
Cloning and Spatiotemporal Expression Analysis of IGF1R Gene cDNA in Alopex lagopus (Arctic Fox)


Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Materials
2.1.1. Experimental Animals
2.1.2. Main Reagents
2.2. Experimental Methods
2.2.1. Tissue Sample Collection
2.2.2. Total RNA Extraction and cDNA Synthesis
2.2.3. Cloning, Sequencing, and Bioinformatics Analysis of IGF1R Gene CDS Region
2.2.4. Spatiotemporal Expression Analysis of IGF1R Gene in the Arctic Fox
3. Results and Analysis
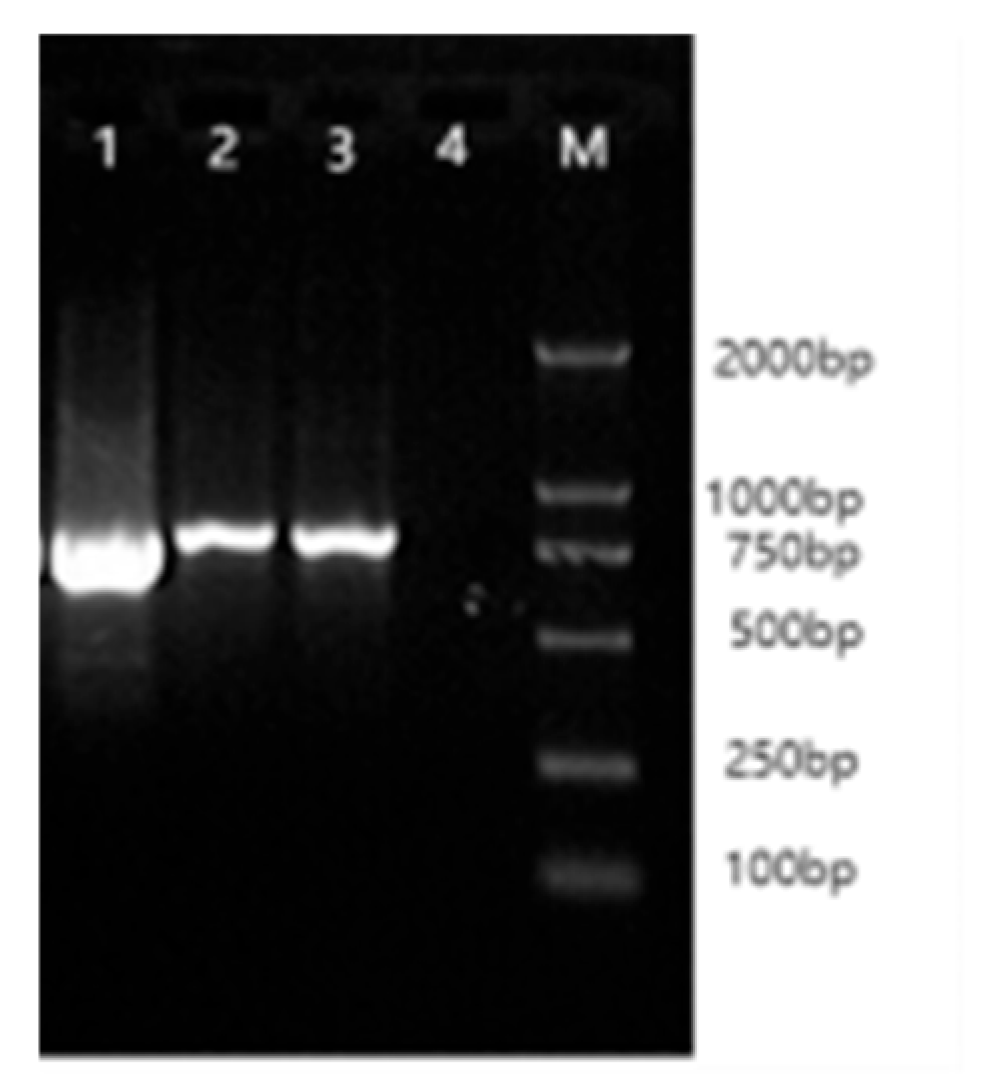
3.1. IGF1R Gene CDS Amplification Results
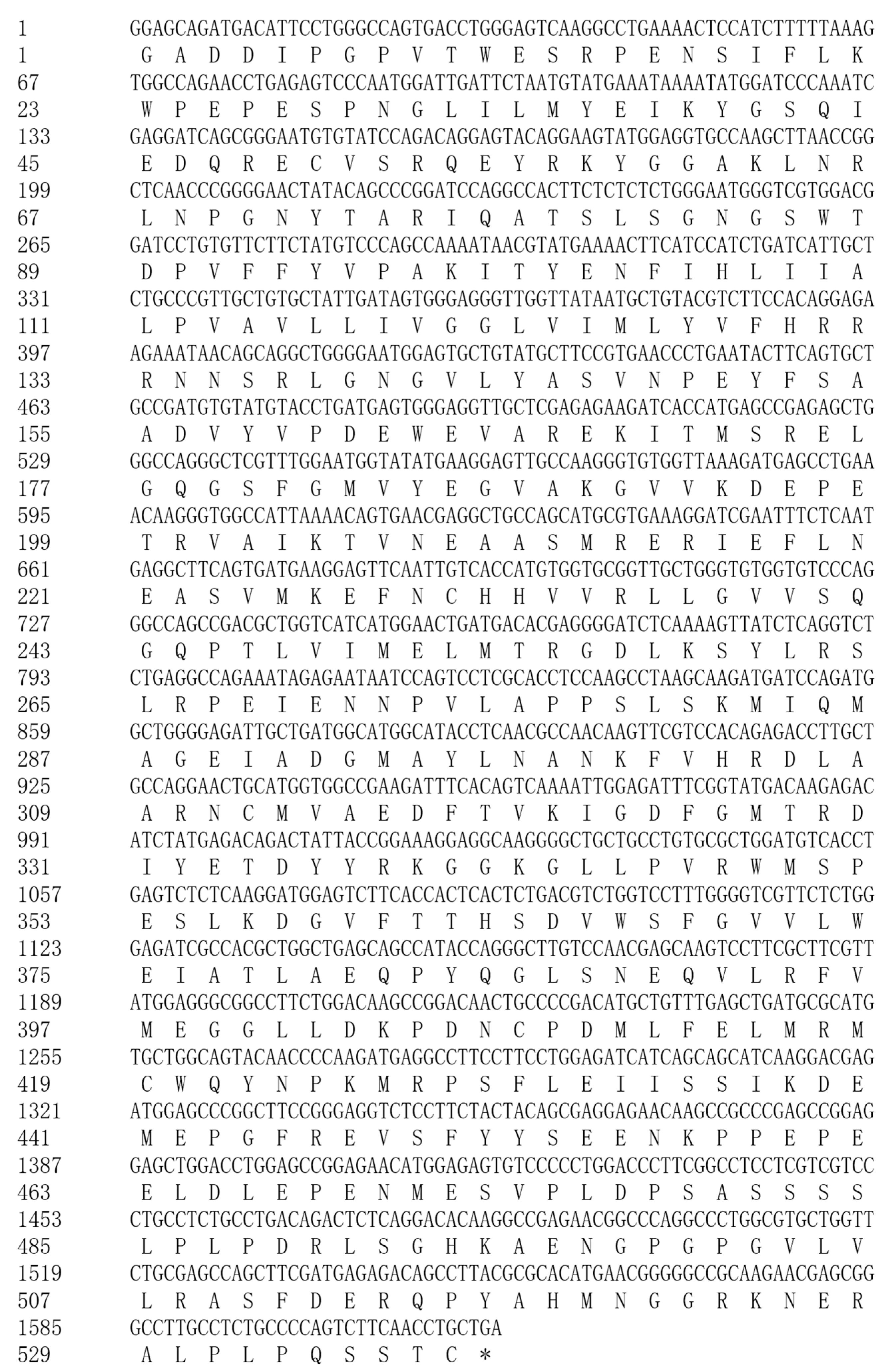
3.2. Sequencing Results and Sequence Analysis
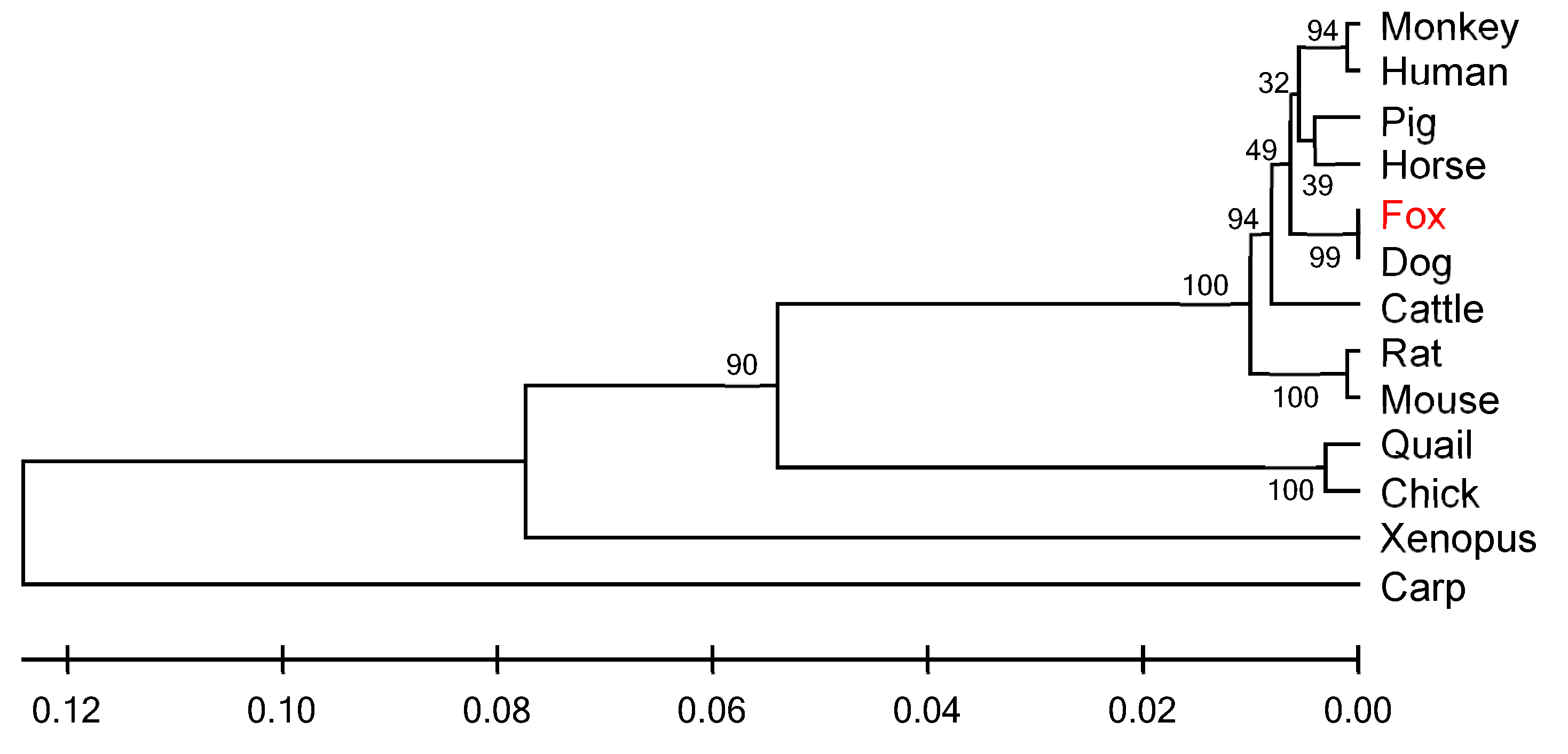
3.3. Similarity Analysis and Phylogenetic Tree Construction of IGF1R Gene
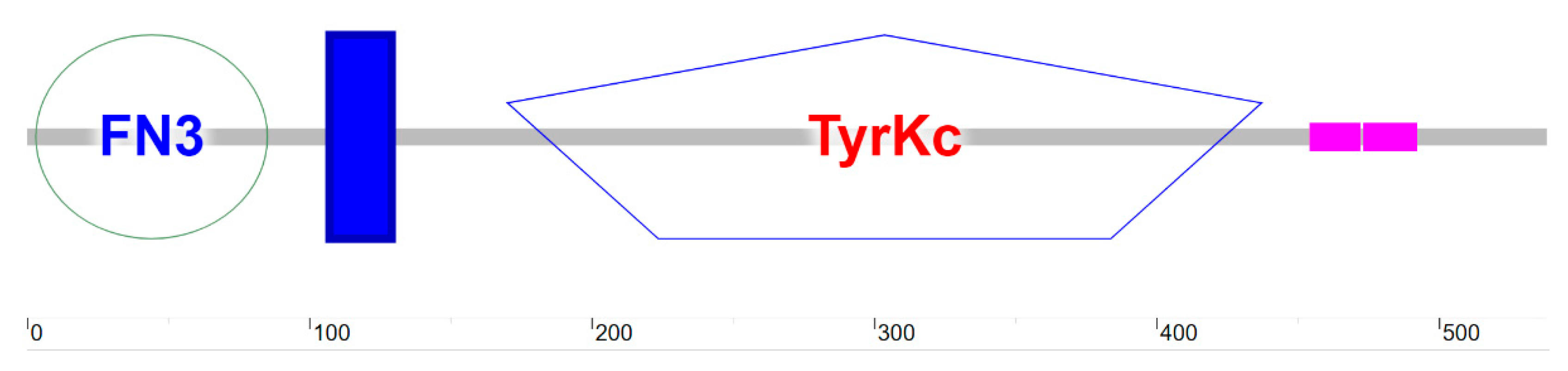
3.4. Structural and Functional Analysis of Arctic Fox IGF1R Protein
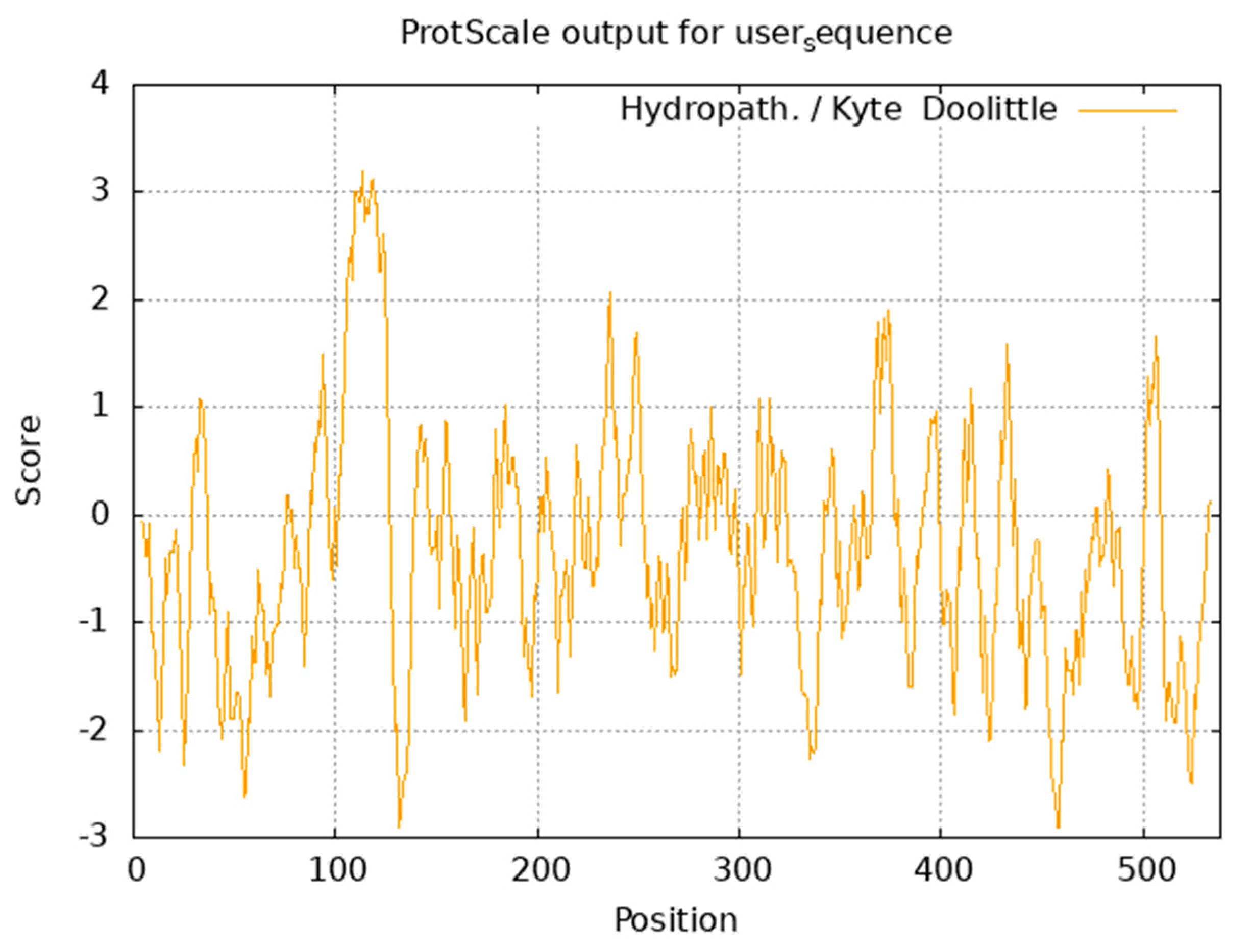
3.4.1. Physicochemical Properties and Hydrophobicity
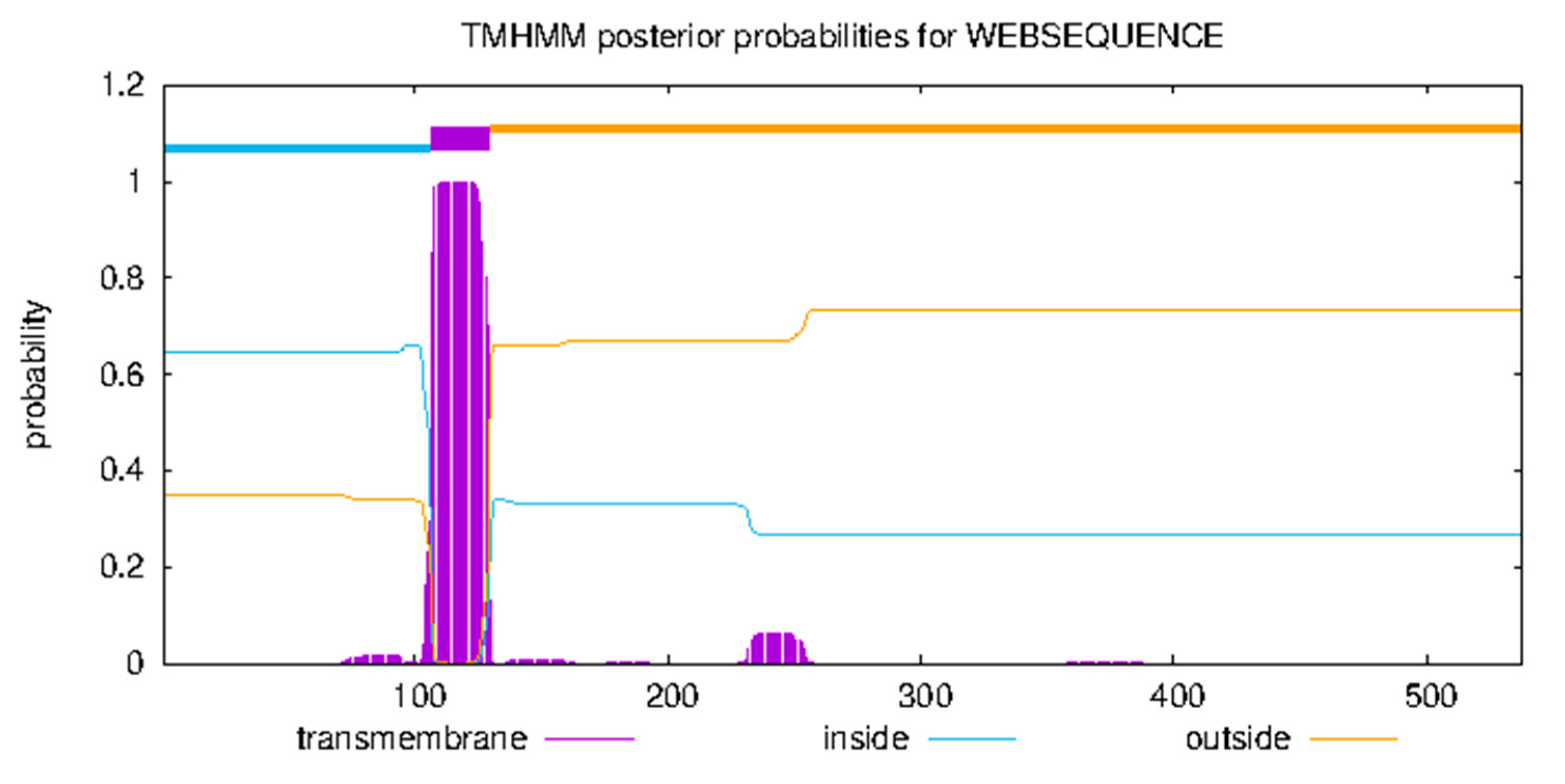
3.4.2. Transmembrane Domain and Subcellular Localization
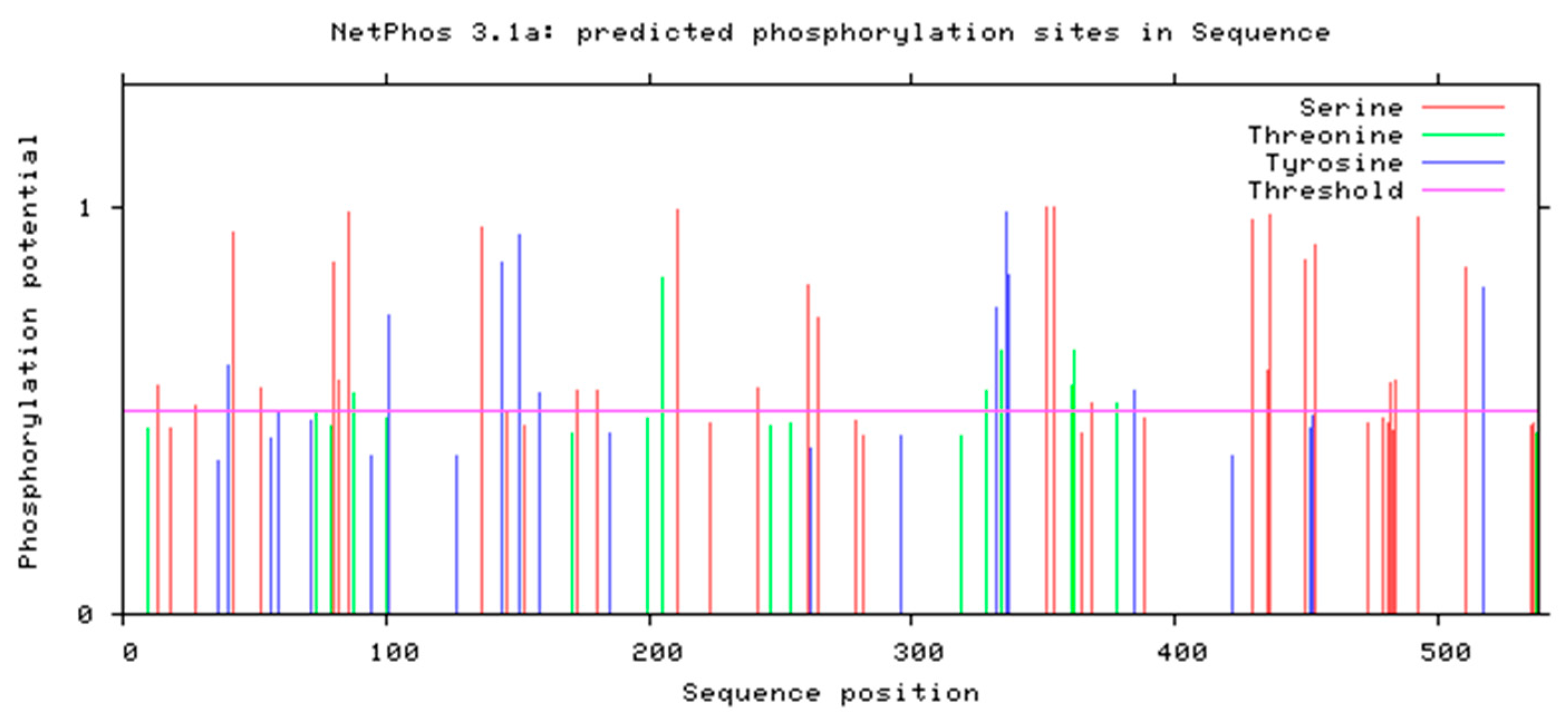
3.4.3. Phosphorylation Site Prediction
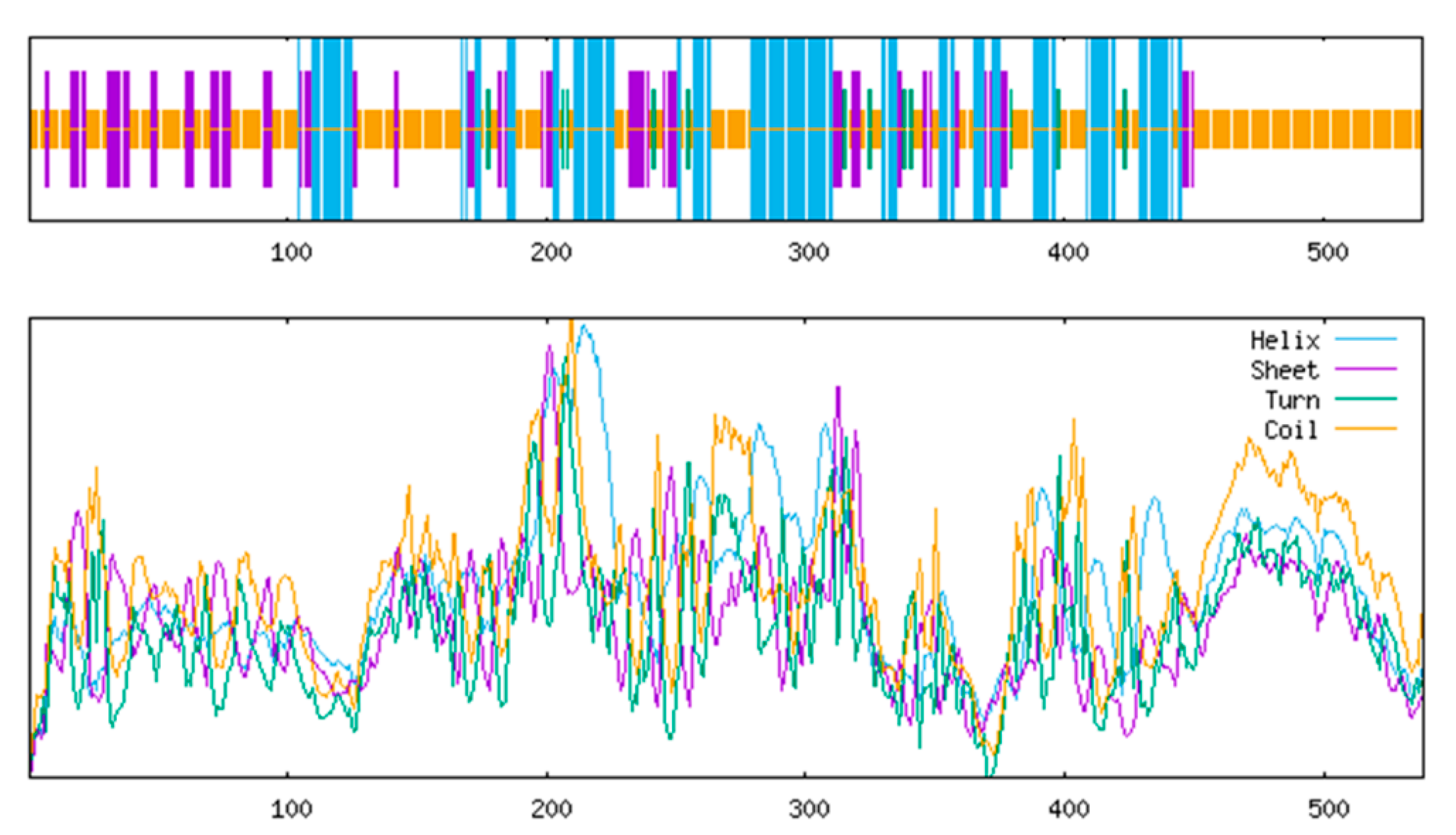
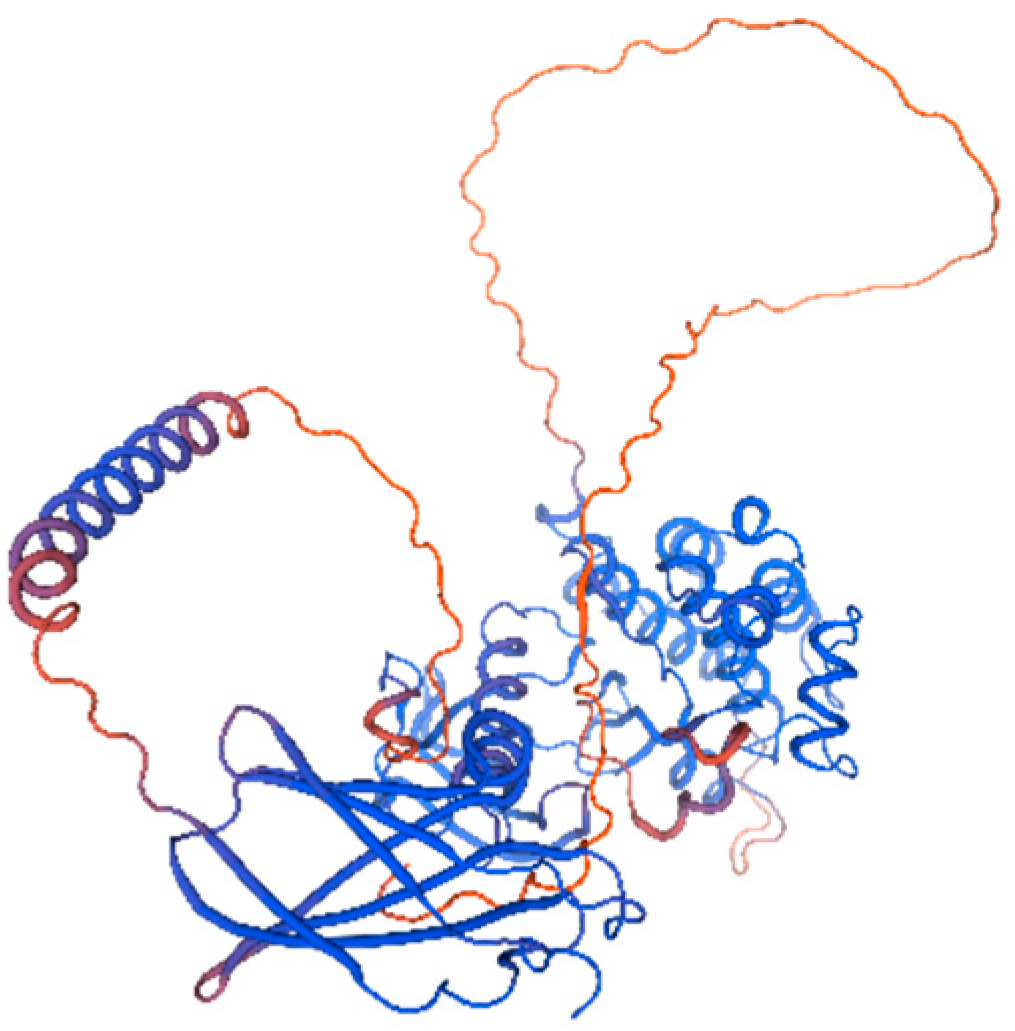
3.4.4. Secondary and Tertiary Structure Prediction
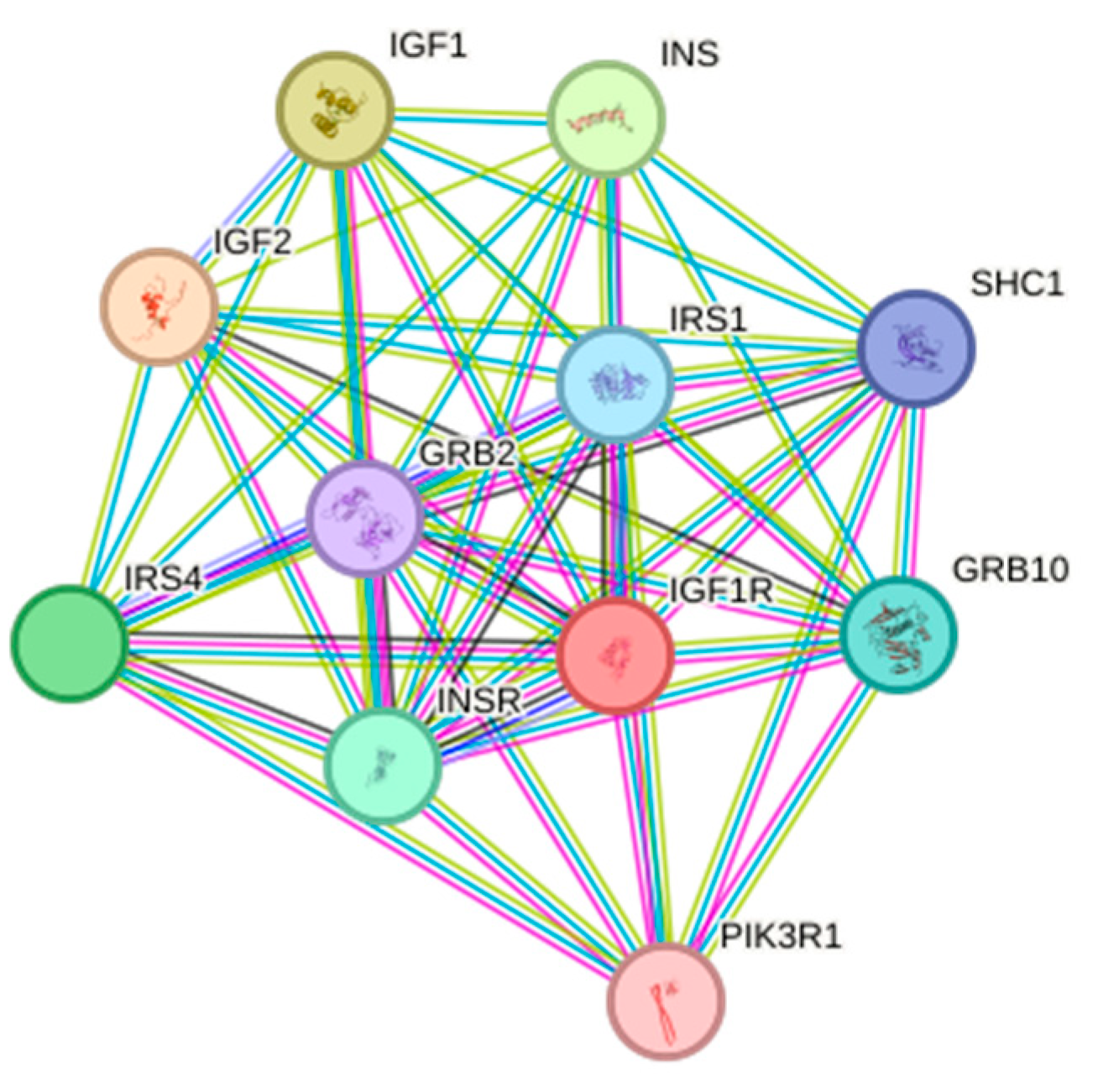
3.4.5. Protein Interaction Analysis
3.5. Tissue-Specific and Spatiotemporal Expression Patterns of IGF1R Gene in the Arctic Fox
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| IGFIR | Insulin-like growth factor 1 receptor |
| cDNA | Complementary DNA |
| IGF1 | Insulin like growth factor 1 |
| IGF2 | Insulin like growth factor 2 |
| MC1R | Melanocortin 1 receptor |
| GAPDH | Glyceraldehyde-3-phosphate dehydrogenase |
| CDS | Coding sequences |
| ORF | Open reading frame |
| DEPC | Diethylpyrocarbonate |
| UTR | Untranslated regions |
| PI | Isoelectric point |
| LSM | Least squares method |
| SNP | Single-nucleotide polymorphism |
| FSH | Follicle-stimulating hormone |
| GH | Growth hormone |
References
