

Impacts of Land Use on Soil Nitrogen-Cycling Microbial Communities: Insights from Community Structure, Functional Gene Abundance, and Network Complexity


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
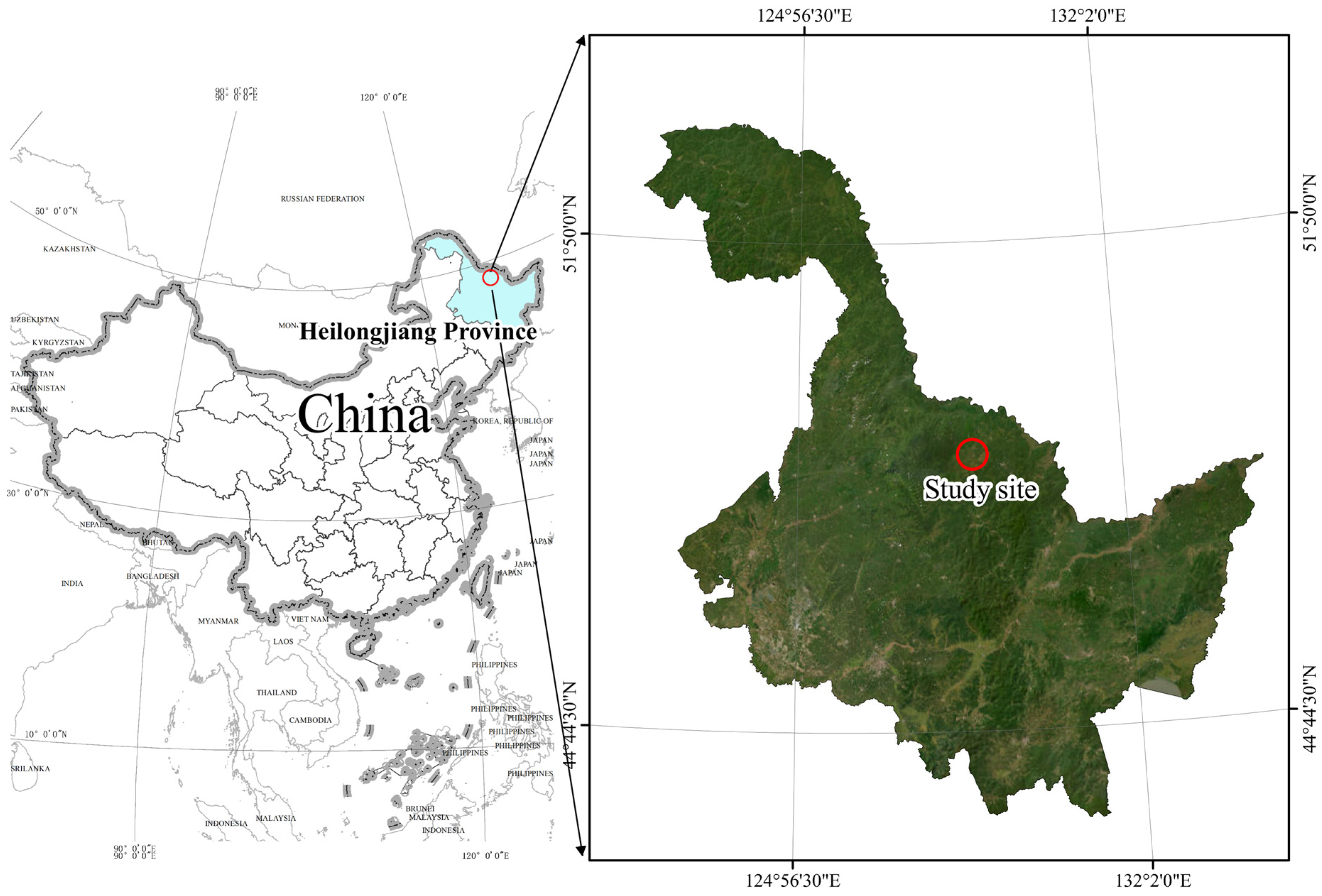
2.1. Site Description
2.2. Experimental Design
2.3. Sample Collection
2.4. Soil Chemical Analysis
2.5. Determination of Soil Enzyme Activity
2.6. Soil DNA Extraction and High-Throughput Assay
2.7. Statistical Analysis
3. Results
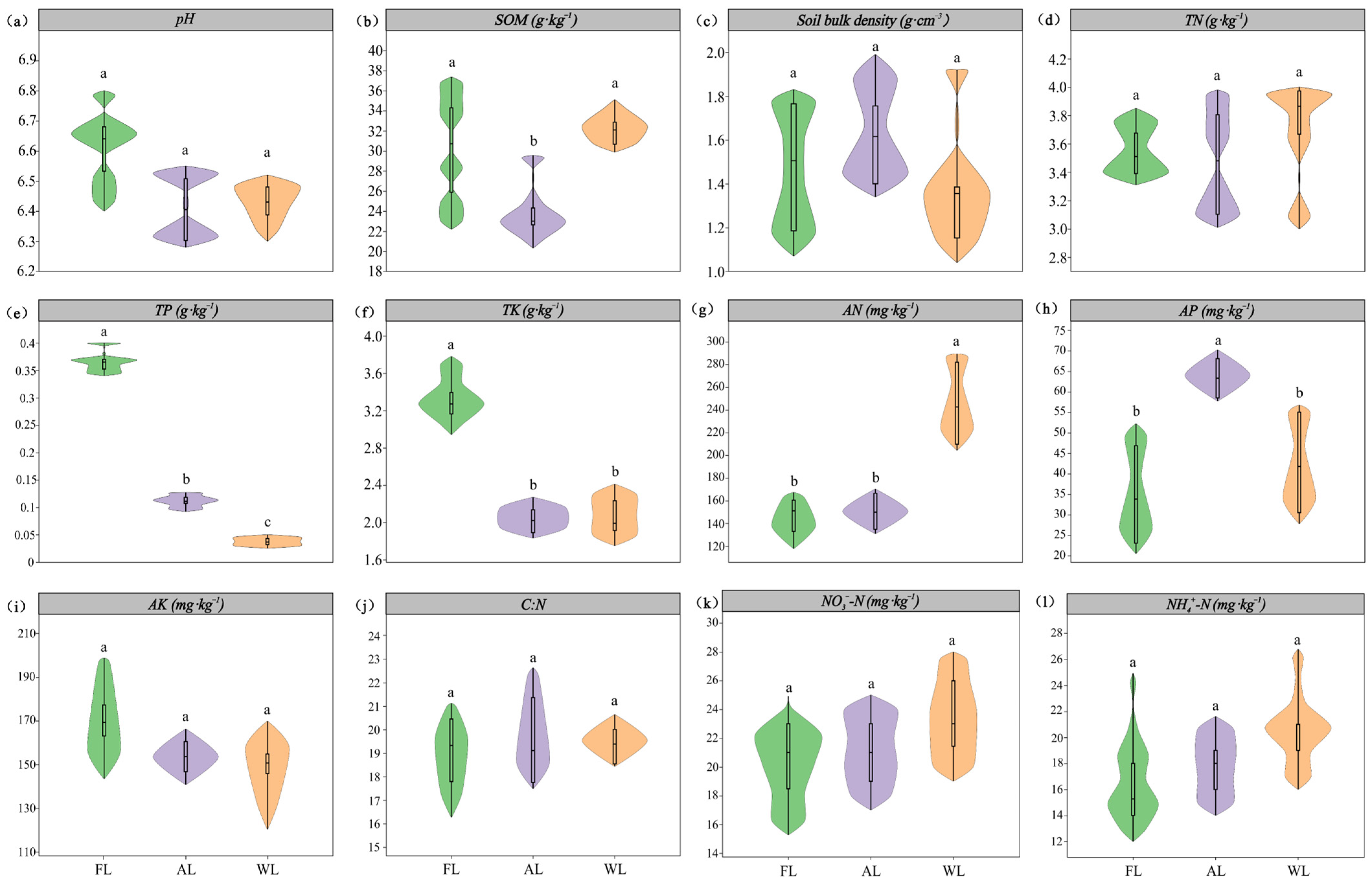
3.1. Effects of Soil Physicochemical Properties Under Land Use Patterns
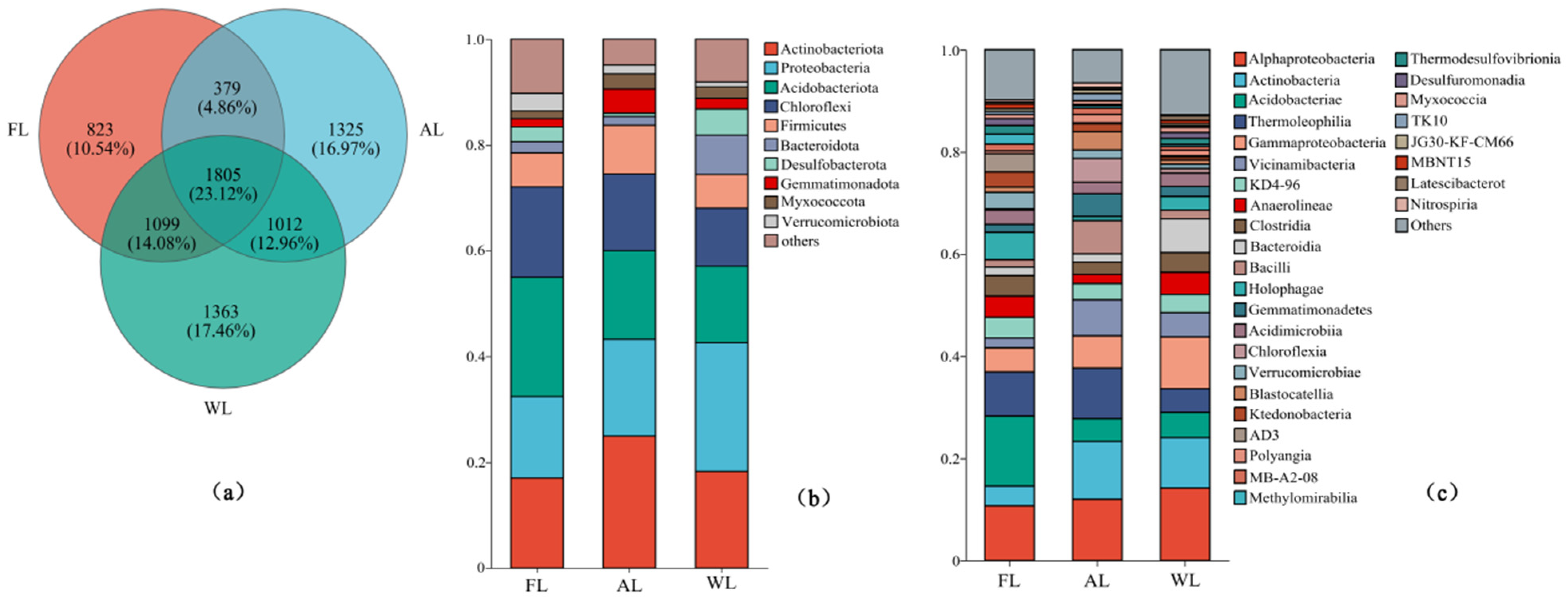
3.2. Composition of Soil Microbial Community
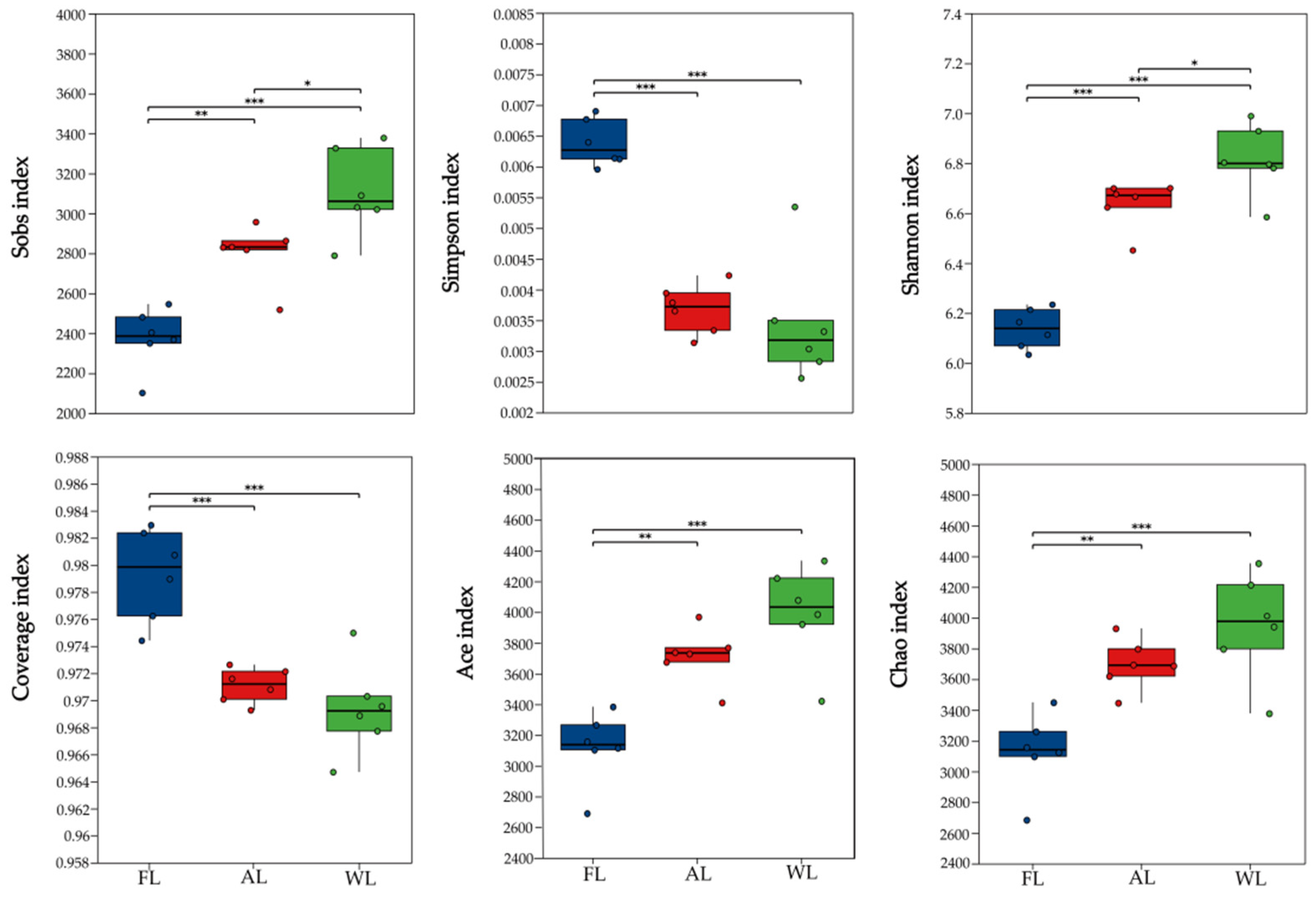
3.3. Analysis of Changes in Soil Microbial Alpha Diversity
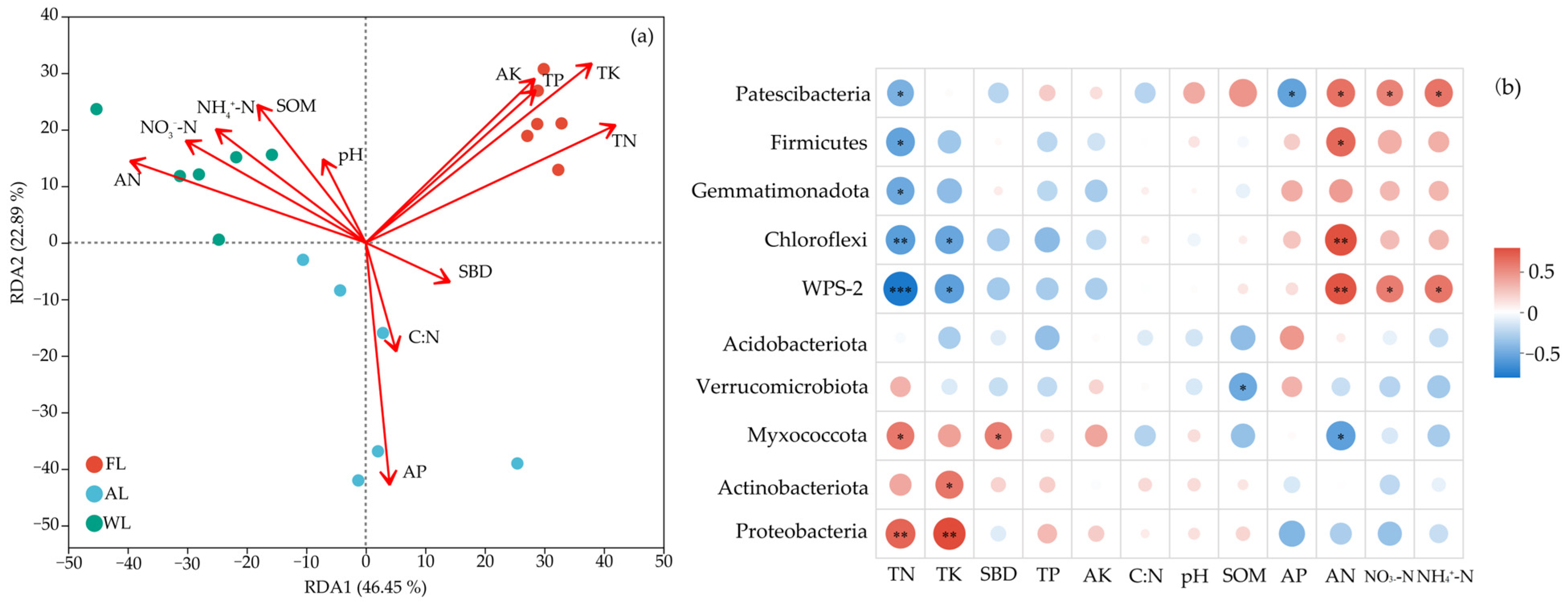
3.4. Correlation Analysis of Environmental Factors
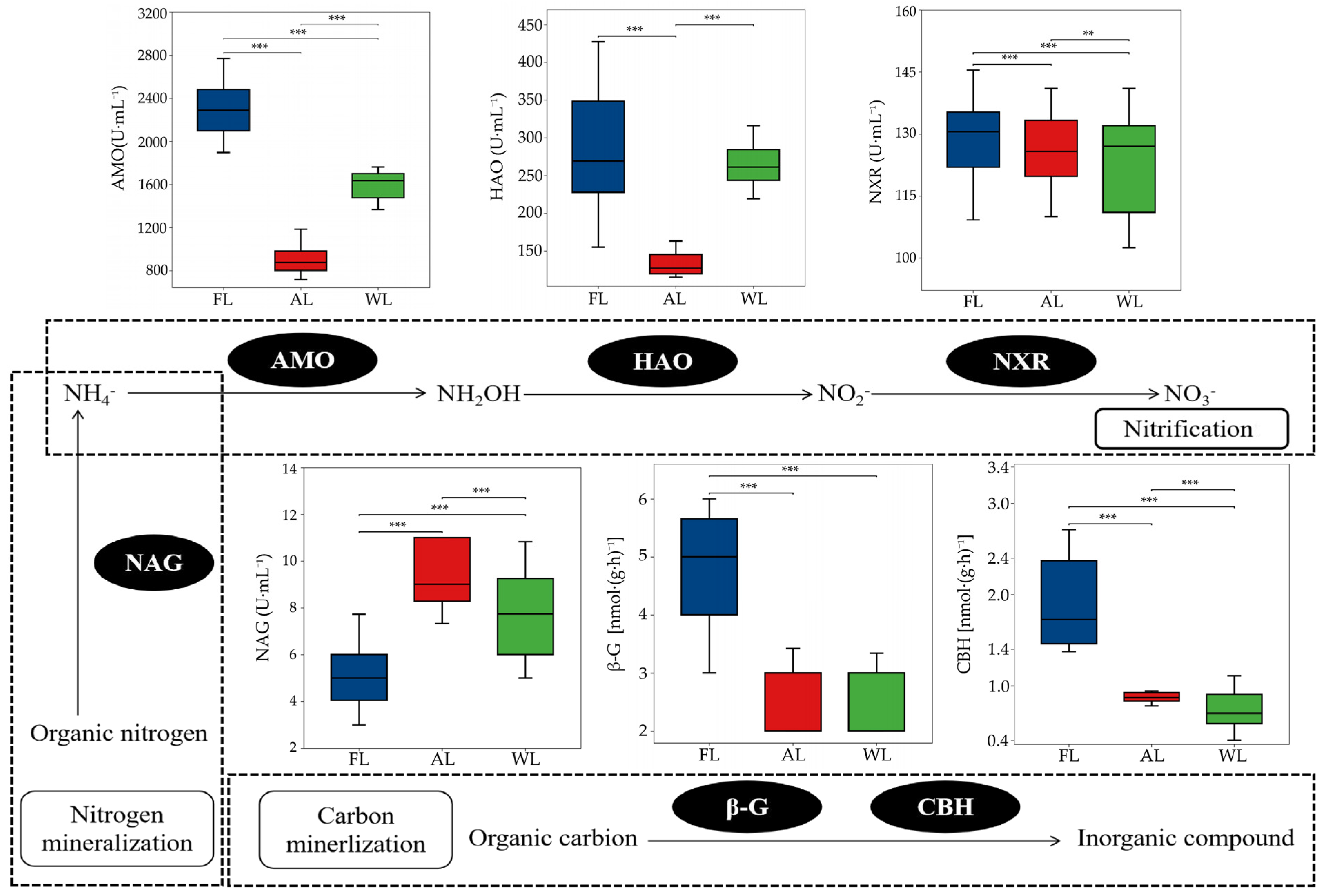
3.5. Effects of Land Use Types on Soil Nitrogen Mineralization, Nitrification, and C-N Cycle-Related Enzymes Activity
3.6. Effects of Major Control Factors on Soil Microbial Properties
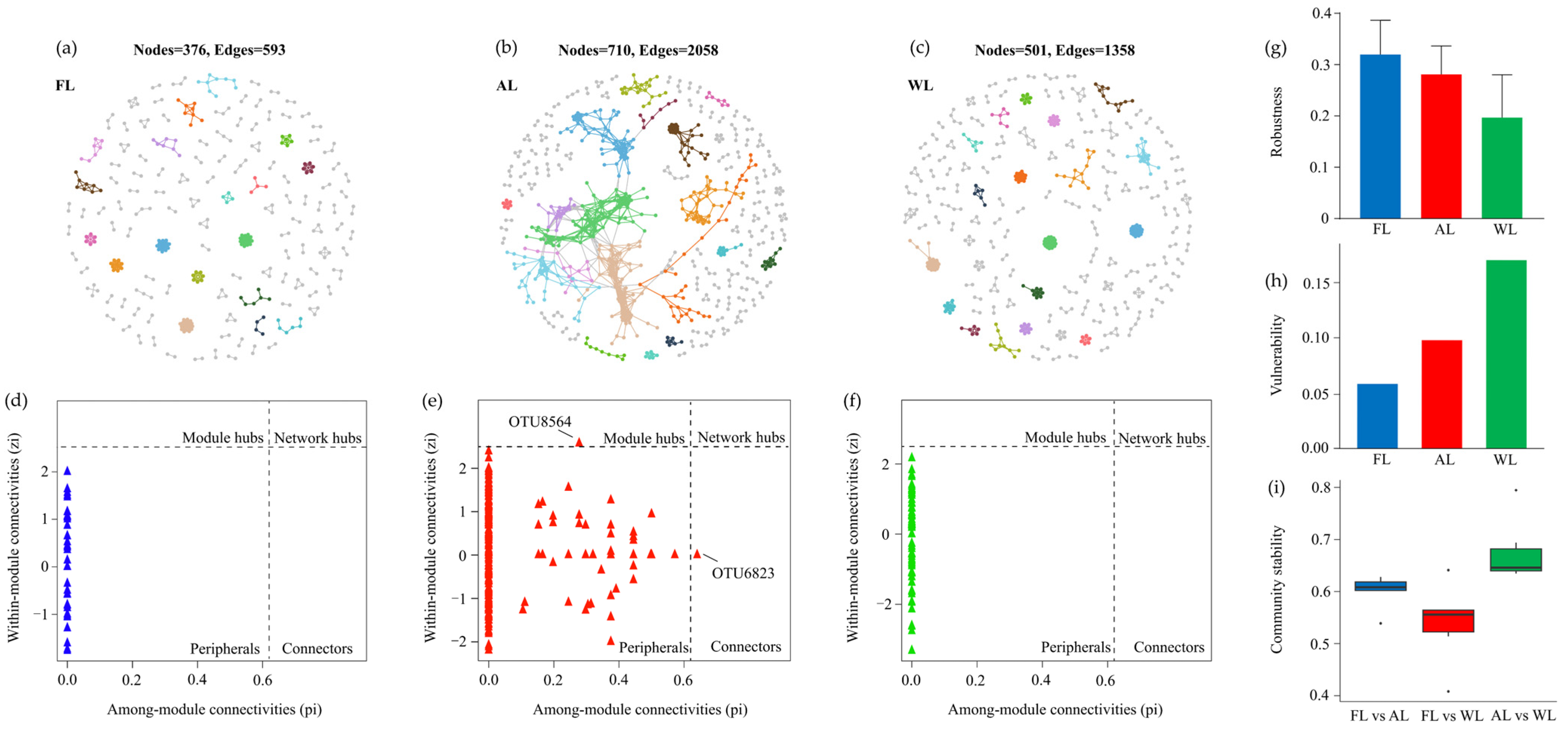
3.7. Effects of Land Use Types on Microbial Co-Occurrence Patterns
4. Discussion
4.1. Impacts of Land-Use Patterns on Soil Bacterial Community Structure
4.2. Effect of Land Use on Soil Bacterial Community Diversity
4.3. Effects of Land Use Patterns on Soil Nitrogen Processes and Microbial Properties
4.4. Land-Use Types Impact Microbial Network Complexity, Connectivity, Robustness, and Stability
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, C.T.; Wang, G.X.; Wang, Y.; Rafique, R.; Ma, L.; Hu, L.; Luo, Y.Q. Fire alters vegetation and soil microbial community in alpine meadow. Land. Degrad. Dev. 2016, 27, 1379–1390. [Google Scholar] [CrossRef]
- Fierer, N.; Leff, J.W.; Adams, B.J.; Nielsen, U.N.; Bates, S.T.; Lauber, C.L.; Owens, S.; Gilbert, J.A.; Wall, D.H.; Caporaso, J.G. Cross-biome metagenomic analyses of soil microbial communities and their functional attributes. Proc. Natl. Acad. Sci. USA 2012, 109, 21390–21395. [Google Scholar] [CrossRef]
