
Age-Related Changes in Stand Structure, Spatial Patterns, and Soil Physicochemical Properties in Michelia macclurei Plantations of South China

Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Experimental Design
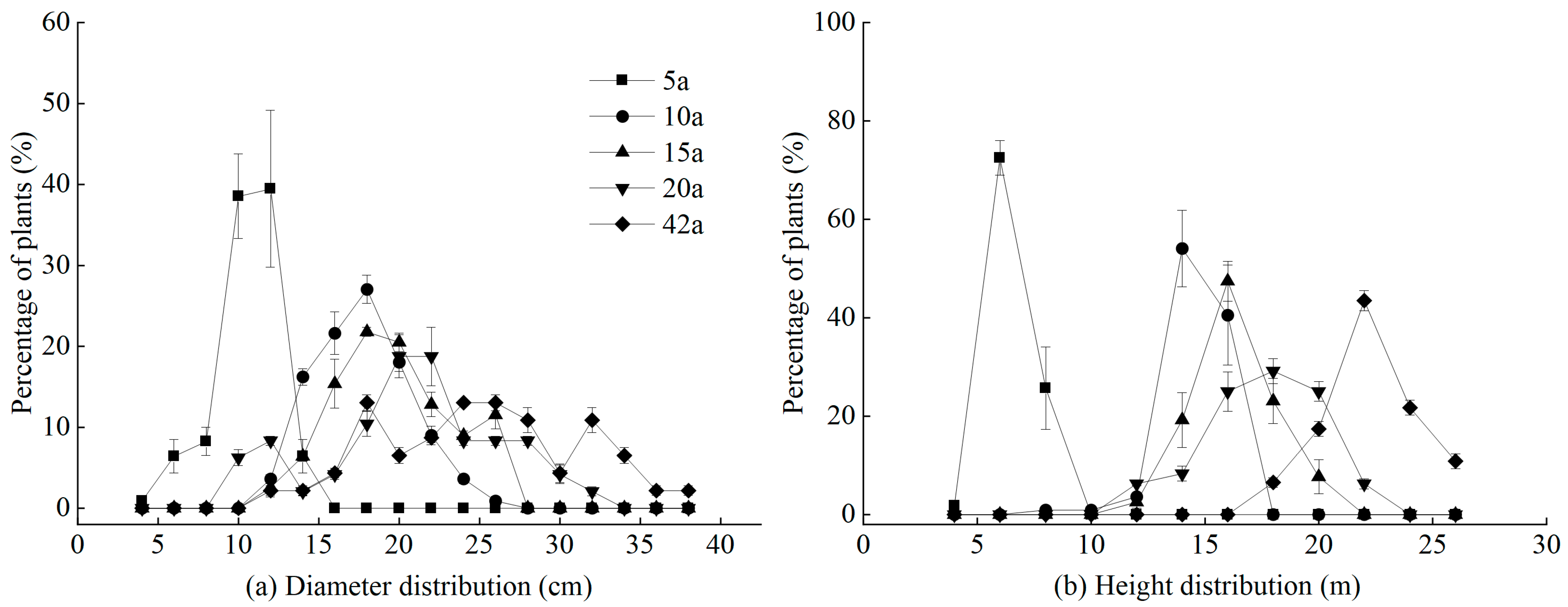
2.3. Stand Diameter Class and Height Class Structure
2.4. Crown Structure
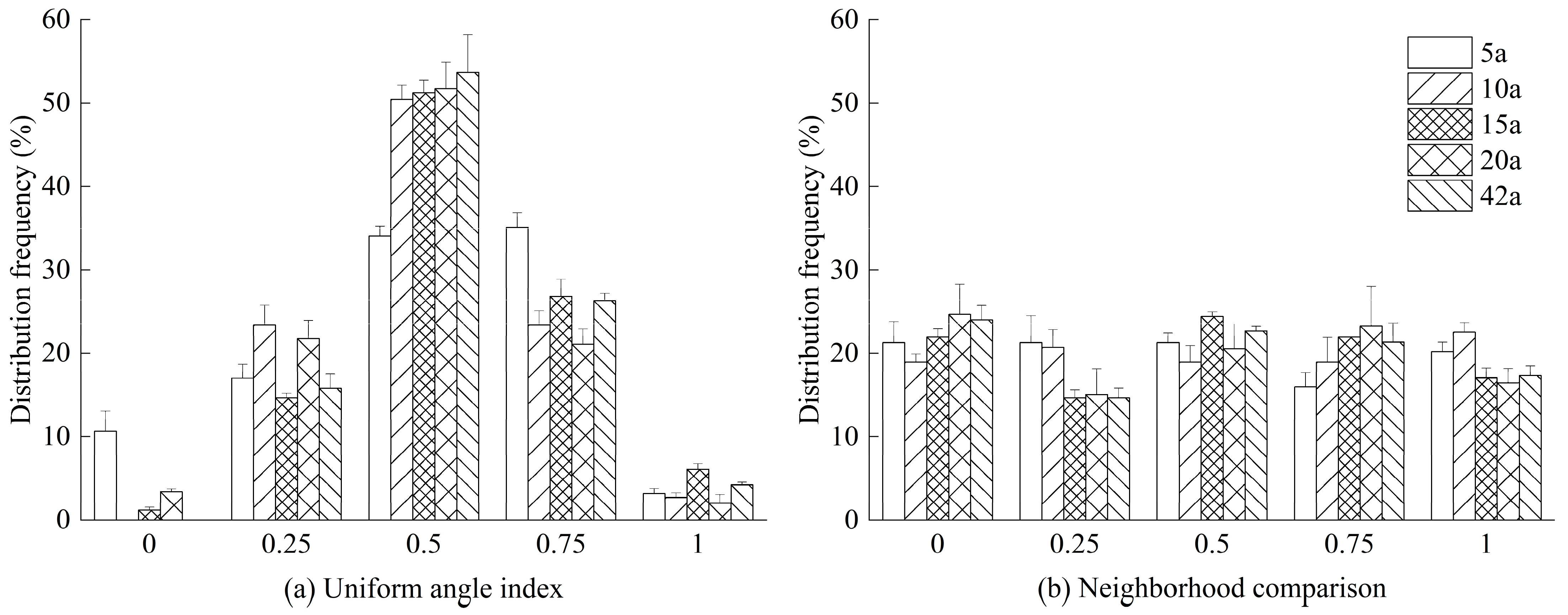
2.5. Stand Spatial Distribution Pattern
2.6. Determination of Soil Physicochemical Properties
2.7. Data Processing and Statistical Analysis
3. Results
3.1. Stand Growth of Michelia macclurei Plantations Across Different Stand Ages
3.2. Diameter Class and Height Class Structures of Michelia macclurei Plantations with Different Stand Ages
3.3. Crown Structure and Spatial Structure of Michelia macclurei Plantations Across Different Stand Ages
3.4. Soil Physical Properties of Michelia macclurei Plantations Across Different Stand Ages
3.5. Soil Chemical Properties of Michelia macclurei Plantations Across Different Stand Ages
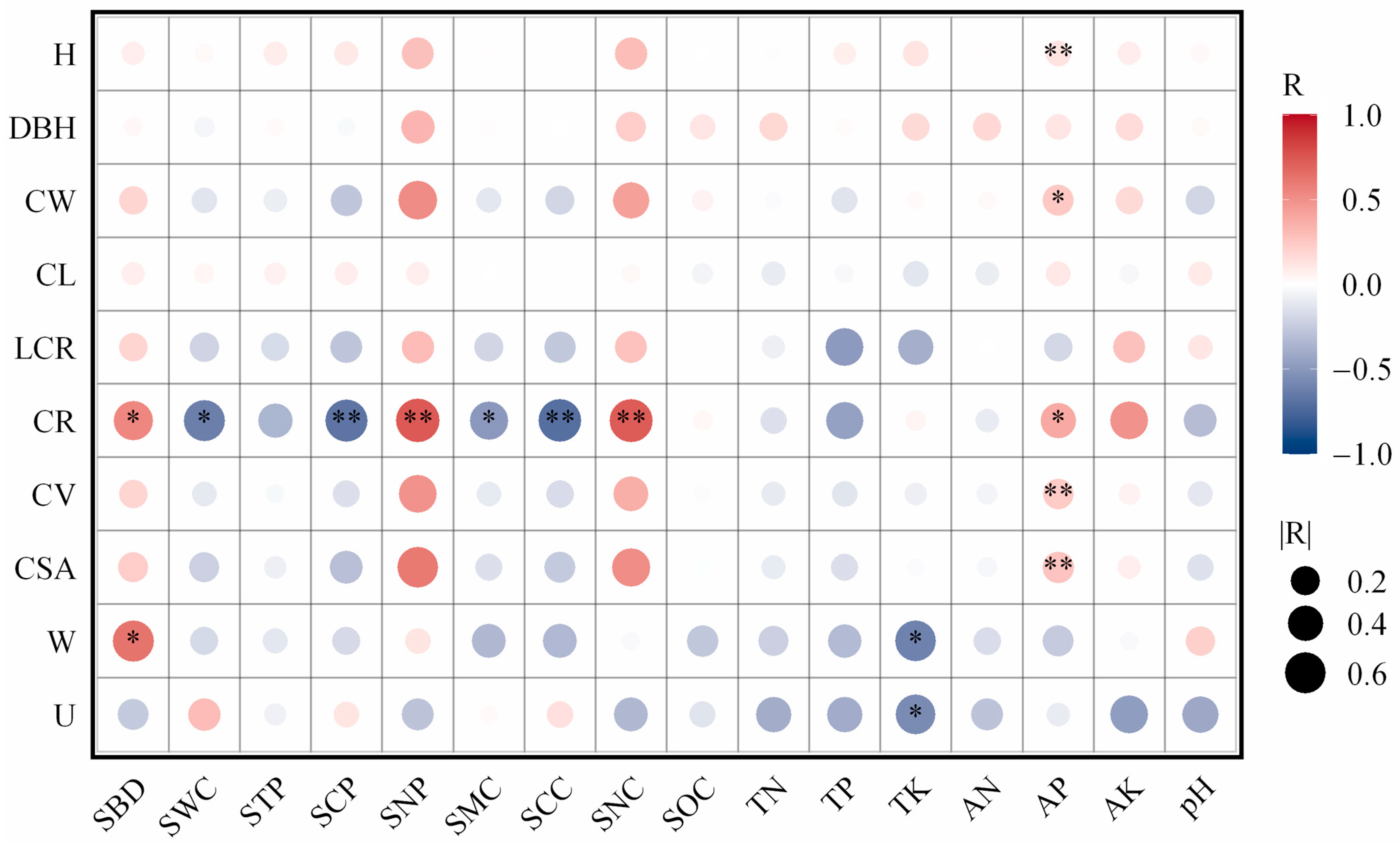
3.6. Relationship Between Stand Structure and Soil Physicochemical Properties
4. Discussion
4.1. Age-Dependent Characteristics of Stand Structure
4.2. Age-Related Characteristics of Soil Degradation
4.3. Relationship Between Stand Structure and Soil Properties
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Pryde, E.C.; Holland, G.J.; Watson, S.J.; Turton, S.M.; Nimmo, D.G. Conservation of tropical forest tree species in a native timber plantation landscape. For. Ecol. Manag. 2015, 339, 96–104. [Google Scholar] [CrossRef]
- Chen, L.; Wang, S.; Chen, C. Degradation mechanism of Chinese fir plantation. J. Appl. Ecol. 2004, 15, 1953–1957. [Google Scholar]
- Qiu, X.; Peng, D.; Li, W.; Jiang, H. Soil physicochemical properties of Pinus tabuliformis plantations of different ages in Yanqing, Beijing. Chin. J. Appl. Environ. Biol. 2018, 24, 221–231. [Google Scholar]
- Cui, N.-J.; Zhang, D.-J.; Liu, Y.; Zhang, J.; Ou, J.; Zhang, J.; Deng, C.; Ji, T.-W. Plant diversity and soil physicochemical properties under different aged Pinus massoniana plantations. Chin. J. Ecol. 2014, 33, 2610. [Google Scholar]
- Vittori Antisari, L.; Papp, R.; Vianello, G.; Marinari, S. Effects of Douglas fir stand age on soil chemical properties, nutrient dynamics, and enzyme activity: A case study in Northern Apennines, Italy. Forests 2018, 9, 641. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, S.; Gao, H.; Yu, X. Dynamics of soil active organic matter in Chinese fir plantations. J. Appl. Ecol. 2005, 16, 1270–1274. [Google Scholar]