- Alex, P.; Kanakkaparambil, R.; Gopalakrishnan, R.; Ramasamy, C.; Thazhathuveettil, A. The effect of insulin-like growth factor 1 receptor gene single nucleotide polymorphism on growth and milk production traits in two native Indian tropical goat breeds. Anim. Biotechnol. 2023, 34, 4828–4836. [Google Scholar] [CrossRef] [PubMed]
- Baker, J.; Liu, J.P.; Robertson, E.J.; Efstratiadis, A. Role of insulin-like growth factors in embryonic and postnatal growth. Cell 1993, 75, 73–82. [Google Scholar] [CrossRef]
- Constância, M.; Hemberger, M.; Hughes, J.; Dean, W.; Ferguson-Smith, A.; Fundele, R.; Stewart, F.; Kelsey, G.; Fowden, A.; Sibley, C.; et al. Placental-specific IGF-II is a major modulator of placental and fetal growth. Nature 2002, 417, 945–948. [Google Scholar] [CrossRef] [PubMed]
- Woods, K.A.; Camacho-Hübner, C.; Savage, M.O.; Clark, A.J. Intrauterine growth retardation and postnatal growth failure associated with deletion of the insulin-like growth factor I gene. N. Engl. J. Med. 1996, 335, 1363–1367. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.P.; Baker, J.; Perkins, A.S.; Robertson, E.J.; Efstratiadis, A. Mice carrying null mutations of the genes encoding insulin-like growth factor I (Igf-1) and type 1 IGF receptor (IGF1R). Cell 1993, 75, 59–72. [Google Scholar] [CrossRef]
- Galifi, C.A.; Wood, T.L. Insulin-like growth factor-1 receptor crosstalk with integrins, cadherins, and the tumor microenvironment: Sticking points in understanding IGF1R function in cancer. Endocr.-Relat. Cancer 2023, 30, e230031. [Google Scholar] [CrossRef]
- James, H.A.; O’Neill, B.T.; Nair, K.S. Insulin Regulation of Proteostasis and Clinical Implications. Cell Metab. 2017, 26, 310–323. [Google Scholar] [CrossRef]
- Pierzchała, M.; Pareek, C.S.; Urbański, P.; Goluch, D.; Kamyczek, M.; Różycki, M.; Smoczynski, R.; Horbańczuk, J.O.; Kurył, J. Study of the differential transcription in liver of growth hormone receptor (GHR), insulin-like growth factors (IGF1, IGF2) and insulin-like growth factor receptor (IGF1R) genes at different postnatal developmental ages in pig breeds. Mol. Biol. Rep. 2012, 39, 3055–3066. [Google Scholar] [CrossRef]
