Wolbachia-Mitochondrial DNA Associations in Transitional Populations of Rhagoletis cerasi




{kind=link}
Simple Summary
Abstract
1. Introduction
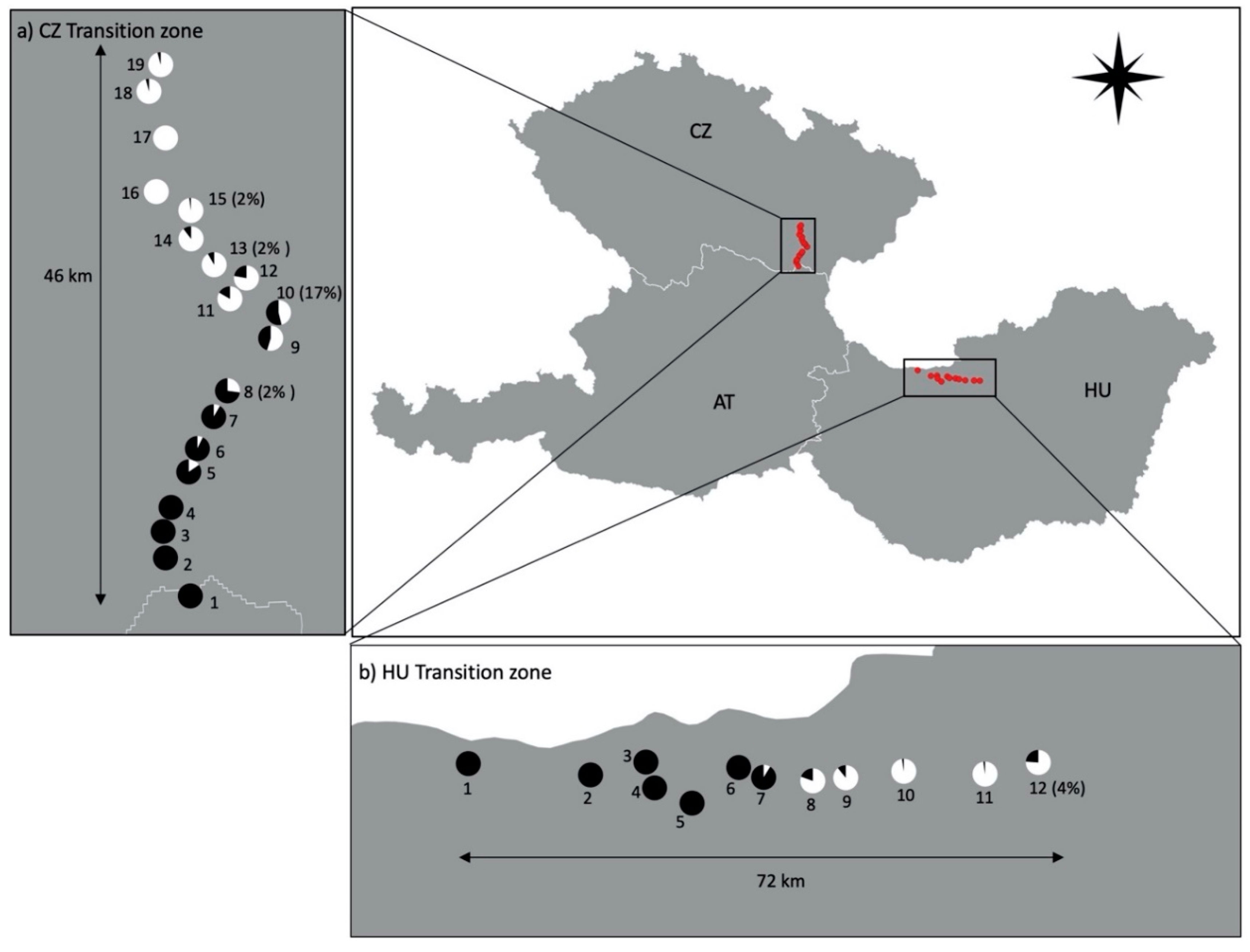
2. Materials and Methods
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Weinert, L.A.; Araujo-Jnr, E.V.; Ahmed, M.Z.; Welch, J.J. The incidence of bacterial endosymbionts in terrestrial arthropods. Proc. R. Soc. B Biol. Sci. 2015, 282, 20150249. [Google Scholar] [CrossRef] [PubMed]
- Werren, J.H.; Windsor, D.M. Wolbachia infection frequencies in insects: Evidence of a global equilibrium? Proc. R. Soc. B Biol. Sci. 2000, 267, 1277–1285. [Google Scholar] [CrossRef] [PubMed]
- Werren, J.H.; Baldo, L.; Clark, M.E. Wolbachia: Master manipulators of invertebrate biology. Nat. Rev. Microbiol. 2008, 6, 741–751. [Google Scholar] [CrossRef] [PubMed]
- Nikolouli, K.; Colinet, H.; Renault, D.; Enriquez, T.; Mouton, L.; Gibert, P.; Sassu, F.; Cáceres, C.; Stauffer, C.; Pereira, R.; et al. Sterile insect technique and Wolbachia symbiosis as potential tools for the control of the invasive species Drosophila suzukii. J. Pest Sci. 2018, 91, 489–503. [Google Scholar] [CrossRef]
- Bishop, C.; Parry, R.; Asgari, S. Effect of Wolbachia wAlbB on a positive-sense RNA negev-like virus: A novel virus persistently infecting Aedes albopictus mosquitoes and cells. J. Gen. Virol. 2020, 101, 216–225. [Google Scholar] [CrossRef]
- Iturbe-Ormaetxe, I.; Walker, T.; O’Neill, S.L. Wolbachia and the biological control of mosquito-borne disease. EMBO Rep. 2011, 12, 508–518. [Google Scholar] [CrossRef]
- Schmidt, T.L.; Barton, N.H.; Rasic, G.; Turley, A.P.; Montgomery, B.L.; Iturbe-Ormaetxe, I.; Cook, P.E.; Ryan, P.A.; Ritchie, S.A.; Hoffmann, A.A.; et al. Local introduction and heterogeneous spatial spread of dengue-suppressing Wolbachia through an urban population of Aedes aegypti. PLoS Biol. 2017, 15, e2001894. [Google Scholar] [CrossRef]
- Walker, T.; Moreira, L.A. Can Wolbachia be used to control Malaria? Memorias do Instituto Oswaldo Cruz 2011, 106, 212–217. [Google Scholar] [CrossRef]
- Zabalou, S.; Riegler, M.; Theodarakopoulou, M.; Stauffer, C.; Savakis, C.; Bourtzis, K. Wolbachia-induced cytoplasmic incompatibility as a means for insect pest population control. Proc. Natl. Acad. Sci. USA 2004, 101, 15042–15045. [Google Scholar] [CrossRef]
- Schofield, P. Spatially explicit models of Turelli-Hoffmann Wolbachia invasive wave fronts. J. Theor. Biol. 2002, 215, 121–131. [Google Scholar] [CrossRef]