- Ouyang, S.; Xiang, W.; Wang, X.; Xiao, W.; Chen, L.; Li, S.; Sun, H.; Deng, X.; Forrester, D.I.; Zeng, L.; et al. Effects of stand age, richness and density on productivity in subtropical forests in China. J. Ecol. 2019, 107, 2266–2277. [Google Scholar] [CrossRef]
- Hu, Y.; Li, J.; Cao, X.; Chen, J. Dynamic changes and their relationship of spatial structure and soil’s water conservation function in Cunninghamia lanceolata forest after stand improvement. J. Cent. South Univ. For. Technol. 2018, 38, 103–109. [Google Scholar]
- Liang, Y.; Qin, W.; Yu, G.; Wei, Z.; Zhang, L. Growth regularity of Michelia macclurei plantation in southeastern of Guangxi. J. Northwest For. Univ. 2010, 26, 150–154+240. [Google Scholar]
- Meng, L.; Huang, Y.; Liang, J.; Yan, Y.; Li, Y.; Feng, Y.; Yang, Z.; Tang, G. Effects of pruning on the form quality of artificial young forests of Michelia macclurei, Castanopsis hystrix and Castanopsis fissa. Sci. Silvae Sin. 2023, 59, 106–116. [Google Scholar]
- Wang, Y.; Wang, Y.; Shen, L.; Wu, Z.; Li, H.; Hu, M.; Liu, Q.; Chen, C.; Hu, X.; Zhong, Y. Evaluation of wood anatomical properties from 18 tree species in the subtropical region of China. Forests 2023, 14, 2344. [Google Scholar] [CrossRef]
- Wang, Q.-K.; Wang, S.-L.; Zhong, M.-C. Ecosystem carbon storage and soil organic carbon stability in pure and mixed stands of Cunninghamia lanceolata and Michelia macclurei. Plant Soil 2013, 370, 295–304. [Google Scholar] [CrossRef]
- Wei, S.; Song, Z.; Luo, S.; Zhong, Y.; Zhou, Y.; Lu, R. Transcriptome analysis reveals the heat stress response genes by fire stimulation in Michelia macclurei Dandy. Forests 2023, 14, 610. [Google Scholar] [CrossRef]
- Qi, D.; Zhang, F.; Wu, Z.; Yang, X.; Lan, G.; Yang, C. Effect of Michelia macclurei intercropping on soil properties and microbial population in young rubber plantation. J. Rubber Res. 2024, 27, 415–427. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, S.; Huang, Y. Leaf litter decomposition in the pure and mixed plantations of Cunninghamia lanceolata and Michelia macclurei in subtropical China. Biol. Fertil. Soils 2009, 45, 371–377. [Google Scholar] [CrossRef]
- Chen, C.; Fang, X.; Xiang, W.; Lei, P.; Ouyang, S.; Kuzyakov, Y. Soil-plant co-stimulation during forest vegetation restoration in a subtropical area of southern China. For. Ecosyst. 2020, 7, 32. [Google Scholar] [CrossRef]
- Chen, S.; Wei, B.; Fu, Y. A study of the chemical composition and biological activity of Michelia macclurei Dandy heartwood: New sources of natural antioxidants, enzyme inhibitors and bacterial inhibitors. Int. J. Mol. Sci. 2023, 24, 7972. [Google Scholar] [CrossRef]