- Zheng, W.; Leng, X.; Vinsky, M.; Li, C.; Jiang, H. Association of body weight gain with muscle, fat, and liver expression levels of growth hormone receptor, insulin-like growth factor I, and beta-adrenergic receptor mRNAs in steers. Domest. Anim. Endocrinol. 2018, 64, 31–37. [Google Scholar] [CrossRef]
- Harumi, T.; Maruyama, K.; Kagami, H.; Sano, A.; Matsubara, Y.; Tagami, T.; Naito, M. Cloning of porcine IGF1 receptor cDNA and detection of sequence polymorphisms using RT-PCR. Anim. Genet. 2001, 32, 386–389. [Google Scholar] [CrossRef]
- Hong, Z.; Wang, D.; Qiao, X.; Xie, Y.; Yang, S.; Hao, K.; Han, C.; Liu, H.; Liu, Z. Wnt5a negatively regulates melanogenesis in primary Arctic fox epidermal melanocytes. Gene 2025, 934, 149045. [Google Scholar] [CrossRef]
- Våge, D.I.; Fuglei, E.; Snipstad, K.; Beheim, J.; Landsem, V.M.; Klungland, H. Two cysteine substitutions in the MC1R generate the blue variant of the Arctic fox (Alopex lagopus) and prevent expression of the white winter coat. Peptides 2005, 26, 1814–1817. [Google Scholar] [CrossRef]
- Jaillardon, L.; Abadie, J.; Godard, T.; Campone, M.; Loussouarn, D.; Siliart, B.; Nguyen, F. The dog as a naturally-occurring model for insulin-like growth factor type 1 receptor-overexpressing breast cancer: An observational cohort study. BMC Cancer 2015, 15, 664. [Google Scholar] [CrossRef] [PubMed]
- Forbes, B.E. Molecular mechanisms underlying insulin-like growth factor action: How mutations in the GH: IGF axis lead to short stature. Pediatr. Endocrinol. Rev. 2011, 8, 374–381. [Google Scholar]
- Holzenberger, M.; Dupont, J.; Ducos, B.; Leneuve, P.; Géloën, A.; Even, P.C.; Cervera, P.; Le Bouc, Y. IGF-1 receptor regulates lifespan and resistance to oxidative stress in mice. Nature 2003, 421, 182–187. [Google Scholar] [CrossRef] [PubMed]
- López, I.P.; Rodriguez-de la Rosa, L.; Pais, R.S.; Piñeiro-Hermida, S.; Torrens, R.; Contreras, J.; Varela-Nieto, I.; Pichel, J.G. Differential organ phenotypes after postnatal IGF1R gene conditional deletion induced by tamoxifen in UBC-CreERT2; IGF1R fl/fl double transgenic mice. Transgenic Res. 2015, 24, 279–294. [Google Scholar] [CrossRef]
