
Are Dried and Powdered Moringa oleifera Lam. Leaves Susceptible to Moths That Feed on Stored Products?


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Rearing
2.2. Egg Collection
2.3. Tests
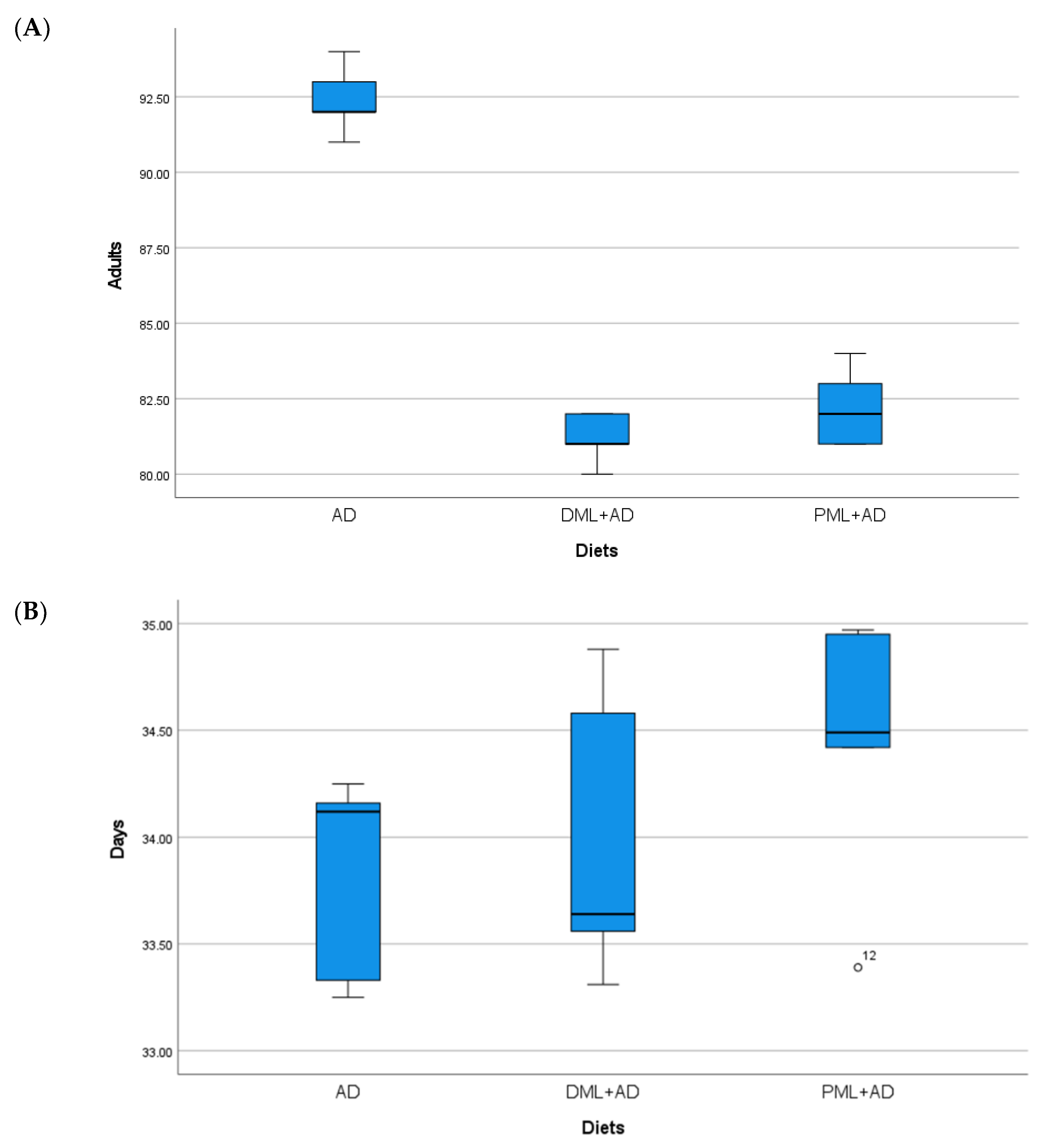
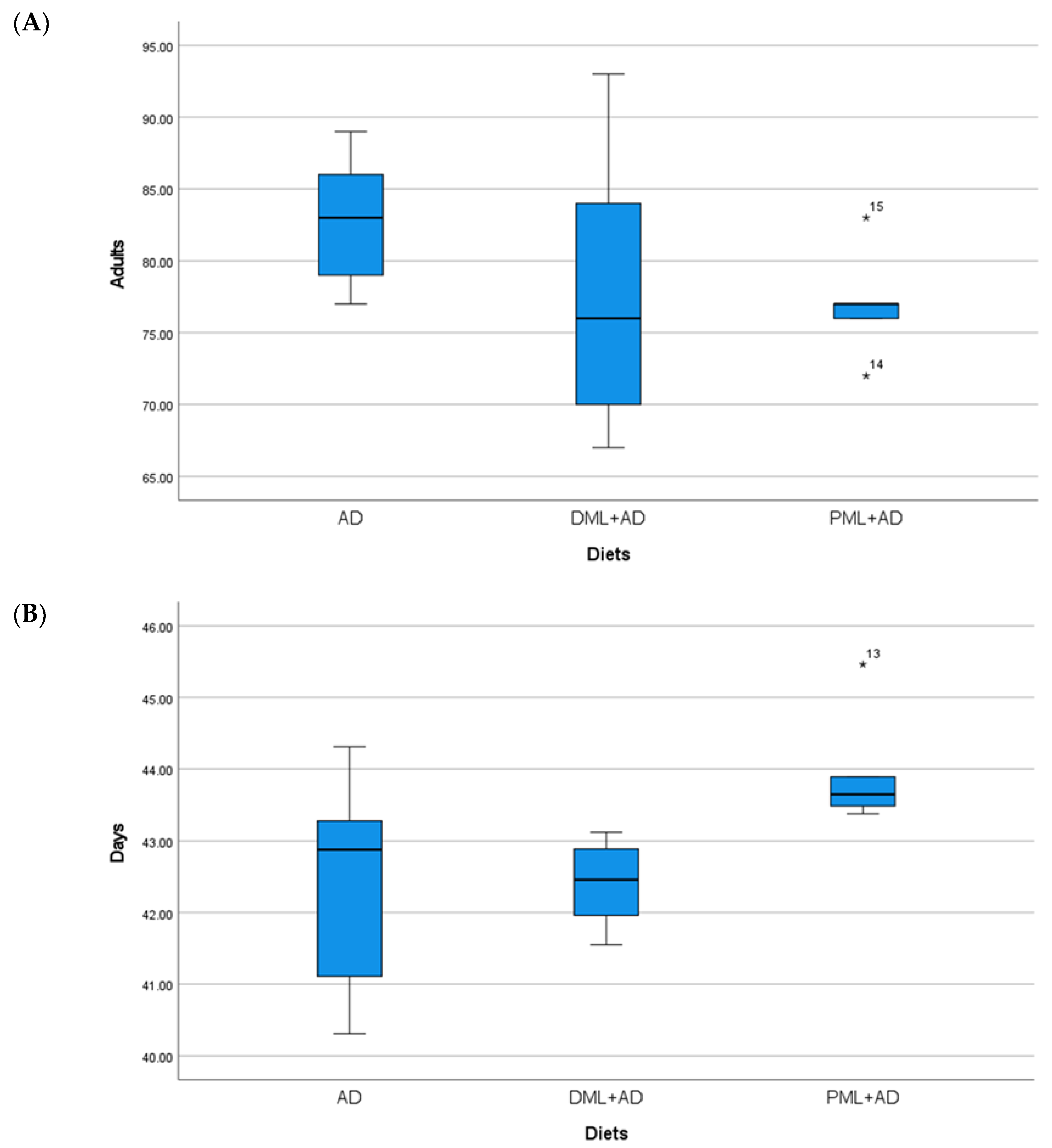
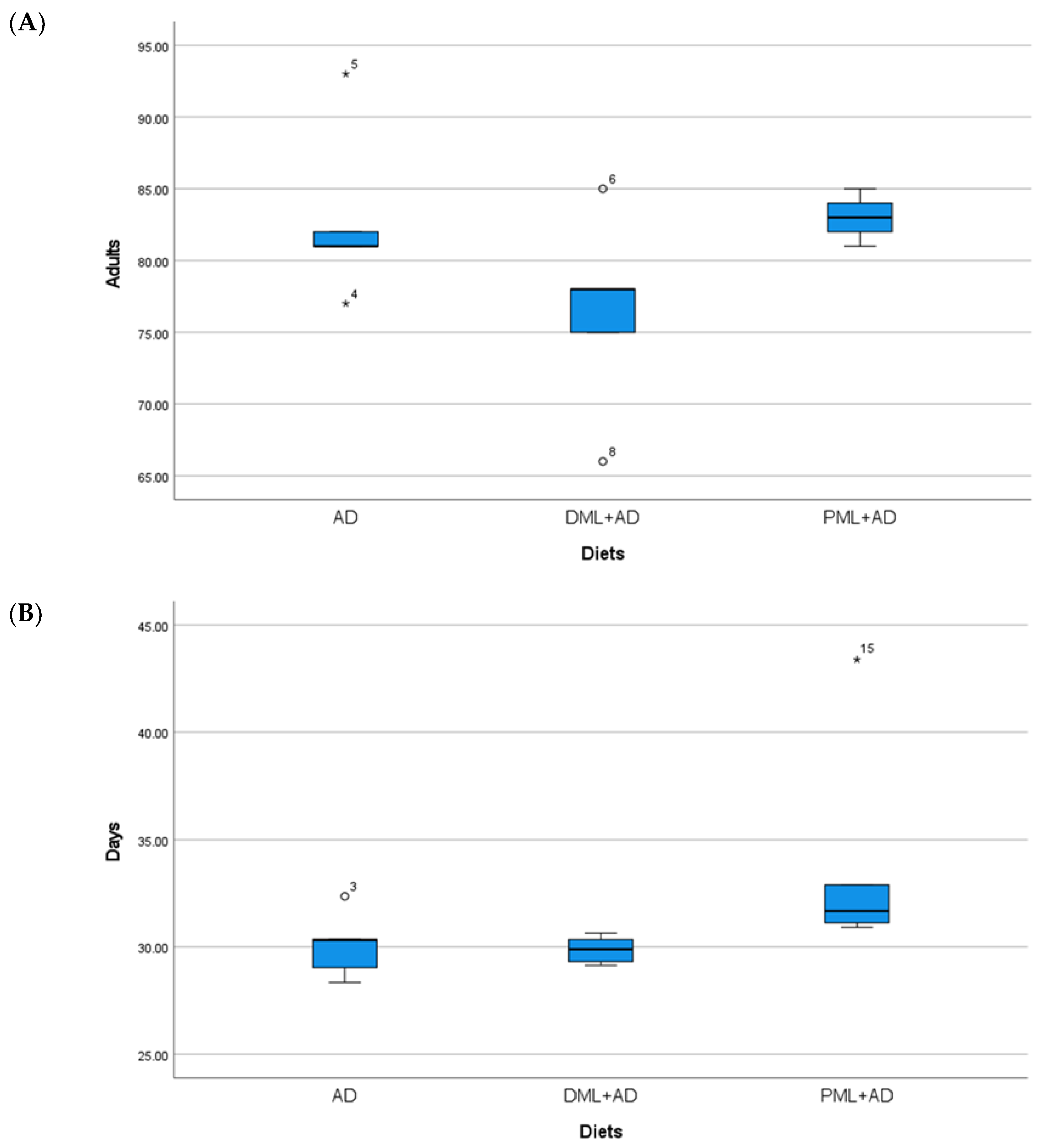
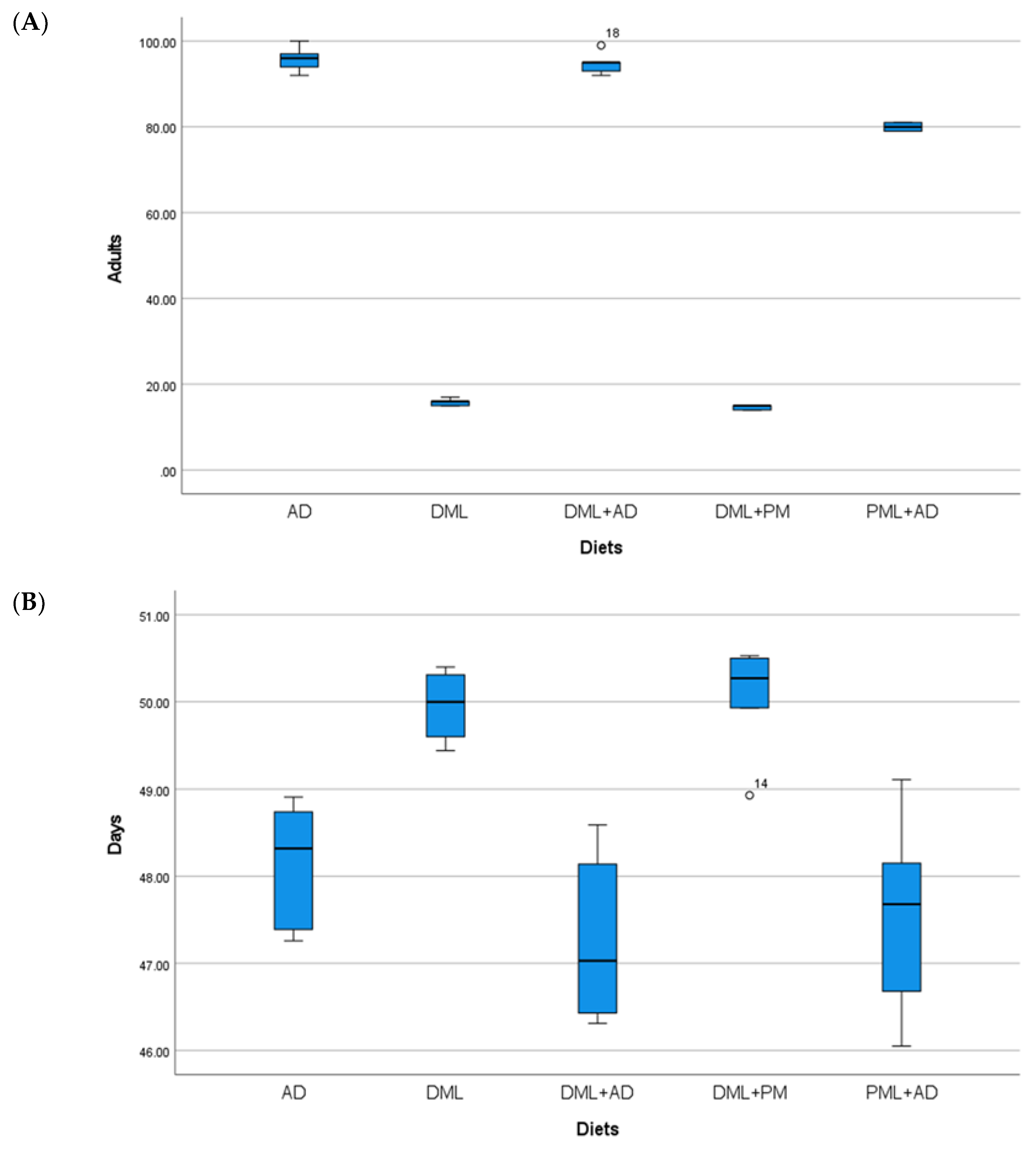
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Maida, A.; Rarooq, A.; Razyia, N.; Umer, R.; Kazi, T.G.; Nadeem, M. Mineral composition of Moringa oleifera leaves and pods from different regions of Punjab, Pakistan. Asian J. Plant Sci. 2005, 4, 417–421. [Google Scholar]
- Jongrungruangchok, S.; Bunrathep, S.; Songsak, T. Nutrients and minerals content of eleven different samples of Moringa oleifera cultivated in Thailand. J. Health Res. 2018, 24, 123–127. [Google Scholar]
- Leone, A.; Spada, A.; Battezzati, A.; Schiraldi, A.; Aristil, J.; Bertoli, S. Moringa oleifera seeds and oil: Characteristics and uses for human health. Int. J. Mol. Sci. 2016, 17, 2141. [Google Scholar] [CrossRef] [Green Version]
