
Evaluating the Potential of Using Spodoptera litura Eggs for Mass-Rearing Telenomus remus, a Promising Egg Parasitoid of Spodoptera frugiperda



Abstract
Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. Insect Culture
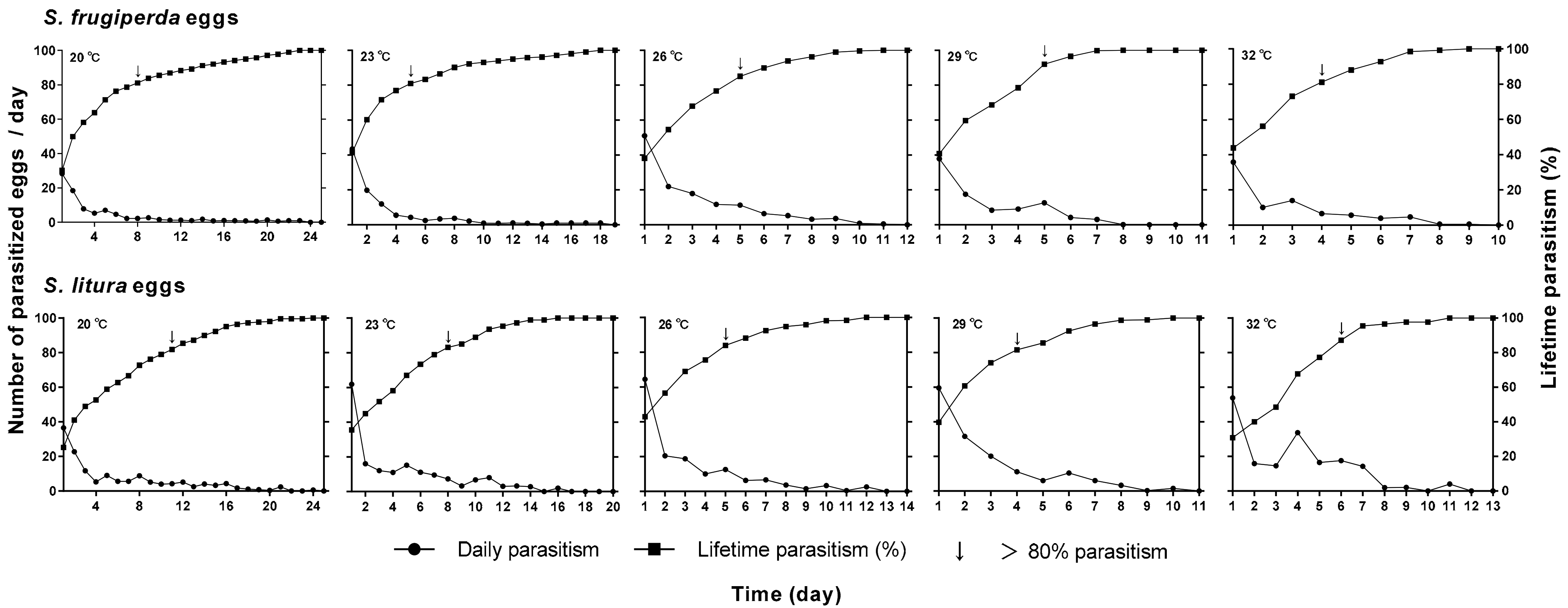
2.2. Effects of Temperature and Host Species on T. remus Development
2.3. Effects of Temperature and Host Species on the Fecundity and Lifespan of T. remus
2.4. Statistical Analysis
3. Results
3.1. Development of T. remus Reared on Eggs of S. frugiperda or S. litura
3.2. Thermal Requirements of T. remus Reared on the Eggs of S. frugiperda or S. litura
3.3. Biological Characteristics of T. remus Reared on Eggs of S. frugiperda or S. litura
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- CABI. Spodoptera Frugiperda (Fall Armyworm). Available online: https://www.cabi.org/isc/datasheet/29810#94987198-9f50-4173-8bbd-30bd93840e73?tdsourcetag=s_pcqq_aiomsg (accessed on 1 September 2020).
