Overwintering Larval Cold Tolerance of Sirex noctilio (Hymenoptera: Siricidae): Geographic Variation in Northeast China


 and
and
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Supercooling Point
2.3. Lower Lethal Temperature
2.4. Historical Temperature Data and Microclimatological Measurements
2.5. Data Analysis
3. Results
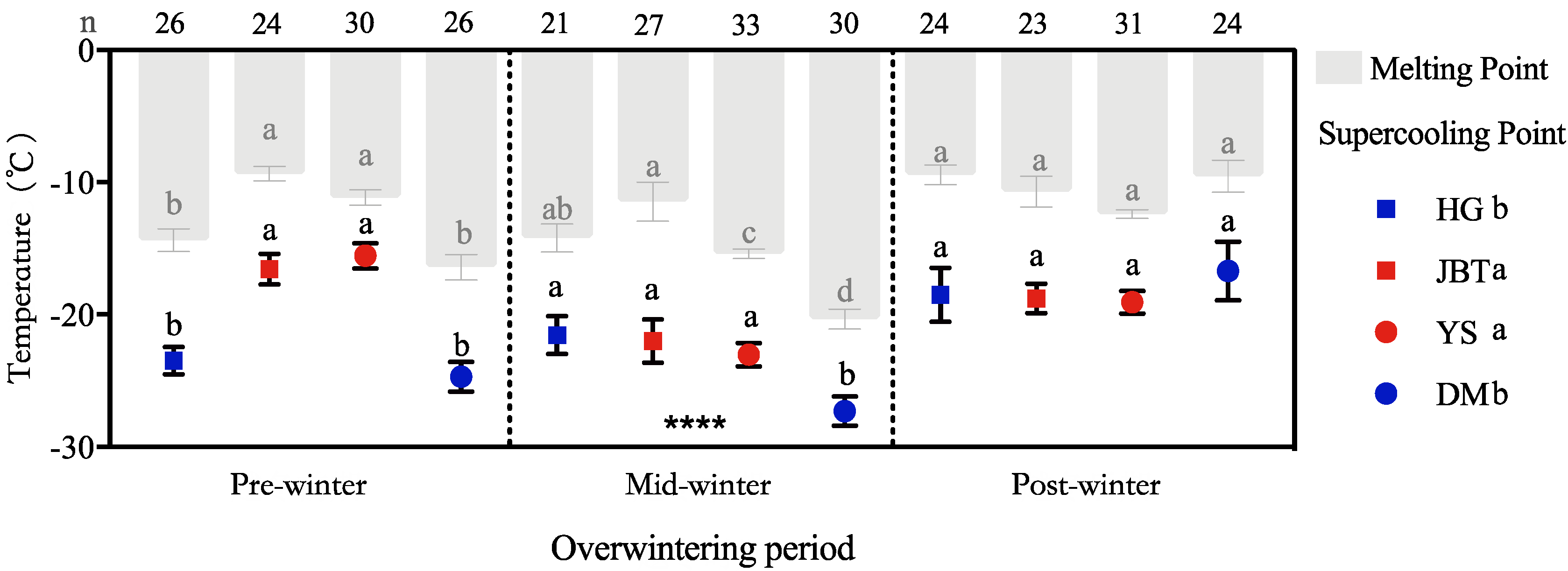
3.1. Supercooling and Melting Points of Overwintering Larvae
3.2. Diversity of SCPs from Different Geographic Populations
3.3. LLTs of Overwintering Larvae during Mid-Winter
3.4. Historical Climate Data of Sample Collection Sites
3.5. Relationship Between Historical Climate Data and the Cold Tolerance of S. noctilio Populations
4. Discussion
4.1. Cold Tolerance Strategy of S. noctilio Overwintering Larvae
4.2. In Relation to Historical Climate Data, the Cold Hardiness of Populations Varies on a Spatiotemporal Scale
4.3. Low Temperature Parameters Limiting the Distribution of S. noctilio
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Kirk, A.A. Bioclimates of Australian Pinus Radiata Areas and Sirex Noctilio Localities in the Northern Hemisphere. Aust. For. 1974, 37, 126–131. [Google Scholar] [CrossRef]
- Carnegie, A.J.; Matsuki, M.; Haugen, D.A.; Hurley, B.P.; Ahumada, R.; Klasmer, P.; Sun, J.; Iede, E.T. Predicting the potential distribution of Sirex noctilio (Hymenoptera: Siricidae), a significant exotic pest of Pinus plantations. Ann. For. Sci. 2006, 63, 119–128. [Google Scholar] [CrossRef]
- Hurley, B.P.; Slippers, B.; Croft, P.K.; Hatting, H.J.; Van Der Linde, M.; Morris, A.R.; Dyer, C.; Wingfield, M.J. Factors influencing parasitism of Sirex noctilio (Hymenoptera: Siricidae) by the nematode Deladenus siricidicola (Nematoda: Neotylenchidae) in summer rainfall areas of South Africa. Biol. Control. 2008, 45, 450–459. [Google Scholar] [CrossRef]
- Ireland, K.B.; Bulman, L.; Hoskins, A.J.; Pinkard, E.A.; Mohammed, C.; Kriticos, D.J. Estimating the potential geographical range of Sirex noctilio: Comparison with an existing model and relationship with field severity. Biol. Invasions 2018, 20, 2599–2622. [Google Scholar] [CrossRef]
- Slippers, B.; De Groot, P.; Wingfield, M.J. The Sirex Woodwasp and Its Fungal Symbiont; Springer: Dordrecht, The Netherlands; Heidelberg, Germany; London, UK; New York, NY, USA, 2012; pp. 167–247. [Google Scholar]