- Heykal, A.; El Wahab, A.E.A.; Asran, A.M. Biological studies on Plodia interpunctella Hbn. on certain medical and aromatical dried plants (Lepidoptera: Phycitidae). Agric. Res. Rev. 1978, 56, 147–149. [Google Scholar]
- Wahab, A.E.A.; El Halfawy, M.A.; Heykal, A.; Asran, A.M. Biology of Anagasta kuehniella Zell. on certain medical and aromatical dried plants (Lepidoptera; Phycitidae). Agric. Res. Rev. 1978, 56, 143–146. [Google Scholar]
- Stampini, M. Development observations of Plodia interpunctella (Hbn.) (Lepidoptera Pyralidae) on some dried medicinal plants. Boll. Zool. Agr. Bachic. 2009, 41, 41–50. [Google Scholar]
- Jagadish, P.S.; Nirmala, P.; Rashmi, M.A.; Neelu, N. Biology of rice moth Corcyra cephalonica Stainton on foxtail millet. Karnataka J. Agric. Sci. 2009, 22, 674–675. [Google Scholar]
- Soderstrom, E.; Hinsch, R.; Bongers, A.; Brandl, D.; Hoogendorn, H. Detecting adult Phycitinae (Lepidoptera: Pyralidae) infesta-tions in a raisin-marketing channel. J. Econ. Entomol. 1987, 80, 1229–1232. [Google Scholar] [CrossRef]
- Sinha, R.N.; Watters, F.L. Insect Pests of Flour Mills, Grain Elevators, and Feed Mills and Their Control; Agriculture Canada Publication: Ottawa, ON, Canada, 1985; pp. 1–290. [Google Scholar]