- Epaud, R.; Aubey, F.; Xu, J.; Chaker, Z.; Clemessy, M.; Dautin, A.; Ahamed, K.; Bonora, M.; Hoyeau, N.; Fléjou, J.F.; et al. Knockout of insulin-like growth factor-1 receptor impairs distal lung morphogenesis. PLoS ONE 2012, 7, e48071. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.L.; Wen, Y.F.; Cao, X.K.; Cheng, J.; Huang, Y.Z.; Ma, Y.; Hu, L.Y.; Lei, C.Z.; Qi, X.L.; Cao, H.; et al. Copy number variation (CNV) in the IGF1R gene across four cattle breeds and its association with economic traits. Arch. Anim. Breed. 2019, 62, 171–179. [Google Scholar] [CrossRef]
- Ding, N.; Tian, D.; Li, X.; Zhang, Z.; Tian, F.; Liu, S.; Han, B.; Liu, D.; Zhao, K. Genetic Polymorphisms of IGF1 and IGF1R Genes and Their Effects on Growth Traits in Hulun Buir Sheep. Genes 2022, 13, 666. [Google Scholar] [CrossRef]
- Jin, S.; Chen, S.; Li, H.; Lu, Y.; Xu, G.; Yang, N. Associations of polymorphisms in GHRL, GHSR, and IGF1R genes with feed efficiency in chickens. Mol. Biol. Rep. 2014, 41, 3973–3979. [Google Scholar] [CrossRef]
- Shah, A.; Ahmad, I.; Ahmad, I.; Amin, A.; Rather, M.A. Gene Characterization, Molecular Docking and Dynamic Simulations of Insulin-Like Growth Factor Receptor (IGF-1Ra) in Common Carp, Cyprinus carpio. Proc. Zool. Soc. 2023, 76, 382–396. [Google Scholar] [CrossRef]
- Niu, S.; Wang, Z.; Ge, D.; Zhang, G.; Li, Y. Prediction of functional phosphorylation sites by incorporating evolutionary information. Protein Cell 2012, 3, 675–690. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Zeng, S.; Xu, C.; Qiu, W.; Liang, Y.; Joshi, T.; Xu, D. MusiteDeep: A deep-learning framework for general and kinase-specific phosphorylation site prediction. Bioinformatics 2017, 33, 3909–3916. [Google Scholar] [CrossRef] [PubMed]
- Skorstengaard, K.; Jensen, M.S.; Sahl, P.; Petersen, T.E.; Magnusson, S. Complete primary structure of bovine plasma fibronectin. Eur. J. Biochem. 1986, 161, 441–453. [Google Scholar] [CrossRef]
- Hou, Q.; Gong, R.; Liu, X.; Mao, H.; Xu, X.; Liu, D.; Dai, Z.; Wang, H.; Wang, B.; Hu, C. Poly I:C facilitates the phosphorylation of Ctenopharyngodon idellus type I IFN receptor subunits and JAK kinase. Fish Shellfish. Immunol. 2017, 60, 13–20. [Google Scholar] [CrossRef]