- Li, D.; Shi, J.; Lu, M.; Ren, L.; Zhen, C.; Luo, Y. Detection and Identification of the Invasive Sirex noctilio (Hymenoptera: Siricidae) Fungal Symbiont, Amylostereum areolatum (Russulales: Amylostereacea), in China and the Stimulating Effect of Insect Venom on Laccase Production by A. areolatum YQL03. J. Econ. Entomol. 2015, 108, 1136–1147. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Tao, J.; Ren, L.; Shi, J.; Luo, Y. Identification of Sirex noctilio (Hymenoptera: Siricidae) Using a Species-Specific Cytochrome C Oxidase Subunit I PCR Assay. J. Econ. Entomol. 2016, 109, 1424–1430. [Google Scholar] [CrossRef] [PubMed]
- Chown, S.; Nicolson, S. Insect Physiological Ecology; Oxford University Press: Oxford, UK, 2004; pp. 14–46. [Google Scholar]
- Makarieva, A.; Gorshkov, V.; Li, B. Temperature-associated upper limits to body size in terrestrial poikilotherms. Oikos 2005, 111, 425–436. [Google Scholar] [CrossRef]
- Bale, J.S.; Hayward, S.A. Insect overwintering in a changing climate. J. Exp. Biol. 2010, 213, 980–994. [Google Scholar] [CrossRef]
- Shi, P.; Wang, B.; Ayres, M.P.; Ge, F.; Zhong, L.; Li, B.L. Influence of temperature on the northern distribution limits of Scirpophaga incertulas Walker (Lepidoptera: Pyralidae) in China. J. Therm. Biol. 2012, 37, 130–137. [Google Scholar] [CrossRef]
- Madden, J.L. Egg and larval development in the woodwasp, Sirex noctilio F. Aust. J. Zool. 1981, 29, 493–506. [Google Scholar] [CrossRef]
- Li, C.; Wang, L.; Li, J.; Gao, C.; Luo, Y.; Ren, L. Thermal survival limits of larvae and adults of Sirex noctilio (Hymenoptera: Siricidae) in China. PLoS ONE 2019, 14, e0218888. [Google Scholar] [CrossRef] [PubMed]
- Stevens, G.C. The latitudinal gradient in geographical range: How so many species coexist in the tropics. Am. Nat. 1989, 133, 240–256. [Google Scholar] [CrossRef]
- Zachariassen, K.E. Physiology of cold tolerance in insects. Physiol. Rev. 1985, 65, 799–832. [Google Scholar] [CrossRef] [PubMed]
- Asahina, E. Frost Resistance in Insects. Adv. Insect Physiol. 1969, 6, 1–49. [Google Scholar]
- Richard, E.L.J. A primer on insect cold-tolerance. In Low Temperature Biology of Insects; David, L.D., Richard, E.L.J., Eds.; Cambridge University Press: Cambridge, UK, 2010; 404p, ISBN 978-0-521-88635-2. [Google Scholar]
- Sinclair, B.J.; Coello Alvarado, L.E.; Ferguson, L.V. An invitation to measure insect cold tolerance: Methods, approaches, and workflow. J. Therm. Biol. 2015, 53, 180–197. [Google Scholar] [CrossRef]
- Andersen, J.L.; Manenti, T.; Sørensen, J.G.; Macmillan, H.A.; Loeschcke, V.; Overgaard, J. How to assess Drosophila cold tolerance: Chill coma temperate and lower lethal temperature are the best predictors of cold distribution limits. Funct. Ecol. 2015, 29, 55–65. [Google Scholar] [CrossRef]
- Feng, Y.; Xu, L.; Tian, B.; Tao, J.; Wang, J.; Zong, S. Cold hardiness of asian longhorned beetle (coleoptera: Cerambycidae) larvae in different populations. Environ. Entomol. 2014, 5, 1419–1426. [Google Scholar] [CrossRef]