- Cox, P.D. The suitability of dried fruits, almonds and carobs for the development of Ephestia figulilella Gregson, E. calidella (Guenee) and E. cautella (Walker) (Lepidoptera: Phycitidae). J. Stored Prod. Res. 1975, 11, 229–233. [Google Scholar] [CrossRef]
- Na, J.H.; Ryoo, M.I. The influence of temperature on development of Plodia interpunctella (Lepidoptera: Pyralidae) on dried vegetable commodities. J. Stored Prod. Res. 2000, 36, 125–129. [Google Scholar] [CrossRef]
- Burks, C.S.; Johnson, J.A. Biology, behavior, and ecology of stored fruit and nut insects. In Stored Product Protection; Hagstrum, D.W., Phillips, T.W., Cuperus, G., Eds.; Kansas State University: Manhattan, KA, USA, 2012; pp. 21–32. [Google Scholar]
- Binder, M.; Mahler, V.; Hayek, B.; Sperr, W.R.; Scholler, M.; Prozell, S.; Wiedermann, G.; Valent, P.; Valenta, R.; Duchêne, M. Molecular and immunological characterization of arginine kinase from the Indianmeal moth, Plodia interpunctella, a novel cross-reactive invertebrate pan-allergen. J. Immunol. 2001, 167, 5470–5477. [Google Scholar] [CrossRef] [Green Version]
