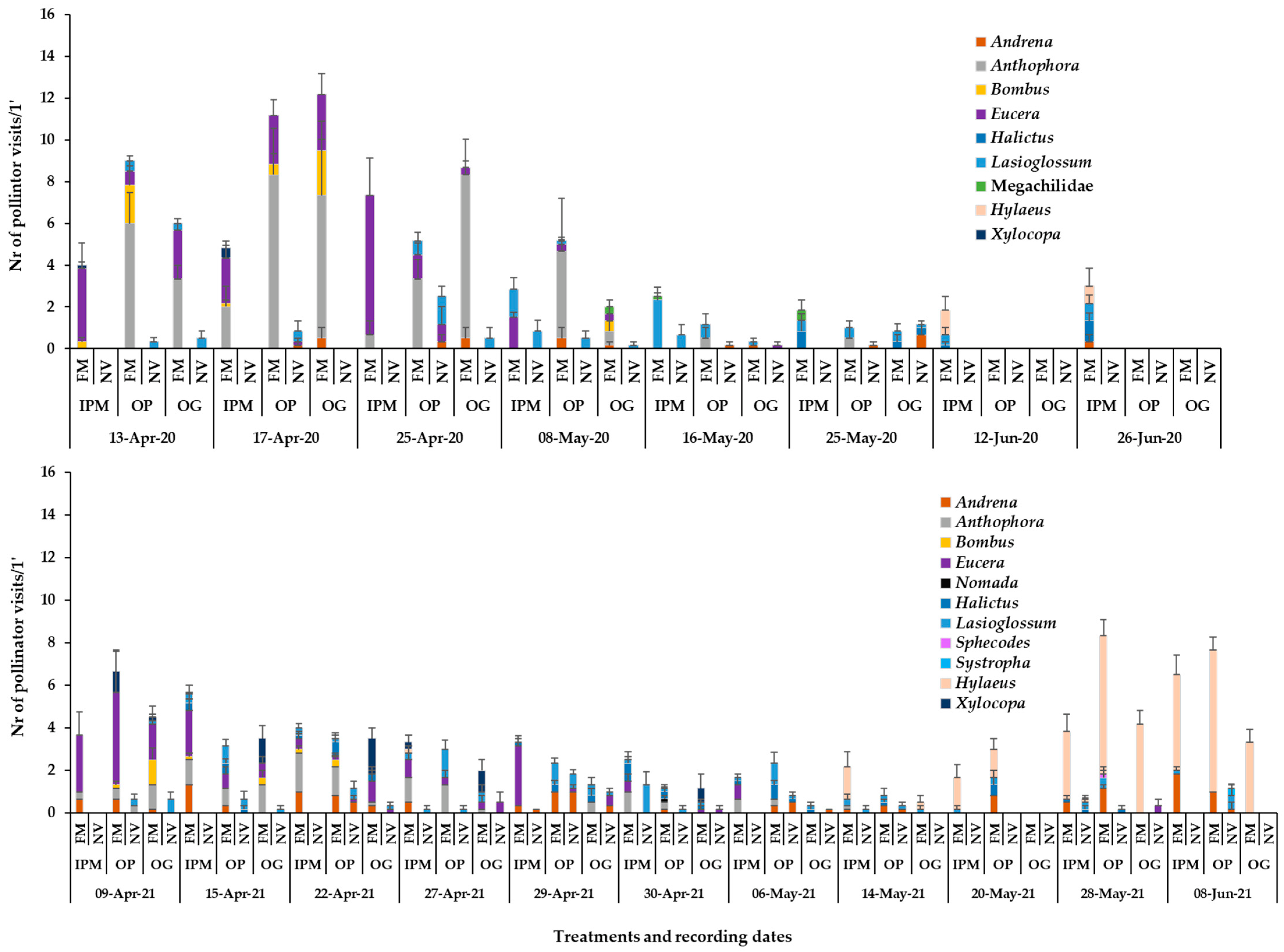
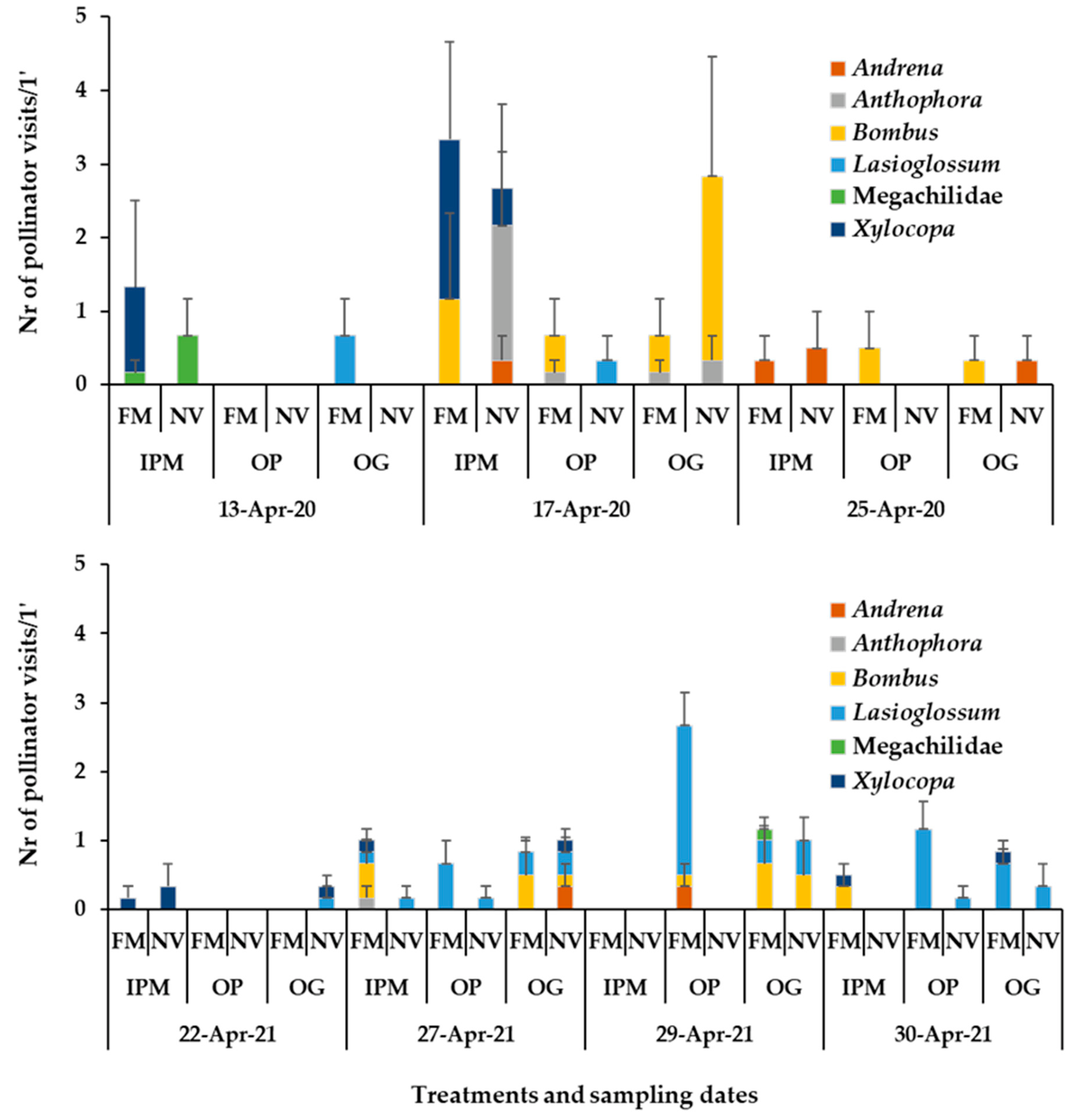
3.2. Presence of Pollinators in Sown/Wild Plant Patches and on Apple Blossom
3.3. Bee Fauna Composition and Other Main Fauna of Flower Visitors in Association to Flower Species
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Losey, J.E.; Vaughan, M. The economic value of ecological services provided by insects. Bioscience 2006, 56, 311–323. [Google Scholar] [CrossRef]
- Kremen, C.; Williams, N.M.; Aizen, M.A.; Gemmill-Herren, B.; LeBuhn, G.; Minckley, R.; Packer, L.; Potts, S.G.; Roulston, T.A.; Steffan-Dewenter, I.; et al. Pollination and other ecosystem services produced by mobile organisms: A conceptual framework for the effects of land-use change. Ecol. Lett. 2007, 10, 299–314. [Google Scholar] [CrossRef]
- Porto, R.G.; de Almeida, R.F.; Cruz-Neto, O.; Tabarelli, M.; Viana, B.F.; Peres, C.A.; Lopes, A.V. Pollination ecosystem services: A comprehensive review of economic values, research funding and policy actions. Food Secur. 2020, 12, 1425–1442. [Google Scholar] [CrossRef]
- Ollerton, J.; Winfree, R.; Tarrant, S. How many flowering plants are pollinated by animals? Oikos 2011, 120, 321–326. [Google Scholar] [CrossRef]
- Potts, S.G.; Imperatriz Fonseca, V.; Ngo, H.T.; Biesmeijer, J.C.; Breeze, T.D.; Dicks, L.; Garibaldi, L.A.; Hill, R.; Settele, J.; Vanbergen, A.J.; et al. Summary for Policymakers of the Assessment Report of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services on Pollinators, Pollination and Food Production; Secretariat of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services: Bonn, Germany, 2016; 36p. [Google Scholar]
- Kearns, C.A.; Inouye, D.; Waser, N.M. Endangered mutualisms: The conservation of plant-pollinator interactions. Annu. Rev. Ecol. Syst. 1998, 29, 83–112. [Google Scholar] [CrossRef]
- Garibaldi, L.A.; Steffan-Dewenter, I.; Kremen, C.; Morales, J.M.; Bommarco, R.; Cunningham, S.A.; Carvalheiro, L.G.; Chacoff, N.P.; Dudenhöffer, J.H.; Greenleaf, S.S.; et al. Stability of pollination services decreases with isolation from natural areas despite honey bee visits. Ecol. Lett. 2011, 14, 1062–1072. [Google Scholar] [CrossRef]
- Isaacs, R.; Tuell, J.; Fiedler, A.; Gardiner, M.; Landis, D. Maximizing arthropod-mediated ecosystem services in agricultural landscapes: The role of native plants. Front. Ecol. Environ. 2009, 7, 196–203. [Google Scholar] [CrossRef]
- Bommarco, R.; Kleijn, D.; Potts, S.G. Ecological intensification: Harnessing ecosystem services for food security. Trends Ecol. Evol. 2013, 28, 230–238. [Google Scholar] [CrossRef]
- Potts, S.G.; Imperatriz-Fonseca, V.; Ngo, H.T.; Aizen, M.A.; Biesmeijer, J.C.; Breeze, T.D. Safeguarding pollinators and their values to human well-being. Nature 2016, 540, 220–229. [Google Scholar] [CrossRef]
- Johnson, C.A.; Dutt, P.; Levine, J.M. Competition for pollinators destabilizes plant coexistence. Nature 2022, 607, 721–725. [Google Scholar] [CrossRef]
- Biesmeijer, J.C.; Roberts, S.P.; Reemer, M.; Ohlemuller, R.; Edwards, M.; Peeters, T.; Schaffers, A.P.; Potts, S.G.; Kleukers, R.J.M.C.; Thomas, C.D.; et al. Parallel declines in pollinators and insect-pollinated plants in Britain and the Netherlands. Science 2006, 313, 351–354. [Google Scholar] [CrossRef]
- Potts, S.G.; Biesmeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global pollinator declines: Trends, impacts and drivers. Trends Ecol. Evol. 2010, 25, 345–353. [Google Scholar] [CrossRef]
- Meek, B.; Loxton, D.; Sparks, T.; Pywell, R.; Pickett, H.; Nowakowski, M. The effect of arable field margin composition on invertebrate biodiversity. Biol. Conserv. 2002, 106, 259–271. [Google Scholar] [CrossRef]
