
Host Specificity and Preliminary Impact of Lepidapion argentatum (Coleoptera, Brentidae), a Biocontrol Candidate for French Broom (Genista monspessulana, Fabaceae)


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
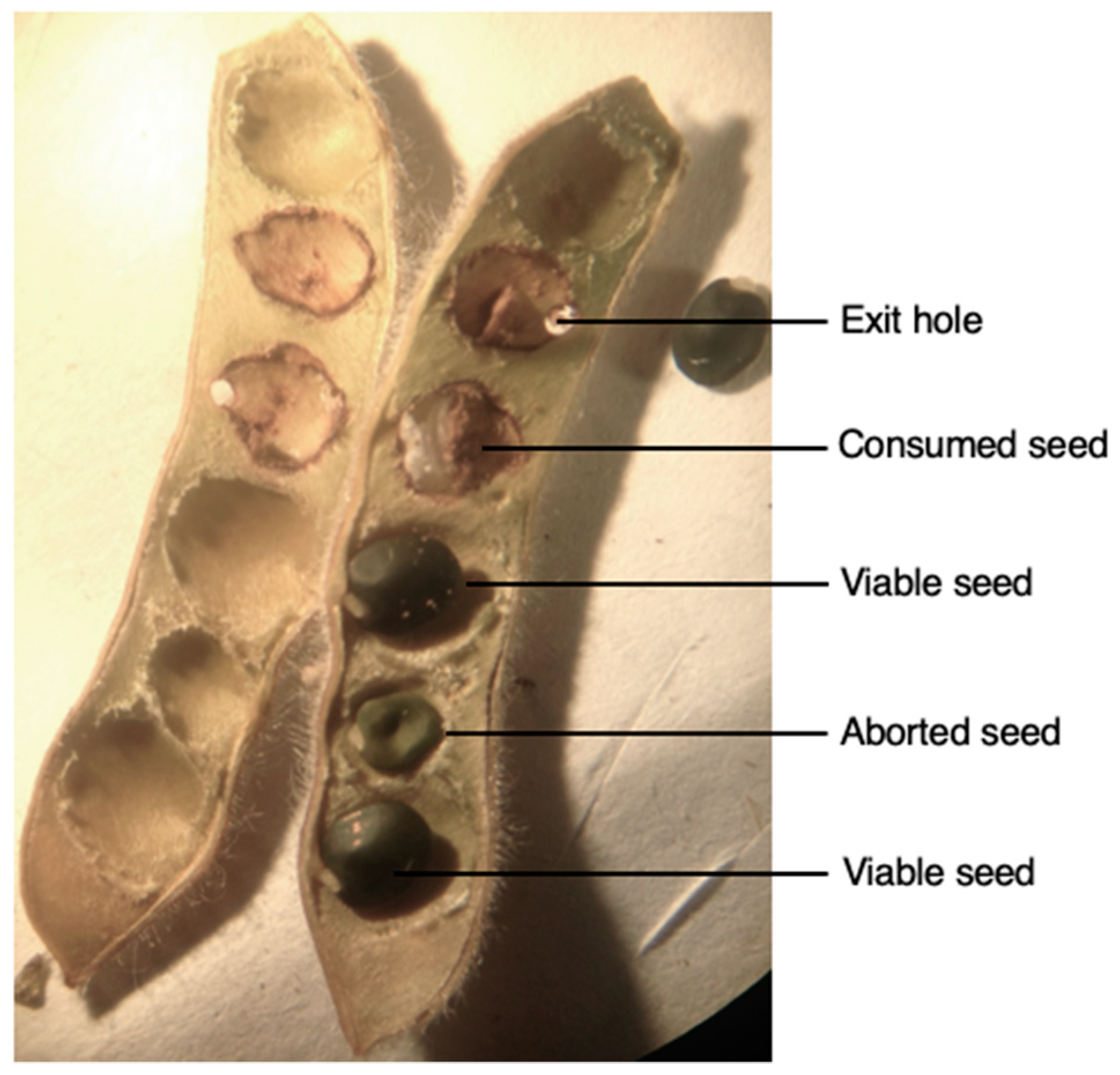
2.1. Evaluation of Weevil Impact on Pods
2.2. Host–Specificity Tests
- No-Choice Tests on Pods in 2012 and 2015
- No-Choice Tests on Stems in 2012 and 2015
3. Results
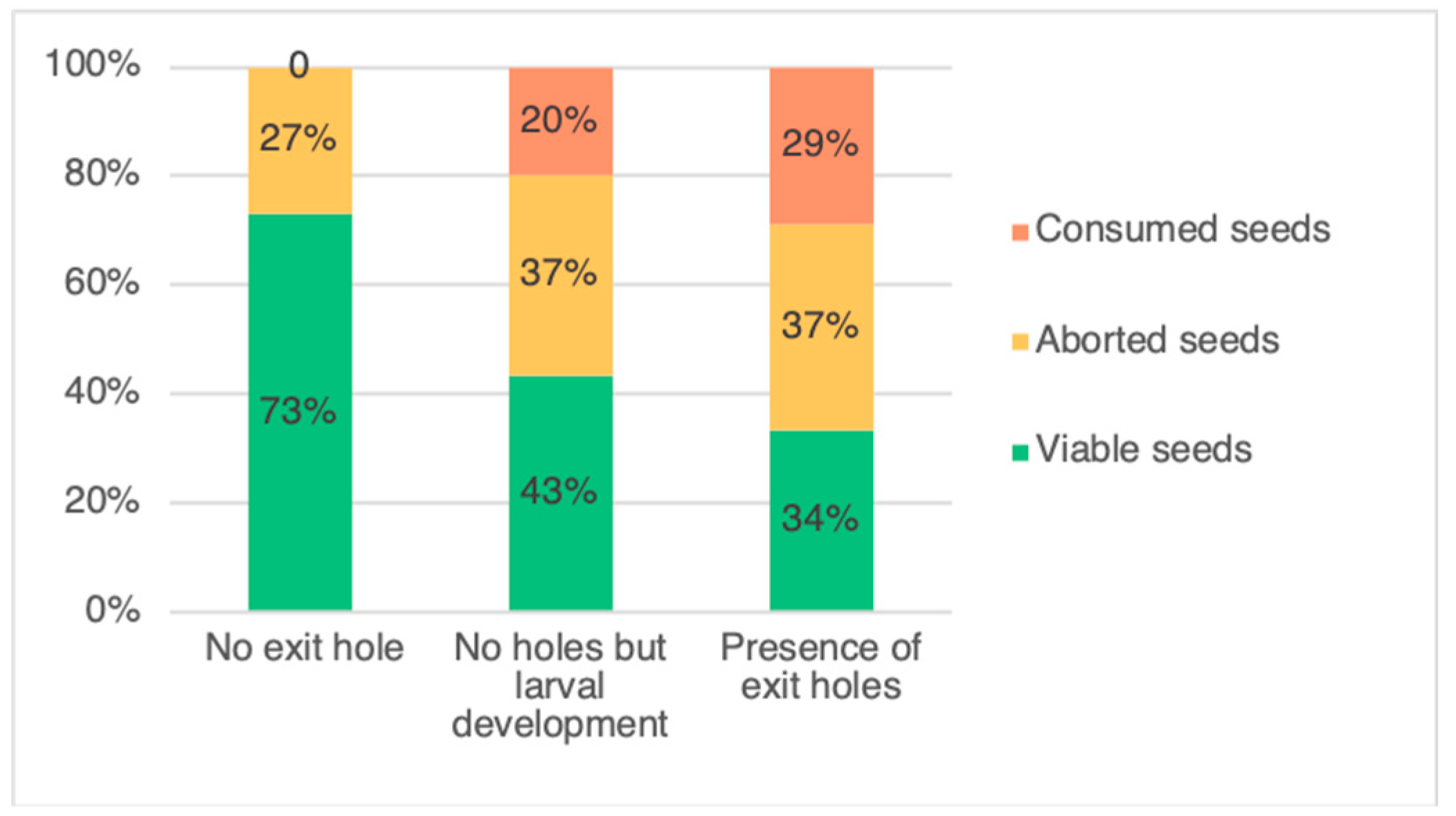
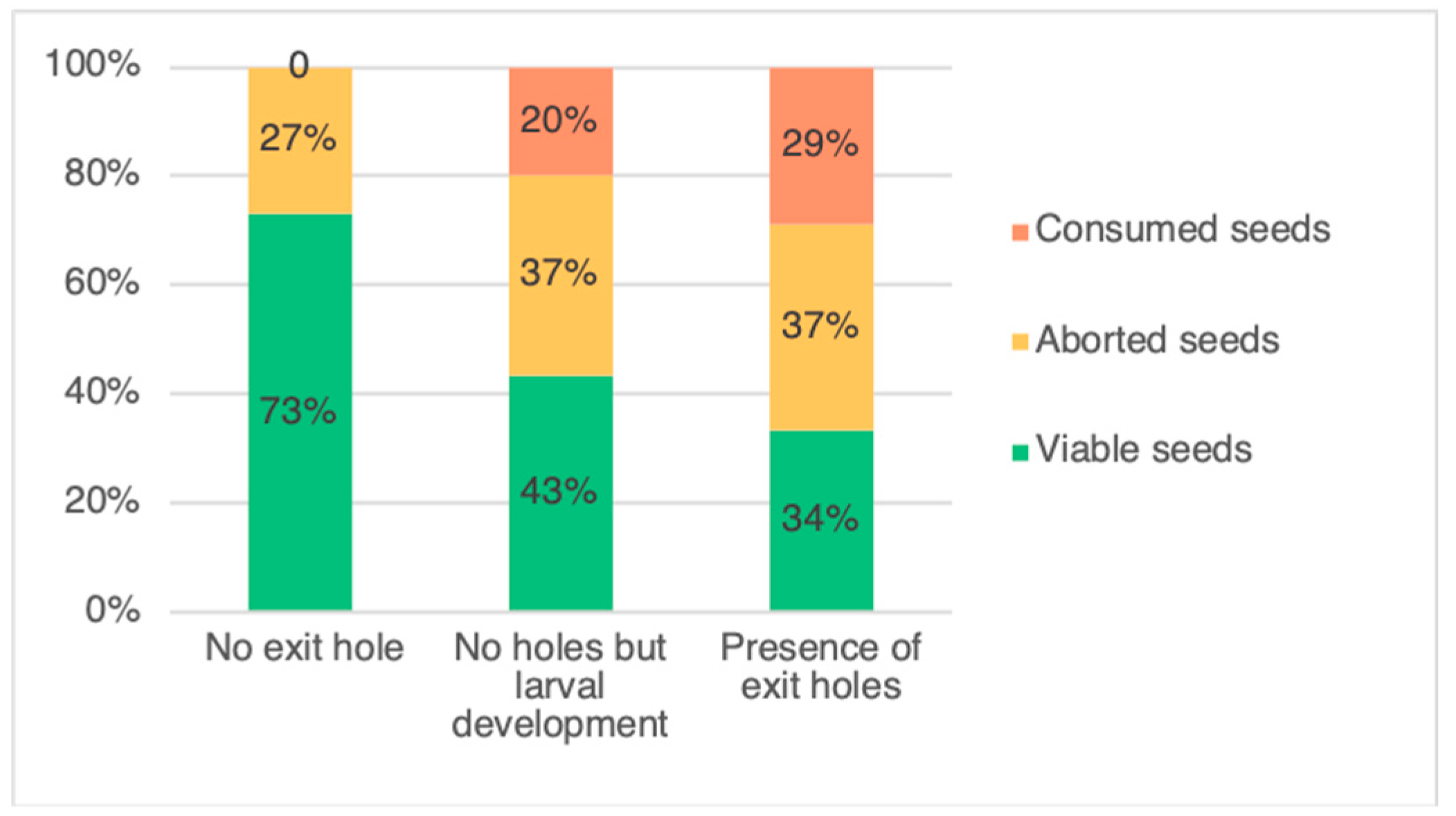
3.1. Evaluation of Weevil Impact on Pods
3.2. Host–Specificity Tests
- No-choice tests on pods in 2012 and 2015
- No-choice tests on stems in 2012 and 2015
- Choice tests
4. Discussion
4.1. Evaluation of Weevil Impact on Pods
4.2. Host–Specificity Tests
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Parsons, W.T.; Cuthbertson, E.G. Noxious Weeds of Australia; CSIRO Publishing: Melbourne, Australia, 2001; pp. 1–689. [Google Scholar]
- Alexander, J.M.; Antonio, C.M.D. Control Methods for the Removal of French and Scotch Broom Tested in Coastal California. Ecol. Restor. 2003, 21, 191–198. [Google Scholar] [CrossRef]
- Bossard, C. Genista monspessulana (L.) L. Johnson. In Invasive Plants of California’s Wildlands; Bossard, C.C., Randall, J.M., Hoshovsky, M.C., Eds.; University of California Press: Berkeley, CA, USA, 2000; pp. 203–208. [Google Scholar]
- Henry, K.; Sheppard, A.; Virtue, J.; Thomann, T.; Ivory, S.; Lush, A. Application for the approval of the Cape broom psyllid, Arytinnis hakani, a potential biological control agent for Cape broom, Genista monspessulana. S. Aust. Res. Dev. Inst. Sustain. Systems. Entomol. 2008, 1–34. [Google Scholar]
- Sullivan, P. A success story: The Cape broom psyllid, Arytinnis hakani Loginova. Plant Prot. Q. 2013, 28, 81–83. [Google Scholar]
- Sheppard, A.W.; Henry, K. Genista monspessulana—Cape broom. In Biological Control of Weeds in Australia 1960 to 2010; Julien, M., McFadyen, R., Cullen, J., Eds.; CSIRO Publishing: Melbourne, Australia, 2012; pp. 267–273. [Google Scholar]
- Kleist, A.; Herrera-Reddy, A.M.; Sforza, R.; Jasieniuk, M. Inferring the complex origins of horticultural invasives: French broom in California. Biol. Invasions 2014, 16, 887–901. [Google Scholar] [CrossRef]
- Sheppard, A. Survey and Selection of Agents for the Biological Control of Montpellier Broom, Genista monspessulana, CSIRO Entomology, Contracted Research Report No. 59 (unpublished) p59; CSIRO Entomology: Canberra, Australia, 2000; pp. 1–59. [Google Scholar]