- Blyth, A.J.; Kirk, N.S.; Forbes, B.E. Understanding IGF-II Action through Insights into Receptor Binding and Activation. Cells 2020, 9, 2276. [Google Scholar] [CrossRef]
- Grochowska, E.; Lisiak, D.; Akram, M.Z.; Adeniyi, O.O.; Lühken, G.; Borys, B. Association of a polymorphism in exon 3 of the IGF1R gene with growth, body size, slaughter and meat quality traits in Colored Polish Merino sheep. Meat Sci. 2021, 172, 108314. [Google Scholar] [CrossRef]
- Wang, C.; Liu, S.; Wu, Q.; Cheng, Y.; Feng, T.; Song, J.; Yang, R.; Geng, H.; Lu, G.; Wang, S.; et al. Porcine IGF-1R synonymous mutations in the intracellular domain affect cell proliferation and alter kinase activity. Int. J. Biol. Macromol. 2020, 152, 147–153. [Google Scholar] [CrossRef]
- Szewczuk, M.; Zych, S.; Wójcik, J.; Czerniawska-Piątkowska, E. Association of two SNPs in the coding region of the insulin-like growth factor 1 receptor (IGF1R) gene with growth-related traits in Angus cattle. J. Appl. Genet. 2013, 54, 305–308. [Google Scholar] [CrossRef]
- Yakar, S.; Adamo, M.L. Insulin-like growth factor 1 physiology: Lessons from mouse models. Endocrinol. Metab. Clin. N. Am. 2012, 41, 231–247. [Google Scholar] [CrossRef]
- Hoyt, J.A.; Cozzi, E.; D’Alessio, D.A.; Thompson, C.C.; Aroda, V.R. A look at duodenal mucosal resurfacing: Rationale for targeting the duodenum in type 2 diabetes. Diabetes Obes. Metab. 2024, 26, 2017–2028. [Google Scholar] [CrossRef] [PubMed]
- Serrat, M.A.; Lovejoy, C.O.; King, D. Age- and site-specific decline in insulin-like growth factor-I receptor expression is correlated with differential growth plate activity in the mouse hindlimb. Anat. Rec. 2007, 290, 375–381. [Google Scholar] [CrossRef] [PubMed]
- Yoon, M.J.; Berger, T.; Roser, J.F. Localization of insulin-like growth factor-I (IGF-I) and IGF-I receptor (IGF-IR) in equine testes. Reprod. Domest. Anim. = Zuchthyg. 2011, 46, 221–228. [Google Scholar] [CrossRef]