- Hubert, J.; Stejskal, V.; Athanassiou, C.G.; Throne, J.E. Health hazards associated with arthropod infestation of stored products. Ann. Rev. Entomol. 2018, 63, 553–573. [Google Scholar] [CrossRef]
- South, R. The Moth of the British Isles; Second Series; Warne: London, UK; New York, NY, USA, 1961; pp. 1–379. [Google Scholar]
- Koch, M. Wir Bestimmen Schmetterlinge; Neumann-Neudamm: Melsunghen, Germany, 1984; pp. 1–792. [Google Scholar]
- Skinner, B. Moth of the British Isles; Viking, Penguin Books: Harmondsworth, UK, 1984; pp. 1–267. [Google Scholar]
- Locatelli, D.P.; Di Egidio, V.; Stampini, M. Observation of the development of Idaea inquinata (Scop.) (Lepidoptera Geometridae) on medicinal plants and other food substrates. Boll. Zool. Agr. Bachic. Ser II 2005, 37, 123–132. [Google Scholar]
- Locatelli, D.P.; Savoldelli, S.; Girgenti, P.; Lucchini, G.A.; Limonta, L. Can environmental dust from silo area allow the development of stored product insects? J. Stored Prod. Res. 2017, 71, 41–46. [Google Scholar] [CrossRef]
- Limonta, L.; Stampini, M.; Locatelli, D.P. Development of rusty wave Idaea inquinata at constant temperatures, relative humidities and photoperiod. Bull. Insectology 2010, 63, 171–174. [Google Scholar]