- Potts, S.G.; Vulliamy, B.; Dafni, A.; Ne’eman, G.; Willmer, P. Linking Bees and Flowers: How Do Floral Communities Structure Pollinator Communities? Ecology 2003, 84, 2628–2642. [Google Scholar] [CrossRef]
- Michener, C.D. The Bees of the World, 2nd ed.; Johns Hopkins University Press: Baltimore, MD, USA, 2007. [Google Scholar]
- Van Rijn, P.C.J.; Kooijman, J.; Wäckers, F.L. The contribution of floral resources and honeydew to the performance of predatory hoverflies (Diptera: Syrphidae). Biol. Control 2013, 67, 32–38. [Google Scholar] [CrossRef]
- Goulson, D.; Hanley, M.E.; Darvill, B.; Ellis, J.S.; Knight, M.E. Causes of rarity in bumblebees. Biol. Conserv. 2005, 122, 1–8. [Google Scholar] [CrossRef]
- Winfree, R.; Aguilar, R.; Vázquez, D.P.; LeBuhn, G.; Aizen, M.A. A meta-analysis of bees’ responses to anthropogenic disturbance. Ecology 2009, 90, 2068–2076. [Google Scholar] [CrossRef]
- Bommarco, R.; Lundin, O.; Smith, H.G.; Rundlöf, M. Drastic historic shifts in bumble-bee community composition in Sweden. Proc. R. Soc. B Biol. Sci. 2012, 279, 309–315. [Google Scholar] [CrossRef]
- Schindler, M.; Diestelhorst, O.; Härtel, S.; Saure, C.; Schanowski, A.; Schwenninger, H.R. Monitoring agricultural ecosystems by using wild bees as environmental indicators. BioRisk 2013, 8, 53–71. [Google Scholar] [CrossRef]
- Baude, M.; Kunin, W.E.; Boatman, N.D.; Conyers, S.; Davies, N.; Gillespie, M.A.K.; Morton, R.D.; Smart, S.M.; Memmott, J. Historical nectar assessment reveals the fall and rise of floral resources in Britain. Nature 2016, 530, 85–88. [Google Scholar] [CrossRef]
- Cole, L.J.; Brocklehurst, S.; Robertson, D.; Harrison, W.; McCracken, D.I. Exploring the interactions between resource availability and the utilisation of semi-natural habitats by insect pollinators in an intensive agricultural landscape. Agric. Ecosyst. Environ. 2017, 246, 157–167. [Google Scholar] [CrossRef]
- Nieto, A.; Roberts, S.P.M.; Kemp, J.; Rasmont, P.; Kuhlmann, M.; García Criado, M.; Biesmeijer, J.C.; Bogusch, P.; Dathe, H.H.; De la Rúa, P.; et al. European Red List of Bees; Publication Office of the European Union: Luxembourg, 2014. [CrossRef]
- Reilly, J.R.; Artz, D.R.; Biddinger, D.; Bobiwash, K.; Boyle, N.K.; Brittain, C.; Brokaw, J.; Campbell, J.W.; Daniels, J.; Elle, E.; et al. Crop production in the USA is frequently limited by a lack of pollinators. Proc. R. Soc. B 2020, 287, 20200922. [Google Scholar] [CrossRef]
- Petanidou, T.; Kallimanis, A.S.; Tzanopoulos, J.; Sgardelis, S.P.; Pantis, J.D. Long-term observation of a pollination network: Fluctuation in species and interactions, relative invariance of network structure and implications for estimates of specialization. Ecol. Lett. 2008, 11, 564–575. [Google Scholar] [CrossRef]
- Patiny, S.; Rasmont, P.; Michez, D. A survey and review of the status of wild bees in the West-Palaearctic region. Apidologie 2009, 40, 313–331. [Google Scholar] [CrossRef]
- Petanidou, T.; Ståhls, G.; Vujić, A.; Olesen, J.M.; Rojo, S.; Thrasyvoulou, A.; Sgardelis, S.; Kallimanis, A.S.; Kokkini, S.; Tscheulin, T. Investigating plant—Pollinator relationships in the Aegean: The approaches of the project POL-AEGIS (The pollinators of the Aegean archipelago: Diversity and threats). J. Apic. Res. 2013, 52, 106–117. [Google Scholar] [CrossRef]
- Melin, A.; Rouget, M.; Midgley, J.J.; Donaldson, J.S. Pollination ecosystem services in South African agricultural systems. S. Afr. J. Sci. 2014, 110, 1–9. [Google Scholar] [CrossRef]
- Meiners, J.M.; Griswold, T.L.; Carril, O.M. Decades of native bee biodiversity surveys at Pinnacles National Park highlight the importance of monitoring natural areas over time. PLoS ONE 2019, 14, e0207566. [Google Scholar] [CrossRef]
- Garibaldi, L.A.; Aizen, M.A.; Cunningham, S.; Klein, A.M. Pollinator shortage and global crop yield: Looking at the whole spectrum of pollinator dependency. Commun. Integr. Biol. 2009, 2, 37–39. [Google Scholar] [CrossRef]