- Li, R.; Pourpak, A.; Morris, S.W. Inhibition of the insulin-like growth factor-1 receptor (IGF1R) tyrosine kinase as a novel cancer therapy approach. J. Med. Chem. 2009, 52, 4981–5004. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Sequences | Annealing Temperature | Application |
|---|---|---|---|
| IGF1R-1 | F: GCAGAAGGAGCAGATGACATTC R: GAGGTGCGAGGACTGGATTAT | 54 | PCR amplification |
| IGF1R-2 | F: AATCCAGTCCTCGCACCTC R: AGGAGGCTGTGGCTAGATTG | 60 | PCR amplification |
| IGF1R-3 | F: GGAGTTCTAACAATCTAGCC R: ACCTGCGTGAATGTAAGAC | 40 | PCR amplification |
| IGF1R | F: GTCCTTCGCTTCGTTATG R: CTTGTTCTCCTCGCTGTAG | 60 | qRT-PCR |
| GAPDH | F: GGGTGATGCTGGTGCTG R: ACGGTCTTCTGGGTGGC | 60 | qRT-PCR |
| Analysis Tools | Websites | Function Prediction |
|---|---|---|
| ExPASy | https://web.expasy.org/protscale/ (accessed on 24 February 2025) | Physicochemical properties |
| ProtSca | https://web.expasy.org/protscale (accessed on 24 February 2025) | Hydrophilicity/hydrophobicity |
| Novopro | https://www.novopro.cn/tools/tmhmm.html (accessed on 24 February 2025) | Transmembrane domain |
| SMART | http://smart.embl-heidelberg.de/ (accessed on 24 February 2025) | Functional domain |
| NetPhos-3.1 | https://services.healthtech.dtu.dk/services/NetPhos-3.1/ (accessed on 25 February 2025) | Phosphorylation sites |
| SOPMA | https://npsa.lyon.inserm.fr/cgi-bin/ (accessed on 25 February 2025) | Secondary structure |
| SWISS-MODEL | https://swissmodel.expasy.org/ (accessed on 25 February 2025) | Tertiary structure |
| STRING | https://cn.string-db.org/ (accessed on 25 February 2025) | Protein–protein interaction |
| Species | Similarity of Mature | Amino Acid Number | GenBank Accession No. |
|---|---|---|---|
| Canis familiaris | 99% | 538 | XP_545828 |
| Sus scrofa | 98% | 538 | NP_999337 |
| Equus caballus | 98% | 538 | XP_001489815 |
| Bos Taurus | 97% | 538 | XP_606794 |
| Homo sapiens | 97% | 538 | EAX02222 |
| Macaca mulatta | 97% | 538 | XP_001100407 |
| Rattus norvegicus | 93% | 538 | AAA41392 |
| Mus musculus | 96% | 538 | NP_034643 |
| Gallus gallus | 84% | 538 | NP_990363 |
| Coturnix japonica | 84% | 538 | BAF73401 |
| Xenopus laevis | 79% | 538 | CAA90517 |
