
Role of B-Cell Activating Factor (BAFF) in Inflammatory Bowel Disease

 ,
,  ,
,  ,
, 
Abstract
1. Introduction
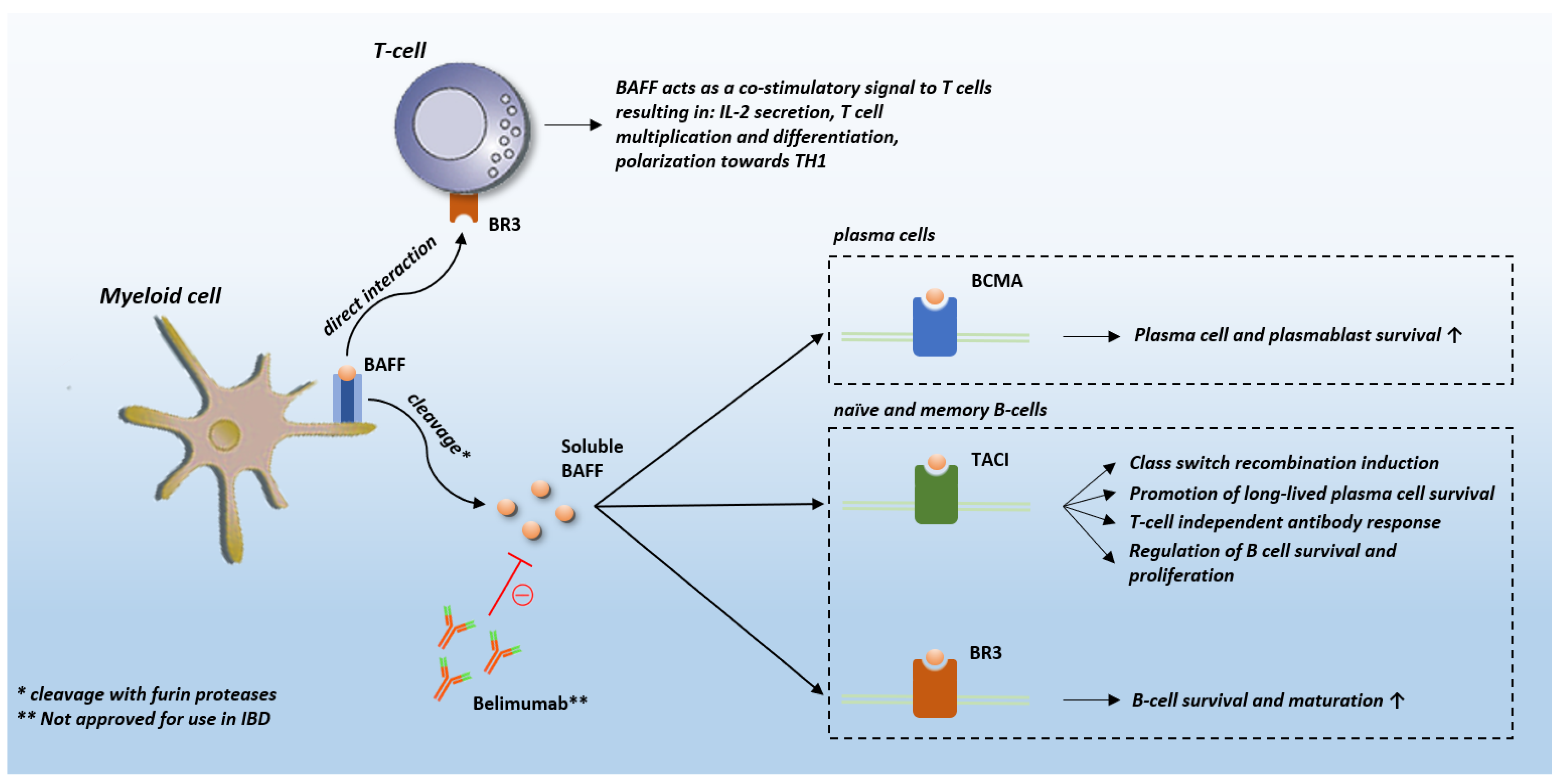
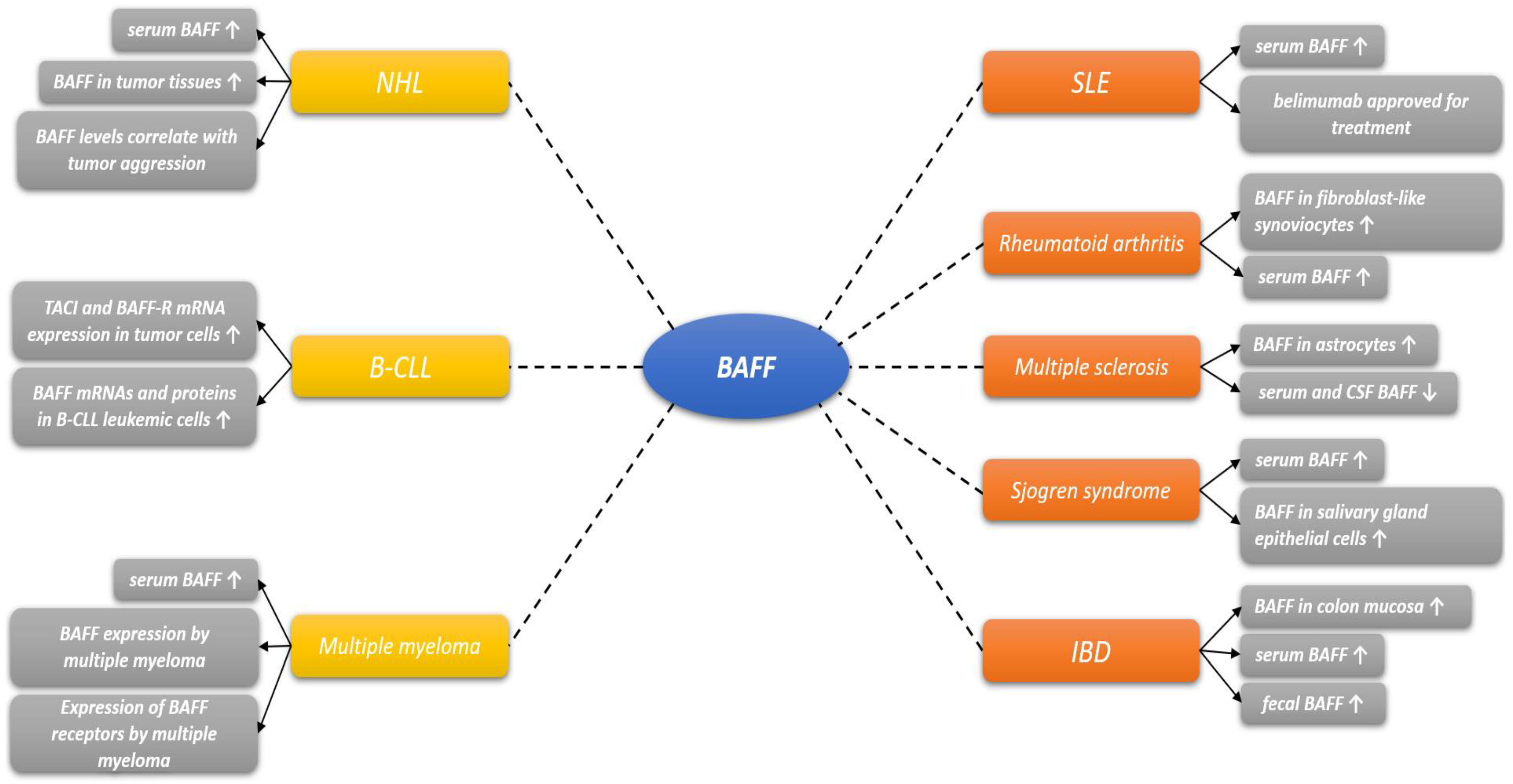
2. BAFF Biology and Role in Other Diseases
3. BAFF in IBD
3.1. Role of B-Cells in IBD
3.2. Pathophysiological Background of BAFF in IBD
3.3. Clinical Implications of BAFF in IBD
4. Future Directions and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Brnić, D.; Martinovic, D.; Zivkovic, P.M.; Tokic, D.; Hadjina, I.T.; Rusic, D.; Vilovic, M.; Supe-Domic, D.; Tonkic, A.; Bozic, J. Serum adropin levels are reduced in patients with inflammatory bowel diseases. Sci. Rep. 2020, 10, 9264. [Google Scholar] [CrossRef]
- Zivkovic, P.M.; Matetic, A.; Hadjina, I.T.; Rusic, D.; Vilovic, M.; Supe-Domic, D.; Borovac, J.A.; Mudnic, I.; Tonkic, A.; Bozic, J. Serum Catestatin Levels and Arterial Stiffness Parameters Are Increased in Patients with Inflammatory Bowel Disease. J. Clin. Med. 2020, 9, 628. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.; Chang, E.B. Inflammatory Bowel Diseases (IBD) and the Microbiome-Searching the Crime Scene for Clues. Gastroenterology 2021, 160, 524–537. [Google Scholar] [CrossRef] [PubMed]
- Ng, S.C.; Shi, H.Y.; Hamidi, N.; Underwood, F.E.; Tang, W.; Benchimol, E.I.; Panaccione, R.; Ghosh, S.; Wu, J.C.Y.; Chan, F.K.L.; et al. Worldwide incidence and prevalence of inflammatory bowel disease in the 21st century: A systematic review of population-based studies. Lancet 2017, 390, 2769–2778. [Google Scholar] [CrossRef]
- Peyrin-Biroulet, L. Why should we define and target early Crohn’s disease? Gastroenterol. Hepatol. 2011, 7, 324–326. [Google Scholar]
- Satsangi, J.; Silverberg, M.S.; Vermeire, S.; Colombel, J.F. The Montreal classification of inflammatory bowel disease: Controversies, consensus, and implications. Gut 2006, 55, 749–753. [Google Scholar] [CrossRef]
- Langhorst, J.; Elsenbruch, S.; Koelzer, J.; Rueffer, A.; Michalsen, A.; Dobos, G.J. Noninvasive markers in the assessment of intestinal inflammation in inflammatory bowel diseases: Performance of fecal lactoferrin, calprotectin, and PMN-elastase, CRP, and clinical indices. Am. J. Gastroenterol. 2008, 103, 162–169. [Google Scholar] [CrossRef]
- Kumric, M.; Kurir, T.T.; Martinovic, D.; Zivkovic, P.M.; Bozic, J. Impact of the COVID-19 pandemic on inflammatory bowel disease patients: A review of the current evidence. World J. Gastroenterol. 2021, 27, 3748–3761. [Google Scholar] [CrossRef]
- Tindemans, I.; Joosse, M.E.; Samsom, J.N. Dissecting the Heterogeneity in T-Cell Mediated Inflammation in IBD. Cells 2020, 9, 110. [Google Scholar] [CrossRef]
- Bromke, M.A.; Neubauer, K.; Kempiński, R.; Krzystek-Korpacka, M. Faecal Calprotectin in Assessment of Mucosal Healing in Adults with Inflammatory Bowel Disease: A Meta-Analysis. J. Clin. Med. 2021, 10, 2203. [Google Scholar] [CrossRef]
- Mumolo, M.G.; Bertani, L.; Ceccarelli, L.; Laino, G.; Di Fluri, G.; Albano, E.; Tapete, G.; Costa, F. From bench to bedside: Fecal calprotectin in inflammatory bowel diseases clinical setting. World J. Gastroenterol. 2018, 24, 3681–3694. [Google Scholar] [CrossRef] [PubMed]
- Kolho, K.L.; Alfthan, H.; Hämäläinen, E. Effect of bowel cleansing for colonoscopy on fecal calprotectin levels in pediatric patients. J. Pediatr. Gastroenterol. Nutr. 2012, 55, 751–753. [Google Scholar] [CrossRef] [PubMed]
- Poullis, A.; Foster, R.; Shetty, A.; Fagerhol, M.K.; Mendall, M.A. Bowel inflammation as measured by fecal calprotectin: A link between lifestyle factors and colorectal cancer risk. Cancer Epidemiol. Biomark. Prev. 2004, 13, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Calafat, M.; Cabré, E.; Mañosa, M.; Lobatón, T.; Marín, L.; Domènech, E. High within-day variability of fecal calprotectin levels in patients with active ulcerative colitis: What is the best timing for stool sampling? Inflamm. Bowel Dis. 2015, 21, 1072–1076. [Google Scholar] [CrossRef] [PubMed]