- Dorigatti, I.; McCormack, C.; Nedjati-Gilani, G.; Ferguson, N.M. Using Wolbachia for dengue control: Insights from modelling. Trends Parasitol. 2018, 34, 102–113. [Google Scholar] [CrossRef] [PubMed]
- Turelli, M.; Hoffmann, A.A.; McKechnie, S.W. Dynamics of cytoplasmic incompatibility and mtDNA variation in natural Drosophila simulans populations. Genetics 1992, 132, 713–723. [Google Scholar]
- Hurst, G.D.D.; Jiggins, F.M. Problems with mitochondrial DNA as a marker in population, phylogeographic and phylogenetic studies: The effects of inherited symbionts. Proc. R. Soc. B Biol. Sci. 2005, 272, 1525–1534. [Google Scholar] [CrossRef] [PubMed]
- Kodandaramaiah, U.; Simonsen, T.J.; Bromilow, S.; Wahlberg, N.; Sperling, F. Deceptive single-locus taxonomy and phylogeography: Wolbachia-associated divergence in mitochondrial DNA is not reflected in morphology and nuclear markers in a butterfly species. Ecol. Evol. 2013, 3, 5167–5176. [Google Scholar] [CrossRef] [PubMed]
- Graham, R.I.; Wilson, K. Male-killing Wolbachia and mitochondrial selective sweep in a migratory African insect. BMC Evol. Biol. 2012, 12, 204. [Google Scholar] [CrossRef] [PubMed]
- Delgado, A.M.; Cook, J.M. Effects of a sex ratio-distorting endosymbiont on mtDNA variation in a global insect pest. BMC Evol. Biol. 2009, 9, 49. [Google Scholar] [CrossRef] [PubMed]
- Roehrdanz, R.L.; Levine, E. Wolbachia bacterial infections linked to mitochondrial DNA reproductive isolation among populations of northern corn rootworm (Coleoptera:Chrysomelidae). Ann. Entomol. Soc. Am. 2007, 100, 522–531. [Google Scholar] [CrossRef]
- Schuler, H.; Köppler, K.; Daxböck-Horvath, S.; Rasool, B.; Krumböck, S.; Schwarz, D.; Hoffmeister, T.S.; Schlick-Steiner, B.C.; Steiner, F.M.; Telschow, A.; et al. The hitchhiker’s guide to Europe: The infection dynamics of an ongoing Wolbachia invasion and mitochondrial selective sweep in Rhagoletis cerasi. Mol. Ecol. 2016, 25, 1595–1609. [Google Scholar] [CrossRef]
- Jaenike, J.; Stahlhut, J.K.; Boelio, L.M.; Unckless, R.L. Association between Wolbachia and Spiroplasma within Drosophila neotestacea: An emerging symbiotic mutualism? Mol. Ecol. 2010, 19, 414–425. [Google Scholar] [CrossRef]
- Karimi, J.; Darsouei, R. Presence of the endosymbiont Wolbachia among some fruit flies (Diptera: Tephritidae) from Iran: A multilocus sequence typing approach. J. Asia Pac. Entomol. 2014, 17, 105–112. [Google Scholar] [CrossRef]