- Herrera, A.M.; Carruthers, R.I.; Mills, N.J. Introduced populations of Genista monspessulana (French broom) are more dense and produce a greater seed rain in California, USA, than native populations in the Mediterranean Basin of Europe. Biol. Invasions 2011, 13, 369–380. [Google Scholar] [CrossRef]
- Sheppard, A.W.; Shaw, R.H.; Sforza, R. Top 20 environmental weeds for classical biological control in Europe: A review of opportunities, regulations and other barriers to adoption. Weed Res. 2006, 46, 93–117. [Google Scholar] [CrossRef]
- Sheppard, A.; Thomann, T. Quantitative field surveys for the selection of biological control agents for Genista monspessulana, based on host range and efficacy assessment. In Proceedings of the XI International Symposium on Biological Control of Weeds, Canberra, Australia, 28 April–2 May 2003; pp. 162–174. [Google Scholar]
- Cook, B.S.; Smith, L. Prerelease efficacy test of the psyllid, Arytinnis hakani, a prospective biological control agent of the invasive weed Genista monspessulana. Biocontrol Sci. Technol. 2014, 24, 641–651. [Google Scholar] [CrossRef]
- Hogg, B.N.; Moran, P.J.; Smith, L. Multi-generational impacts of the psyllid Arytinnis hakani (Hemiptera: Psyllidae) on growth and survival of the invasive weed Genista monspessulana. Biol. Control 2016, 100, 87–93. [Google Scholar] [CrossRef] [Green Version]
- Sanchez, A. Lutte Biologique Contre L’invasion du Genêt de Montpellier en Australie et Aux Etats-Unis; Rapport BTS Technologies Végétales: Montpellier, France, 2005; pp. 1–30. [Google Scholar]
- Sheppard, A.; Thomann, T. International Broom Initiative: Biological Control of French Broom: European Native-Range Studies; Phase III. Final Report; CSIRO: Canberra, Australia, 2006; pp. 1–56. [Google Scholar]
- Alonso-Zarazaga, M.A. Revision of the supraspecific taxa in the Palearctic Apionidae Shoenherr, 1823 (Coleoptera, Curculionidea). 2. Subfamily Apioninea Shoenherr, 1823: Introduction, keys and descriptions. Graellsia 1990, 46, 76–78. [Google Scholar]
- Tutin, T.G.; Heywood, V.H.; Burges, N.A.; Moore, D.M.; Valentine, D.H.; Walters, S.M.; Webb, D.A. Flora Europaea Vol 2. Rosaceae to Umbelliferae; Cambridge University Press: Cambridge, UK, 1968. [Google Scholar]
- Hoffmann, A. Apion (Lepidapion) argentatum. Faune de France, 3ème partie, Coléoptères et Curculionidés 1958, 3, 1542–1544. [Google Scholar]