- Montezano, D.G.; Specht, A.; Sosa-Gómez, D.R.; Roque-Specht, V.F.; Sousa-Silva, J.C.; Paula-Moraes, S.V.; Peterson, J.A.; Hunt, T.E. Host plants of Spodoptera frugiperda (Lepidoptera: Noctuidae) in the Americas. Afr. Entomol. 2018, 26, 286–300. [Google Scholar] [CrossRef]
- Day, R.; Abrahams, P.; Bateman, M.; Beale, T.; Clottey, V.; Cock, M.; Colmenarez, Y.; Corniani, N.; Early, R.; Godwin, J.; et al. Fall armyworm: Impacts and implications for Africa. Outlooks Pest Manag. 2017, 28, 196–201. [Google Scholar] [CrossRef]
- Wu, K.M. Management strategies of fall armyworm (Spodoptera frugiperda) in China. Plant Prot. 2020, 46, 1–5. [Google Scholar] [CrossRef]
- Okuma, D.M.; Bernardi, D.; Horikoshi, R.J.; Bernardi, O.; Silva, A.P.; Omoto, C. Inheritance and fitness costs of Spodoptera frugiperda (Lepidoptera: Noctuidae) resistance to spinosad in Brazil. Pest Manag. Sci. 2017, 78, 1441–1448. [Google Scholar] [CrossRef]
- Guo, J.F.; Wu, S.Y.; Zhang, F.; Huang, C.L.; He, K.L.; Babendreier, D.; Wang, Z.Y. Prospects for microbial control of the fall armyworm Spodoptera frugiperda: A review. BioControl 2020, 65, 647–662. [Google Scholar] [CrossRef]
- Shylesha, A.N.; Jalali, S.K.; Gupta, A.; Varshney, R.; Venkatesan, T.; Shetty, P.; Ojha, R.; Ganiger, P.C.; Navik, O.; Subaharan, K.; et al. Studies on new invasive pest Spodoptera frugiperda (J. E. Smith) (Lepidoptera: Noctuidae) and its natural enemies. J. Biol. Control 2018, 32, 145–151. [Google Scholar] [CrossRef]
- Chen, W.B.; Li, Y.Y.; Wang, M.Q.; Liu, C.X.; Mao, J.J.; Chen, H.Y.; Zhang, L.S. Natural enemy insect resources of the fall armyworm Spodoptera frugiperda, their application status, and existing problems and suggestions. Chin. J. Biol. Control 2019, 34, 658–673. [Google Scholar] [CrossRef]
- Chen, W.B.; Li, Y.Y.; Wang, M.Q.; Liu, C.X.; Mao, J.J.; Chen, H.Y.; Zhang, L.S. Entomopathogen resources of the fall armyworm Spodoptera frugiperda, and their application status. Plant Prot. 2019, 45, 1–9. [Google Scholar] [CrossRef]
- Salazar-Mendoza, P.; Rodriguez-Saona, C.; Fernandes, O.A. Release density, dispersal capacity, and optimal rearing conditions for Telenomus remus, an egg parasitoid of Spodoptera frugiperda, in maize. Biocontrol Sci. Technol. 2020, 30, 1040–1059. [Google Scholar] [CrossRef]
- Kenis, M.; du Plessis, H.; Van den Berg, J.; Ba, M.N.; Goergen, G.; Kwadjo, K.E.; Baoua, I.; Tefera, T.; Buddie, A.; Cafà, G.; et al. Telenomus remus, a candidate parasitoid for the biological control of Spodoptera frugiperda in Africa, is already present on the continent. Insects 2019, 10, 92. [Google Scholar] [CrossRef]
- Cave, R.D. Biology, ecology and use in pest management of Telenomus remus. Biocontrol News Inf. 2000, 21, 21–26. [Google Scholar]