- Sagili, R.R.; Burgett, D.M. Evaluating Honey Bee Colonies for Pollination A Guide for Commercial Growers and Beekeepers; Oregon State University: Corvallis, OR, USA, 2011; Available online: https://catalog.extension.oregonstate.edu/pnw623 (accessed on 1 January 2023).
- DeGrandi-Hoffman, G.; Graham, H.; Ahumada, F.; Smart, M.; Ziolkowski, N. The Economics of Honey Bee (Hymenoptera: Apidae) Management and Overwintering Strategies for Colonies Used to Pollinate Almonds. J. Econ. Entomol. 2019, 112, 2524–2533. [Google Scholar] [CrossRef]
- Devkota, K.; Rijal, P.; Fernando dos Santos, C. The Use of Honeybee Hives May Boost Yields of Some Crops in Nepal. Psyche A J. Entomol. 2021, 2021, 8876388. [Google Scholar] [CrossRef]
- Breeze, T.D.; Vaissière, B.E.; Bommarco, R.; Petanidou, T.; Seraphides, N.; Kozák, L.; Scheper, J.; Biesmeijer, J.C.; Kleijn, D.; Gyldenkærne, S.; et al. Agricultural Policies Exacerbate Honeybee Pollination Service Supply-Demand Mismatches Across Europe. PLoS ONE 2014, 9, e82996. [Google Scholar] [CrossRef]
- Westerkamp, C.; Gottsberger, G. The Costly Crop Pollination Crisis. Pollinating Bees—The Conservation Link between Agriculture and Nature; Kevan, P., Fonseca, V.I., Eds.; Ministry of Environment: Brasilia, Brazil, 2022; pp. 51–56.
- Bond, J.; Plattner, K.; Hunt, K. Pollination-Services Market, Fruit and Tree Nuts Outlook; Situation and Outlook Report No. FTS-357SA; US Department of Agriculture: Washington, DC, USA; Economic Research Service: Washington, DC, USA, 2014. Available online: https://www.ers.usda.gov/publications/pub-details/?pubid=37060 (accessed on 21 December 2022).
- Winfree, R.; Williams, N.M.; Dushoff, J.; Kremen, C. Native bees provide insurance against ongoing honey bee losses. Ecol. Lett. 2007, 10, 1105–1113. [Google Scholar] [CrossRef]
- O’Toole, C. Mason Bees as Managed Pollinators of Orchard Crops. Bee World 2011, 88, 5–8. [Google Scholar] [CrossRef]
- Garibaldi, L.A.; Steffan-Dewenter, I.; Winfree, R.; Aizen, M.A.; Bommarco, R.; Cunningham, S.A.; Kremen, C.; Carvalheiro, L.G.; Harder, L.D.; Afik, O.; et al. Wild Pollinators Enhance Fruit Set of Crops Regardless of Honey Bee Abundance. Science 2013, 339, 1608–1611. [Google Scholar] [CrossRef]
- Pérez-Méndez, N.; Andersson, G.K.S.; Requier, F.; Hipólito, J.; Aizen, M.A.; Morales, C.L.; García, N.; Gennari, G.P.; Garibaldi, L.A. The economic cost of losing native pollinator species for orchard production. J. Appl. Ecol. 2019, 57, 599–608. [Google Scholar] [CrossRef]
- Nicholls, C.I.; Altieri, M.A. Plant biodiversity enhances bees and other insect pollinators in agroecosystems. A review. Agron. Sustain. Dev. 2012, 33, 257–274. [Google Scholar] [CrossRef]
- Dicks, L.V.; Abrahams, A.; Atkinson, J.; Biesmeijer, J.; Bourn, N.; Brown, C.; Brown, M.J.; Carvell, C.; Connolly, C.; Cresswell, J.E.; et al. Identifying key knowledge needs for evidence-based conservation of wild insect pollinators: A collaborative cross-sectoral exercise. Insect Conserv. Divers. 2013, 6, 435–446. [Google Scholar] [CrossRef]
- Giovanetti, M.; Albertazzi, S.; Flaminio, S.; Ranalli, R.; Bortolotti, L.; Quaranta, M. Pollination in Agroecosystems: A Review of the Conceptual Framework with a View to Sound Monitoring. Land 2021, 10, 540. [Google Scholar] [CrossRef]
- Mottershead, D.; Underwood, E. Pollinators in the CAP: Integrating Pollinator Conservation into the Common Agricultural Policy; Institute for European Environmental Policy: Brussels, Belgium, 2020. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations. FAO’s Global Action on Pollination Services for Sustainable Agriculture, Global Pollination Project. Available online: https://www.fao.org/common-pages/search/en/?q=pollinators (accessed on 19 July 2022).