- Kerdellant, E.; Thomann, T.; Vitou, J.; Giusto, C.; Simonot, O.; Sheppard, A.; Sforza, R.F.H. The double life cycle of Lepidapion argentatum, a promising candidate for the biological control of French Broom. Biocontrol Sci. Technol. 2019, 1–13. [Google Scholar] [CrossRef]
- Wapshere, A.J. A strategy for evaluating the safety of organisms for biological weed control. Ann. Appl. Biol. 1974, 77, 201–211. [Google Scholar] [CrossRef]
- Akers, R.P.; Pitcairn, M.J.; Carruthers, R. Non-Target Host Plant List for Host Specificity Testing of French Broom (Genista monspessulana (L.) Johnson) (Fabaceae) CDFA Unpublished Proposal for Approval of a Test Plant List to USDA-APHIS TAG; 2005; pp. 1–26. [Google Scholar]
- Sheppard, A.; Thomann, T.; Vitou, J. Biological control of French Broom: Life cycle of Lepidapion argentatum (Gerstäcker) (Curculionidea: Apionidae) and its host specificity testing on Australian Lupinus cultivars. In RIRDC Biological Control of Weeds in South Eastern Australia; Final Csiro Report; CSIRO: Canberra, Australia, 2013; pp. 1–31. [Google Scholar]
- Sheppard, A.; Thomann, T.; Agret, S. Foreign Exploration & Host Specificity Testing of Biological Control Agents of French Broom in California; CSIRO: Canberra, Australia, 2002. [Google Scholar]
- Mc Kay, F.; Gandolfo, D.; Witt, A.B.R. Biology and host range of Coelocephalapion gandolfoi Kissinger ( Brentidae ), a promising candidate for the biological control of invasive Prosopis species ( Leguminosae ) in South Africa. Afr. Entomol. 2012, 20, 281–291. [Google Scholar] [CrossRef]
- Cowley, J.M. Life cycle of Apion ulicis (Coleoptera: Apionidae),and gorse seed attack around Auckland, New Zealand. N. Z. J. Zool. 1983, 10, 83–86. [Google Scholar] [CrossRef]
- Rodriguez, E.; Peco, B.; Gurrea, M.P. Effect of Scotch broom, Cytisus scoparius, pod size and patch density on Exapion fuscirostre (Coleoptera, Apionidae) seed weevil oviposition. Aust. J. Entomol. 2012, 51, 127–132. [Google Scholar] [CrossRef]
- Bitume, E.V.; Moran, P.J.; Sforza, R.F.H. Impact in quarantine of the galling weevil Lepidapion argentatum on shoot growth of French broom (Genista monspessulana), an invasive weed in the western U.S. Biocontrol Sci. Technol. 2019. [Google Scholar] [CrossRef]
- Gonzáles, W.L.; Caballero, P.P.; Medel, R. Galler-induced reduction of shoot growth and fruit production in the shrub Colliguaja integerrima (Euphorbiaceae). Rev. Chil. De Hist. Nat. 2005, 78, 83–93. [Google Scholar] [CrossRef]