- Prosky, L.; Asp, N.G.; Schweizer, T.F.; DeVries, J.W.; Furda, I. Determination of insoluble, soluble, and total dietary fiber in foods and food products: Interlaboratory study. JAOAC 1988, 71, 1017–1023. [Google Scholar] [CrossRef]
- Rocklin, R.D.; Pohl, C.A. Determination of Carbohydrates by Anion Exchange Chromatography with Pulsed Amperometric Detection. J. Liq. Chromatogr. 1983, 6, 1577–1590. [Google Scholar] [CrossRef]
- Anonymous. AOAC 34.01.05 n.925.31, Nitrogen in eggs. In Official Methods of Analysis of AOAC International, 16th ed.; Cunniff, P., Ed.; Publisher AOAC International: Gaithersburg, MD, USA, 1995; Ch. 34; p. 2. [Google Scholar]
- Anonymous. AOAC 31.04.02 n. 963.15, Fat in cacao products. In Official Methods of Analysis of AOAC International, 16th ed.; Cunniff, P., Ed.; AOAC International: Gaithersburg, MD, USA, 1996; Ch. 31; p. 10. [Google Scholar]
- AACC 08-01.01. AACC Approved Method of Analysis, 11th ed. Available online: http://methods.aaccnet.org/summaries/08-01-01.aspx (accessed on 10 May 2021).
