
Swallowtail Butterflies Use Multiple Visual Cues to Select Oviposition Sites

Abstract
Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Animals
2.2. Behavioral Experiment
2.3. Experimental Setup
2.4. Procedure of Behavioral Experiment
2.5. Measurement of Visual Features
2.6. Statistical Analysis
3. Results
3.1. Oviposition Behavior and Leaf Selection
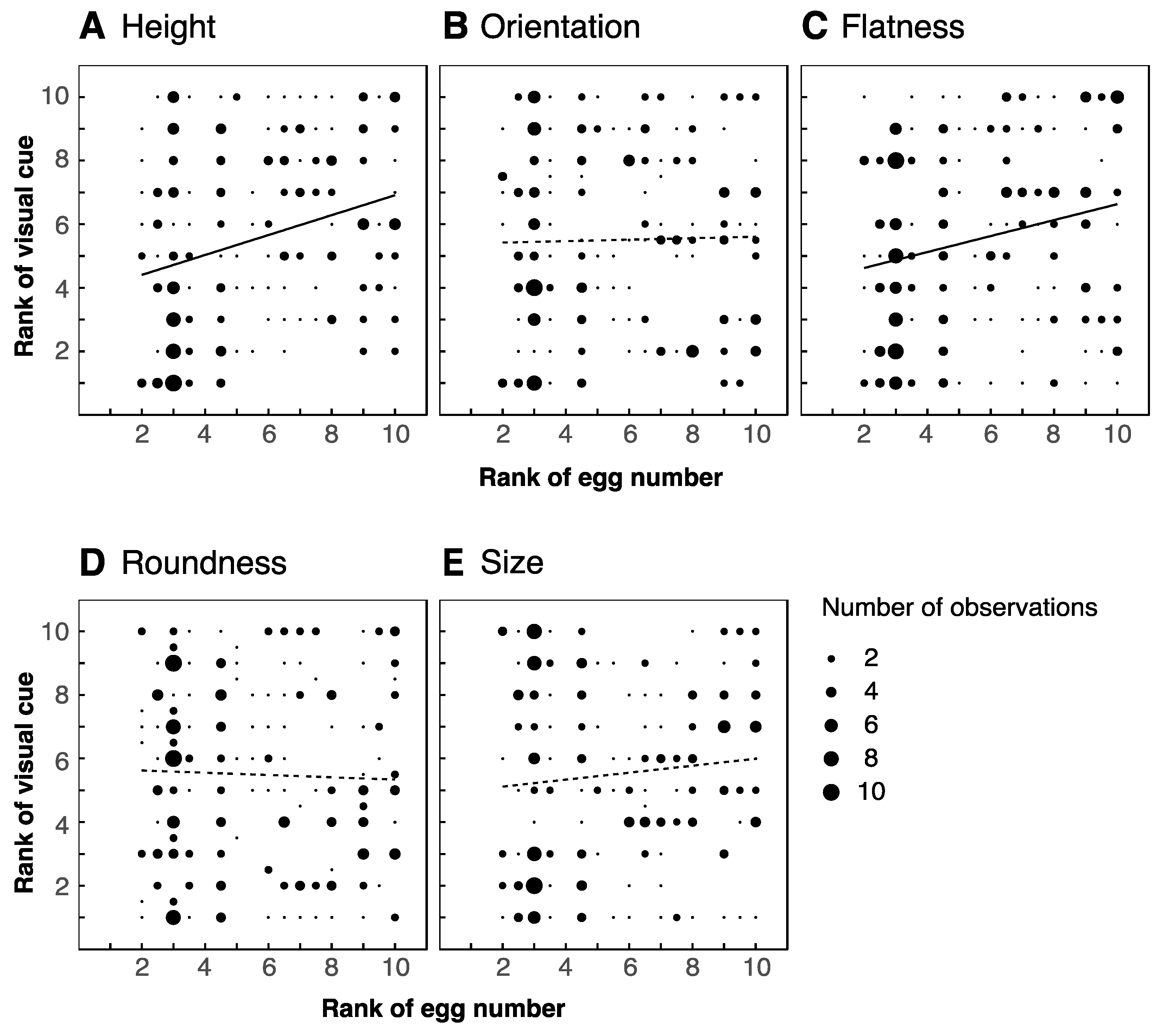
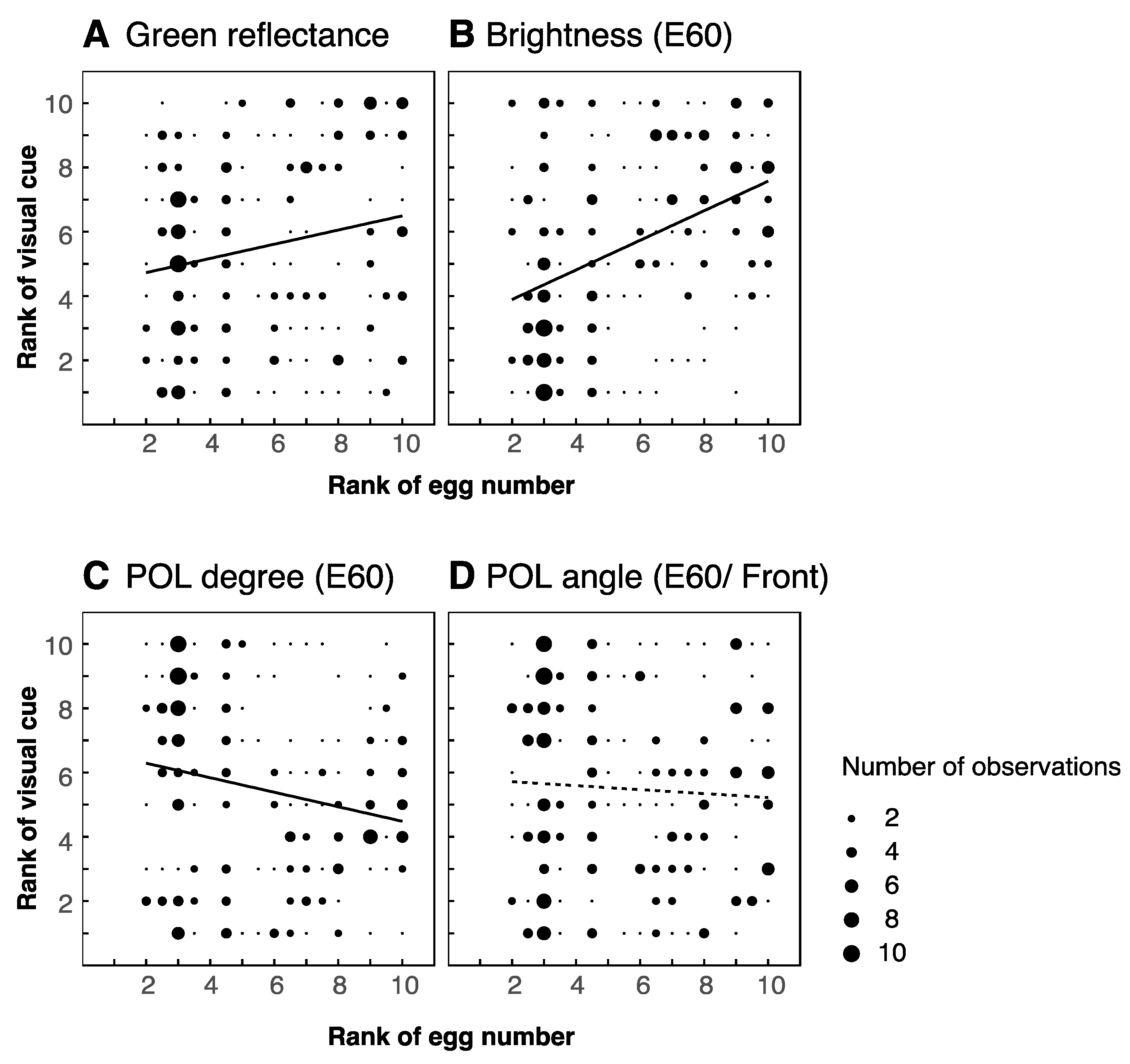
3.2. Visual Features of Leaves
4. Discussion
4.1. Leaf Selection for Oviposition and Visual Cues
4.2. Correlations between Number of Eggs and Visual Cues
4.3. Ecological Perspectives
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Menzel, R.; Backhaus, W. Color vision in honey bees: Phenomena and physiological mechanisms. In Facets of Vision; Stavenga, D.G., Hardie, R.C., Eds.; Springer: Berlin/Heidelberg, Germany, 1989. [Google Scholar]
- Chittka, L.; Raine, N.E. Recognition of flowers by pollinators. Curr. Opin. Plant Biol. 2006, 9, 428–435. [Google Scholar] [CrossRef]
- Hempel de Ibarra, N.; Giurfa, M.; Vorobyev, M. Discrimination of coloured patterns by honeybees through chromatic and achromatic cues. J. Comp. Physiol. A 2002, 188, 503–512. [Google Scholar] [CrossRef] [PubMed]
- Giurfa, M. Cognitive neuroethology: Dissecting non-elemental learning in a honeybee brain. Curr. Opin. Neurobiol. 2003, 13, 726–735. [Google Scholar] [CrossRef] [PubMed]
- Hempel de Ibarra, N.; Vorobyev, M. Flower patterns are adapted for detection by bees. J. Comp. Physiol. A 2009, 195, 319–323. [Google Scholar] [CrossRef] [PubMed]
- Weiss, M.R. Associative colour learning u a nymphalid butterfly. Ecol. Entomol. 1995, 20, 298–301. [Google Scholar] [CrossRef]
- Weiss, M.R. Innate colour preferences and flexible colour learning in the pipivine swallowtail. Anim. Behav. 1997, 53, 1043–1052. [Google Scholar] [CrossRef]
- Kelber, A. Ovipositing butterflies use a red receptor to see green. J. Exp. Biol. 1999, 202 Pt 19, 2619–2630. [Google Scholar] [CrossRef]
- Ômura, H.; Honda, K. Priority of color over scent during flower visitation by adult Vanessa indica butterflies. Oecologia 2005, 142, 588–596. [Google Scholar] [CrossRef]