- Ren, S.; Wang, Z.; Yan, L.; Feng, Q.; Chen, Z.; Zhao, R. Comparison of anatomical characteristics and chemical compositions between sapwood and heartwood of Michelia macclurei. Ind. Crops Prod. 2023, 193, 116190. [Google Scholar] [CrossRef]
- Wu, T.; Song, Y.; Tissue, D.; Su, W.; Luo, H.; Li, X.; Yang, S.; Liu, X.; Yan, J.; Huang, J.; et al. Photosynthetic and biochemical responses of four subtropical tree seedlings to reduced dry season and increased wet season precipitation and variable N deposition. Tree Physiol. 2024, 44, tpad114. [Google Scholar] [CrossRef]
- Zhang, X.-Y.; Bao, N.-W.; Liu, L.-T.; Mo, X.-Y.; Mei, J.-Y.; Xie, Y.-N. Structural characteristics and control techniques of Eucalyptus stand. J. Northwest For. Univ. 2016, 32, 45–51. [Google Scholar]
- Bao, N.; Zhang, S.; Mo, X. Overview of tree crown structure. Eucalypt Sci. Technol. 2021, 38, 68–74. [Google Scholar]
- Li, G.-D.; Zhang, Y.-X.; Fu, H.-M.; Li, D.-D.; Yue, Z.-W.; Xi, B.-Y.; Wang, Y. Characteristics of crown architecture of young triploid Populus tomentosa B301 clones. J. Northwest For. Univ. 2021, 36, 94–101. [Google Scholar]
- Liu, Y.; Li, C.-X.; Wang, Z.-C.; Zhao, Y.-Z.; Li, Y.-X. Stand spatial structure characteristics of two main natural secondary forests in Daxing’an Mountain. J. Northeast For. Univ. 2020, 48, 128–134. [Google Scholar]
- Ying, H.G.; Gadow, K.; Yanbo, H. The optimum standard angle of the uniform angle index. For. Res. 2004, 17, 687–692. [Google Scholar]
- Hu, X.; Zhang, H.; Yang, G.; Qiu, H.; Lei, K.; Yang, T.; Liu, Y.; Zuo, Y.; Wang, J.; Cui, Z. Visual simulation research on growth polymorphism of Chinese fir stand based on different comprehensive grade models of spatial structure parameters. Forests 2023, 14, 617. [Google Scholar] [CrossRef]
- Bao, S.D. Soil Agrochemical Analysis; China Agricultural Publishing House: Beijing, China, 2000; pp. 45–50. ISBN 9787109066441. [Google Scholar]
- Wang, L.; Yu, L.; Zhang, J.; Liu, L.; Du, J.; Gao, S.; Zhang, W. Analysis of water conservation functions of different forest types in the upper reaches of Hunhe River. J. Soil Water Conserv. 2015, 29, 249–255. [Google Scholar]
- Zhang, J.; Xu, Z.; Wen, X.; Wang, Q.; Ye, J. Spatial structure analysis and comprehensive index evaluation of fir plantation stands. J. Northwest For. Univ. 2024, 39, 10–16. [Google Scholar]
- Zhang, Y.; Li, H.; Zhang, H.; Huang, B.; Liu, C.; Jiang, Z.; Ma, X. Soil C: N: P stoichiometry and its relationship with the soil physicochemical properties of different aged Chinese fir (Cunninghamia lanceolata) plantations. Acta Ecol. Sin. 2019, 39, 2520–2531. [Google Scholar]