- Leone, A.; Fiorillo, G.; Criscuoli, F.; Ravasenghi, S.; Santagostini, L.; Fico, G.; Spadafranca, A.; Battezzati, A.; Schiraldi, A.; Pozzi, F.; et al. Nutritional characterization and phenolic profiling of Moringa oleifera leaves grown in Chad, Sahrawi Refugee Camps, and Haiti. Int. J. Mol. Sci. 2015, 16, 18923–18937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silhacek, D.; Murphy, C. Moisture content in a wheat germ diet and its effect on the growth of Plodia interpunctella (Hubner). J. Stored Prod. Res. 2008, 44, 36–40. [Google Scholar] [CrossRef]
- Ashfaq, M.; Shahzad, M.A.; Basra Ashfaq, U. Moringa: A miracle plant for agro-forestry. J. Agri. Sci. 2012, 8, 115–122. [Google Scholar]
- Musa, A.K. Influence of plant powders on infestation by adults and larvae of Khapra Beetle, Trogoderma granarium Everts (Coleoptera: Dermestidae) in stored groundnut. AJBAS 2013, 7, 427–432. [Google Scholar]
- Kasolo, J.N.; Bimenya, G.S.; Ojok, L.; Ochieng, J.; Ogwa-Okeng, J.W. Phytochemical and uses of Moringa oleifera leaves in Ugandan rural communities. J. Med. Plants Res. 2010, 4, 753–757. [Google Scholar]
- Moreira, X.; Galman, A.; Francisco, M.; Castagneyrol, B.; Abdala-Roberts, L. Host plant frequency and secondary metabolites are concurrently associated with insect herbivory in a dominant riparian tree. Biol. Lett. 2018, 14, 20180281. [Google Scholar] [CrossRef] [Green Version]
- Su, Q.; Zhou, Z.; Zhang, J.; Shi, C.; Zhang, G.; Jin, Z.; Wang, W.; Li, C. Effect of plant secondary metabolites on common cutworm, Spodoptera litura (Lepidoptera: Noctuidae). Entomol. Res. 2018, 48, 18–26. [Google Scholar] [CrossRef]
- Motevalli-Haghi, S.F.; Fathi, M.; Ebrahimzadeh, M.A.; Eslami, S.; Karamie, M.; Eslamifar, M.; Dehghan, O. Evaluation of Phytochemical, total phenolic and flavonoid content, antioxidant activities and repelling property of Sambucus ebulus. JMPB 2020, 1, 97–105. [Google Scholar]
- Qasim, M.; Islam, W.; Ashraf, J.H.; Ali, I.; Wang, L. Saponins in Insect Pest Control. In Co-Evolution of Secondary Metabolites; Springer Nature Switzerland AG: Cham, Switzerland, 2020; pp. 898–924. [Google Scholar]
- Juntheikki, M.R.; Julkunene-Tiitto, R. Inibition of beta-glucosidase and esterase by tannins from Betula, Salix and Pinus species. J. Chem. Ecol. 2000, 26, 1151–1165. [Google Scholar] [CrossRef]
- Pospisil, J. The response of Leptinotarsa decemlineata (Coleoptera) to tannin as an antifeedant. Acta Entomol. Bohemoslov. 1982, 6, 429–434. [Google Scholar]
- Manuwoto, S.; Scriber, J.M. Effects of hydrolysable and condensed tannin on growth and development of two species of polyphagous Lepidoptera: Spodoptera eridania and Callosomia promethean. Oecology 1986, 69, 225–230. [Google Scholar] [CrossRef]