- Harris, P.; Shorthouse, J.D. Effectiveness of Gall Inducers in Weed Biological Control. Can. Entomol. 1996, 128, 1021–1055. [Google Scholar] [CrossRef] [Green Version]
- Hartnett, D.C.; Abrahamson, W.G. The effects of stem gall insects on life history patterns in Solidago canadensis. Ecology 1979, 60, 910–917. [Google Scholar] [CrossRef]
- Korotyaev, B.A.; Konstantinov, A.S.; Lingafelter, S.W.; Mandelshtam, M.Y.; Volkovitsh, M.G. Gall-inducing Coleoptera. Biol. Ecol. Evol. Gall-Inducing Arthropods 2005, 1, 239–271. [Google Scholar]
- De Souza, A.L.T.; Fernandes, G.W.; Figueira, J.E.C.; Tanaka, M.O. Natural history of a gall-inducing weevil Collabismus clitellae (Coleoptera: Curculionidae) and some effects on its host plant Solanum lycocarpum (Solanaceae) in southeastern Brazil. Ann. Entomol. Soc. Am. 1998, 91, 404–409. [Google Scholar]
- Baars, J.R.; Hill, M.P.; Heystek, F.; Neser, S.; Urban, A.J. Biology, oviposition preference and impact in quarantine of the petiole-galling weevil, Coelocephalapion camarae Kissinger, a promising candidate agent for biological control of Lantana camara. Biol. Control 2007, 40, 187–195. [Google Scholar] [CrossRef]
- Ulmer, B.J.; Duncan, R.E.; Prena, J.; Pena, J.E. A Weevil, Eurhinus magnificus Gyllenhal (Insecta: Coleoptera: Curculionidae) 1; Department of Entomology and Nematology, University of Florida/IFAS Extension: Gainesville, FL, USA, 2007; pp. 1–6. [Google Scholar]
- Vergara-Pineda, S.; Jones, R.W.; Malda-barrera, G.; Caltzontzin-Fernandez, K.; Obregon-Zuniga, A.; Cambron-Sandoval, V.H. Coelocephalapion subornatum (Fall) (Coleoptera: Brentidae: Apioninae) Forms Galls in Stems of Acacia farnesiana (L.) Willd. in Central Mexico. Southwest. Entomol. 2015, 40, 223–226. [Google Scholar] [CrossRef]
- McClay, A.S.; Balciunas, J.K. The role of pre-release efficacy assessment in selecting classical biological control agents for weeds—Applying the Anna Karenina principle. Biol. Control 2005, 35, 197–207. [Google Scholar] [CrossRef]
- Pearson, D.E.; Callaway, R.M. Indirect nontarget effects of host-specific biological control agents: Implications for biological control. Biol. Control 2005, 35, 288–298. [Google Scholar] [CrossRef]
- Smith, L. Cause and effect, and how to make a better biocontrol agent. News Inf. 2006, 27, 28N–30N. [Google Scholar]