- Li, L.; Wu, Y.; Xiao, Z.; Ji, Y.; Lin, W.; Zhu, X.; Fan, Z. Characterization of soil ecological stoichiometry in plantation forests of different stand ages of Sonneratia apetala. For. Grassl. Resour. Res. 2023, 6, 113–119. [Google Scholar]
- Wang, D.-Z.; Zhang, D.-Y.; Zhang, Z.-D.; Mu, H.-X.; Zhang, Y.-Z. Stand structure and natural regeneration of Larix principis-rupprechtii plantation in north mountain of Hebei. J. Cent. South Univ. For. Technol. 2015, 35, 33–39. [Google Scholar]
- Li, X.; Liao, L.; Yang, H.; Huang, Y.; He, J.A.; Ye, S. Spatial pattern of artificial mixed forest of Cunninghamia lanceolata and Michelia macclurei. J. Southwest For. Univ. 2020, 40, 1–9. [Google Scholar]
- Lv, Z.; Duan, A.; Zhang, J. Influence of forest age, tree size, and climate factors on biomass and carbon storage allocation in Chinese fir forests. Ecol. Indic. 2024, 163, 112096. [Google Scholar] [CrossRef]
- Uria-Diez, J.; Pommerening, A. Crown plasticity in Scots pine (Pinus sylvestris L.) as a strategy of adaptation to competition and environmental factors. Ecol. Model. 2017, 356, 117–126. [Google Scholar] [CrossRef]
- Wan, Z.; Li, X.; Liu, C.; Sun, H.; Wang, X.; Fang, L. Dynamic analysis of relationship among tree height, diameter at the breast height and crown of artificial Sassafras tzumu forest at different development stages. Genom. Appl. Biol. 2017, 36, 331–339. [Google Scholar]
- Whittinghill, K.A.; Hobbie, S.E. Effects of pH and calcium on soil organic matter dynamics in Alaskan tundra. Biogeochemistry 2012, 111, 569–581. [Google Scholar] [CrossRef]
- Wu, Y.; Huang, Q.; Liu, X.; Zheng, R.; Cen, J.; Ding, B.; Zhang, Y.; Fu, Y. Effects of Pinus massoniana plantation age on soil physical and chemical properties in Karst areas in southwest China. J. Nanjing For. Univ. 2024, 48, 99–107. [Google Scholar]
- Yin, X.; Zhao, L.; Fang, Q.; Ding, G. Differences in soil physicochemical properties in different-aged Pinus massoniana plantations in Southwest China. Forests 2021, 12, 987. [Google Scholar] [CrossRef]
- Zhang, X.; Yang, Z.; Zha, T.; Zhang, Z.; Wang, G.; Zhu, Y.; Lü, Z. Changes in the physical properties of soil in forestlands after 22 years under the influence of the Conversion of Cropland into Farmland Project in Loess region, Western Shanxi Province. Acta Ecol. Sin. 2017, 37, 416–424. [Google Scholar]