- Zhou, H.; Wang, Z.Y.; Li, C.; Yuan, H.W.; Hu, L.; Zeng, P.; Yang, W.T.; Liao, B.H.; Gu, J.F. Straw removal reduces Cd availability and rice Cd accumulation in Cd-contaminated paddy soil: Cd fraction, soil microorganism structure and porewater DOC and Cd. J. Hazard. Mater. 2024, 476, 135189. [Google Scholar] [CrossRef] [PubMed]
- Hao, G.; Chen, X.Q.; Du, Z.B.; Yang, N.; Li, M.Y.; Gao, Y.; Liang, J.A.; Chen, L.; Li, H.Y. Phylogenetic and functional diversity consistently increase engineered ecosystem functioning under nitrogen enrichment: The example of green roofs. Ecol. Eng. 2025, 212, 107499. [Google Scholar] [CrossRef]
- Lu, M.; Xiao, Z. Wetland ecology and climate change: Addressing global challenges with countermeasures. Adv. Resour. Res. 2024, 4, 67–88. [Google Scholar]
- Mao, J.L.; Hu, G.J.; Deng, W.; Zhao, M.; Li, J.B. Industrial wastewater treatment using floating wetlands: A review. Environ. Sci. Pollu Res. 2024, 31, 5043–5070. [Google Scholar] [CrossRef]
- Szostek, M.; Szpunar-Krok, E.; Pawlak, R.; Stanek-Tarkowska, J.; Ilek, A. Effect of different tillage systems on soil organic carbon and enzymatic activity. Agronomy 2022, 12, 208. [Google Scholar] [CrossRef]
- Jing, Y.L.; Zhang, Y.H.; Han, I.; Wang, P.; Mei, Q.W.; Huang, Y.J. Effects of different straw biochars on soil organic carbon, nitrogen, available phosphorus, and enzyme activity in paddy soil. Sci. Rep. 2020, 10, 8837. [Google Scholar] [CrossRef]
- Zhang, H.X.; Huang, Y.M.; Shen, J.K.; Xu, F.J.; Hou, H.Y.; Xie, C.J.; Wang, B.R.; An, S.S. Mechanism of polyethylene and biodegradable microplastic aging effects on soil organic carbon fractions in different land-use types. Sci. Total Environ. 2024, 912, 168961. [Google Scholar] [CrossRef]