- Feng, Y.; Xu, L.; Li, W.; Xu, Z.; Cao, M.; Wang, J.; Tao, J.; Zong, S. Seasonal changes in supercooling capacity and major cryoprotectants of overwintering asian longhorned beetle (anoplophora glabripennis) larvae. Agric. For. Entomol. 2016, 18, 302–312. [Google Scholar] [CrossRef]
- Javal, M.; Roques, A.; Roux, G.; Laparie, M. Respiration-based monitoring of metabolic rate following cold-exposure in two invasive anoplophora species depending on acclimation regime. Comp. Biochem. Phys. A 2018, 216, 20–27. [Google Scholar]
- Li, J.; Shi, J.; Xue, Y.; Mao, H.; Luo, Y. Major physiological adjustments in freezing-tolerant grey tiger longicorn beetle (Xylotrechus rusticus) during overwintering period. J. For. Res. 2014, 25, 653–659. [Google Scholar] [CrossRef]
- Ma, R.; Hao, S.; Kong, W.; Sun, J.; Kang, L. Cold hardiness as a factor for assessing the potential distribution of the Japanese pine sawyer Monochamus alternatus (Coleoptera: Cerambycidae) in China. Ann. For. Sci. 2006, 63, 449–456. [Google Scholar] [CrossRef][Green Version]
- EPPO. PQR Database. Paris, France: European and Mediterranean Plant Protection Organization. 2014. Available online: http://www.eppo.int/DATABASES/pqr/pqr.html (accessed on 6 June 2020).
- David, L.D.; Richard, E.L.J. Cryobiology. Book Reviews: Insects at Low Temperature. Science 1991, 252, 1326–1327. [Google Scholar]
- Sunday, J.M.; Bates, A.E.; Dulvy, N.K. Global analysis of thermal tolerance and latitude in ectotherms. Proc. Biol. Sci. 2011, 278, 1823–1830. [Google Scholar] [CrossRef] [PubMed]
- Sheldon, K.S.; Tewksbury, J.J. The impact of seasonality in temperature on thermal tolerance and elevational range size. Ecology 2014, 95, 2134–2143. [Google Scholar] [CrossRef] [PubMed]
- Sunday, J.; Bennett, J.M.; Calosi, P.; Clusella-Trullas, S.; Gravel, S.; Hargreaves, A.L.; Leiva, F.P.; Verberk, W.C.E.P.; Olalla-Tárraga, M.Á.; Morales-Castilla, I. Thermal tolerance patterns across latitude and elevation. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2019, 374, 20190036. [Google Scholar] [CrossRef] [PubMed]
- Käfer, H.; Kovac, H.; Simov, N.; Battisti, A.; Erregger, B.; Schmidt, A.K.D.; Stabentheiner, A. Temperature tolerance and thermal environment of European seed bugs. Insects 2020, 11, 197. [Google Scholar]
- Oyen, K.J.; Giri, S.; Dillon, M.E. Altitudinal variation in bumble bee (Bombus) critical thermal limits. J. Therm. Biol. 2016, 59, 52–57. [Google Scholar] [CrossRef]
- Taylor, K.L. The Sirex woodwasp: Ecology and control of an introduced forest insect (pine pest). Ecol. Pests 1981, 12, 231–248. [Google Scholar]
