Aquatic Insects Are Dramatically Underrepresented in Genomic Research


{kind=link}
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
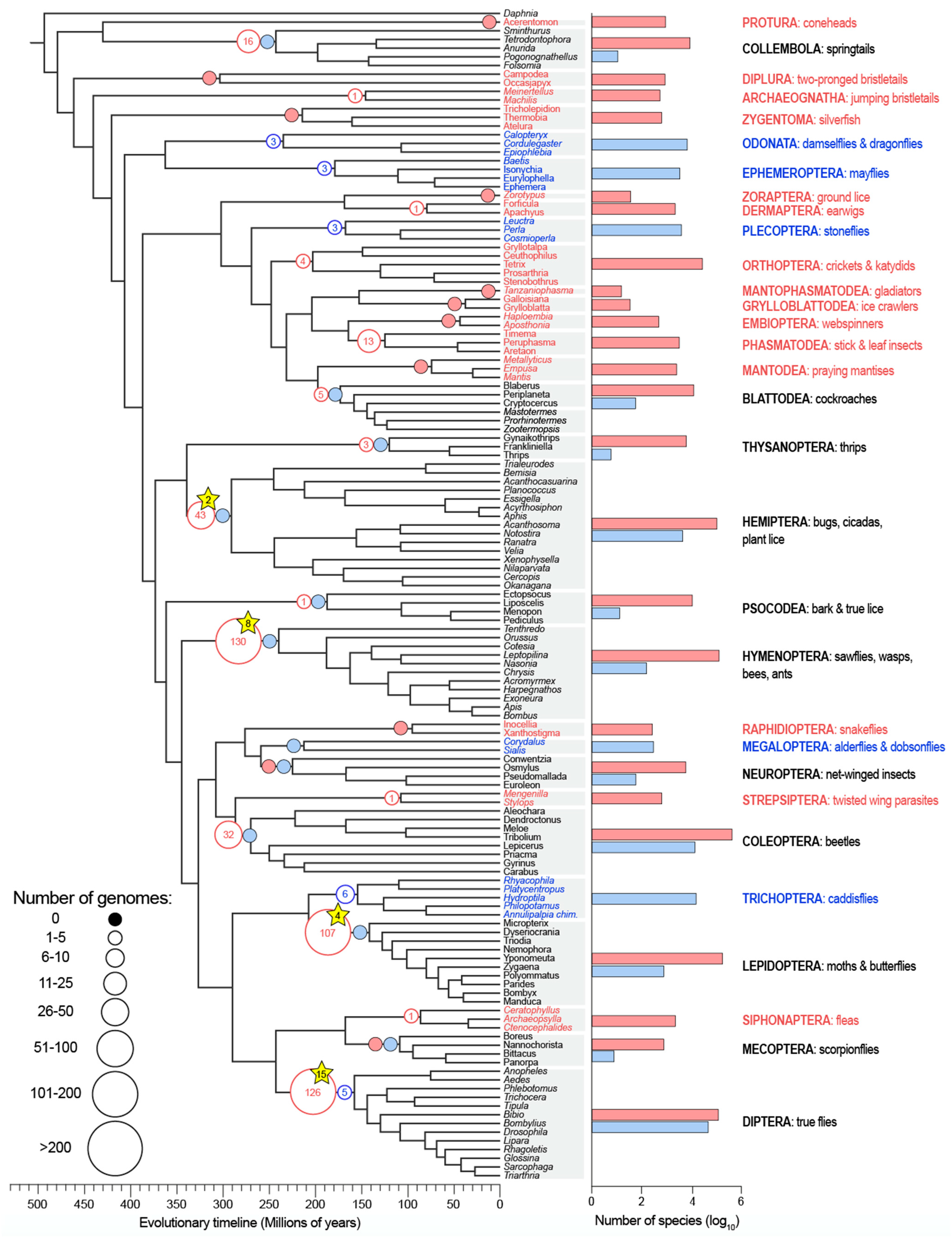
3. Results and Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zhang, Z.-Q. Animal Biodiversity: An Outline of Higher-Level Classification and Survey of Taxonomic Richness; Magnolia Press: Auckland, New Zealand, 2011. [Google Scholar]
- Dijkstra, K.-D.B.; Monaghan, M.T.; Pauls, S.U. Freshwater biodiversity and aquatic insect diversification. Ann. Rev. Entomol. 2014, 59. [Google Scholar] [CrossRef] [PubMed]
- Hotaling, S.; Kelley, J.L. The rising tide of high-quality genomic resources. Mol. Ecol. Resour. 2020, 19, 567–569. [Google Scholar] [CrossRef] [PubMed]
- i5K Consortium. The i5K Initiative: Advancing arthropod genomics for knowledge, human health, agriculture, and the environment. J. Hered. 2013, 104, 595–600. [Google Scholar] [CrossRef] [PubMed]
- May, M.L. Odonata: Who they are and what they have done for us lately: Classification and ecosystem services of dragonflies. Insects 2019, 10, 62. [Google Scholar] [CrossRef]
- Epanchin, P.N.; Knapp, R.A.; Lawler, S.P. Nonnative trout impact an alpine-nesting bird by altering aquatic-insect subsidies. Ecology 2010, 91, 2406–2415. [Google Scholar] [CrossRef]
- Miguel, T.B.; Oliveira-Junior, J.M.B.; Ligeiro, R.; Juen, L. Odonata (Insecta) as a tool for the biomonitoring of environmental quality. Ecol. Indic. 2017, 81, 555–566. [Google Scholar] [CrossRef]
- Mauvisseau, Q.; Davy-Bowker, J.; Bulling, M.; Brys, R.; Neyrinck, S.; Troth, C.; Sweet, M. Combining ddPCR and environmental DNA to improve detection capabilities of a critically endangered freshwater invertebrate. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef]
- Balian, E.; Segers, H.; Lévèque, C.; Martens, K. The Freshwater Animal Diversity Assessment: An overview of the results. Hydrobiologia 2008, 595, 627–637. [Google Scholar] [CrossRef]
- Toews, D.P.; Brelsford, A. The biogeography of mitochondrial and nuclear discordance in animals. Mol. Ecol. 2012, 21, 3907–3930. [Google Scholar] [CrossRef]
- Rueda, L.M. Global Diversity of Mosquitoes (Insecta: Diptera: Culicidae) in Freshwater. In Freshwater Animal Diversity Assessment; Springer: Berlin, Germany, 2007; pp. 477–487. [Google Scholar]
- Ha, Y.R.; Ryu, J.; Yeom, E.; Lee, S.J. Comparison of the tracheal systems of Anopheles sinensis and Aedes togoi larvae using synchrotron X-ray microscopic computed tomography (respiratory system of mosquito larvae using SR-µCT). Microsc. Res. Tech. 2017, 80, 985–993. [Google Scholar] [CrossRef]