- Liu, C.; Hu, S.G.; Wu, S.; Song, J.R.; Li, H.Y. County-level land use carbon emissions in China: Spatiotemporal patterns and impact factors. Sustain. Cities Soc. 2024, 105, 105304. [Google Scholar] [CrossRef]
- Feng, J.; Wen, Y.L.; Zhang, H.Y.; Duan, W.; Hao, H.G. Wetland restoration, household income, and livelihood structure of farmers. Front. Sustain. Food Syst. 2024, 8, 1256115. [Google Scholar] [CrossRef]
- Pedrinho, A.; Mendes, L.W.; de Araujo Pereira, A.P.; Araujo, A.S.F.; Vaishnav, A.; Karpouzas, D.G.; Singh, B.K. Soil microbial diversity plays an important role in resisting and restoring degraded ecosystems. Plant Soil 2024, 500, 325–349. [Google Scholar] [CrossRef]
- Wang, C.W.; Liu, Z.K.; Yu, W.Y.; Ye, X.H.; Ma, L.N.; Wang, R.Z.; Huang, Z.Y.; Liu, G.F. Grassland degradation has stronger effects on soil fungal community than bacterial community across the Semi-Arid region of northern China. Plants 2022, 24, 3488. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wang, X.T.; Wang, J.; Liao, L.R.; Lei, S.L.; Liu, G.B.; Zhang, C. Alpine meadow degradation depresses soil nitrogen fixation by regulating plant functional groups and diazotrophic community composition. Plant Soil 2022, 473, 319–335. [Google Scholar] [CrossRef]
- Zhang, M.X.; Hu, J.P.; Li, J.L.; Che, Z.; Li, L.; Lü, Z.L.; Dong, W.Q.; Zhang, J.Q.; Yao, T.; Duan, T.Y.; et al. Responses of legume-associated rhizobacterial communities to plant diversity and soil traits in alpine grassland. Land. Degrad. Dev. 2024, 2, 441–453. [Google Scholar] [CrossRef]
- Pechlivanis, N.; Karakatsoulis, G.; Kyritsis, K.; Tsagiopoulou, M.; Sgardelis, S.; Kappas, I.; Psomopoulos, F. Microbial co-occurrence network demonstrates spatial and climatic trends for global soil diversity. Sci. Data 2024, 11, 672. [Google Scholar] [CrossRef]
- Yuan, M.M.; Guo, X.; Wu, L.W.; Zhang, Y.; Xiao, N.J.; Ning, D.L.; Shi, Z.; Zhou, X.S.; Wu, L.Y.; Yang, Y.F.; et al. Climate warming enhances microbial network complexity and stability. Nat. Clim. Change 2021, 11, 343–348. [Google Scholar] [CrossRef]