- Striz, I. B Cell-Activating Factor (BAFF) in Inflammatory Bowel Disease: BAFFling No Longer? Dig. Dis. Sci. 2016, 61, 2456–2458. [Google Scholar] [CrossRef][Green Version]
- Sun, M.; He, C.; Cong, Y.; Liu, Z. Regulatory immune cells in regulation of intestinal inflammatory response to microbiota. Mucosal Immunol. 2015, 8, 969–978. [Google Scholar] [CrossRef]
- Scheinberg, M.A.; Hislop, C.M.; Martin, R.S. Blisibimod for treatment of systemic lupus erythematosus: With trials you become wiser. Expert Opin. Biol. Ther. 2016, 16, 723–733. [Google Scholar] [CrossRef]
- Blair, H.A.; Duggan, S.T. Belimumab: A Review in Systemic Lupus Erythematosus. Drugs 2018, 78, 355–366. [Google Scholar] [CrossRef]
- Hengeveld, P.J.; Kersten, M.J. B-cell activating factor in the pathophysiology of multiple myeloma: A target for therapy? Blood Cancer J. 2015, 5, e282. [Google Scholar] [CrossRef]
- Gross, J.A.; Johnston, J.; Mudri, S.; Enselman, R.; Dillon, S.R.; Madden, K.; Xu, W.; Parrish-Novak, J.; Foster, D.; Lofton-Day, C.; et al. TACI and BCMA are receptors for a TNF homologue implicated in B-cell autoimmune disease. Nature 2000, 404, 995–999. [Google Scholar] [CrossRef]
- Medema, J.P.; Planelles-Carazo, L.; Hardenberg, G.; Hahne, M. The uncertain glory of APRIL. Cell Death Differ. 2003, 10, 1121–1125. [Google Scholar] [CrossRef]
- Wu, Y.; Bressette, D.; Carrell, J.A.; Kaufman, T.; Feng, P.; Taylor, K.; Gan, Y.; Cho, Y.H.; Garcia, A.D.; Gollatz, E.; et al. Tumor necrosis factor (TNF) receptor superfamily member TACI is a high affinity receptor for TNF family members APRIL and BLyS. J. Biol. Chem. 2000, 275, 35478–35485. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.S.; Bixler, S.A.; Qian, F.; Vora, K.; Scott, M.L.; Cachero, T.G.; Hession, C.; Schneider, P.; Sizing, I.D.; Mullen, C.; et al. BAFF-R, a newly identified TNF receptor that specifically interacts with BAFF. Science 2001, 293, 2108–2111. [Google Scholar] [CrossRef]
- Woodland, R.T.; Fox, C.J.; Schmidt, M.R.; Hammerman, P.S.; Opferman, J.T.; Korsmeyer, S.J.; Hilbert, D.M.; Thompson, C.B. Multiple signaling pathways promote B lymphocyte stimulator dependent B-cell growth and survival. Blood 2008, 111, 750–760. [Google Scholar] [CrossRef] [PubMed]
- Mackay, F.; Schneider, P. Cracking the BAFF code. Nat. Rev. Immunol. 2009, 9, 491–502. [Google Scholar] [CrossRef] [PubMed]
- Striz, I.; Brabcova, E.; Kolesar, L.; Sekerkova, A. Cytokine networking of innate immunity cells: A potential target of therapy. Clin. Sci. 2014, 126, 593–612. [Google Scholar] [CrossRef]
- Huard, B.; Arlettaz, L.; Ambrose, C.; Kindler, V.; Mauri, D.; Roosnek, E.; Tschopp, J.; Schneider, P.; French, L.E. BAFF production by antigen-presenting cells provides T cell co-stimulation. Int. Immunol. 2004, 16, 467–475. [Google Scholar] [CrossRef]
- Boulé, M.W.; Broughton, C.; Mackay, F.; Akira, S.; Marshak-Rothstein, A.; Rifkin, I.R. Toll-like receptor 9-dependent and independent dendritic cell activation by chromatin-immunoglobulin G complexes. J. Exp Med. 2004, 199, 1631–1640. [Google Scholar] [CrossRef]
- Chu, V.T.; Enghard, P.; Riemekasten, G.; Berek, C. In vitro andin vivo activation induces BAFF and APRIL expression in B cells. J. Immunol. 2007, 179, 5947–5957. [Google Scholar] [CrossRef]
- Gorelik, L.; Gilbride, K.; Dobles, M.; Kalled, S.L.; Zandman, D.; Scott, M.L. Normal B cell homeostasis requires B cell activation factor production by radiation-resistant cells. J. Exp. Med. 2003, 198, 937–945. [Google Scholar] [CrossRef]