- Zaccardi, G.; Kelber, A.; Sison-Mangus, M.P.; Briscoe, A.D. Color discrimination in the red range with only one long-wavelength sensitive opsin. J. Exp. Biol. 2006, 209, 1944–1955. [Google Scholar] [CrossRef] [PubMed]
- Kandori, I.; Yamaki, T.; Okuyama, S.; Sakamoto, N.; Yokoi, T. Interspecific and intersexual learning rate differences in four butterfly species. J. Exp. Biol. 2009, 212, 3810–3816. [Google Scholar] [CrossRef]
- Blackiston, D.; Briscoe, A.D.; Weiss, M.R. Color vision and learning in the monarch butterfly, Danaus plexippus (Nymphalidae). J. Exp. Biol. 2011, 214, 509–520. [Google Scholar] [CrossRef]
- Kinoshita, M.; Arikawa, K. Color and polarization vision in foraging Papilio. J. Comp. Physiol. A 2014, 200, 513–526. [Google Scholar] [CrossRef]
- Arikawa, K. Spectral organization of the eye of a butterfly, Papilio. J. Comp. Physiol. A 2003, 189, 791–800. [Google Scholar] [CrossRef]
- Stewart, F.J.; Kinoshita, M.; Arikawa, K. The roles of visual parallax and edge attraction in the foraging behavior of the butterfly Papilio xuthus. J. Exp. Biol. 2015, 218, 1725–1732. [Google Scholar] [CrossRef] [PubMed]
- Stewart, F.J.; Kinoshita, M.; Arikawa, K. Monopolatic motion vision in the butterfly Papilio xuthus. J. Exp. Biol. 2019, 222, jeb191957. [Google Scholar] [CrossRef] [PubMed]
- Nishida, R.; Ohsugi, T.; Kokubo, S.; Fukami, H. Oviposition stimulants of a Citrus-feeding swallowtail butterfly, Papilio xuthus L. Experientia 1987, 43, 342–344. [Google Scholar] [CrossRef]
- Renwick, J.A.A. Chemical ecology of oviposition in phytophagous insects. Experientia 1989, 45, 223–228. [Google Scholar] [CrossRef]
- Itoh, Y.; Okumura, O.; Fujii, T.; Ishikawa, H.; Ômura, H. Effects of mating on host selection by female small white butterflies Pieris rapae (Lepidoptera: Pieridae). J. Comp. Physiol. A 2018, 204, 245–255. [Google Scholar] [CrossRef]
- Abe, H.; Teramoto, Y.; Ichinose, T. Relationship between host plant ranges of the three Papilionid butterflies and ovipsition-inducing contact chemicals in their host plants. Appl. Entomol. Zool. 1981, 16, 493–496. [Google Scholar] [CrossRef][Green Version]