- El-Halfawy, M.A. Entomofauna of dried medical and aromatical plants with a short note on their occurrence and populations. Agric. Res. Rev. 1977, 55, 103–160. [Google Scholar]
- Kalinovič, I.; Rozman, V. Infestation of stored medicinal plants and herbal tea by insects and mites. Plant Prot. Sci. 2000, 36, 21–22. [Google Scholar] [CrossRef]
- Arbogast, R.T.; Kendra, P.E.; Mankin, R.W.; McDonald, R.C. Insect infestation of a botanicals warehouse in north-central Florida. J. Stored Prod. Res. 2002, 38, 349–363. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diets. | Cadra cautella | Corcyra cephalonica | Plodia interpunctella | Idaea inquinata | ||||
|---|---|---|---|---|---|---|---|---|
| Adults | Days | Adults | Days | Adults | Days | Adults | Days | |
| AD | 92.4 ± 0.51 a | 33.8 ± 0.22 | 82.8 ± 2.20 | 42.4 ± 0.73 | 82.8 ± 2.69 a | 30.1 ± 0.69 a,b | 95.8 ± 1.36 a | 48.1 ± 0.34 b |
| DML | 0.0 ± 0.00 | - | 0.0 ± 0.00 | - | 0.0 ± 0.00 | - | 15.8 ± 0.37 c | 49.9 ± 0.18 a |
| PML | 0.0 ± 0.00 | - | 0.0 ± 0.00 | - | 0.0 ± 0.00 | - | 0.0 ± 0.00 | - |
| ½DML + ½PML | 0.0 ± 0.00 | - | 0.0 ± 0.00 | - | 0.0 ± 0.00 | - | 14.6 ± 0.24 c | 50.0 ± 0.29 a |
| ¼DML + ¾AD | 81.2 ± 0.37 b | 34.2 ± 0.30 | 78.0 ± 4.74 | 42.4 ± 0.29 | 76.4 ± 3.07 b | 29.9 ± 0.29 b | 94.8 ± 1.20 a | 47.3 ± 0.47 b |
| ¼PML + ¾AD | 82.2 ± 0.58 b | 34.4 ± 0.29 | 77.0 ± 1.76 | 44.0 ± 0.38 | 83.0 ± 0.71 a | 32.0 ± 0.50 a | 80.0 ± 0.45 b | 47.5 ± 0.54 b |
| Nutrients | Idaea | Pyralidae |
|---|---|---|
| Proteins | 13.9 ± 0.261 | 8.6 ± 0.05 |
| Lipids | 2.9 ± 0.12 | 2.0 ± 0.19 |
| Starch | 22.0 ± 0.79 | 29.3 ± 1.13 |
| Soluble carbohydrates * | 8.1 ± 0.27 | 13.8 ± 0.21 |
| Total fiber | 27.6 ± 1.01 | 9.7 ± 0.37 |
| Ashes | 3.6 ± 0.08 | 1.4 ± 0.02 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Limonta, L.; Locatelli, D.P. Are Dried and Powdered Moringa oleifera Lam. Leaves Susceptible to Moths That Feed on Stored Products? Insects 2021, 12, 610. https://doi.org/10.3390/insects12070610
Limonta L, Locatelli DP. Are Dried and Powdered Moringa oleifera Lam. Leaves Susceptible to Moths That Feed on Stored Products? Insects. 2021; 12(7):610. https://doi.org/10.3390/insects12070610
Chicago/Turabian StyleLimonta, Lidia, and Daria Patrizia Locatelli. 2021. "Are Dried and Powdered Moringa oleifera Lam. Leaves Susceptible to Moths That Feed on Stored Products?" Insects 12, no. 7: 610. https://doi.org/10.3390/insects12070610
APA StyleLimonta, L., & Locatelli, D. P. (2021). Are Dried and Powdered Moringa oleifera Lam. Leaves Susceptible to Moths That Feed on Stored Products? Insects, 12(7), 610. https://doi.org/10.3390/insects12070610