- Pomari, A.F.; Bueno, A.F.; Bueno, R.C.O.F.; Junior, M.; de Oliceriras, A.; Fonseca, A.C.P.F. Releasing number of Telenomus remus (Nixon) (Hymenoptera: Platygastridae) against Spodoptera frugiperda Smith (Lepidoptera: Noctuidae) in corn, cotton and soybean. Ciência Rural. 2013, 43, 377–382. [Google Scholar] [CrossRef]
- Huo, L.X.; Zhou, J.C.; Ning, S.F.; Zhao, Q.; Zhang, L.X.; Zhang, Z.T.; Zhang, L.S.; Dong, H. Biological characteristics of Telenomus remus against Spodoptera frugiperda and Spodoptera litura eggs. Plant Prot. 2019, 45, 60–64. [Google Scholar] [CrossRef]
- Queiroz, A.P.; Bueno, A.F.; Pomari-Fernandes, A.; Grande, M.L.M.; Bortolotto, O.C.; Silva, D.M. Quality control of Telenomus remus (Hymenoptera: Platygastridae) reared on the factitious host Corcyra cephalonica (Lepidoptera: Pyralidae) for successive generations. Bull. Entomol. Res. 2017, 107, 791–798. [Google Scholar] [CrossRef]
- Queiroz, A.P.; Bueno, A.F.; Pomari-Fernandes, A.; Grande, M.L.M.; Bortolotto, O.C.; Silva, D.M. Low temperature storage of Telenomus remus (Nixon) (Hymenoptera: Platygastridae) and its factitious host Corcyra cephalonica (Stainton) (Lepidoptera: Pyralidae). Neotrop. Entomol. 2017, 46, 182–192. [Google Scholar] [CrossRef]
- Jalali, S.K.; Venkatesan, T.; Murthy, K.S.; Biswas, S.R.; Lalitha, Y. Influence of temperature and host density on functional response of Telenomus remus Nixon, an egg parasitoid of Spodoptera litura Fabricius. Entomon-Trivandrum 2005, 30, 193–199. [Google Scholar]
- Zhao, S.Y.; Luo, Q.M.; Sun, X.X.; Yang, X.M.; Jiang, Y.Y.; Wu, K.M. Comparison of morphological and biological characteristics between Spodoptera frugiperda and Spodoptera litura. Chin. Plant Prot. 2019, 39, 26–35. [Google Scholar]
- Iranipour, S.; Bonab, Z.N.; Michaud, J.P. Thermal requirements of Trissolcus grandis (Hymenoptera: Scelionidae), an egg parasitoid of sunn pest. Eur. J. Entomol. 2010, 107, 47–53. [Google Scholar] [CrossRef]
- Hougardy, E.; Hogg, B.N.; Wang, X.G.; Daane, K.M. Comparison of thermal performances of two Asian larval parasitoids of Drosophila suzukii. Biol. Control. 2019, 136, 10400. [Google Scholar] [CrossRef]
- Bueno, R.C.O.F.; Carneiro, T.R.; Pratissoli, D.; Bueno, A.F.; Fernandes, O.A. Biology and thermal requirements of Telenomus remus reared on fall armyworm Spodoptera frugiperda eggs. Cienc. Rural. 2008, 38, 1–6. [Google Scholar] [CrossRef]
- Greene, G.L.; Leppla, N.C.; Dickerson, W.A. Velvetbean caterpillar: A rearing procedure and artificial medium. J. Econ. Entomol. 1976, 69, 487–488. [Google Scholar] [CrossRef]
- Chen, Q.J.; Li, G.H.; Pang, Y. A simple artificial diet for mass rearing of some noctuid species. Entomol. Knowl. 2000, 37, 325–327. [Google Scholar]
- Campbell, A.; Frazer, B.D.; Gilbert, N.; Mackauer, M. Temperature requirements of some aphids and their parasites. J. Appl. Ecol. 1974, 11, 431–438. [Google Scholar] [CrossRef]