- European Commission. Eurostat Regional Yearbook: 2020; European Commission: Brussels, Belgium, 2020.
- Nielsen, A.; Steffan-Dewenter, I.; Westphal, C.; Messinger, O.; Potts, S.G.; Roberts, S.P.; Settele, J.; Szentgyörgyi, H.; Vaissière, B.E.; Vaitis, M.; et al. Assessing bee species richness in two Mediterranean communities: Importance of habitat type and sampling techniques. Ecol. Res. 2011, 26, 969–983. [Google Scholar] [CrossRef]
- Drossart, M.; Gérard, M. Beyond the Decline of Wild Bees: Optimizing Conservation Measures and Bringing Together the Actors. Insects 2020, 11, 649. [Google Scholar] [CrossRef]
- Krahner, A.; Schmidt, J.; Maixner, M.; Porten, M.; Schmitt, T. Evaluation of four different methods for assessing bee diversity as ecological indicators of agro-ecosystems. Ecol. Indic. 2021, 125, 107573. [Google Scholar] [CrossRef]
- Brandl, M.; Hussain, R.I.; Maas, B.; Rabl, D.; Pachinger, B.; Holzinger, W.; Krautzer, B.; Moser, D.; Frank, T. Improving insect conservation values of agri-environment schemes through diversified seed mixtures. Biol. Conserv. 2022, 269, 109530. [Google Scholar] [CrossRef]
- Cano, D.; Martínez-Núñez, C.; Pérez, A.J.; Salido, T.; Rey, P.J. Small floral patches are resistant reservoirs of wild floral visitor insects and the pollination service in agricultural landscapes. Biol. Conserv. 2022, 276, 109789. [Google Scholar] [CrossRef]
- Winfree, R. The conservation and restoration of wild bees. Ann. N. Y. Acad. Sci. 2010, 1195, 169–197. [Google Scholar] [CrossRef]
- Scheper, J.; Bommarco, R.; Holzschuh, A.; Potts, S.G.; Riedinger, V.; Roberts, S.P.M.; Rundlöf, M.; Smith, H.G.; Steffan-Dewenter, I.; Wickens, J.B.; et al. Local and landscape-level floral resources explain effects of wildflower strips on wild bees across four European countries. J. Appl. Ecol. 2015, 52, 1165–1175. [Google Scholar] [CrossRef]
- Beyer, N.; Gabriel, D.; Kirsch, F.; Schulz-Kesting, K.; Dauber, J.; Westphal, C. Functional groups of wild bees respond differently to faba bean Vicia faba L. cultivation at landscape scale. J. Appl. Ecol. 2020, 57, 2499–2508. [Google Scholar] [CrossRef]
- Pywell, R.F.; Warman, E.A.; Carvell, C.; Sparks, T.H.; Dicks, L.V.; Bennett, D.; Wright, A.; Critchley, C.N.R.; Sherwood, A. Providing foraging resources for bumblebees in intensively farmed landscapes. Biol. Conserv. 2005, 121, 479–494. [Google Scholar] [CrossRef]
- Ambrosino, M.D.; Luna, J.M.; Jepson, P.C.; Wratten, S.D. Relative frequencies of visits to selected insectary plants by predatory hoverflies (Diptera: Syrphidae), other beneficial insects, and herbivores. Environ. Entomol. 2006, 35, 394–400. [Google Scholar] [CrossRef]
- Carvell, C.; Meek, W.R.; Pywell, R.F.; Goulson, D.; Nowakowski, M. Comparing the efficacy of agri-environment schemes to enhance bumble bee abundance and diversity on arable field margins. J. Appl. Ecol. 2007, 44, 29–40. [Google Scholar] [CrossRef]
- Carvalheiro, L.G.; Seymour, C.L.; Nicolson, S.W.; Veldtman, R. Creating patches of native flowers facilitates crop pollination in large agricultural fields: Mango as a case study. J. Appl. Ecol. 2012, 49, 1373–1383. [Google Scholar] [CrossRef]
- Campbell, A.; Sutton, P.; Wilby, A.; Wackers, F. Improving pest control and pollination services in cider apple orchards by means of ’multi-functional’ flowering strips. Environ. Manag. Farml. 2013, 118, 283–290. [Google Scholar]
- Blaauw, B.R.; Isaacs, R. Flower plantings increase wild bee abundance and the pollination services provided to a pollination-dependent crop. J. Appl. Ecol. 2014, 51, 890–898. [Google Scholar] [CrossRef]
- Karamaouna, F.; Kati, V.; Volakakis, N.; Varikou, K.; Garantonakis, N.; Economou, L.; Birouraki, A.; Markellou, E.; Liberopoulou, S.; Edwards, M. Ground cover management with mixtures of flowering plants to enhance insect pollinators and natural enemies of pests in olive groves. Agric. Ecosyst. Environ. 2019, 274, 76–89. [Google Scholar] [CrossRef]
- Kati, V.; Karamaouna, F.; Economou, L.; Mylona, P.V.; Samara, M.; Mitroiu, M.-D.; Barda, M.; Edwards, M.; Liberopoulou, S. Sown Wildflowers Enhance Habitats of Pollinators and Beneficial Arthropod in a Tomato Field Margin. Plants 2021, 10, 1003. [Google Scholar] [CrossRef]
- Karamaouna, F.; Kati, V.; Economou, L.; Troyanos, G.; Samara, M.; Liberopoulou, S.; Barda, M.; Mitroiu, M.-D.; Edwards, M. Selected flowering plants as a habitat for pollinators and natural enemies in field margins of a watermelon crop—Implications for crop yield. Int. J. Pest Manag. 2022. [Google Scholar] [CrossRef]
- Delaplane, K.S.; Mayer, D.F. Crop Pollination by Bees; CABI Publishing: Wallingford, UK, 2000. [Google Scholar]
- Garratt, M.P.; Breeze, T.D.; Jenner, N.; Polce, C.; Biesmeijer, J.C.; Potts, S.G. Avoiding a bad apple: Insect pollination enhances fruit quality and economic value. Agric. Ecosyst. Environ. 2014, 184, 34–40. [Google Scholar] [CrossRef]
- Free, J.B. Pollinating efficiency of honey-bee visits to apple flowers. J. Hortic. Sci. Biotechnol. 1966, 41, 91–94. [Google Scholar] [CrossRef]
- Samnegård, U.; Hambäck, P.A.; Smith, H.G. Pollination treatment affects fruit set and modifies marketable and storable fruit quality of commercial apples. R. Soc. Open Sci. 2019, 6, 190326. [Google Scholar] [CrossRef]