- Chen, Y.; Lin, L.; Wang, C.; Yeh, C.; Hwang, S. Response of two Pieris (Lepidoptera: Pieridae) species to fertilization of a host plant. Zool. Stud. 2004, 43, 778–786. [Google Scholar]
- Jaumann, S.; Snell-Rood, E.C. Trade-offs between fecundity and choosiness in ovipositing butterflies. Anim. Behav. 2017, 123, 433–440. [Google Scholar] [CrossRef]
- Reudler-Talsma, J.H.; Biere, A.; Harvey, J.A.; van Nouhuys, S. Oviposition cues for a specialist butterfly-plant chemistry and size. J. Chem. Ecol. 2008, 34, 1202–1212. [Google Scholar] [CrossRef] [PubMed]
- Ilse, D. New observations on responses to colours in egg laying butterflies. Nature 1937, 544–545. [Google Scholar] [CrossRef]
- Kolb, G.; Scherer, C. Experiments on wavelength specific behavior of Pieris brassicae L. During drumming and egg-laying. J. Comp. Physiol. A 1982, 149, 325–332. [Google Scholar] [CrossRef]
- Scherer, C.; Kolb, G. Behavioral experiments on the visual processing of color stimuli in Pieris brassicae L. (Lepidoptera). J. Comp. Physiol. A 1987, 160, 645–656. [Google Scholar] [CrossRef]
- Kelber, A. Why ‘false’ colours are seen by butterflies. Nature 1999, 402, 251. [Google Scholar] [CrossRef]
- Kelber, A.; Thunell, C.; Arikawa, K. Polarisation-dependent colour vision in Papilio butterflies. J. Exp. Biol. 2001, 204, 2469–2480. [Google Scholar] [CrossRef]
- Siegel, S.; Castellan, N.J. Nonparametric Statistics for the Behavioral Scuence; MacGraw-Hill: New York, NY, USA, 1988; ISBN 10:70573573. [Google Scholar]
- Arikawa, K.; Takagi, N. Genital photoreceptors have crucial role in oviposition in Japanese yellow swallowtail butterfly, Papilio xuthus. Zool. Sci. 2001, 18, 175–179. [Google Scholar] [CrossRef]
- Weiss, M.R.; Papaj, D.R. Colour learning in two behavioural contexts: How much can a butterfly keep in mind? Anim. Behav. 2003, 65, 425–434. [Google Scholar] [CrossRef]
- Ichinose, T.; Honda, H. Ovipositional behavior of Papilio protenor demetrius CRAMER and the factors involved in its host plants. Appl. Ent. Zool. 1978, 13, 103–114. [Google Scholar] [CrossRef]