- Rhie, A.; McCarthy, S.A.; Fedrigo, O.; Damas, J.; Formenti, G.; Koren, S.; Uliano-Silva, M.; Chow, W.; Fungtammasan, A.; Gedman, G.L. Towards complete and error-free genome assemblies of all vertebrate species. BioRxiv 2020. [Google Scholar] [CrossRef]
- Simão, F.A.; Waterhouse, R.M.; Ioannidis, P.; Kriventseva, E.V.; Zdobnov, E.M. BUSCO: Assessing genome assembly and annotation completeness with single-copy orthologs. Bioinformatics 2015, 31, 3210–3212. [Google Scholar] [CrossRef] [PubMed]
- Heckenhauer, J.; Frandsen, P.B.; Gupta, D.K.; Paule, J.; Prost, S.; Schell, T.; Schneider, J.V.; Stewart, R.J.; Pauls, S.U. Annotated draft genomes of two caddisfly species Plectrocnemia conspersa CURTIS and Hydropsyche tenuis NAVAS (Insecta: Trichoptera). Genome Biol. Evol. 2019, 11, 3445–3451. [Google Scholar] [CrossRef] [PubMed]
- Stork, N.E. How many species of insects and other terrestrial arthropods are there on Earth? Ann. Rev. Entomol. 2018, 63, 31–45. [Google Scholar] [CrossRef]
- Bennett, A.M. Global diversity of hymenopterans (Hymenoptera; Insecta) in freshwater. In Freshwater Animal Diversity Assessment; Springer: Berlin, Germany, 2007; pp. 529–534. [Google Scholar]
- Del-Claro, K.; Guillermo, R. Aquatic Insects: Behavior and Ecology; Springer: Berlin, Germany, 2019; p. 438. [Google Scholar]
- Mey, W.; Speidel, W. Global Diversity of Butterflies (Lepidotera) in Freshwater. In Freshwater Animal Diversity Assessment; Springer: Berlin, Germany, 2007; pp. 521–528. [Google Scholar]
- Mound, L.A. The aquatic thrips Organothrips indicus Bhatti (Thysanoptera: Thripidae) in Queensland, and a new species, O. wrighti, from tropical Australia. Aust. J. Entomol. 2000, 39, 10–14. [Google Scholar] [CrossRef]
- Misof, B.; Liu, S.; Meusemann, K.; Peters, R.S.; Donath, A.; Mayer, C.; Frandsen, P.B.; Ware, J.; Flouri, T.; Beutel, R.G.; et al. Phylogenomics resolves the timing and pattern of insect evolution. Science 2014, 346, 763–767. [Google Scholar] [CrossRef]
- Faddeeva-Vakhrusheva, A.; Kraaijeveld, K.; Derks, M.F.; Anvar, S.Y.; Agamennone, V.; Suring, W.; Kampfraath, A.A.; Ellers, J.; le Ngoc, G.; van Gestel, C.A. Coping with living in the soil: The genome of the parthenogenetic springtail Folsomia candida. BMC Genom. 2017, 18, 493. [Google Scholar] [CrossRef]
- Kingan, S.B.; Heaton, H.; Cudini, J.; Lambert, C.C.; Baybayan, P.; Galvin, B.D.; Durbin, R.; Korlach, J.; Lawniczak, M.K. A high-quality de novo genome assembly from a single mosquito using PacBio sequencing. Genes 2019, 10, 62. [Google Scholar] [CrossRef]