- Vicens, N.; Bosch, J. Pollinating efficacy of Osmia cornuta and Apis mellifera (Hymenoptera: Megachilidae, Apidae) on ’red Delicious’ apple. Environ. Entomol. 2000, 29, 235–240. [Google Scholar] [CrossRef]
- Radzevičiūtė, R.; Theodorou, P.; Schlegel, M.; Paxton, R.J. A two-part modelling approach reveals a positive effect of pollinator biodiversity in boosting the pollination of apple flowers. Agric. Ecosyst. Environ. 2021, 306, 107197. [Google Scholar] [CrossRef]
- Sheffield, C.; Westby, S.; Smith, R.; Kevan, P. Potential of bigleaf lupine for building and sustaining Osmia lignaria populations for pollination of apple. Can. Entomol. 2008, 140, 589–599. [Google Scholar] [CrossRef]
- Sedivy, C.; Dorn, S. Towards a sustainable management of bees of the subgenus Osmia (Megachilidae; Osmia) as fruit tree pollinators. Apidologie 2013, 45, 88–105. [Google Scholar] [CrossRef]
- Viana, B.F.; da Encarnação Coutinho, J.G.; Garibaldi, L.A.; Castagnino, G.L.B.; Gramacho, K.P.; Silva, F.O. Stingless bees further improve apple pollination and production. J. Pollinat. Ecol. 2014, 14, 261–269. [Google Scholar] [CrossRef]
- Sapir, G.; Baras, Z.; Azmon, G.; Goldway, M.; Shafir, S.; Allouche, A.; Stern, E.; Stern, R.A. Synergistic effects between bumblebees and honey bees in apple orchards increase cross pollination, seed number and fruit size. Sci. Hortic. 2017, 219, 107–117. [Google Scholar] [CrossRef]
- Tepedino, V.J.; Alston, D.G.; Bradley, B.A.; Toler, T.R.; Griswold, T.L. Orchard pollination in Capitol Reef National Park, Utah, USA. Honey bees or native bees? Biodivers. Conserv. 2007, 16, 3083–3094. [Google Scholar] [CrossRef]
- Marini, L.; Quaranta, M.; Fontana, P.; Biesmeijer, J.C.; Bommarco, R. Landscape context and elevation affect pollinator communities in intensive apple orchards. Basic Appl. Ecol. 2012, 13, 681–689. [Google Scholar] [CrossRef]
- Campbell, A.J.; Wilby, A.; Sutton, P.; Wäckers, F.L. Do sown flower strips boost wild pollinator abundance and pollination services in a spring-flowering crop? A case study from UK cider apple orchards. Agric. Ecosyst. Environ. 2017, 239, 20–29. [Google Scholar] [CrossRef]
- Miñarro, M.; García, D. Complementarity and redundancy in the functional niche of cider apple pollinators. Apidologie 2018, 49, 789–802. [Google Scholar] [CrossRef]
- Porcel, M.; Andersson, G.K.S.; Pålsson, J.; Tasin, M. Organic management in apple orchards: Higher impacts on biological control than on pollination. J. Appl. Ecol. 2018, 55, 2779–2789. [Google Scholar] [CrossRef]
- Pardo, A.; Borges, P.A.V. Worldwide importance of insect pollination in apple orchards: A review. Agric. Ecosyst. Environ. 2020, 293, 106839. [Google Scholar] [CrossRef]
- FAO. World Food and Agriculture—Statistical Yearbook 2021; FAO: Rome, Italy, 2021. [CrossRef]
- Hellenic Statistical Authority. (24 June 2021) Areas and Production, Annual Agricultural Statistical Survey: Year 2019. Available online: https://www.statistics.gr/documents/20181/96929c5c-54d8-ccca-22fe-1c7af02f8bf3 (accessed on 18 July 2022).
- European Union, Commission Regulation (EC) No. 1065/97 of 12 June 1997 Supplementing the Annex to Regulation (EC) No. 1107/96 on the Registration of Geographical Indications and Designations of Origin under the Procedure Laid Down in Article 17 of Council Regulation (EEC) No. 2081/92. Available online: https://op.europa.eu/en/publication-detail/-/publication/01708409-532d-4e55-9f88-cb277630453f/language-en (accessed on 21 December 2022).
- Kizos, T.; Vakoufaris, H. Alternative agri-food geographies? Geographic indications in greece. Tijdschr. Voor Econ. En Soc. Geogr. 2011, 102, 220–235. [Google Scholar] [CrossRef]
- Mitropoulos, D.; Lambrinos, G. Changes in firmness of apples affected by moisture loss during storage. J. Hortic. Sci. Biotechnol. 2005, 80, 399–402. [Google Scholar] [CrossRef]
- Mitropoulos, D.; Lambrinos, G. “Delicious Pilafa” apple density changes as a quality index of mass loss degradation during storage. J. Food Qual. 2007, 30, 527–537. [Google Scholar] [CrossRef]
- Thanopoulos, R.; Chatzigeorgiou, T.; Argyropoulou, K.; Kostouros, N.-M.; Bebeli, P.J. State of Crop Landraces in Arcadia (Greece) and In-Situ Conservation Potential. Diversity 2021, 13, 558. [Google Scholar] [CrossRef]
- Villa, T.; Maxted, N.; Scholten, M.; Ford-Lloyd, B. Defining and identifying crop landraces. Plant Genet. Resour. 2005, 3, 373–384. [Google Scholar] [CrossRef]
- Veteläinen, M.; Negri, V.; Maxted, N. (Eds.) European Landraces: On-Farm Conservation, Management and Use; Bioversity Technical Bulletin; Bioversity International: Rome, Italy, 2009; Volume 15, p. 358. ISBN 978-92-9043-805-2. Available online: https://hdl.handle.net/10568/106154 (accessed on 27 January 2023).
- Park, M.; Danforth, B.; Losey, J.; Biddinger, D.; Vaughan, M.; Dollar, J.; Edwin Rajotte, E.; Agnello, A. Wild Pollinators of Eastern Apple Orchards and How to Conserve Them; College of Agriculture and Life Sciences, Cornell University: Ithaca, NY, USA, 2012; Available online: http://www.northeastipm.org/park2012 (accessed on 30 October 2020).