- Wagg, C.; Schlaeppi, K.; Banerjee, S.; Kuramae, E.E.; van der Heijden, M.G.A. Fungal-bacterial diversity and microbiome complexity predict ecosystem functioning. Nat Commun. 2019, 10, 4841. [Google Scholar] [CrossRef]
- Zhou, L.; Liu, S.; Lin, D.; Hu, H.W.; He, J.Z. Divergent changes in diversity and network complexity across different trophic-level organisms drive soil multifunctionality of fire-impacted subtropical forests. For Ecosyst. 2024, 11, 100227. [Google Scholar] [CrossRef]
- Liu, X.; Xiong, Z.; Ouyang, L.; He, G.; Liu, W.; Cai, M. Macrohabitat and microhabitat mediate the relationships between wetland multifaceted biodiversity and multifunctionality. CATENA 2024, 241, 108023. [Google Scholar] [CrossRef]
- Ma, Y.; Yang, K.; Yu, H.; Tan, W.; Gao, Y.; Lv, B. Effects and mechanism of microplastics on organic carbon and nitrogen cycling in agricultural soil: A review. Soil Use Manag. 2024, 40, e12971. [Google Scholar] [CrossRef]
- Stein, L.Y.; Klotz, M.G. The nitrogen cycle. Curr. Biol. 2016, 26, 94–98. [Google Scholar] [CrossRef]
- Coskun, D.; Britto, D.T.; Shi, W.; Kronzucker, H.J. How plant root exudates shape the nitrogen cycle. Trends Plant Sci. 2017, 22, 661–673. [Google Scholar] [CrossRef]
- Li, Z.; Liu, X.W.; Zhang, M.H.; Xing, F. Plant diversity and fungal richness regulate the changes in soil multifunctionality in a semi-arid grassland. Biology 2022, 6, 870. [Google Scholar] [CrossRef] [PubMed]
- Rashti, M.R.; Nelson, P.N.; Lan, Z.; Su, N.; Esfandbod, M.; Liu, X.Y.; Goloran, J.; Zhang, H.Z.; Chen, C.R. Sugarcane cultivation altered soil nitrogen cycling microbial processes and decreased nitrogen bioavailability in tropical Australia. J. Soils Sediment 2024, 24, 946–955. [Google Scholar] [CrossRef]
- Zaki, M.; Khalil, H.P.S.A.; Sabaruddin, F.A.; Bairwan, R.D.; Oyekanmi, A.A.; Alfatah, T.; Danish, M.; Mistar, E.M.; Abdullah, C.K. Microbial treatment for nanocellulose extraction from marine algae and its applications as sustainable functional material. Bioresour. Technol. Rep. 2021, 16, 100811. [Google Scholar] [CrossRef]