- Suckling, D.M.; Sforza, R.F.H. What magnitude are observed non-target impacts from weed biocontrol? PLoS ONE 2014, 9, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheppard, A.W.; Haines, M.; Thomann, T. Native range research assists risk analysis for non-targets in weed biological control: The cautionary tale of the broom seed beetle. Aust. J. Entomol. 2006, 45, 292–297. [Google Scholar] [CrossRef]
- Francisco-Ortega, J.; Fernandez-Galvan, M.; Santos-Guerra, A. A literature survey (1696–1991) on the fodder shrubs tagasaste and escobon (Chamaecytisus proliferus(L.fil) Linksensu lato) (Fabaceae: Genisteae). N. Z. J. Agric. Res. 2011, 34, 471–488. [Google Scholar] [CrossRef]
- Nilsen, E.T.; Karpa, D.; Mooney, H.A.; Field, C. Patterns of stem photosynthesis in two invasive legumes (Spartinum junceum, Cytisus scoparius) of the California coastal region. Am. J. Bot. 1993, 80, 1126–1136. [Google Scholar] [CrossRef]
- Pardo, C.; Cubas, P.; Tahiri, H. Molecular phylogeny and systematics of Genista (Leguminosae) and related genera based on nucleotide sequences of nrDNA (ITS region) and cpDNA (trnL-trnF intergenic spacer). Plant Syst. Evol. 2004, 244, 93–119. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Number of Collected Pods | Number of Exit Holes per Pod | Number of Lepidapion argentatum per Plant (Mean ± S.D.) | Number of Parasitoids per Plant | Parasitism Rate per Plant | |
|---|---|---|---|---|---|
| (Mean ± S.D.) | ♂ | ♀ | (Mean ± S.D.) | (Mean ± S.D.) | |
| 81 (per plant) | 1.02 ± 1.06 | 19.3 ± 11.77 | 20.3 ± 11.55 | 10.5 ± 8.44 | 0.22 ± 0.10 |
| N = 810 | N = 825 | N = 193 | N = 203 | N = 105 | |
| Number of Replicates | Exposure Time (Days) | Number of Living Adults (Mean ± S.D.) | Number of Dissected Pods | Pods with Eggs/Total Pods | Number of Eggs | Number of Eggs per Pod | ||
|---|---|---|---|---|---|---|---|---|
| ♂ | ♀ | (Mean ± S.D.) | (Mean ± S.D.) | (Mean ± S.D.) | ||||
| Species Tested in 2012 | ||||||||
| Cytisus-racemosus | 9 | 10 to 16 | 4.67 ± 1.00 | 4.89 ± 0.33 | 6.22 ± 2.86 | 0.73 | 8.11 ± 4.55 | 1.32 ± 0.38 |
| Genista monspessulana | 3 | 12 to 14 | 4.67 ± 0.58 | 5.00 ± 0.00 | 8.33 ± 3.06 | 0.40 | 9.33 ± 6.03 | 1.23 ± 0.84 |
| Lupinus atlanticus consentinii | 6 | 13 to 18 | 3.67 ± 1.03 | 4.33 ± 0.52 | 6.33 ± 2.16 | 0 | 0 | 0 |
| Lupinus albus | 4 | 12 | 2.50 ± 1.73 | 3.75 ± 1.26 | 1.50 ± 0.58 | 0 | 0 | 0 |
| Lupinus angustifolius | 6 | 12 | 2.67 ± 1.37 | 4.50 ± 0.55 | 2.83 ± 1.17 | 0 | 0 | 0 |
| Lupinus consentinii | 12 | 12 to 18 | 2.58 ± 1.51 | 4.00 ± 1.35 | 2.92 ± 1.73 | 0 | 0 | 0 |
| Lupinus luteus | 4 | 12 to 14 | 2.25 ± 1.71 | 4.00 ± 0.82 | 2.25 ± 0.96 | 0 | 0 | 0 |