| Cyprinus carpio | 71% | 538 | AAN52152 |
| Quail | Chick | Monkey | Human | Pig | Cattle | Horse | Fox | Dog | Rat | Mouse | Xenopus | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Chick | 0.006 | |||||||||||
| Monkey | 0.134 | 0.132 | ||||||||||
| Human | 0.137 | 0.134 | 0.002 | |||||||||
| Pig | 0.128 | 0.125 | 0.008 | 0.010 | ||||||||
| Cattle | 0.134 | 0.132 | 0.018 | 0.020 | 0.014 | |||||||
| Horse | 0.134 | 0.132 | 0.012 | 0.014 | 0.008 | 0.016 | ||||||
| Fox | 0.137 | 0.134 | 0.018 | 0.020 | 0.014 | 0.024 | 0.010 | |||||
| Dog | 0.137 | 0.134 | 0.018 | 0.020 | 0.014 | 0.024 | 0.010 | 0.000 | ||||
| Rat | 0.146 | 0.144 | 0.032 | 0.035 | 0.028 | 0.035 | 0.028 | 0.032 | 0.032 | |||
| Mouse | 0.148 | 0.146 | 0.037 | 0.039 | 0.032 | 0.039 | 0.032 | 0.037 | 0.037 | 0.004 | ||
| Xenopus | 0.165 | 0.157 | 0.203 | 0.205 | 0.198 | 0.205 | 0.203 | 0.205 | 0.205 | 0.208 | 0.210 | |
| Carp | 0.248 | 0.240 | 0.276 | 0.274 | 0.271 | 0.271 | 0.274 | 0.271 | 0.271 | 0.287 | 0.284 | 0.266 |
| Amino Acids | Number | Frequency (%) | Amino Acids | Number | Frequency (%) |
|---|---|---|---|---|---|
| Ala (A) | 31 | 5.8 | Lys (K) | 23 | 4.3 |
| Arg (R) | 33 | 6.1 | Met (M) | 22 | 4.1 |
| Asn (N) | 27 | 5.0 | Phe (F) | 20 | 3.7 |
| Asp (D) | 24 | 4.5 | Pro (P) | 39 | 7.2 |
| Cys (C) | 6 | 1.1 | Ser (S) | 40 | 7.4 |
| Gln (Q) | 14 | 2.6 | Thr (T) | 17 | 3.2 |
| Glu (E) | 49 | 9.1 | Trp (W) | 8 | 1.5 |
| Gly (G) | 40 | 7.4 | Tyr (Y) | 22 | 4.1 |
| His (H) | 8 | 1.5 | Val (V) | 41 | 7.6 |
| Ile (I) | 25 | 4.6 | Pyl (O) | 0 | 0 |
| Leu (L) | 49 | 9.1 | Sec (U) | 0 | 0 |
| Histophysiology | GAPDH (Ct1) | IGF1R (Ct2) | Relative Expression (2Ct1-Ct2) | Brain Expression-Normalized (Set to 1) |
|---|---|---|---|---|
| Liver | 22.123 | 23.060 | 0.522 ± 0.109 | 1.906 ± 0.398 |
| Spleen | 23.057 | 24.960 | 0.267 ± 0.085 | 0.976 ± 0.309 |
| Kidney | 22.873 | 23.520 | 0.639 ± 0.230 | 2.331 ± 0.840 |
| Lung | 19.745 | 22.150 | 0.189 ± 0.078 | 0.690 ± 0.286 |
| Stomach | 20.323 | 26.160 | 0.018 ± 0.003 | 0.064 ± 0.011 |
| Pancreas | 18.157 | 21.137 | 0.127 ± 0.107 | 0.463 ± 0.389 |
| Brain | 19.767 | 21.633 | 0.274 ± 0.155 | 1.000 ± 0.565 |