- Alsharif, W.; Saad, M.M.; Hirt, H. Desert microbes for boosting sustainable agriculture in extreme environments. Front. Microbiol. 2020, 11, 1666. [Google Scholar] [CrossRef]
- Ouyang, Y.; Evans, S.E.; Friesen, M.L.; Tiemann, L.K. Effect of nitrogen fertilization on the abundance of nitrogen cycling genes in agricultural soils: A meta-analysis of field studies. Soil Biol. Biochem. 2018, 127, 71–78. [Google Scholar] [CrossRef]
- Pupin, B.; Rangel, D.E.N.; Nahas, E. Evaluation of Soil Microbial and Enzymatic Activity in Ecosystems in a Coastal Region of Brazil. Wetlands 2024, 44, 64. [Google Scholar] [CrossRef]
- Luo, G.W.; Friman, V.P.; Chen, H.; Liu, M.Q.; Wang, M.; Guo, S.W.; Liang, N.; Shen, Q.N. Long-term fertilization regimes drive the abundance and composition of N-cycling-related prokaryotic groups via soil particle-size differentiation. Soil Biol. Biochem. 2018, 116, 213–223. [Google Scholar] [CrossRef]
- Kowalchuk, G.A.; Stephen, J.R. Ammonia-oxidizing bacteria: A model for molecular microbial ecology. Annu. Rev. Microbiol. 2001, 55, 485–529. [Google Scholar] [CrossRef] [PubMed]
- Hussain, M.Z.; Bhardwaj, A.K.; Basso, B.; Robertson, G.P.; Hamilton, S.K. Nitrate leaching from continuous corn, perennial grasses, and poplar in the US Midwest. J. Environ. Qual. 2019, 48, 1849–1855. [Google Scholar] [CrossRef]
- Bouabid, R.; Soudi, B.; Badraoui, M. Nitrogen Dynamics and Management in Rainfed Drylands Issues and Challenges. In Soil and Fertilizers; CRC Press: Boca Raton, FL, USA, 2020; pp. 285–315. [Google Scholar]
- Barton, L.; Hoyle, F.C.; Grace, P.R.; Schwenke, G.D.; Scanlan, C.A.; Armstrong, R.D.; Bell, M.J. Soil nitrogen supply and N fertilizer losses from Australian dryland grain cropping systems. Adv. Agron. 2022, 174, 1–52. [Google Scholar]