- Zhang, J.; Cai, D.; Lu, L.; Li, Y.; Li, H.; Min, H.; Yang, B.; Nong, Y.; Huang, B. Soil ecological stoichiometry of different aged Teak (Tectona grandis) plantations. Acta Ecol. Sin. 2020, 40, 5718–5728. [Google Scholar]
- Zhang, J.; Chang, Q.; Zhang, J. Effect of different age forests on soil fertility quality of degraded ecosystems. Chin. J. Soil Sci. 2006, 37, 429–433. [Google Scholar]
- Zheng, X.; She, H.; Xue, L.; Cai, J. Effects of N and P additions on leaf litter decomposition in Michelia macclurei woodland. J. South China Agric. Univ. 2018, 39, 98–104. [Google Scholar]
- Yu, M.; Zheng, X.; Xue, L. Vertical distribution of soil carbon and nutrient storages under a Michelia macclurei plantation. J. South China Agric. Univ. 2019, 40, 77–82. [Google Scholar]
- Zheng, X.; Cai, J.; Xue, L.; She, H. Effects of external N and P on decomposition of mixed leaf litters of Michelia macclurei and Pinus massoniana woodlands. J. South China Agric. Univ. 2017, 38, 97–104. [Google Scholar]
- Jiao, J.-H.; Fu, X.; Zhang, S.; Liu, W.; Zhou, J.-J.; Wu, X.-Y.; Lin, X.-R.; Tian, Y.-L.; Tang, G.-H.; Li, P.-Q. Physiochemical properties and microorganism community structure of Zanthoxylum bungeanum rhizosphere soil at different ages. J. Northwest For. Univ. 2023, 38, 156–165. [Google Scholar]
- Sun, J.; Gao, P.; Li, C.; Wang, R.; Niu, X.; Wang, B. Ecological stoichiometry characteristics of the leaf–litter–soil continuum of Quercus acutissima Carr. and Pinus densiflora Sieb. in Northern China. Environ. Earth Sci. 2019, 78, 20. [Google Scholar] [CrossRef]
- Meng, C.; Zheng, X.-X.; Wang, W. Correlation between stand structure and soil of water conservation forest in Badaling forest farm of Beijing. J. Northwest For. Univ. 2016, 31, 99–105. [Google Scholar]
- Zhang, C.; Zhao, Q.; Cai, Y.; Li, X.; He, T. Effect of litter input on root exudative carbon input and rhizosphere nitrogen. J. For. Environ. 2021, 41, 263–271. [Google Scholar]
- Niu, S.; Zhou, Y.; Liu, L.; Qin, S.; Yin, Y.; Song, X.; Xiao, W. Soil properties in Pinus sylvestris var. mongolica plantation of different ages. J. Northeast For. Univ. 2015, 43, 47–50+62. [Google Scholar]
- Lei, L.-Q.; Lu, L.-H.; Nong, Y.; Ming, A.-G.; Liu, S.-L.; He, Y. Stoichiometry characterization of soil C, N and P of Pinus massoniana plantations at different age stages. For. Res. 2017, 3, 954–960. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
| Stand Age (a) | Altitude (m) | Slope (°) | Slope Aspect | Canopy Density | Stand Density (Plant·hm−2) | Understory Vegetation Shannon–Wiener Index |
|---|---|---|---|---|---|---|
| 5 | 322 | 28 | Southeast | 0.89 | 900.00 ± 72.17 | 2.9631 |