- Uzzan, M.; Colombel, J.F.; Cerutti, A.; Treton, X.; Mehandru, S. B Cell-Activating Factor (BAFF)-Targeted B Cell Therapies in Inflammatory Bowel Diseases. Dig. Dis. Sci. 2016, 61, 3407–3424. [Google Scholar] [CrossRef]
- Moore, P.A.; Belvedere, O.; Orr, A.; Pieri, K.; LaFleur, D.W.; Feng, P.; Soppet, D.; Charters, M.; Gentz, R.; Parmelee, D.; et al. BLyS: Member of the tumor necrosis factor family and B lymphocyte stimulator. Science 1999, 285, 260–263. [Google Scholar] [CrossRef]
- Hsu, B.L.; Harless, S.M.; Lindsley, R.C.; Hilbert, D.M.; Cancro, M.P. Cutting edge: BLyS enables survival of transitional and mature B cells through distinct mediators. J. Immunol. 2002, 168, 5993–5996. [Google Scholar] [CrossRef] [PubMed]
- Mackay, F.; Woodcock, S.A.; Lawton, P.; Ambrose, C.; Baetscher, M.; Schneider, P.; Tschopp, J.; Browning, J.L. Mice transgenic for BAFF develop lymphocytic disorders along with autoimmune manifestations. J. Exp. Med. 1999, 190, 1697–1710. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.S.; Schneider, P.; Kalled, S.L.; Wang, L.; Lefevre, E.A.; Cachero, T.G.; MacKay, F.; Bixler, S.A.; Zafari, M.; Liu, Z.Y.; et al. BAFF binds to the tumor necrosis factor receptor-like molecule B cell maturation antigen and is important for maintaining the peripheral B cell population. J. Exp. Med. 2000, 192, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Sandborn, W.J. Serologic markers in inflammatory bowel disease: State of the art. Rev. Gastroenterol. Disord. 2004, 4, 167–174. [Google Scholar] [PubMed]
- Abokor, A.A.; McDaniel, G.H.; Golonka, R.M.; Campbell, C.; Brahmandam, S.; Yeoh, B.S.; Joe, B.; Vijay-Kumar, M.; Saha, P. Immunoglobulin A, an Active Liaison for Host-Microbiota Homeostasis. Microorganisms 2021, 9, 2117. [Google Scholar] [CrossRef]
- Mackay, F.; Leung, H. The role of the BAFF/APRIL system on T cell function. Semin. Immunol. 2006, 18, 284–289. [Google Scholar] [CrossRef]
- Groom, J.; Kalled, S.L.; Cutler, A.H.; Olson, C.; Woodcock, S.A.; Schneider, P.; Tschopp, J.; Cachero, T.G.; Batten, M.; Wheway, J.; et al. Association of BAFF/ BLyS overexpression and altered B cell differentiation with Sjogren’s syndrome. J. Clin. Investig. 2002, 109, 59–68. [Google Scholar] [CrossRef]
- Lied, G.A.; Lillestøl, K.; Valeur, J.; Berstad, A. Intestinal B cell-activating factor: An indicator of non-IgE-mediated hypersensitivity reactions to food? Aliment. Pharmacol. Ther. 2010, 32, 66–73. [Google Scholar] [CrossRef]
- Ohata, J.; Zvaifler, N.J.; Nishio, M.; Boyle, D.L.; Kalled, S.L.; Carson, D.A.; Kipps, T.J. Fibroblast-like synoviocytes of mesenchymal origin express functional B cell-activating factor of the TNF family in response to proinflammatory cytokines. J. Immunol. 2005, 174, 864–870. [Google Scholar] [CrossRef] [PubMed]
- Ittah, M.; Miceli-Richard, C.; Eric Gottenberg, J.; Lavie, F.; Lazure, T.; Ba, N.; Sellam, J.; Lepajolec, C.; Mariette, X. B cell-activating factor of the tumor necrosis factor family (BAFF) is expressed under stimulation by interferon in salivary gland epithelial cells in primary Sjogren’s syndrome. Arthritis Res. Ther. 2006, 8, R51. [Google Scholar] [CrossRef] [PubMed]
- Abe, M.; Kido, S.; Hiasa, M.; Nakano, A.; Oda, A.; Amou, H.; Matsumoto, T. BAFF and APRIL as osteoclast derived survival factors for myeloma cells: A rationale for TACI-Fc treatment in patients with multiple myeloma. Leukemia 2006, 20, 1313–1315. [Google Scholar] [CrossRef] [PubMed]
- Thangarajh, M.; Gomes, A.; Masterman, T.; Hillert, J.; Hjelmström, P. Expression of B-cell-activating factor of the TNF family (BAFF) and its receptors in multiple sclerosis. J. Neuroimmunol. 2004, 152, 183–190. [Google Scholar] [CrossRef]