- Li, S.L.; Liu, X.; Yue, F.J.; Yan, Z.; Wang, T.; Li, S.; Liu, C.Q. Nitrogen dynamics in the Critical Zones of China. Prog. Phys. Geog. 2022, 46, 869–888. [Google Scholar] [CrossRef]
- Jin, X.; Ma, J.; Cai, T.; Sun, X. Non-use value assessment for wetland ecosystem service of Hongxing National Nature Reserve in northeast China. J. For. Res. 2016, 27, 1435–1442. [Google Scholar] [CrossRef]
- IGAC. Soil Classification in Sucre Municipalities. Inst. Geogr. Agric. Classif. Rep. 2016, 34, 112–127. [Google Scholar]
- Ding, J.N.; Xu, N. Variations of soil bacterial microbial community and functional structure under different land-uses. Rev. Bras. Cienc. Solo 2022, 46, e0220090. [Google Scholar] [CrossRef]
- Patel, A.; Kumar, A.; Gupta, R.; Singh, S. High-throughput sequencing reveals SOC dynamics in soil microbial communities. Appl. Soil Ecol. 2023, 182, 104519. [Google Scholar]
- Ma, Q.; Qu, Y.Y.; Shen, W.L.; Zhang, Z.J.; Wang, J.W.; Liu, Z.Y.; Li, D.X.; Li, H.J.; Zhou, J.T. Bacterial community compositions of coking wastewater treatment plants in steel industry revealed by Illumina high-throughput sequencing. Bioresour. Technol. 2015, 179, 436–443. [Google Scholar] [CrossRef]
- Sanchez, D.; Armas, C.; Pueyo, J.J.; Trasar-Cepeda, C.; Hernández, T. Soil total nitrogen and its relation to land use changes. Geoderma 2023, 411, 115699. [Google Scholar]
- Yuan, J.; Li, X.; Wang, H.; Zhang, Y.; Chen, L. Effects of land use on soil total phosphorus and potassium. Soil Syst. 2022, 6, 98–109. [Google Scholar]
- Xie, Z.; Zhang, W.; Li, L. Optimization of the Ammonium Acetate Extraction Method for Available Potassium in Soils. Soil Sci. Plant Nutr. 2020, 66, 561–567. [Google Scholar]
- Corwin, D.L.; Plant, R.E. Applications of apparent soil electrical conductivity in precision agriculture. Comput. Electron. Agric. 2005, 46, 1–10. [Google Scholar] [CrossRef]
- Lu, R.K. Soil and Agro-Chemical Analytical Methods; China Agricultural Science and Technology Press: Beijing, China, 1999. [Google Scholar]