- Mateos, M.; Martinez Montoya, H.; Lanzavecchia, S.B.; Conte, C.; Guillén, K.; Morán-Aceves, B.M.; Toledo, J.; Liedo, P.; Asimakis, E.D.; Doudoumis, V.; et al. Wolbachia pipientis associated with tephritid fruit fly pests: From basic research to applications. Front. Microbiol. 2020, 11, 1080. [Google Scholar] [CrossRef] [PubMed]
- Baldo, L.; Ayoub, N.A.; Hayashi, C.Y.; Russel, J.A.; Stahlhut, J.K.; Werren, J.H. Insight into the routes of Wolbachia invasion: High levels of horizontal transfer in the spider genus Agelenopsis revealed by Wolbachia strain and mitochondrial DNA diversity. Mol. Ecol. 2008, 17, 557–569. [Google Scholar] [CrossRef] [PubMed]
- Yeap, H.L.; Rašić, G.; Endersby-Harshman, N.M.; Lee, S.F.; Arguni, E.; Le Nguyen, H.; Hoffmann, A.A. Mitochondrial DNA variants help monitor the dynamics of Wolbachia invasion into host populations. Heredity 2016, 116, 265–276. [Google Scholar] [CrossRef]
- Turelli, M.; Hoffmann, A.A. Rapid spread of an inherited incompatibility factor in California Drosophila. Nature 1991, 353, 440–442. [Google Scholar] [CrossRef] [PubMed]
- Kriesner, P.; Hoffmann, A.A.; Lee, S.F.; Turelli, M.; Weeks, A.R. Rapid sequential spread of two Wolbachia variants in Drosophila simulans. PLoS Pathog. 2013, 9, e1003607. [Google Scholar] [CrossRef] [PubMed]
- Chan, M.H.; Kim, P.S. Modelling a Wolbachia invasion using a slow-fast dispersal reaction-diffusion approach. Bull. Math. Biol. 2013, 75, 1501–1523. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Tang, M.; Yu, J. Wolbachia infection dynamics by reaction-diffusion equations. Sci. China Math. 2015, 58, 77–96. [Google Scholar] [CrossRef]
- Fimiani, P. Mediterranean region. In Fruit Flies: Their Biology, Natural Enemies and Control; Robinson, A.S., Hopper, G., Eds.; Elsevier: Amsterdam, The Netherlands, 1989; pp. 39–50. [Google Scholar]
- Bakovic, V.; Schuler, H.; Schebeck, M.; Feder, J.L.; Stauffer, C.; Ragland, G.J. Host plant-related genomic differentiation in the European cherry fruit fly, Rhagoletis cerasi. Mol. Ecol. 2019, 28, 4648–4666. [Google Scholar] [CrossRef]
- Riegler, M.; Stauffer, C. Wolbachia infections and superinfections in cytoplasmically incompatible populations of the European cherry fruit fly Rhagoletis cerasi (Diptera, Tephritidae). Mol. Ecol. 2002, 11, 2425–2434. [Google Scholar] [CrossRef]
- Arthofer, W.; Riegler, M.; Schneider, D.; Krammer, M.; Miller, W.J.; Stauffer, C. Hidden Wolbachia diversity in field populations of the European cherry fruit fly, Rhagoletis cerasi (Diptera, Tephritidae). Mol. Ecol. 2009, 18, 3816–3830. [Google Scholar] [CrossRef]