- Cheema, G.S.; Roschke, V.; Hilbert, D.M.; Stohl, W. Elevated serum B lymphocyte stimulator levels in patients with systemic immune based rheumatic diseases. Arthritis Rheum. 2001, 44, 1313–1319. [Google Scholar] [CrossRef]
- Seyler, T.M.; Park, Y.W.; Takemura, S.; Bram, R.J.; Kurtin, P.J.; Goronzy, J.J.; Weyand, C.M. BLyS and APRIL in rheumatoid arthritis. J. Clin. Investig. 2005, 115, 3083–3092. [Google Scholar] [CrossRef] [PubMed]
- Mariette, X.; Roux, S.; Zhang, J.; Bengoufa, D.; Lavie, F.; Zhou, T.; Kimberly, R. The level of BLyS (BAFF) correlates with the titre of autoantibodies in human Sjogren’s syndrome. Ann. Rheum. Dis. 2003, 62, 168–171. [Google Scholar] [CrossRef]
- Szodoray, P.; Jonsson, R. The BAFF/APRIL system in systemic autoimmune diseases with a special emphasis on Sjogren’s syndrome. Scand. J. Immunol. 2005, 62, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Novak, A.J.; Grote, D.M.; Stenson, M.; Ziesmer, S.C.; Witzig, T.E.; Habermann, T.M.; Harder, B.; Ristow, K.M.; Bram, R.J.; Jelinek, D.F.; et al. Expression of BLyS and its receptors in B-cell non-Hodgkin lymphoma: Correlation with disease activity and patient outcome. Blood 2004, 104, 2247–2253. [Google Scholar] [CrossRef]
- Kern, C.; Cornuel, J.F.; Billard, C.; Tang, R.; Rouillard, D.; Stenou, V.; Defrance, T.; Ajchenbaum-Cymbalista, F.; Simonin, P.Y.; Feldblum, S.; et al. Involvement of BAFF and APRIL in the resistance to apoptosis of B-CLL through an autocrine pathway. Blood 2004, 103, 679–688. [Google Scholar] [CrossRef]
- Novak, A.J.; Darce, J.R.; Arendt, B.K.; Harder, B.; Henderson, K.; Kindsvogel, W.; Gross, J.A.; Greipp, P.R.; Jelinek, D.F. Expression of BCMA, TACI, and BAFF-R in multiple myeloma: A mechanism for growth and survival. Blood 2004, 103, 689–694. [Google Scholar] [CrossRef] [PubMed]
- Krumbholz, M.; Theil, D.; Derfuss, T.; Rosenwald, A.; Schrader, F.; Monoranu, C.M.; Kalled, S.L.; Hess, D.M.; Serafini, B.; Aloisi, F.; et al. BAFF is produced by astrocytes and up-regulated in multiple sclerosis lesions and primary central nervous system lymphoma. J. Exp. Med. 2005, 201, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Ng, L.G.; Sutherland, A.P.; Newton, R.; Qian, F.; Cachero, T.G.; Scott, M.L.; Thompson, J.S.; Wheway, J.; Chtanova, T.; Groom, J.; et al. B cell-activating factor belonging to the TNF family (BAFF)-R is the principal BAFF receptor facilitating BAFF costimulation of circulating T and B cells. J. Immunol. 2004, 173, 807–817. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.Z.; Li, Y.Y. Inflammatory bowel disease: Pathogenesis. World J. Gastroenterol. 2014, 20, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Schirbel, A.; Fiocchi, C. Inflammatory bowel disease: Established and evolving considerations on its etiopathogenesis and therapy. J. Dig. Dis. 2010, 11, 266–276. [Google Scholar] [CrossRef]
- Zhao, J.; Lu, Q.; Liu, Y.; Shi, Z.; Hu, L.; Zeng, Z.; Tu, Y.; Xiao, Z.; Xu, Q. Th17 Cells in Inflammatory Bowel Disease: Cytokines, Plasticity, and Therapies. J. Immunol. Res. 2021, 2021, 8816041. [Google Scholar] [CrossRef]
- Yan, J.B.; Luo, M.M.; Chen, Z.Y.; He, B.H. The Function and Role of the Th17/Treg Cell Balance in Inflammatory Bowel Disease. J. Immunol. Res. 2020, 2020, 8813558. [Google Scholar] [CrossRef]
- Ware, C.F. Decoy receptors thwart B cells. Nature 2000, 404, 949–950. [Google Scholar] [CrossRef]
- Okai, S.; Usui, F.; Ohta, M.; Mori, H.; Kurokawa, K.; Matsumoto, S.; Kato, T.; Miyauchi, E.; Ohno, H.; Shinkura, R. Intestinal IgA as a modulator of the gut microbiota. Gut Microbes. 2017, 8, 486–492. [Google Scholar] [CrossRef]
- Yu, G.; Boone, T.; Delaney, J.; Hawkins, N.; Kelley, M.; Ramakrishnan, M.; McCabe, S.; Qiu, W.R.; Kornuc, M.; Xia, X.Z.; et al. APRIL and TALL-I and receptors BCMA and TACI: System for regulating humoral immunity. Nat. Immunol. 2000, 1, 252–256. [Google Scholar] [CrossRef]