| 10 | 122 | 22 | Southeast | 0.92 | 925.00 ± 28.87 | 1.7463 |
| 15 | 151 | 31 | Southwest | 0.93 | 650.00 ± 62.92 | 1.8930 |
| 20 | 243 | 30 | Southwest | 0.89 | 400.00 ± 62.92 | 2.1030 |
| 42 | 178 | 36 | Southeast | 0.88 | 383.33 ± 46.40 | 1.9039 |
| Stand Age (a) | Tree Height (m) | DBH (cm) | Crown Width (m) | Individual Tree Volume (m3) |
|---|---|---|---|---|
| 5 | 6.48 ± 0.16 d | 10.60 ± 0.43 d | 2.84 ± 0.23 c | 0.03 ± 0.00 d |
| 10 | 14.63 ± 0.45 c | 17.96 ± 0.29 c | 5.01 ± 0.21 c | 0.19 ± 0.01 c |
| 15 | 16.30 ± 0.67 bc | 19.85 ± 0.21 bc | 3.63 ± 0.09 b | 0.25 ± 0.01 bc |
| 20 | 17.97 ± 0.74 b | 21.51 ± 1.16 b | 5.29 ± 0.78 b | 0.32 ± 0.04 b |
| 42 | 21.96 ± 0.49 a | 25.52 ± 1.47 a | 7.58 ± 0.33 a | 0.53 ± 0.06 a |
| Stand Age (a) | Crown Width (m) | Crown Length (m) | Live Crown Ratio | Crown Projection Ratio | Crown Volume (m3) | Crown Surface Area (m2) |
|---|---|---|---|---|---|---|
| 5 | 2.84 ± 0.23 c | 4.42 ± 0.08 d | 0.68 ± 0.02 a | 0.28 ± 0.01 ab | 10.46 ± 1.06 d | 9.74 ± 1.72 c |
| 10 | 4.01 ± 0.21 bc | 9.51 ± 0.10 c | 0.65 ± 0.03 a | 0.23 ± 0.02 b | 31.02 ± 1.48 c | 42.33 ± 3.53 c |
| 15 | 4.63 ± 0.09 b | 9.91 ± 0.96 bc | 0.60 ± 0.04 a | 0.24 ± 0.00 b | 38.53 ± 3.49 bc | 61.70 ± 6.60 bc |
| 20 | 5.29 ± 0.78 b | 11.38 ± 0.48 b | 0.65 ± 0.02 a | 0.26 ± 0.04 ab | 53.82 ± 11.37 b | 111.67 ± 39.47 b |
| 42 | 7.54 ± 0.33 a | 14.84 ± 0.54 a | 0.67 ± 0.04 a | 0.31 ± 0.01 a | 91.76 ± 3.24 a | 229.98 ± 13.55 a |
| Stand Age (a) | Uniform Angle Index | Neighborhood Comparison |
|---|---|---|
| 5 | 0.5720 ± 0.0280 a | 0.4854 ± 0.0154 a |
| 10 | 0.5143 ± 0.0135 ab | 0.5137 ± 0.0106 a |
| 15 | 0.5552 ± 0.0187 ab | 0.4936 ± 0.0110 a |
| 20 | 0.4883 ± 0.0120 b | 0.4795 ± 0.0060 a |
| 42 | 0.5010 ± 0.0305 ab | 0.4902 ± 0.0151 a |
| Stand Age (a) | Layer (cm) | Soil Bulk Density (g·cm−3) | Soil Total Porosity (%) | Capillary Porosity (%) | Non-Capillary Porosity (%) | Saturated Water-Holding Capacity (t·hm−2) | Capillary Water-Holding Capacity (t·hm−2) | Non-Capillary Water-Holding Capacity (t·hm−2) |
|---|---|---|---|---|---|---|---|---|
| 5 | 0–20 | 0.99 ± 0.07 a | 43.95 ± 2.21 a | 40.53 ±1.47 ab | 3.42 ± 0.77 ab | 904.77 ± 106.41 a | 832.28 ± 85.00 a | 72.49 ± 21.47 ab |
| 20–40 | 1.38 ± 0.10 a | 40.87 ± 0.41 ab | 37.81 ± 0.94 c | 3.07 ± 0.56 ab | 600.47 ± 52.14 a | 556.3 ± 54.44 a | 44.18 ± 5.82 b | |
| 40–60 | 1.40 ± 0.05 a | 40.55 ± 1.46 ab | 37.27 ± 1.31 b | 3.28 ± 0.48 ab | 582.25 ± 35.27 b | 535.43 ± 34.70 b | 46.82 ± 6.25 ab | |
| 10 | 0–20 | 1.07 ± 0.01 b | 41.08 ± 0.47 ab | 40.40 ± 0.41 ab | 0.68 ± 0.08 b | 769.06 ± 5.99 a | 756.20 ± 6.79 a | 12.86 ± 1.21 b |