| Lupinus mutabilis | 8 | 10 to 17 | 1.75 ± 0.89 | 1.88 ± 1.36 | 3.13 ± 1.36 | 0 | 0 | 0 |
| Lupinus pilosus | 3 | 14 to 16 | 4.00 ± 1.00 | 5.00 ± 0.00 | 4.67 ± 1.15 | 0 | 0 | 0 |
| Species tested in 2015 | ||||||||
| Genista monspessulana | 11 | 14 | 4.54 ± 0.82 | 4.72 ± 0.46 | 6.27 ± 4.10 | 0.66 | 8.36 ± 10.37 | 1.12 ± 0.84 |
| Crotalaria sagitalis 1 | 5 | 14 | 2.20 ± 1.48 | 3.40 ± 2.07 | 2.40 ± 1.14 | 0 | 0 | 0 |
| Cytisus scoparius | 5 | 14 | 4.20 ± 0.45 | 3.80 ± 0.45 | NA | 0 | 0 | 0 |
| Glycine max | 5 | 14 | 0 | 0.60 ± 0.89 | 4.60 ± 1.52 | 0 | 0 | 0 |
| Lupinus albus | 5 | 14 | 3.80 ± 0.84 | 3.40 ± 0.89 | 1.20 ± 1.10 | 0 | 0 | 0 |
| Lupinus angustifolius | 5 | 14 | 5.00 ± 0.00 | 5.00 ± 0.00 | 2.00 ± 0.71 | 0 | 0 | 0 |
| Lupinus luteus | 3 | 14 | 3.40 ± 1.34 | 4.40 ± 0.55 | 2.00 ± 1.22 | 0 | 0 | 0 |
| Trifolium repens | 5 | 14 | 0 | 1.20 ± 1.30 | NA | 0 | 0 | 0 |
| Number of Replicates | Number of Living Adults (Mean ± S.D.) | Mean of Impact Damage on Leaves * | Total Number of Exposed Stems | Number of Stems with Galls | Stems with Galls/Total Exposed Stems | Number of Galls | Number of Larvae | ||
|---|---|---|---|---|---|---|---|---|---|
| ♂ | ♀ | (Mean ± S.D.) | (Mean ± S.D.) | ||||||
| Species tested in 2012 | |||||||||
| Genista monspessulana (France) | 5 | 0.80 ± 0.84 | 1.60 ± 1.14 | 2.6 | n/a | NA | NA | 7.80 ± 2.05 | 4.80 ± 0.84 |
| Lupinus albus | 5 | 0.60 ± 0.55 | 1.20 ± 0.84 | 1 | n/a | 0 | n/a | 0 | 0 |
| Lupinus angustifolius | 5 | 1.00 ± 0.71 | 1.60 ± 1.14 | 0.6 | n/a | 0 | n/a | 0 | 0 |
| Lupinus atlanticus consentinii | 5 | 0.40 ± 0.55 | 0.60 ± 0.55 | 0.6 | n/a | 0 | n/a | 0 | 0 |
| Lupinus consentinii | 5 | 0.60 ± 0.55 | 0.60 ± 0.89 | 0.8 | n/a | 0 | n/a | 0 | 0 |
| Lupinus luteus | 5 | 0.40 ± 0.55 | 0.80 ± 0.45 | 0.2 | n/a | 0 | n/a | 0 | 0 |
| Lupinus mutabilis | 5 | 0 | 0 | 0.4 | n/a | 0 | n/a | 0 | 0 |
| Lupinus pilosus | 5 | 0.40 ± 0.55 | 0.80 ± 0.45 | 0.6 | n/a | 0 | n/a | 0 | 0 |
| Species tested in 2015 | |||||||||
| Genista monspessulana (USA) | 5 | 5.00 ± 0.00 | 5.00 ± 0.00 | 3 | 8.60 ± 3.78 | 8.00 ± 3.74 | 0.93 | 14.20 ± 11.86 | 14.20 ± 11.86 |
| Genista monspessulana (France) | 25 | 4.92 ± 0.28 | 4.76 ± 0.43 | 2.96 | 8.88 ± 3.94 | 8.12 ± 3.49 | 0.91 | 13.28 ± 5.43 | 13.08 ± 5.18 |
| Genista linifolia | 5 | 2.40 ± 0.89 | 4.20 ± 0.84 | 2.8 | 8.20 ± 1.92 | 7.40 ± 1.14 | 0.90 | 12.00 ± 1.58 | 12.00 ± 1.58 |
| Cytisus proliferus | 5 | 4.80 ± 0.45 | 5.00 ± 0.00 | 2.4 | 18.2 ± 1.48 | 5.20 ± 4.60 | 0.29 | 7.40 ± 7.30 | 7.40 ± 7.30 |