- Bellon, M.R.; Gotor, E.; Caracciolo, F. Conserving landraces and improving livelihoods: How to assess the success of on-farm conservation projects? Int. J. Agric. Sustain. 2014, 13, 167–182. [Google Scholar] [CrossRef]
- Goulson, D.; Lye, G.C.; Darvill, B. Decline and conservation of bumble bees. Annu. Rev. Entomol. 2008, 53, 191–208. [Google Scholar] [CrossRef]
- Hanley, M.E.; Franco, M.; Pichon, S.; Darvill, B.; Goulson, D. Breeding system, pollinator choice and variation in pollen quality in British herbaceous plants. Funct. Ecol. 2008, 22, 592–598. [Google Scholar] [CrossRef]
- Kleijn, D.; Raemakers, I. Ecological society of America A retrospective analysis of pollen host plant use by stable and declining bumble bee species. Ecol. Soc. Am. 2008, 89, 1811–1823. [Google Scholar] [CrossRef]
- Scheper, J.; Reemer, M.; van Kats, R.; Ozinga, W.A.; van der Linden, G.T.J.; Schaminée, J.H.J.; Siepel, H.; Kleijn, D. Museum specimens reveal loss of pollen host plants as key factor driving wild bee decline in The Netherlands. Proc. Natl. Acad. Sci. USA 2014, 111, 17552–17557. [Google Scholar] [CrossRef]
- Cole, L.J.; Baddeley, J.A.; Robertson, D.; Topp, C.F.E.; Walker, R.L.; Watson, C.A. Supporting wild pollinators in agricultural landscapes through targeted legume mixtures. Agric. Ecosyst. Environ. 2022, 323, 107648. [Google Scholar] [CrossRef]
- Pfiffner, L.; Laurent, J.; Cahenzli, F.; Maren, K.; Weronika, S.; Lene, S. Perennial Flower Strips—A Tool for Improving Pest Control in Fruit Orchards, Technical Guide No. 1096. 2018; 16p. Available online: www.fibl-shop.org (accessed on 16 March 2019).
- Cresswell, C.J.; Cunningham, H.M.; Wilcox, A.; Randall, N.P. A trait-based approach to plant species selection to increase functionality of farmland vegetative strips. Ecol. Evol. 2019, 9, 4532–4543. [Google Scholar] [CrossRef]
- Shavit, O.; Dafni, A.; Neeman, G. Competition between honey bees (Apis mellifera) and native solitary bees in the Mediterranean region of Israel—Implications for conservation. Isr. J. Plant Sci. 2009, 57, 171–183. [Google Scholar] [CrossRef]
- Kennedy, C.M.; Lonsdorf, E.; Neel, M.C.; Williams, N.M.; Ricketts, T.H.; Winfree, R.; Bommarco, R.; Brittain, C.; Burley, A.L.; Cariveau, D.; et al. A global quantitative synthesis of local and landscape effects on wild bee pollinators in agroecosystems. Ecol. Lett. 2013, 16, 584–599. [Google Scholar] [CrossRef]
- Gleiciani, P.-R.; Campos, M. Aspects of Landscape and Pollinators—What is Important to Bee Conservation? Diversity 2014, 6, 158–175. [Google Scholar] [CrossRef]
- Raderschall, C.A.; Bommarco, R.; Lindström, S.A.M.; Lundin, O. Landscape crop diversity and semi-natural habitat affect crop pollinators, pollination benefit and yield. Agric. Ecosyst. Environ. 2021, 306, 107189. [Google Scholar] [CrossRef]
- Tutin, T.G.E.A. (Ed.) Flora Europaea v. 5; Cambridge University Press: Cambridge, UK, 1980. [Google Scholar]
- Collins, A.G. Key to the Genera of British Bees; BWARS: London, UK, 2012; Available online: https://www.bwars.com/content/bees-britain (accessed on 5 September 2019).
- SAS Institute Inc. JMP v7; SAS Institute Inc.: Cary, NC, USA. Available online: https://www.jmp.com/en_ph/company/about-us.html (accessed on 27 January 2023).
- Feltham, H.; Park, K.; Minderman, J.; Goulson, D. Experimental evidence that wildflower strips increase pollinator visits to crops. Ecol. Evol. 2015, 5, 3523–3530. [Google Scholar] [CrossRef]
- Königslöw, V.; Fornoff, F.; Klein, A.M. Wild bee communities benefit from temporal complementarity of hedges and flower strips in apple orchards. J. Appl. Ecol. 2022, 59, 2814–2824. [Google Scholar] [CrossRef]
- Free, J.B. Insect Pollination of Crops By J. B. Free London: Academic Press (1970), pp. 544, £7.25. Exp. Agric. 1971, 7, 367–368. [Google Scholar] [CrossRef]
- Kirk, W.D.J. Faba bean: Vicia faba. Bee World 2004, 85, 60–62. [Google Scholar] [CrossRef]
- Aouar-Sadli, M.; Louadi, K.; Doumandji, S. Pollination of the broad bean (Vicia faba L. var. major) (Fabaceae) by wild bees and honey bees (Hymenoptera: Apoidea) and its impact on the seed production in the Tizi-Ouzou area (Algeria). Afr. J. Agric. Res. 2018, 3, 266–272. [Google Scholar]
- Marzinzig, B.; Brünjes, L.; Biagioni, S.; Behling, H.; Link, W.; Westphal, C. Bee pollinators of faba bean (Vicia faba L.) differ in their foraging behaviour and pollination efficiency. Agric. Ecosyst. Environ. 2018, 264, 24–33. [Google Scholar] [CrossRef]
- Boyle, R.M.D.; Philogène, B.J.R. The native pollinators of an apple orchard: Variations and significance. J. Hortic. Sci. 1983, 58, 355–363. [Google Scholar] [CrossRef]
- Wei, S.G.; Wang, R.; Smirle, M.J.; Xu, H.L. Release of Osmia excavata and Osmia jacoti (Hymenoptera: Megachilidae) for apple pollination. Can. Entomol. 2002, 134, 369–380. [Google Scholar] [CrossRef]
- Adamson, N.L.; Roulston, T.H.; Fell, R.D.; Mullins, D.E. From April to August—Wild bees pollinating crops through the growing season in Virginia, USA. Environ. Entomol. 2012, 41, 813–821. [Google Scholar] [CrossRef]
- Campbell, A.; Wilby, A.; Sutton, P.; Wäckers, F. Getting More Power from Your Flowers: Multi-Functional Flower Strips Enhance Pollinators and Pest Control Agents in Apple Orchards. Insects 2017, 8, 101. [Google Scholar] [CrossRef]
- Carvell, C.; Osborne, J.L.; Bourke, A.F.G.; Freeman, S.N.; Pywell, R.F.; Heard, M.S. Bumble bee species’ responses to a targeted conservation measure depend on landscape context and habitat quality. Ecol. Appl. 2011, 21, 1760–1771. [Google Scholar] [CrossRef]
- Underwood, E.; Tucker, G. Ecological Focus Area choices and their potential impacts on biodiversity. In Report for BirdLife Europe and the European Environmental Bureau; Institute for European Environmental Policy: London, UK, 2016. [Google Scholar] [CrossRef]
- Hart, K.; Mottershead, D.; Tucker, G.; Underwood, E.; Mar´echal, A.; Menet, L.; Martin, I.; Dayde, C.; Bresson, C.; Deniel, E.; et al. European Commission, Directorate-General for Agriculture and Rural Development, Evaluation Study of the Payment for Agricultural Practices Beneficial for the Climate and the Environment; Final Report. Publications Office: Luxembourg, 2017. Available online: https://data.europa.eu/doi/10.2762/71725 (accessed on 28 December 2022).