- Pomari, A.F.; Bueno, A.F.; Bueno, R.C.O.F.; Junior, A.O.M. Biological characteristics and thermal requirements of the biological control agent Telenomus remus (Hymenoptera: Platygastridae) reared on eggs of different species of the Genus Spodoptera (Lepidoptera: Noctuidae). Ann. Entomol. Soc. Am. 2012, 105, 73–81. [Google Scholar] [CrossRef]
- Pinto, J.R.L.; Fernandes, O.A. Parasitism capacity of Telenomus remus and Trichogramma pretiosum on eggs of moth pests of peanut. Bull. Insectol. 2020, 73, 71–78. [Google Scholar]
- Pak, G.A.; Buis, H.C.E.M.; Heck, I.C.C.; Hermans, M.L.G. Behavioural variations among strains of Trichogramma spp. Host-age selection. Entomol. Exp. Appl. 1986, 40, 247–258. [Google Scholar] [CrossRef]
- Bueno, R.C.O.F.; Parra, J.R.P.; Bueno, A.F. Biological characteristics and thermal requirements of a Brazilian strain of the parasitoid Trichogramma pretiosum reared on eggs of Pseudoplusia includens and Anticarsia gemmatalis. Biol. Control. 2009, 51, 355–361. [Google Scholar] [CrossRef]
- CABI. Spodoptera litura (Taro Caterpillar). Available online: https://www.cabi.org/isc/datasheet/44520 (accessed on 11 December 2020).
- Bai, B.; Luck, R.F.; Forster, L.; Stephens, B.; Janssen, J.A.M. The effect of host size on quality attributes of the egg parasitoid, Trichogramma pretiosum. Entomol. Exp. Appl. 1992, 64, 37–48. [Google Scholar] [CrossRef]
- Bueno, R.C.O.F.; Carneiro, T.R.; Bueno, A.F.; Pratissoli, D.; Fernandes, O.A.; Vieira, S.S. Parasitism capacity of Telenomus remus Nixon (Hymenoptera: Scelionidae) on Spodoptera frugiperda (Smith) (Lepidoptera: Noctuidae) eggs. Braz. Arch. Biol. Techn. 2010, 53, 133–139. [Google Scholar] [CrossRef]
- Kang, L.; Chen, B.; Wei, J.N.; Liu, T.X. Roles of thermal adaptation and chemical ecology in Liriomyza distribution and control. Annu. Rev. Entomol. 2009, 54, 127–145. [Google Scholar] [CrossRef]
- Sunday, J.M.; Bates, A.E.; Kearney, M.R.; Colwell, R.K.; Dulvy, N.K.; Longino, J.T.; Huey, R.B. Thermal-safety margins and the necessity of thermoregulatory behavior across latitude and elevation. Proc. Natl. Acad. Sci. USA 2014, 111, 5610–5615. [Google Scholar] [CrossRef]
- Krechemer, F.S.; Foerster, L.A. Tuta absoluta (Lepidoptera: Gelechiidae): Thermal requirements and effect of temperature on development, survival, reproduction and longevity. Eur. J. Entomol. 2015, 112, 658–663. [Google Scholar] [CrossRef]
- Pomari, A.F.; Bueno, A.F.; Bueno, R.C.O.F.; Menezes, A.O. Telenomus remus Nixon egg parasitization of three species of Spodoptera under different temperatures. Neotrop. Entomol. 2013, 42, 399–406. [Google Scholar] [CrossRef]
- Visser, B.; Ellers, J. Lack of lipogenesis in parasitoids: A review of physiological mechanisms and evolutionary implications. J. Insect. Physiol. 2008, 54, 1315–1322. [Google Scholar] [CrossRef]