| Genista stenopetala | 5 | 4.80 ± 0.45 | 4.80 ± 0.45 | 3 | 11.2 ± 3.77 | 4.60 ± 1.82 | 0.41 | 5.60 ± 2.70 | 5.60 ± 2.7 |
| Spartium junceum | 5 | 5.00 ± 0.00 | 5.00 ± 0.00 | 2.2 | 13.4 ± 2.88 | 5.40 ± 2.51 | 0.40 | 6.60 ± 2.97 | 6.60 ± 2.97 |
| Cytisus villosus | 5 | 5.00 ± 0.00 | 4.60 ± 0.55 | 2.2 | 12.8 ± 2.86 | 1.80 ± 1.79 | 0.14 | 2.40 ± 2.07 | 2.40 ± 2.07 |
| Lupinus chamissonis 1 | 5 | 3.80 ± 1.10 | 4.60 ± 0.55 | 1.2 | 9.80 ± 1.92 | 1.60 ± 3.05 | 0.16 | 3.40 ± 7.06 | 3.40 ± 7.06 |
| Lupinus arboreus blue 1 | 5 | 4.60 ± 0.55 | 5.00 ± 0.00 | 1.6 | 8.40 ± 0.55 | 0.60 ± 0.89 | 0.07 | 0.60 ± 0.89 | 0.60 ± 0.89 |
| Amorpha fruticosa1 | 5 | 0 | 0 | 0 | 8.20 ± 1.92 | 0 | 0 | 0 | 0 |
| Astragalus canadensis1 | 5 | 2.20 ± 1.10 | 2.40 ± 1.14 | 0 | 9.20 ± 2.17 | 0 | 0 | 0 | 0 |
| Astragalus nuttalii1 | 5 | 0.40 ± 0.89 | 0.60 ± 0.89 | 0 | 7.80 ± 1.30 | 0 | 0 | 0 | 0 |
| Astragalus trichopodus1 | 5 | 1.00 ± 1.22 | 1.60 ± 2.30 | 0 | 8.40 ± 1.14 | 0 | 0 | 0 | 0 |
| Baptisia australis1 | 2 | 0.50 ± 0.71 | 3.00 ± 0.00 | 0 | 2.50 ± 2.12 | 0 | 0 | 0 | 0 |
| Baptisia bracteata1 | 5 | 1.40 ± 1.14 | 3.00 ± 1.41 | 0 | 1.00 ± 0.00 | 0 | 0 | 0 | 0 |
| Cladrastis lutea1 | 2 | 0 | 1.50 ± 2.12 | 1 | 3.50 ± 0.70 | 0 | 0 | 0 | 0 |
| Crotalaria sagitalis1 | 5 | 0.20 ± 0.45 | 4.00 ± 0.71 | 0.2 | 7.60 ± 1.14 | 0 | 0 | 0 | 0 |
| Cytisus scoparius | 5 | 4.40 ± 0.89 | 4.20 ± 0.84 | 0.6 | 9.80 ± 3.83 | 0 | 0 | 0 | 0 |
| Cytisus striatus | 5 | 4.40 ± 0.55 | 4.80 ± 0.45 | 0.6 | 14.8 ± 4.44 | 0 | 0 | 0 | 0 |
| Glycine max | 5 | 0 | 0.80 ± 0.84 | 0 | 4.60 ± 0.89 | 0 | 0 | 0 | 0 |
| Hoita macrostachya1 | 5 | 0 | 0 | 0 | 10.6 ± 1.14 | 0 | 0 | 0 | 0 |
| Lathyrus vestitus1 | 5 | 0 | 0.40 ± 0.55 | 0 | 5.80 ± 1.79 | 0 | 0 | 0 | 0 |
| Lotus scoparius1 | 5 | 0 | 0 | 0 | 19.2 ± 4.44 | 0 | 0 | 0 | 0 |
| Lupinus albus | 5 | 4.80 ± 0.45 | 5.00 ± 0.00 | 2.2 | 12.8 ± 3.49 | 0 | 0 | 0 | 0 |
| Lupinus angustifolius | 5 | 3.00 ± 0.71 | 4.00 ± 0.71 | 0.8 | 31.25 ± 12.12 | 0 | 0 | 0 | 0 |
| Lupinus luteus | 5 | 1.60 ± 1.82 | 2.80 ± 2.17 | 1.2 | 7.60 ± 3.91 | 0 | 0 | 0 | 0 |
| Lupinus microcarpus densiflorus1 | 5 | 0 | 0.40 ± 0.55 | 0.4 | 7.00 ± 1.41 | 0 | 0 | 0 | 0 |
| Lupinus pilosus | 5 | 3.00 ± 1.87 | 4.80 ± 0.45 | 0.2 | 9.80 ± 5.81 | 0 | 0 | 0 | 0 |
| Lupinus texensis1 | 5 | 4.00 ± 1.00 | 4.40 ± 0.55 | 1.4 | 12.4 ± 2.70 | 0 | 0 | 0 | 0 |
| Sesbania exaltata1 | 5 | 0.25 ± 0.45 | 0.25 ± 0.45 | 0 | 17.0 ± 4.64 | 0 | 0 | 0 | 0 |
| Thermopsis macrophylla1 | 5 | 3.40 ± 1.14 | 3.60 ± 1.14 | 0.8 | 6.40 ± 0.55 | 0 | 0 | 0 | 0 |