- Do, R.K.; Chen-Kiang, S. Mechanism of BLyS action in B cell immunity. Cytokine Growth Factor Rev. 2002, 13, 19–25. [Google Scholar] [CrossRef]
- Morimoto, M.; Watanabe, T.; Yamori, M.; Takebe, M.; Wakatsuki, Y. Isoflavones regulate innate immunity and inhibit experimental colitis. J. Gastroenterol. Hepatol. 2009, 24, 1123–1129. [Google Scholar] [CrossRef]
- McDonnell, M.; Liang, Y.; Noronha, A.; Coukos, J.; Kasper, D.L.; Farraye, F.A.; Ganley-Leal, L.M. Systemic Toll-like receptor ligands modify B-cell responses in human inflammatory bowel disease. Inflamm. Bowel Dis. 2011, 17, 298–307. [Google Scholar] [CrossRef]
- Goetz, M.; Atreya, R.; Ghalibafian, M.; Galle, P.R.; Neurath, M.F. Exacerbation of ulcerative colitis after rituximab salvage therapy. Inflamm. Bowel Dis. 2007, 13, 1365–1368. [Google Scholar] [CrossRef] [PubMed]
- Olson, T.S.; Bamias, G.; Naganuma, M.; Rivera-Nieves, J.; Burcin, T.L.; Ross, W.; Morris, M.A.; Pizarro, T.T.; Ernst, P.B.; Cominelli, F.; et al. Expanded B cell population blocks regulatory T cells and exacerbates ileitis in a murine model of Crohn disease. J. Clin. Investig. 2004, 114, 389–398. [Google Scholar] [CrossRef] [PubMed]
- Takahasi, F.; Shah, H.S.; Wise, L.S.; Das, K.M. Circulating antibodies against human colonic extract enriched with a 40 kDa protein in patients with ulcerative colitis. Gut 1990, 31, 1016–1020. [Google Scholar] [CrossRef]
- Chao, L.P.; Steele, J.; Rodrigues, C.; Lennard-Jones, J.; Stanford, J.L.; Spiliadis, C.; Rook, G.A. Specificity of antibodies secreted by hybridomas generated from activated B cells in the mesenteric lymph nodes of patients with inflammatory bowel disease. Gut 1988, 29, 35–40. [Google Scholar] [CrossRef]
- Russell, M.W.; Reinholdt, J.; Kilian, M. Anti-inflammatory activity of human IgA antibodies and their Faba fragments: Inhibition of IgG-mediated complement activation. Eur. J. Immunol. 1989, 19, 2243–2249. [Google Scholar] [CrossRef]
- Han, X.; Uchida, K.; Jurickova, I.; Koch, D.; Willson, T.; Samson, C.; Bonkowski, E.; Trauernicht, A.; Kim, M.O.; Tomer, G.; et al. Granulocyte-macrophage colony-stimulating factor autoantibodies in murine ileitis and progressive ileal Crohn’s disease. Gastroenterology 2009, 136, 1261–1271. [Google Scholar] [CrossRef]
- Larabi, A.; Barnich, N.; Nguyen, H.T.T. New insights into the interplay between autophagy, gut microbiota and inflammatory responses in IBD. Autophagy 2020, 16, 38–51. [Google Scholar] [CrossRef]
- Selmi, C.; Generali, E.; Massarotti, M.; Bianchi, G.; Sciré, C.A. New treatments for inflammatory rheumatic disease. Immunol. Res. 2014, 60, 277–288. [Google Scholar] [CrossRef] [PubMed]
- Dumoitier, N.; Terrier, B.; London, J.; Lofek, S.; Mouthon, L. Implication of B lymphocytes in the pathogenesis of ANCA-associated vasculitides. Autoimmun. Rev. 2015, 14, 996–1004. [Google Scholar] [CrossRef] [PubMed]
- Jayasekera, P.; Parslew, R.; Al-Sharqi, A. A case of tumour necrosis factor-a inhibitor- and rituximab-induced plantar pustular psoriasis that completely resolved with tocilizumab. Br. J. Dermatol. 2014, 171, 1546–1549. [Google Scholar] [CrossRef] [PubMed]
- Fiorillo, L.; Wang, C.; Hemmati, I. Rituximab induced psoriasis in an infant. Pediatr. Dermatol. 2014, 31, e149–e151. [Google Scholar] [CrossRef] [PubMed]
- Wolf, S.D.; Dittel, B.N.; Hardardottir, F.; Janeway, C.A. Experimental autoimmune encephalomyelitis induction in genetically B cell-deficient mice. J. Exp. Med. 1996, 184, 2271–2278. [Google Scholar] [CrossRef]
- Ray, A.; Mann, M.K.; Basu, S.; Dittel, B.N. A case for regulatory B cells in controlling the severity of autoimmune-mediated inflammation in experimental autoimmune encephalomyelitis and multiple sclerosis. J. Neuroimmunol. 2011, 230, 1–9. [Google Scholar] [CrossRef]