- Cole, L.J.; Kleijn, D.; Dicks, L.V.; Stout, J.C.; Potts, S.G.; Albrecht, M.; Balzan, M.V.; Bartomeus, I.; Bebeli, P.J.; Bevk, D.; et al. A critical analysis of the potential for EU Common Agricultural Policy measures to support wild pollinators on farmland. J. Appl. Ecol. 2020, 57, 681–694. [Google Scholar] [CrossRef]
- Shakeel, M.; Ali, H.; Ahmad, S.; Said, F.; Khan, K.A.; Bashir, M.A.; Anjum, S.I.; Islam, W.; Ghramh, H.A.; Ansari, M.J.; et al. Insect pollinators diversity and abundance in Eruca sativa Mill. (Arugula) and Brassica rapa L. (Field mustard) crops. Saudi J. Biol. Sci. 2018, 26, 1704–1709. [Google Scholar] [CrossRef]
- El-Berry, A.A.; Moustafa, M.A.; Abdel-Gawaad, A.A.; El-Bialey, S. Pollinators other than honey bees visiting certain vegetable plants in Egypt. Z. Für Angew. Entomol. 1974, 77, 106–110. [Google Scholar] [CrossRef]
- Dunn, L.; Lequerica, M.; Reid, C.R.; Latty, T. Dual ecosystem services of syrphid flies (Diptera: Syrphidae): Pollinators and biological control agents. Pest Manag. Sci. 2020, 76, 1973–1979. [Google Scholar] [CrossRef]
- Boroumand, R.Z.; Kouchaki, A. Evaluation of cardinal temperature for three species of medicinal plants, Ajowan (Trachyspermum ammi), Fennel (Foeniculum vulgare) and Dill (Anethum graveolens). Desert J. 2006, 11, 11–16. [Google Scholar]
- Hussein, M.H.; Abdel-Aal, S.A. Wild and honeybees as pollinators of 10 plant species in Assiut area. Egypt. Z. Fur Angew. Entomol. 1982, 93, 342–346. [Google Scholar] [CrossRef]
- Bendifallah, L.; Louadi, K.; Doumandji, S. Bee fauna potential visitors of coriander flowers Coriandrum sativum L. (Apiaceae) in the Mitidja area (Algeria). J. Apic. Sci. 2013, 57, 59–70. [Google Scholar] [CrossRef]
- Bretagnolle, V.; Gaba, S. Weeds for bees? A review. Agron. Sustain. Dev. 2015, 35, 891–909. [Google Scholar] [CrossRef]
- Štefanić, E.; Rašić, S.; Panjković, B.; Kovačević, V.; Zima, D.; Antunović, S.; Štefanić, I. The role of weeds from field margins in supporting crop pollinators. J. Cent. Eur. Agric. 2020, 21, 602–608. [Google Scholar] [CrossRef]
- Carvell, C.; Meek, W.R.; Pywell, R.F.; Nowakowski, M. The response of foraging bumblebees to successional change in newly created arable field margins. Biol. Conserv. 2004, 118, 327–339. [Google Scholar] [CrossRef]
- Nichols, R.N.; Goulson, D.; Holland, J.M. The best wildflowers for wild bees. J. Insect Conserv. 2019, 23, 819–830. [Google Scholar] [CrossRef]
- Ara, S.; Rather, Z.A.; Paray, M.A. Hang around flora-the pollination enhancers of apple, of Kashmir Himalaya. J. Pharmacogn. Phytochem. 2018, 7, 1462–1467. [Google Scholar]
- Lysenkov, S.N. Difficulties in Studies of the Ecological Links between Plants and Flower-Visiting Insects: One and a Half Centuries of Research on Veronica chamaedrys L. Biol. Bull. Rev. 2021, 11, 488–497. [Google Scholar] [CrossRef]
- Pesenko, Y.A. Contributions to the halictid fauna of the Eastern Palaearctic Region: Genus Lasioglossum Curtis (Hymenoptera: Halictidae, Halictinae). Zoosystematica Ross. 2006, 15, 133–166. [Google Scholar] [CrossRef]
- Nutt, P.; Ziermann, J.; Hintz, M.; Neuffer, B.; Theißen, G. Capsella as a model system to study the evolutionary relevance of floral homeotic mutants. Plant Syst. Evol. 2006, 259, 217–235. [Google Scholar] [CrossRef]
- Rosa García, R.; Miñarro, M. Role of floral resources in the conservation of pollinator communities in cider-apple orchards. Agric. Ecosyst. Environ. 2014, 183, 118–126. [Google Scholar] [CrossRef]
- Lynn, A.; Piotter, E.; Harrison, E.; Galen, C. Sexual and natural selection on pollen morphology in Taraxacum. Am. J. Bot. 2020, 107, 364–374. [Google Scholar] [CrossRef]
- Hattori, M.; Nagano, Y.; Itino, T. Geographic Variation in Flower Size and Flower-Visitor Composition of Two Bumblebee-Pollinated, Spring-Flowering Herbs, Lamium album L. var. barbatum (Lamiaceae) and Meehania urticifolia (Lamiaceae). Am. J. Plant Sci. 2015, 6, 737–745. [Google Scholar] [CrossRef]
- Waddington, K.D. Foraging patterns of halictid bees at flowers of Convolvulus arvensis. Psyche 1976, 83, 112–119. [Google Scholar] [CrossRef]
- Holm, L.G.; Plucknett, D.L.; Pancho, J.V.; Herberger, J.P. The World’s Worst Weeds. Distribution and Biology; University Press of Hawaii: Honolulu, HI, USA, 1977. [Google Scholar]
- Skinner, K.; Smith, L.; Rice, P. Using noxious weed lists to prioritize targets for developing weed management strategies. Weed Sci. 2000, 48, 640–644. [Google Scholar] [CrossRef]
- Albrecht, M.; Kleijn, D.; Williams, N.M.; Tschumi, M.; Blaauw, B.R.; Bommarco, R.; Campbell, A.J.; Dainese, M.; Drummond, F.A.; Entling, M.H.; et al. The effectiveness of flower strips and hedgerows on pest control, pollination services and crop yield: A quantitative synthesis. Ecol. Lett. 2020, 23, 1488–1498. [Google Scholar] [CrossRef]
- McKerchar, M.; Potts, S.G.; Fountain, M.T.; Garratt, M.P.D.; Westbury, D.B. The potential for wildflower interventions to enhance natural enemies and pollinators in commercial apple orchards is limited by other management practices. Agric. Ecosyst. Environ. 2020, 301, 107034. [Google Scholar] [CrossRef]
- Wood, T.J.; Holland, J.M.; Goulson, D. Pollinator-friendly management does not increase the diversity of farmland bees and wasps. Biol. Conserv. 2015, 187, 120–126. [Google Scholar] [CrossRef]
- Morse, R.A.; Calderone, N.W. The Value of Honey Bees as Pollinators of US Crops in 2000. Bee Cult. 2000, 128, 1–15. [Google Scholar]
- Prendergast, K.S.; Leclercq, N.; Vereecken, N.J. Honey bees (Hymenoptera: Apidae) outnumber native bees in Tasmanian apple orchards: Perspectives for balancing crop production and native bee conservation. Austral Entomol. 2021, 60, 422–435. [Google Scholar] [CrossRef]