- Hellsten, U.; Harland, R.M.; Gilchrist, M.J.; Hendrix, D.; Jurka, J.; Kapitonov, V.; Ovcharenko, I.; Putnam, N.H.; Shu, S.; Taher, L. The genome of the Western clawed frog Xenopus tropicalis. Science 2010, 328, 633–636. [Google Scholar] [CrossRef]
- Nowoshilow, S.; Schloissnig, S.; Fei, J.-F.; Dahl, A.; Pang, A.W.; Pippel, M.; Winkler, S.; Hastie, A.R.; Young, G.; Roscito, J.G. The axolotl genome and the evolution of key tissue formation regulators. Nature 2018, 554, 50–55. [Google Scholar] [CrossRef]
- Gregory, T.R. Animal Genome Size Database. 2020. Available online: http://www.genomesize.com (accessed on 13 July 2020).
- Macdonald, H.C.; Cunha, L.; Bruford, M.W. Development of genomic resources for four potential environmental bioindicator species: Isoperla grammatica, Amphinemura sulcicollis, Oniscus asellus and Baetis rhodani. BioRxiv 2016. [Google Scholar] [CrossRef]
- Luo, S.; Tang, M.; Frandsen, P.B.; Stewart, R.J.; Zhou, X. The genome of an underwater architect, the caddisfly Stenopsyche tienmushanensis Hwang (Insecta: Trichoptera). GigaScience 2018, 7, giy143. [Google Scholar] [CrossRef] [PubMed]
- Hotaling, S.; Kelley, J.L.; Weisrock, D.W. Nuclear and mitochondrial genomic resources for the meltwater stonefly (Plecoptera: Nemouridae), Lednia tumana (Ricker, 1952). Aquat. Insects 2019, 40, 362–369. [Google Scholar] [CrossRef]
- Almudi, I.; Vizueta, J.; Wyatt, C.D.; de Mendoza, A.; Marlétaz, F.; Firbas, P.N.; Feuda, R.; Masiero, G.; Medina, P.; Alcaina-Caro, A. Genomic adaptations to aquatic and aerial life in mayflies and the origin of insect wings. Nat. Commun. 2020, 11, 1–11. [Google Scholar] [CrossRef]
- Sayers, E.W.; Cavanaugh, M.; Clark, K.; Ostell, J.; Pruitt, K.D.; Karsch-Mizrachi, I. GenBank. Nucleic Acids Res. 2020, 48, D84–D86. [Google Scholar]

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hotaling, S.; Kelley, J.L.; Frandsen, P.B. Aquatic Insects Are Dramatically Underrepresented in Genomic Research. Insects 2020, 11, 601. https://doi.org/10.3390/insects11090601
Hotaling S, Kelley JL, Frandsen PB. Aquatic Insects Are Dramatically Underrepresented in Genomic Research. Insects. 2020; 11(9):601. https://doi.org/10.3390/insects11090601
Chicago/Turabian StyleHotaling, Scott, Joanna L. Kelley, and Paul B. Frandsen. 2020. "Aquatic Insects Are Dramatically Underrepresented in Genomic Research" Insects 11, no. 9: 601. https://doi.org/10.3390/insects11090601
APA StyleHotaling, S., Kelley, J. L., & Frandsen, P. B. (2020). Aquatic Insects Are Dramatically Underrepresented in Genomic Research. Insects, 11(9), 601. https://doi.org/10.3390/insects11090601