| 20–40 | 1.08 ± 0.02 a | 40.87 ± 1.87 a | 40.22 ± 1.78 ab | 0.65 ± 0.10 b | 758.88 ± 38.88 a | 746.74 ± 37.44 a | 12.13 ± 1.90 b | |
| 40–60 | 1.31 ± 0.00 a | 43.85 ± 0.23 a | 43.17 ± 0.15 a | 0.69 ± 0.23 c | 668.89 ± 3.36 a | 658.45 ± 4.21 a | 10.44 ± 3.50 b | |
| 15 | 0–20 | 1.05 ± 0.07 a | 43.85 ± 4.63 a | 43.05 ± 4.63 a | 0.80 ± 0.18 b | 863.58 ± 119.72 a | 848.52 ± 120.63 a | 15.06 ± 2.87 b |
| 20–40 | 1.25 ± 0.06 ab | 41.86 ± 0.49 a | 40.45±0.09ab | 1.41 ± 0.58 b | 675.83 ± 26.64 ab | 653.95 ± 33.96 a | 21.88 ± 7.89 b | |
| 40–60 | 1.30 ± 0.04 a | 42.25±0.32a | 40.82 ± 0.64 ab | 1.44 ± 0.37 bc | 655.08 ± 14.61 a | 632.75 ± 17.66 a | 22.33 ± 5.70 b | |
| 20 | 0–20 | 0.97 ± 0.05 a | 38.20 ± 1.10 a | 37.18 ± 0.92 ab | 1.02 ± 0.30 b | 796.45 ± 27.83 a | 775.84 ± 32.68 a | 20.61 ± 5.05 b |
| 20–40 | 1.13 ± 0.11 a | 40.67 ± 0.96 ab | 39.34 ± 0.62 bc | 1.33 ± 0.67 bc | 739.7 ± 87.77 a | 716.92 ± 90.14 a | 22.78 ± 9.87 b | |
| 40–60 | 1.26 ± 0.07 ab | 43.20 ± 0.58 a | 41.90 ± 0.86 a | 1.30 ± 0.35 b | 691.58 ± 42.15 ab | 670.49 ± 37.47 a | 21.09 ± 6.88 b | |
| 42 | 0–20 | 1.10 ± 0.07 a | 41.07 ± 1.91 a | 34.55±0.45b | 6.53 ± 2.16 a | 757.70 ± 85.37 a | 632.96 ± 38.59 a | 124.74 ± 49.89 a |
| 20–40 | 1.24 ± 0.08 a | 38.66 ± 2.08 b | 33.79 ± 1.40 d | 4.87 ± 1.00 a | 634.75 ± 69.38 a | 554.18 ± 53.77 a | 80.57 ± 19.17 a | |
| 40–60 | 1.30 ± 0.12 ab | 37.78 ± 1.69 b | 31.58 ± 2.37 c | 6.20 ± 2.30 a | 598.48 ± 82.55 b | 494.25 ± 46.12 b | 104.20 ± 47.02 a |
| Stand Age (a) | Layer (cm) | Organic Matter (g·kg−1) | Total Nitrogen (g·kg−1) | Total Phosphorus (g·kg−1) | Total Potassium (g·kg−1) | Alkali-Hydrolyzable Nitrogen (mg·kg−1) | Available Phosphorus (mg·kg−1) | Available Potassium (mg·kg−1) | pH (H2O) |
|---|---|---|---|---|---|---|---|---|---|
| 5 | 0–20 | 47.52 ± 7.98 a | 1.76 ± 0.25 a | 0.19 ± 0.02 ab | 8.62 ± 0.42 b | 185.56 ± 24.71 a | 0.75 ± 0.20 a | 38.73 ± 10.64 a | 4.21 ± 0.02 ab |
| 20–40 | 17.84 ± 3.44 a | 0.73 ± 0.13 b | 0.14 ± 0.01 b | 9.76 ± 0.74 b | 75.94 ± 12.07 a | 0.39 ± 0.03 a | 25.43 ± 2.29 a | 4.30 ± 0.02 bc | |
| 40–60 | 12.65 ± 3.11 a | 0.55 ± 0.04 b | 0.14 ± 0.01c | 10.25 ± 0.86 b | 51.12 ± 6.16 a | 0.35 ± 0.09 a | 18.16±0.96a | 4.33 ± 0.06 b | |
| 10 | 0–20 | 26.32 ± 7.14 b | 0.96 ± 0.18 c | 0.19±0.02 ab | 6.65 ± 1.17 b | 102.49 ± 14.35 b | 0.75 ± 0.30 a | 16.04 ± 1.56 b | 4.15 ± 0.11 b |
| 20–40 | 18.24 ± 3.49 a | 0.65 ± 0.08 b | 0.18 ± 0.02 ab | 7.12 ± 1.37 b | 71.28 ± 5.98 a | 0.42 ± 0.1 a | 12.77 ± 1.35 b | 4.23 ± 0.05 c | |