- Geslin, B.; Aizen, M.A.; Garcia, N.; Pereira, A.-J.; Vaissière, B.E.; Garibaldi, L.A. The impact of honey bee colony quality on crop yield and farmers’ profit in apples and pears. Agric. Ecosyst. Environ. 2017, 248, 153–161. [Google Scholar] [CrossRef]
- Blitzer, E.J.; Gibbs, J.; Park, M.G.; Danforth, B.N. Pollination services for apple are dependent on diverse wild bee communities. Agric. Ecosyst. Environ. 2016, 221, 1–7. [Google Scholar] [CrossRef]
- Földesi, R.; Kovács-Hostyánszki, A.; Kőrösi, Á.; Somay, L.; Elek, Z.; Markó, V.; Sárospataki, M.; Bakos, R.; Varga, Á.; Nyisztor, K.; et al. Relationships between wild bees, hoverflies and pollination success in apple orchards with different landscape contexts. Agric. For. Entomol. 2016, 18, 68–75. [Google Scholar] [CrossRef]
- Lowe, E.B.; Groves, R.; Gratton, C. Impacts of field-edge flower plantings on pollinator conservation and ecosystem service delivery—A meta-analysis. Agric. Ecosyst. Environ. 2021, 310, 107290. [Google Scholar] [CrossRef]
- Watson, J.C.; Wolf, A.T.; Ascher, J.S. Forested landscapes promote richness and abundance of native bees (Hymenoptera: Apoidea: Anthophila) in Wisconsin apple orchards. Environ. Entomol. 2011, 40, 621–632. [Google Scholar] [CrossRef]
- Gervais, A.; Bélisle, M.; Mazerolle, M.J.; Fournier, V. Landscape Enhancements in Apple Orchards: Higher Bumble Bee Queen Species Richness, But No Effect on Apple Quality. Insects 2021, 12, 421. [Google Scholar] [CrossRef]
- Senapathi, D.; Biesmeijer, J.C.; Breeze, T.D.; Kleijn, D.; Potts, S.G.; Carvalheiro, L.G. Pollinator conservation—The difference between managing for pollination services and preserving pollinator diversity. Curr. Opin. Insect Sci. 2015, 12, 93–101. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Plant Species | Seed Percentage (%) | |
|---|---|---|---|
| Year | |||
| 2020 | 2021 | ||
| Apiaceae | Anethum graveolens L. | 15 | 10 |
| Apiaceae | Coriandrum sativum L. | 15 | 20 |
| Apiaceae | Foeniculum vulgare Mill. | - | 10 |
| Brassicaceae | Eruca sativa Mill. | 15 | 25 |
| Fabaceae | Lathyrus sativus L. (PGI ‘Fava Feneou’) | 15 | - |
| Fabaceae | Vicia faba L. (landrace ‘Episkopis’) | 15 | 7 |
| Fabaceae | Vicia sativa L. | 15 | 23 |
| Poaceae | Triticum aestivum L. (landrace ‘Zoolitsa’) | 15 | - |
| Poaceae | Triticum aestivum L. (landrace ‘Asprositi’) | - | 5 |
| Family | Genus | Species | Associated Plants |
|---|---|---|---|
| Andrenidae | Andrena | Andrena sp1 (♀) Andrena sp2 (♀) Andrena sp3 (♀) Andrena sp4 (♀) Andrena sp5 (♀) Andrena sp6 (♀,♂) Andrena sp7 (♀) Andrena sp8 (♀) Andrena sp9 (♀) Andrena sp10 (♀) Andrena sp11 (♂) Andrena sp12 (♀,♂) | Taraxacum sp., Daucus carota Gallium aparine Veronica sp. Stellaria apetala Malus domestica FM, Medicago arabica Eruca. sativa Trifolium repens Coriandrum sativum Coriandrum sativum Veronica sp. FM, Coriandrum sativum, Cardaria draba |
| Apidae | Apis | A. mellifera (♀) | M. domestica, Vicia faba, Coriandrum sativum, Veronica spp., E. sativa, Ranunculus repens, Calepina irregularis, Lamium amplexicaule, Cardaria draba, Triffolium repens, Convolvulus arvense, Raphanus sp. |
| Anthophora | A. plumipes (♀) Anthophora spp. (♀) | Vicia faba, Eruca sativa Eruca sativa, Lamium amplexicaule | |
| Bombus | B. argillaceus (♀) B. terrestris (♀,♂) Bombus sp1 (♀) Bombus sp2 (♀) | Vicia faba Vicia faba, Malus domestica Malus domestica Vicia faba | |
| Eucera | E. nigrescens (♀,♂) Eucera sp1 (♂) Eucera sp2 (♂) | FM, Torilis sp., V. villosa, V. faba, E. sativa Medicago arabica Eruca sativa | |
| Nomada | Nomada sp. (♀) | NV | |
| Xylocopa | X. violaceae (♀,♂) | Vicia faba, Vicia sativa | |
| Colletidae | Hylaeus | H. variegatus (♀) Hylaeus sp1 (♀,♂) Hylaeus sp2 (♀) | Coriandrum sativum Coriandrum sativum Eruca sativa |
| Halictidae | Halictus | Halictus sp1 (♀) Halictus sp2 (♀) Halictus sp3 (♀) | Crepis sp. Coriandrum sativum Coriandrum sativum |
| Lasioglossum | L. marginatum (♀) Lasioglossum sp1 (♀) Lasioglossum sp2 (♀) Lasioglossum sp3 (♀) Lasioglossum sp4 (♀) Lasioglossum sp5 (♀) Lasioglossum sp6 (♀) Lasioglossum sp7 (♀) Lasioglossum sp8 (♀) Lasioglossum sp9 (♀) Lasioglossum sp10 (♀) Lasioglossum sp11 (♀) Lasioglossum sp12 (♀) Lasioglossum sp13 (♀) | Malus domestica Veronica sp., Sonchus oleraceus Eruca sativa, Veronica sp. Lathyrus sativus Eruca sativa Malus domestica Veronica sp. Eruca sativa Capsella bursa-pastoris Medicago arabica Ranunculus repens Coriandrum sativum Convolvulus arvense NV | |
| Systropha | S. curvicornis (♀,♂) | Convolvulus arvense | |
| Sphecodes | Sphecodes sp. (♀) | Coriandrum sativum |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barda, M.; Karamaouna, F.; Kati, V.; Perdikis, D. Do Patches of Flowering Plants Enhance Insect Pollinators in Apple Orchards? Insects 2023, 14, 208. https://doi.org/10.3390/insects14020208
Barda M, Karamaouna F, Kati V, Perdikis D. Do Patches of Flowering Plants Enhance Insect Pollinators in Apple Orchards? Insects. 2023; 14(2):208. https://doi.org/10.3390/insects14020208
Chicago/Turabian StyleBarda, Myrto, Filitsa Karamaouna, Vaya Kati, and Dionysios Perdikis. 2023. "Do Patches of Flowering Plants Enhance Insect Pollinators in Apple Orchards?" Insects 14, no. 2: 208. https://doi.org/10.3390/insects14020208
APA StyleBarda, M., Karamaouna, F., Kati, V., & Perdikis, D. (2023). Do Patches of Flowering Plants Enhance Insect Pollinators in Apple Orchards? Insects, 14(2), 208. https://doi.org/10.3390/insects14020208