- Ray, A.; Basu, S.; Williams, C.B.; Salzman, N.H.; Dittel, B.N. A novel IL-10-independent regulatory role for B cells in suppressing autoimmunity by maintenance of regulatory T cells via GITR ligand. J. Immunol. 2012, 188, 3188–3198. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Shimoda, M.; Ono, Y.; Villalobos, I.B.; Mitra, A.; Konia, T.; Grando, S.A.; Zone, J.J.; Maverakis, E. Persistence of autoreactive IgA-secreting B cells despite multiple immunosuppressive medications including Rituximab. JAMA Dermatol. 2015, 151, 646–650. [Google Scholar] [CrossRef]
- Cupi, M.L.; Sarra, M.; Marafini, I.; Monteleone, I.; Franzè, E.; Ortenzi, A.; Colantoni, A.; Sica, G.; Sileri, P.; Rosado, M.M.; et al. Plasma cells in the mucosa of patients with inflammatory bowel disease produce granzyme B and possess cytotoxic activities. J. Immunol. 2014, 192, 6083–6091. [Google Scholar] [CrossRef]
- Leiper, K.; Martin, K.; Ellis, A.; Subramanian, S.; Watson, A.J.; Christmas, S.E.; Howarth, D.; Campbell, F.; Rhodes, J.M. Randomised placebo-controlled trial of rituximab (anti-CD20) in active ulcerative colitis. Gut 2011, 60, 1520–1526. [Google Scholar] [CrossRef]
- Swaminath, A.; Magro, C.M.; Dwyer, E. Refractory urticarial vasculitis as a complication of ulcerative colitis successfully treated with rituximab. J. Clin. Rheumatol. 2011, 17, 281–283. [Google Scholar] [CrossRef] [PubMed]
- Ardelean, D.S.; Gonska, T.; Wires, S.; Cutz, E.; Griffiths, A.; Harvey, E.; Tse, S.M.; Benseler, S.M. Severe ulcerative colitis after rituximab therapy. Pediatrics 2010, 126, e243–e246. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.H.; Brant, S.R. Recent insights into the genetics of inflammatory bowel disease. Gastroenterology 2011, 140, 1704–1712. [Google Scholar] [CrossRef] [PubMed]
- Kristjánsson, V.B.; Lund, S.H.; Gröndal, G.; Sveinsdóttir, S.V.; Agnarsson, H.R.; Jónasson, J.G.; Björnsson, E.S. Increased risk of inflammatory bowel disease among patients treated with rituximab in Iceland from 2001 to 2018. Scand. J. Gastroenterol. 2021, 56, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Vincent, F.B.; Morand, E.F.; Mackay, F. BAFF and innate immunity: New therapeutic targets for systemic lupus erythematosus. Immunol. Cell Biol. 2012, 90, 293–303. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.K.; Arendt, B.K.; Darce, J.R.; Wu, X.; Jelinek, D.F. A role for BLyS in the activation of innate immune cells. Blood 2006, 108, 2687–2694. [Google Scholar] [CrossRef] [PubMed]
- Jostins, L.; Ripke, S.; Weersma, R.K.; Duerr, R.H.; McGovern, D.P.; Hui, K.Y.; Lee, J.C.; Schumm, L.P.; Sharma, Y.; Anderson, C.A.; et al. Host-microbe interactions have shaped the genetic architecture of inflammatory bowel disease. Nature 2012, 491, 119–124. [Google Scholar] [CrossRef]
- Steinbach, E.C.; Plevy, S.E. The role of macrophages and dendritic cells in the initiation of inflammation in IBD. Inflamm. Bowel Dis. 2014, 20, 166–175. [Google Scholar] [CrossRef]
- Thosani, N.; Singh, H.; Kapadia, A.; Ochi, N.; Lee, J.H.; Ajani, J.; Swisher, S.G.; Hofstetter, W.L.; Guha, S.; Bhutani, M.S. Diagnostic accuracy of EUS in differentiating mucosal versus submucosal invasion of superficial esophageal cancers: A systematic review and meta-analysis. Gastrointest. Endosc. 2012, 75, 242–253. [Google Scholar] [CrossRef]
- Mowat, A.M.; Bain, C.C. Mucosal macrophages in intestinal homeostasis and inflammation. J. Innate Immun. 2011, 3, 550–564. [Google Scholar] [CrossRef]
- Liu, Z.; Davidson, A. BAFF and selection of autoreactive B cells. Trends Immunol. 2011, 32, 388–394. [Google Scholar] [CrossRef] [PubMed]