- Augustinos, A.A.; Asimakopoulou, A.K.; Moraiti, C.A.; Mavragani-Tsipidou, P.; Papadopoulos, N.T.; Bourtzis, K. Microsatellite and Wolbachia analysis in Rhagoletis cerasi natural populations: Population structuring and multiple infections. Ecol. Evol. 2014, 4, 1943–1962. [Google Scholar] [CrossRef] [PubMed]
- Bakovic, V.; Schebeck, M.; Telschow, A.; Stauffer, C.; Schuler, H. Spatial spread of Wolbachia in Rhagoletis cerasi populations. Biol. Lett. 2018, 14, e20180161. [Google Scholar] [CrossRef] [PubMed]
- Boller, E.F.; Russ, K.; Vallo, V.; Bush, G.L. Incompatible races of European cherry fruit fly, Rhagoletis cerasi (Diptera:Tephritidae), their origin and potential use in biological control. Entomol. Exp. App. 1976, 20, 237–247. [Google Scholar] [CrossRef]
- Riegler, M.; Charlat, S.; Stauffer, C.; Mercot, H. Wolbachia transfer from a true fly into the real fruit fly: Investigating the outcomes of host/symbiont co-evolution. App. Environ. Microbiol. 2004, 70, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Schebeck, M.; Feldkirchner, L.; Stauffer, C.; Schuler, H. Dynamics of an ongoing Wolbachia spread in the European cherry fruit fly, Rhagoletis cerasi (Diptera: Tephritidae). Insects 2019, 10, 172. [Google Scholar] [CrossRef]
- Simon, C.; Frati, F.; Beckenbach, A.; Crespi, B.; Liu, H.; Flook, P. Evolution, weighting, and phylogenetic utility of mitochondrial gene-sequences and a compilation of conserved polymerase chain-reaction primers. Ann. Entomol. Soc. Am. 1994, 87, 651–701. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Carrington, L.B.; Lipkowitz, J.R.; Hoffmann, A.A.; Turelli, M. A re-examination of Wolbachia-induced cytoplasmic incompatibility in California Drosophila simulans. PLoS ONE 2011, 6, e22565. [Google Scholar] [CrossRef]
- Schuler, H.; Bertheau, C.; Egan, S.P.; Feder, J.L.; Riegler, M.; Schlick-Steiner, B.C.; Steiner, F.M.; Johannesen, J.; Kern, P.; Tuba, K.; et al. Evidence for a recent horizontal transmission and spatial spread of Wolbachia from endemic Rhagoletis cerasi (Diptera: Tephritidae) to invasive Rhagoletis cingulata in Europe. Mol. Ecol. 2013, 22, 4101–4111. [Google Scholar] [CrossRef]
- Schuler, H.; Kern, P.; Arthofer, W.; Vogt, H.; Fischer, M.; Stauffer, C.; Riegler, M. Wolbachia in parasitoids attacking native european and introduced eastern cherry fruit flies in Europe. Environ. Enthomol. 2016, 45, 1424–1431. [Google Scholar] [CrossRef]