- Bueno, R.C.O.F.; Bueno, A.F.; Xavier, M.F.C.; Carvalho, M.M. Telenomus remus (Hymenoptera: Platygastridae) parasitism on eggs of Anticarsia gemmatalis (Lepidoptera: Eribidae) compared with its natural host Spodoptera frugiperda (Lepidoptera: Noctuidae). Ann. Entomol. Soc. Am. 2014, 107, 799–808. [Google Scholar] [CrossRef]
- Pak, G.A.; Oatman, E.R. Biology of Trichogramma brevicapillum. Entomol. Exp. Appl. 1982, 32, 61–67. [Google Scholar] [CrossRef]
- Bari, M.N.; Jahan, M.; Islam, K.S. Effects of temperature on the life table parameters of Trichogramma zahiri (Hymenoptera: Trichogrammatidae), an egg parasitoid of Dicladispa armigera (Chrysomelidae: Coleoptera). Environ. Entomol. 2015, 44, 368–378. [Google Scholar] [CrossRef]
- Gerling, D. The developmental biology of Telenomus remus Nixon (Hym., Scelionidae). Bull. Entomol. Res. 1972, 61, 385–388. [Google Scholar] [CrossRef]
- Chia, S.Y.; Tanga, C.M.; Khamis, F.M.; Mohamed, S.A.; Salifu, D.; Sevgan, S.; Fiaboe, K.K.M.; Niassy, S.; van Loon, J.J.; Dicke, M.; et al. Threshold temperatures and thermal requirements of black soldier fly Hermetia illucens: Implications for mass production. PLoS ONE 2018, 13, e206097. [Google Scholar] [CrossRef]
- Honek, A. Geographical variation in thermal requirements for insect development. Eur. J. Entomol. 1996, 93, 303–312. [Google Scholar]
- Corbet, S.A. Insect chemosensory responses: A chemical legacy hypothesis. Ecol. Entomol. 1985, 10, 143–153. [Google Scholar] [CrossRef]
- Queiroz, A.P.; Bueno, A.F.; Pomari-Fernande, A.; Bortolotto, O.C.; Mikami, A.Y.; Olive, L. Influence of host preference, mating, and release density on the parasitism of Telenomus remus (Nixon) (Hymenoptera, Platygastridae). Rev. Bras. Entomol. 2017, 61, 86–90. [Google Scholar] [CrossRef]
- Jaraleño-Teniente, J.; Lomeli-Flores, J.R.; Rodríguez-Leyva, E.; Bujanos-Muñiz, R.; Rodríguez-Rodríguez, S.E. Egg parasitoids survey of Spodoptera frugiperda (Smith) (Lepidoptera: Noctuidae) in maize and sorghum in central Mexico. Insects 2020, 11, 157. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | T | H | T × H | Error | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| F | df | p | F | df | p | F | df | p | ||
| Egg-first instar larvae | 3378.102 | 4 | <0.0001 | 7.579 | 1 | 0.006 | 1.550 | 4 | 0.187 | 490 |
| Second instar larvae | 218.185 | 4 | <0.0001 | 0.256 | 1 | 0.613 | 0.071 | 4 | 0.991 | 490 |
| Prepupae | 324.844 | 4 | <0.0001 | 1.153 | 1 | 0.283 | 0.222 | 4 | 0.926 | 490 |
| Pupae | 13,226.875 | 4 | <0.0001 | 27.431 | 1 | <0.0001 | 39.061 | 4 | <0.0001 | 490 |
| Generation | 43,620.894 | 4 | <0.0001 | 8.006 | 1 | 0.005 | 42.906 | 4 | <0.0001 | 490 |
| Number of parasitized eggs | 5.214 | 4 | <0.0001 | 148.405 | 1 | <0.0001 | 7.406 | 4 | <0.0001 | 274 |