- Zhang, L.L.; Wu, Z.J.; Chen, L.J.; Li, D.P.; Ma, X.Z.; Shi, Y.F. A microplate fluorimetric assay for sacchariase activity measurement. Spectrosc. Spect. Anal. 2009, 29, 1341–1354. [Google Scholar]
- Gao, G.F.; Li, P.F.; Zhong, J.X.; Shen, Z.J.; Chen, J.; Li, Y.T.; Isabwe, A.; Zhu, X.Y.; Ding, Q.S.; Zhang, S.; et al. Spartina alterniflora invasion alters soil bacterial communities and enhances soil N2O emissions by stimulating soil denitrification in mangrove wetland. Sci. Total Environ. 2019, 653, 231–240. [Google Scholar] [CrossRef]
- Zeng, Z.B.; Zhu, B.; Zhu, X.M.; Liu, X.F.; Wang, Y. Effects of fertilization on N2O emission and denitrification in purple soil during summer maize season in the Sichuan basin. Acta Pedol. Sin. 2013, 50, 130–137. [Google Scholar]
- Sinsabaugh, R.L.; Reynolds, H.; Long, T.M. Rapid assay for amidohydrolase (urease) activity in environmental samples. Soil Biol. Biochem. 2000, 32, 2095–2097. [Google Scholar] [CrossRef]
- Edwards, K.A.; McCulloch, J.; Kershaw, G.P. Soil microbial and nutrient dynamics in a wet Arctic sedge meadow in late winter and early spring. Soil Biol. Biochem. 2006, 38, 2843–2851. [Google Scholar] [CrossRef]
- Yang, Y.L.; Wu, F.Z.; He, Z.H.; Xu, Z.F.; Liu, Y.; Yang, W.Q.; Tan, B. Effects of snow pack removal on soil microbial biomass carbon and nitrogen and the number of soil culturable microorganisms during wintertime in alpine Abies faxoniana forest of western Sichuan, Southwest China. Chin. J. Appl. Ecol. 2012, 23, 1809–1816. [Google Scholar]
- Ding, J.N. Soil nitrogen transformation and functional microbial abundance in an agricultural soil amended with biochar. Rev. Bras. Cienc. Solo 2023, 47, e0220156. [Google Scholar] [CrossRef]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Kassambara, A. ggpubr: ‘ggplot2’ Based Publication Ready Plots; R package Version 0.4.0, 2020. Available online: https://cran.r-project.org/package=ggpubr (accessed on 16 November 2024).