- Tedore, C.; Nilsson, N. Avian UV vision enhances leaf surface contrasts in forest environments. Nat. Commun. 2019, 10, 238. [Google Scholar] [CrossRef]
- Labhart, T. Can invertebrates see the e-vector of polarization as a separate modality of light? J. Exp. Biol. 2016, 219, 3844–3856. [Google Scholar] [CrossRef] [PubMed]
- Marshall, N.J.; Powell, S.B.; Cronin, T.W.; Caldwell, R.L.; Johnsen, S.; Gruev, V.; Chiou, T.S.; Roberts, N.W.; How, M.J. Polarisation signals: A new currency for communication. J. Exp. Biol. 2019, 222, jeb134213. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Morphology | |||||
|---|---|---|---|---|---|
| Leaf Type | Height (cm) | Orientation (−90° < O < +90°) | Flatness (0 < F < 1) | Roundness (0 < R < 1) | Size (cm2) |
| Favored | 86.10 ± 1.11 | 2.80 ± 4.47 | 0.93 ± 0.01 | 0.50 ± 0.02 | 49.58 ± 3.37 |
| Unfavored | 82.73 ± 2.10 | 2.52 ± 5.40 | 0.90 ± 0.01 | 0.46 ± 0.02 | 47.57 ± 4.40 |
| p value | 0.37 | 0.96 | 0.19 | 0.37 | 0.59 |
| Reflected Light | |||||
| Leaf Type | Rel. Green Reflectance (0 < G < 1) | Brightness (E60) (0 < B < 1) | POL Angle (E60/Front) (Degree) | POL Degree (E60) (0 < D < 1) | |
| Favored | 0.74 ± 0.03 | 0.54 ± 0.03 | 104.30 ± 11.15 | 0.10 ± 0.01 | |
| Unfavored | 0.72 ± 0.02 | 0.50 ± 0.02 | 91.32 ± 12.93 | 0.128 ± 0.01 | |
| p value | 0.73 | 0.30 | 0.19 | 0.10 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nagaya, H.; Stewart, F.J.; Kinoshita, M. Swallowtail Butterflies Use Multiple Visual Cues to Select Oviposition Sites. Insects 2021, 12, 1047. https://doi.org/10.3390/insects12111047
Nagaya H, Stewart FJ, Kinoshita M. Swallowtail Butterflies Use Multiple Visual Cues to Select Oviposition Sites. Insects. 2021; 12(11):1047. https://doi.org/10.3390/insects12111047
Chicago/Turabian StyleNagaya, Hiromi, Finlay J. Stewart, and Michiyo Kinoshita. 2021. "Swallowtail Butterflies Use Multiple Visual Cues to Select Oviposition Sites" Insects 12, no. 11: 1047. https://doi.org/10.3390/insects12111047
APA StyleNagaya, H., Stewart, F. J., & Kinoshita, M. (2021). Swallowtail Butterflies Use Multiple Visual Cues to Select Oviposition Sites. Insects, 12(11), 1047. https://doi.org/10.3390/insects12111047