- Sinclair, B.J.; Vernon, P.; Klok, C.J.; Chown, S.L. Insects at low temperatures: An ecological perspective. Trends Ecol. Evol. 2003, 18, 257–262. [Google Scholar] [CrossRef]
- Storey, K.B.; Storey, J.M. Freeze tolerance: Constraining forces, adaptive mechanisms. Revue Can. Zool. 1988, 66, 1122–1127. [Google Scholar]
- Delisle, J.; Labrecque, A.; Royer, L.; Bernier-Cardou, M.; Bauce, É.; Charest, M.; Larrivée, S. Impact of short-term exposure to low subzero temperatures on egg hatch in the hemlock looper. Lambdina Fiscellaria. Entomol. Exp. Appl. 2013, 149, 206–218. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Annual Average Temp. (°C) | Extreme Low Temp. (°C) | Coldest Month Average Temp. (°C) | Coldest Month Average Low Temp. (°C) | Coldest Month Average High Temp. (°C) |
|---|---|---|---|---|---|
| HG | 3.8 | −31.9 | −16.6 | −20 | −12.1 |
| JBT | 6.8 | −32.7 | −12.9 | −17.8 | −6.7 |
| YS | 5 | −36.7 | −17.1 | −22 | −11.6 |
| DM | 4.6 | −36.8 | −18 | −22.4 | −12.6 |
| Exposure Time (h) | Sample Size | Slope + SE | LLT50 (°C) | 50% Fiducial Limit | LLT99 (°C) | 99% Fiducial Limit | Chi-Square | ||
|---|---|---|---|---|---|---|---|---|---|
| Lower | Upper | Lower | Upper | ||||||
| 2 | 77 | −0.30 + 0.09 | −22.09 | −24.52 | −19.75 | −29.77 | −39.79 | −26.56 | 0.67 |
| 4 | 79 | −0.41 + 0.14 | −19.23 | −21.45 | −16.94 | −24.84 | −35.79 | −22.25 | 0.56 |
| 24 | 80 | −0.21 + 0.05 | −10.00 | −12.49 | −7.51 | −21.07 | −30.17 | −17.21 | 0.83 |
| 48 | 81 | −0.16 + 0.04 | −3.53 | −6.25 | −0.41 | −17.89 | −28.57 | −13.28 | 0.92 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, C.; Pei, J.; Li, J.; Liu, X.; Ren, L.; Luo, Y. Overwintering Larval Cold Tolerance of Sirex noctilio (Hymenoptera: Siricidae): Geographic Variation in Northeast China. Insects 2021, 12, 116. https://doi.org/10.3390/insects12020116
Li C, Pei J, Li J, Liu X, Ren L, Luo Y. Overwintering Larval Cold Tolerance of Sirex noctilio (Hymenoptera: Siricidae): Geographic Variation in Northeast China. Insects. 2021; 12(2):116. https://doi.org/10.3390/insects12020116
Chicago/Turabian StyleLi, Chengcheng, Jiahe Pei, Jiale Li, Xiaobo Liu, Lili Ren, and Youqing Luo. 2021. "Overwintering Larval Cold Tolerance of Sirex noctilio (Hymenoptera: Siricidae): Geographic Variation in Northeast China" Insects 12, no. 2: 116. https://doi.org/10.3390/insects12020116
APA StyleLi, C., Pei, J., Li, J., Liu, X., Ren, L., & Luo, Y. (2021). Overwintering Larval Cold Tolerance of Sirex noctilio (Hymenoptera: Siricidae): Geographic Variation in Northeast China. Insects, 12(2), 116. https://doi.org/10.3390/insects12020116