| Thermopsis montana1 | 5 | 4.20 ± 0.84 | 4.60 ± 0.55 | 0.8 | 5.80 ± 1.10 | 0 | 0 | 0 | 0 |
| Vicia faba | 5 | 0 | 0.60 ± 0.89 | 0 | 6.60 ± 1.14 | 0 | 0 | 0 | 0 |
| Number of Replicates (Cages) | Number of Adults Introduced in Each Cage (and Exposure Time) | Number of Living Adults (Mean ± S.D.) | Mean Impact Damage on Leaves * | Stems with Galls/Total Exposed Stems | Number of Galls | Number of Larvae | |||
|---|---|---|---|---|---|---|---|---|---|
| Species | Phase 1 | Phase 2 | ♂ | ♀ | (Mean ± S.D.) | (Mean ± S.D.) | |||
| Genista monspessulana (Control 1) | 5 | 18 ♀ + 15 ♂ (3 days) | 0 | 1.60 ± 1.82 | 2.00 ± 2.91 | 1.20 | 0.20 | 3.60 ± 8.05 | 3.60 ± 8.05 |
| Lupinus albus | 15 ♀ + 12 ♂ (10 days) | 0.60 ± 0.89 | 1.80 ± 1.09 | 0.80 | 0 | 0 | 0 | ||
| Lupinus angustifolius | 0 | 0 | 0.40 | 0 | 0 | 0 | |||
| Lupinus arboreus blue1 | 0.40 ± 0.89 | 0.40 ± 0.89 | 0.20 | 0 | 0 | 0 | |||
| Lupinus luteus | 0.40 ± 0.89 | 2.00 ± 3.46 | 0.80 | 0 | 0 | 0 | |||
| Lupinus microcarpus densiflorus1 | 0.20 ± 0.45 | 1.00 ± 1.22 | 0.40 | 0 | 0 | 0 | |||
| Genista monspessulana (Control 2) | 5 | 0 | 3 ♀ + 3 ♂ (10 days) | 2.20 ± 0.84 | 2.60 ± 0.55 | 2.00 | 0.87 | 15.20 ± 7.05 | 15.20 ± 7.05 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kerdellant, E.; Thomann, T.; Sheppard, A.; Sforza, R.F.H. Host Specificity and Preliminary Impact of Lepidapion argentatum (Coleoptera, Brentidae), a Biocontrol Candidate for French Broom (Genista monspessulana, Fabaceae). Insects 2021, 12, 691. https://doi.org/10.3390/insects12080691
Kerdellant E, Thomann T, Sheppard A, Sforza RFH. Host Specificity and Preliminary Impact of Lepidapion argentatum (Coleoptera, Brentidae), a Biocontrol Candidate for French Broom (Genista monspessulana, Fabaceae). Insects. 2021; 12(8):691. https://doi.org/10.3390/insects12080691
Chicago/Turabian StyleKerdellant, Elven, Thierry Thomann, Andy Sheppard, and René F. H. Sforza. 2021. "Host Specificity and Preliminary Impact of Lepidapion argentatum (Coleoptera, Brentidae), a Biocontrol Candidate for French Broom (Genista monspessulana, Fabaceae)" Insects 12, no. 8: 691. https://doi.org/10.3390/insects12080691
APA StyleKerdellant, E., Thomann, T., Sheppard, A., & Sforza, R. F. H. (2021). Host Specificity and Preliminary Impact of Lepidapion argentatum (Coleoptera, Brentidae), a Biocontrol Candidate for French Broom (Genista monspessulana, Fabaceae). Insects, 12(8), 691. https://doi.org/10.3390/insects12080691