| 40–60 | 10.42 ± 2.11 a | 0.50 ± 0.04 b | 0.17 ± 0.03 abc | 7.45 ± 1.38 b | 48.42 ± 8.24 a | 0.22 ± 0.06 a | 10.28 ± 0.87 bc | 4.27 ± 0.04 b | |
| 15 | 0–20 | 39.03 ± 3.67 ab | 1.44 ± 0.08 abc | 0.23 ± 0.01 a | 8.16 ± 0.20 b | 132.72 ± 7.46 b | 1.51 ± 1.05 a | 14.14 ± 1.39 b | 4.37 ± 0.04 ab |
| 20–40 | 13.31 ± 2.04 a | 0.65 ± 0.06 b | 0.20 ± 0.01 a | 9.18 ± 0.41 b | 57.27 ± 8.15 a | 1.30 ± 0.75 a | 8.55 ± 0.53 b | 4.39 ± 0.04 ab | |
| 40–60 | 11.25 ± 1.78 a | 0.56 ± 0.04 b | 0.20 ± 0.01 ab | 9.54 ± 0.60 b | 51.61 ± 6.85 a | 0.19 ± 0.03 a | 7.78 ± 0.42c | 4.38 ± 0.04 ab | |
| 20 | 0–20 | 36.71 ± 2.33 ab | 1.55 ± 0.06 ab | 0.22 ± 0.02 a | 17.43 ± 0.64 a | 135.42 ± 7.99 b | 0.85 ± 0.33 a | 26.18 ± 2.12 ab | 4.20±0.09ab |
| 20–40 | 23.56 ± 5.06 a | 1.17 ± 0.16 a | 0.21 ± 0.01 a | 18.2 ± 0.81 a | 93.64 ± 14.77 a | 1.93 ± 1.66 a | 21.32 ± 1.24 a | 4.20 ± 0.03 c | |
| 40–60 | 12.81 ± 2.17 a | 0.86 ± 0.09 a | 0.22 ± 0.01 a | 20.27 ± 1.06 a | 60.71 ± 8.30 a | 1.15 ± 0.74 a | 17.23 ± 0.96 ab | 4.26 ± 0.04 b | |
| 42 | 0–20 | 31.83 ± 2.10 ab | 1.08 ± 0.06 bc | 0.16 ± 0.02 b | 7.37 ± 3.99 a | 105.44 ± 6.65 b | 0.75 ± 0.28 a | 22.33 ± 1.63 ab | 4.39 ± 0.03 a |
| 20–40 | 21.63 ± 4.49 a | 0.83 ± 0.14 ab | 0.15 ± 0.02 b | 18.97 ± 4.89 a | 79.14 ± 15.45 a | 0.45 ± 0.12 a | 22.51 ± 2.78 a | 4.44 ± 0.02 a | |
| 40–60 | 11.54 ± 2.66 a | 0.54 ± 0.09 b | 0.15 ± 0.02 bc | 19.61 ± 5.38 a | 55.79 ± 12.08 a | 0.42 ± 0.10 a | 19.21 ± 4.90 a | 4.51 ± 0.02 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, J.; Fang, J.; Lu, D.; Li, C.; Shuai, X.; Zheng, F.; Chen, H. Age-Related Changes in Stand Structure, Spatial Patterns, and Soil Physicochemical Properties in Michelia macclurei Plantations of South China. Life 2025, 15, 917. https://doi.org/10.3390/life15060917
Yang J, Fang J, Lu D, Li C, Shuai X, Zheng F, Chen H. Age-Related Changes in Stand Structure, Spatial Patterns, and Soil Physicochemical Properties in Michelia macclurei Plantations of South China. Life. 2025; 15(6):917. https://doi.org/10.3390/life15060917
Chicago/Turabian StyleYang, Jiaman, Jianbo Fang, Dehao Lu, Cheng Li, Xiaomai Shuai, Fenglin Zheng, and Honyue Chen. 2025. "Age-Related Changes in Stand Structure, Spatial Patterns, and Soil Physicochemical Properties in Michelia macclurei Plantations of South China" Life 15, no. 6: 917. https://doi.org/10.3390/life15060917
APA StyleYang, J., Fang, J., Lu, D., Li, C., Shuai, X., Zheng, F., & Chen, H. (2025). Age-Related Changes in Stand Structure, Spatial Patterns, and Soil Physicochemical Properties in Michelia macclurei Plantations of South China. Life, 15(6), 917. https://doi.org/10.3390/life15060917