| Pituitary gland | 20.793 | 21.853 | 0.480 ± 0.154 | 1.750 ± 0.561 |
| Testis | 22.333 | 20.883 | 2.732 ± 0.486 | 9.971 ± 1.773 |
| Heart | 21.737 | 24.197 | 0.182 ± 0.052 | 0.663 ± 0.190 |
| Muscle | 20.443 | 21.273 | 0.563 ± 0.057 | 2.053 ± 0.208 |
| Histophysiology | GAPDH (Ct1) | IGF1R (Ct2) | Relative Expression (2Ct1-Ct2) | Muscle Expression in 4-Month-Old Male Foxes-Normalized (Set to 1) |
|---|---|---|---|---|
| 2-month-old male fox liver | 22.030 | 26.527 | 0.044 ± 0.017 | 0.043 ± 0.016 |
| 4-month-old male fox liver | 18.353 | 19.450 | 0.467 ± 0.095 | 0.451 ± 0.090 |
| 6-month-old male fox liver | 22.123 | 23.060 | 0.522 ± 0.127 | 0.504 ± 0.120 |
| 2-month-old male fox testis | 21.120 | 19.760 | 2.567 ± 0.264 | 2.480 ± 0.250 |
| 4-month-old male fox testis | 21.107 | 20.020 | 2.124 ± 0.323 | 2.052 ± 0.306 |
| 6-month-old male fox testis | 22.333 | 20.883 | 2.732 ± 0.225 | 2.640 ± 0.213 |
| 2-month-old male fox muscle | 20.220 | 22.993 | 0.146 ± 0.008 | 0.141 ± 0.007 |
| 4-month-old male fox muscle | 18.460 | 18.410 | 1.035 ± 0.264 | 1.000 ± 0.250 |
| 6-month-old male fox muscle | 20.443 | 21.273 | 0.563 ± 0.066 | 0.544 ± 0.063 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, W.; Fu, H.; Meng, X.; Sun, Y.; Ning, F.; Du, Z. Cloning and Spatiotemporal Expression Analysis of IGF1R Gene cDNA in Alopex lagopus (Arctic Fox). Life 2025, 15, 796. https://doi.org/10.3390/life15050796
Xu W, Fu H, Meng X, Sun Y, Ning F, Du Z. Cloning and Spatiotemporal Expression Analysis of IGF1R Gene cDNA in Alopex lagopus (Arctic Fox). Life. 2025; 15(5):796. https://doi.org/10.3390/life15050796
Chicago/Turabian StyleXu, Wei, Hualin Fu, Xiangyu Meng, Yiwen Sun, Fangyong Ning, and Zhiheng Du. 2025. "Cloning and Spatiotemporal Expression Analysis of IGF1R Gene cDNA in Alopex lagopus (Arctic Fox)" Life 15, no. 5: 796. https://doi.org/10.3390/life15050796
APA StyleXu, W., Fu, H., Meng, X., Sun, Y., Ning, F., & Du, Z. (2025). Cloning and Spatiotemporal Expression Analysis of IGF1R Gene cDNA in Alopex lagopus (Arctic Fox). Life, 15(5), 796. https://doi.org/10.3390/life15050796