| Lifespan | 68.836 | 4 | <0.0001 | 10.788 | 1 | 0.001 | 3.552 | 4 | 0.008 | 274 |
| Percentage of female | 1.935 | 4 | 0.105 | 0.080 | 1 | 0.777 | 1.611 | 4 | 0.172 | 274 |
| Emergence rate | 5.379 | 4 | <0.0001 | 55.252 | 1 | <0.0001 | 7.079 | 4 | <0.0001 | 274 |
| Stage | Host | Development Duration (h) (Mean ± SE) | F | df | p | ||||
|---|---|---|---|---|---|---|---|---|---|
| 20 °C | 23 °C | 26 °C | 29 °C | 32 °C | |||||
| Egg-first instar larvae | SF | 148.3 ± 1.4 Aa | 101.2 ± 0.9 Ab | 71.0 ± 0.9 Ac | 65.4 ± 0.7 Ad | 54.3 ± 0.8 Ae | 1530.136 | 4, 245 | <0.0001 |
| SL | 145.2 ± 1.2 Aa | 101.0 ± 0.6 Ab | 71.0 ± 0.8 Ac | 64.1 ± 0.9 Ad | 50.9 ± 0.7 Be | 1889.782 | 4, 245 | <0.0001 | |
| Second instar larvae | SF | 40.1 ± 1.0 Aa | 29.1 ± 0.8 Ab | 25.2 ± 0.8 Ac | 21.6 ± 0.7 Ad | 18.4 ± 0.7 Ae | 107.630 | 4, 245 | <0.0001 |
| SL | 39.8 ± 0.9 Aa | 28.4 ± 0.8 Ab | 25.2 ± 0.7 Ac | 21.6 ± 0.7 Ad | 18.2 ± 0.8 Ae | 110.701 | 4, 245 | <0.0001 | |
| Prepupae | SF | 36.2 ± 1.0 Aa | 26.6 ± 0.6 Ab | 23.2 ± 0.8 Ac | 12.7 ± 0.8 Ad | 12.3 ± 0.6 Ad | 166.942 | 4, 245 | <0.0001 |
| SL | 37.4 ± 1.1 Aa | 27.5 ± 0.8 Ab | 23.0 ± 0.8 Ac | 13.1 ± 0.7 Ad | 12.8 ± 0.6 Ad | 158.575 | 4, 245 | <0.0001 | |
| Pupae | SF | 257.9 ± 0.7 Ba | 189.8 ± 1.6 Ab | 138.6 ± 0.6 Ac | 101.7 ± 0.6 Bd | 96.8 ± 0.6 Ae | 5445.219 | 4, 245 | <0.0001 |
| SL | 263.8 ± 0.8 Aa | 193.6 ± 1.1 Ab | 134.1 ± 0.6 Bc | 114.7 ± 0.6 Ad | 92.4 ± 0.6 Be | 8372.082 | 4, 245 | <0.0001 | |
| Generation | SF | 482.5 ± 0.7 Ba | 346.7 ± 1.6 Ab | 258.0 ± 0.6 Ac | 201.4 ± 0.6 Bd | 181.8 ± 0.6 Ae | 18,101.270 | 4, 245 | <0.0001 |
| SL | 486.2 ± 0.8 Aa | 350.5 ± 1.1 Ab | 253.3 ± 0.6 Bc | 213.5 ± 0.6 Ad | 174.3 ± 0.6 Be | 27,294.323 | 4, 245 | <0.0001 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, W.; Li, Y.; Wang, M.; Mao, J.; Zhang, L. Evaluating the Potential of Using Spodoptera litura Eggs for Mass-Rearing Telenomus remus, a Promising Egg Parasitoid of Spodoptera frugiperda. Insects 2021, 12, 384. https://doi.org/10.3390/insects12050384
Chen W, Li Y, Wang M, Mao J, Zhang L. Evaluating the Potential of Using Spodoptera litura Eggs for Mass-Rearing Telenomus remus, a Promising Egg Parasitoid of Spodoptera frugiperda. Insects. 2021; 12(5):384. https://doi.org/10.3390/insects12050384
Chicago/Turabian StyleChen, Wanbin, Yuyan Li, Mengqing Wang, Jianjun Mao, and Lisheng Zhang. 2021. "Evaluating the Potential of Using Spodoptera litura Eggs for Mass-Rearing Telenomus remus, a Promising Egg Parasitoid of Spodoptera frugiperda" Insects 12, no. 5: 384. https://doi.org/10.3390/insects12050384
APA StyleChen, W., Li, Y., Wang, M., Mao, J., & Zhang, L. (2021). Evaluating the Potential of Using Spodoptera litura Eggs for Mass-Rearing Telenomus remus, a Promising Egg Parasitoid of Spodoptera frugiperda. Insects, 12(5), 384. https://doi.org/10.3390/insects12050384