- Ahmed, M.Z.; Li, S.J.; Xue, X.; Yin, X.J.; Ren, S.X.; Jiggins, F.M.; Greeff, J.M.; Qiu, B.L. The intracellular bacterium Wolbachia uses parasitoid wasps as phoretic vectors for efficient horizontal transmission. PLoS Pathog. 2015, 11, e1004672. [Google Scholar] [CrossRef] [PubMed]
- Lathrop, F.H.; Newton, R.C. The biology of Opius melleus Gahan, a parasite of the Blueberry Maggot. J. Agric. Res. 1933, 46, 143–160. [Google Scholar]
- Feder, J.F. The effect of parasitoids on sympatric host races of Rhagoletis pomonella (Diptera: Tephritidae). Ecology 1995, 76, 801–813. [Google Scholar] [CrossRef]
- Meijer, K.; Smit, C.; Schilthuizen, M.; Beukeboom, L.W. Fitness benefits of the fruit fly Rhagoletis alternata on a non-native rose host. Oecologia 2016, 181, 185–192. [Google Scholar] [CrossRef]
- Park, S.T.; Kim, J. Trends in next-generation sequencing and a new era for whole genome sequencing. Int. Neurourol. J. 2016, 20, S76. [Google Scholar] [CrossRef]
- Morrow, J.L.; Schneider, D.I.; Klasson, L.; Janitz, C.; Miller, W.J.; Riegler, M. Parallel sequencing of Wolbachia wCer2 from donor and novel hosts reveals multiple incompatibility factors and genome stability after host transfers. Gen. Biol. Evol. 2020, 12, 720–735. [Google Scholar] [CrossRef]
- Shropshire, J.D.; Bordenstein, S.R. Two-By-One model of cytoplasmic incompatibility: Synthetic recapitulation by transgenic expression of cifA and cifB in Drosophila. PLoS Genet. 2019, 15, e1008221. [Google Scholar] [CrossRef]
- Ellegaard, K.M.; Klasson, L.; Näslund, K.; Bourtzis, K.; Andersson, S.G. Comparative genomics of Wolbachia and the bacterial species concept. PLoS Genet. 2013, 9, e1003381. [Google Scholar] [CrossRef]

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bakovic, V.; Schebeck, M.; Stauffer, C.; Schuler, H. Wolbachia-Mitochondrial DNA Associations in Transitional Populations of Rhagoletis cerasi. Insects 2020, 11, 675. https://doi.org/10.3390/insects11100675
Bakovic V, Schebeck M, Stauffer C, Schuler H. Wolbachia-Mitochondrial DNA Associations in Transitional Populations of Rhagoletis cerasi. Insects. 2020; 11(10):675. https://doi.org/10.3390/insects11100675
Chicago/Turabian StyleBakovic, Vid, Martin Schebeck, Christian Stauffer, and Hannes Schuler. 2020. "Wolbachia-Mitochondrial DNA Associations in Transitional Populations of Rhagoletis cerasi" Insects 11, no. 10: 675. https://doi.org/10.3390/insects11100675
APA StyleBakovic, V., Schebeck, M., Stauffer, C., & Schuler, H. (2020). Wolbachia-Mitochondrial DNA Associations in Transitional Populations of Rhagoletis cerasi. Insects, 11(10), 675. https://doi.org/10.3390/insects11100675