- Mishima, Y.; Ishihara, S.; Amano, Y.; Oshima, N.; Kadota, C.; Otani, A.; Moriyama, I.; Li, Y.Y.; Aziz, M.M.; Kinoshita, Y. Alterations of peripheral blood CD5+ B cells in inflammatory bowel disease. Scand. J. Gastroenterol. 2009, 44, 172–179. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhu, Y.; Zhang, M.; Wang, H.; Jiang, Y.; Gao, P. Ulcerative Colitis Is Characterized by a Decrease in Regulatory B Cells. J. Crohn’s Colitis 2016, 10, 1212–1223. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Liu, X.; Guo, A.; Xiong, J.; Fu, Y.; Zou, K. B cell-activating factor as a new potential marker in inflammatory bowel disease. Dig. Dis. Sci. 2016, 61, 2608–2618. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, S.; Vargas, J.; Hoffmann, A. Signaling via the NFκB system. Wiley Interdiscip. Rev. Syst. Biol. Med. 2016, 8, 227–241. [Google Scholar] [CrossRef]
- Hayden, M.S.; West, A.P.; Ghosh, S. NF-kappaB and the immune response. Oncogene 2006, 25, 6758–6780. [Google Scholar] [CrossRef]
- Ordaś, I.; Eckmann, L.; Talamini, M.; Baumgart, D.C.; Sandborn, W.J. Ulcerative colitis. Lancet 2012, 380, 1606–1619. [Google Scholar] [CrossRef]
- Baumgart, D.C.; Sandborn, W.J. Crohn’s disease. Lancet 2012, 380, 1590–1605. [Google Scholar] [CrossRef]
- Rogler, G.; Brand, K.; Vogl, D.; Page, S.; Hofmeister, R.; Andus, T.; Knuechel, R.; Baeuerle, P.A.; Schölmerich, J.; Gross, V. Nuclear factor kappaB is activated in macrophages and epithelial cells of inflamed intestinal mucosa. Gastroenterology 1998, 115, 357–369. [Google Scholar] [CrossRef]
- Neurath, M.F.; Pettersson, S.; Zum Büschenfelde, K.H.M.; Strober, W. Local administration of antisense phosphorothioate oligonucleotides to the p65 subunit of NF-kappa B abrogates established experimental colitis in mice. Nat. Med. 1996, 2, 998–1004. [Google Scholar] [CrossRef]
- Schreiber, S.; Nikolaus, S.; Hampe, J. Activation of nuclear factor kappa B inflammatory bowel disease. Gut 1998, 42, 477–484. [Google Scholar] [CrossRef] [PubMed]
- Demchenko, Y.N.; Kuehl, W.M. A critical role for the NFkB pathway in multiple myeloma. Oncotarget 2010, 1, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Kane, S. Fecal lactoferrin is a sensitive and specific marker in identifying intestinal inflammation. Am. J. Gastroenterol. 2003, 98, 1309–1314. [Google Scholar] [CrossRef] [PubMed]
- Suleiman, S.; Sonnenberg, A. Cost-effectiveness of endoscopy in irritable bowel syndrome. Arch. Intern. Med. 2001, 161, 369–375. [Google Scholar] [CrossRef] [PubMed]
- Lacy, B.E.; Pimentel, M.; Brenner, D.M.; Chey, W.D.; Keefer, L.A.; Long, M.D.; Moshiree, B. ACG Clinical Guideline: Management of Irritable Bowel Syndrome. Am. J. Gastroenterol. 2021, 116, 17–44. [Google Scholar] [CrossRef] [PubMed]
- Thakkar, K.; El-Serag, H.B.; Mattek, N.; Gilger, M. Complications of pediatric colonoscopy: A five-year multicenter experience. Clin. Gastroenterol. Hepatol. 2008, 6, 515–520. [Google Scholar] [CrossRef]
- Ricciuto, A.; Griffiths, A.M. Clinical value of fecal calprotectin. Crit. Rev. Clin. Lab. Sci. 2019, 56, 307–320. [Google Scholar] [CrossRef]
- Fu, Y.; Wang, L.; Xie, C.; Zou, K.; Tu, L.; Yan, W.; Hou, X. Comparison of non-invasive biomarkers faecal BAFF, calprotectin and FOBT in discriminating IBS from IBD and evaluation of intestinal inflammation. Sci. Rep. 2017, 7, 2669. [Google Scholar] [CrossRef]
- Hussein, H.A.; Mohamed, R.S. Fecal B-cell-activating factor as a new noninvasive marker in the evaluation of ulcerative colitis Egyptian patients: A comparative cross-sectional study. Egypt J. Intern. Med. 2019, 31, 563–572. [Google Scholar]
- Xie, C.; Quan, R.; Wang, L.; Chen, C.; Yan, W.; Fu, Y. Diagnostic value of fecal B cell activating factor in patients with abdominal discomfort. Clin. Exp. Immunol. 2019, 198, 131–140. [Google Scholar] [CrossRef]
- Gisbert, J.P.; Bermejo, F.; Pérez-Calle, J.L.; Taxonera, C.; Vera, I.; McNicholl, A.G.; Algaba, A.; López, P.; López-Palacios, N.; Calvo, M.; et al. Fecal calprotectin and lactoferrin for the prediction of inflammatory bowel disease relapse. Inflamm. Bowel Dis. 2009, 15, 1190–1198. [Google Scholar] [CrossRef]
- Singh, H.K.; Ee, L.C. Recurrent Abdominal Pain in Children: Is Colonoscopy Indicated? J. Pediatr. Gastroenterol. Nutr. 2019, 68, 214–217. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, V.; Day, A.S.; Connor, S.; Leach, S.T.; Brown, G.; Singh, R.; Friedman, A.; Zekry, A.; Craig, P.I. Role of capsule endoscopy and fecal biomarkers in small-bowel Crohn’s disease to assess remission and predict relapse. Gastrointest. Endosc. 2017, 86, 1070–1078. [Google Scholar] [CrossRef] [PubMed]
- Kopylov, U.; Yablecovitch, D.; Lahat, A.; Neuman, S.; Levhar, N.; Greener, T.; Klang, E.; Rozendorn, N.; Amitai, M.M.; Ben-Horin, S.; et al. Detection of small bowel mucosal healing and deep remission in patients with known small bowel Crohn’s disease using biomarkers, capsule endoscopy, and imaging. Am. J. Gastroenterol. 2015, 110, 1316–1323. [Google Scholar] [CrossRef]
- El-Matary, W.; Abej, E.; Deora, V.; Singh, H.; Bernstein, C.N. Impact of fecal calprotectin measurement on decision-making in children with inflammatory bowel disease. Front. Pediatr. 2017, 5, 7. [Google Scholar] [CrossRef]
- Fodor, I.; Serban, O.; Serban, D.E.; Farcau, D.; Man, S.C.; Dumitrascu, D.L. B cell-activating factor (BAFF) in children with inflammatory bowel disease. Pediatr. Res. 2021, 89, 1798–1803. [Google Scholar] [CrossRef]
- Andreou, N.P.; Legaki, E.; Dovrolis, N.; Boyanov, N.; Georgiou, K.; Gkouskou, K.; Gazouli, M. B-cell activating factor (BAFF) expression is associated with Crohn’s disease and can serve as a potential prognostic indicator of disease response to Infliximab treatment. Dig. Liver Dis. 2021, 53, 574–580. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Study | Population | Purpose | Results |
|---|---|---|---|
| Zhang et al. [94] | 78 UC 37 CD 12 IBS 44 HC |
|
|
| Fu et al. [108] | 49 UC 44 CD 27 IBS 26 HC |
|
|
| Hussein et al. [109] | 50 UC * |
|
|
| Xie et al. [110] | 148 ** 44 HC | discriminative value of fecal BAFF (IBD/malignancy from healthy control) |
|
| Fodor et al. [116] | 32 CD 16 UC 10 IBS 26 HC | discriminative value of fecal BAFF (IBD from IBS in pediatric population) |
|
| Andreou et al. [117] | 112 CD 164 HC | BAFF as a potential prognostic indicator of therapeutic response to Infliximab treatment in CD |
|
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kumric, M.; Zivkovic, P.M.; Ticinovic Kurir, T.; Vrdoljak, J.; Vilovic, M.; Martinovic, D.; Bratanic, A.; Lizatovic, I.K.; Bozic, J. Role of B-Cell Activating Factor (BAFF) in Inflammatory Bowel Disease. Diagnostics 2022, 12, 45. https://doi.org/10.3390/diagnostics12010045
Kumric M, Zivkovic PM, Ticinovic Kurir T, Vrdoljak J, Vilovic M, Martinovic D, Bratanic A, Lizatovic IK, Bozic J. Role of B-Cell Activating Factor (BAFF) in Inflammatory Bowel Disease. Diagnostics. 2022; 12(1):45. https://doi.org/10.3390/diagnostics12010045
Chicago/Turabian StyleKumric, Marko, Piero Marin Zivkovic, Tina Ticinovic Kurir, Josip Vrdoljak, Marino Vilovic, Dinko Martinovic, Andre Bratanic, Ivan Kresimir Lizatovic, and Josko Bozic. 2022. "Role of B-Cell Activating Factor (BAFF) in Inflammatory Bowel Disease" Diagnostics 12, no. 1: 45. https://doi.org/10.3390/diagnostics12010045
APA StyleKumric, M., Zivkovic, P. M., Ticinovic Kurir, T., Vrdoljak, J., Vilovic, M., Martinovic, D., Bratanic, A., Lizatovic, I. K., & Bozic, J. (2022). Role of B-Cell Activating Factor (BAFF) in Inflammatory Bowel Disease. Diagnostics, 12(1), 45. https://doi.org/10.3390/diagnostics12010045