- Chen, H.; Boutros, P.C. VennDiagram: A Package for the Generation of Highly-Customizable Venn and Euler Diagrams in R. BMC Bioinform. 2011, 12, 35. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; O’hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.; Wagner, H. Vegan: Community Ecology Package; R Package Version 2.5-7, 2020. Available online: https://cran.r-project.org/package=vegan (accessed on 16 November 2024).
- Csardi, G.; Nepusz, T. The igraph Software Package for Complex Network Research. Inter J. Complex Syst. 2006, 1695, 1–9. Available online: https://igraph.org (accessed on 16 November 2024).
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Harrell, F.E. Hmisc: Harrell Miscellaneous; R Package Version 4.4-0, 2020. Available online: https://cran.r-project.org/package=Hmisc (accessed on 16 November 2024).
- Chen, X.H.; Song, Y.X.; Ling, C.; Shen, Y.; Zhan, X.H.; Xing, B.S. Fate of emerging antibiotics in soil-plant systems: A case on fluoroquinolones. Sci. Total Environ. 2024, 951, 175487. [Google Scholar] [CrossRef] [PubMed]
- Zhu, P.C.; Yang, S.R.; Wu, Y.X.; Ru, Y.N.; Yu, X.N.; Wang, L.S.; Guo, W.H. Shifts in soil microbial community composition, function, and co-occurrence network of Phragmites australis in the Yellow River Delta. Front. Microbiol. 2022, 13, 858125. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.Y.; Liu, Z.D.; Song, Y.Y.; Wang, X.W.; Yuan, J.B.; Li, M.T.; Lou, Y.J.; Gao, Z.L.; Song, C.C. Soil microbial functional diversity is primarily affected by soil nitrogen, salinity and alkalinity in wetland ecosystem. Appl. Soil Ecol. 2024, 199, 105407. [Google Scholar] [CrossRef]
- Dincă, L.C.; Grenni, P.; Onet, C.; Onet, A. Fertilization and soil microbial community: A review. Appl. Sci. 2022, 12, 1198. [Google Scholar] [CrossRef]
- Zheng, X.X.; Cai, R.H.; Yao, H.W.; Zhuo, X.C.; He, C.; Zheng, Q.; Shi, Q.; Jiao, N.Z. Experimental insight into the enigmatic persistence of marine refractory dissolved organic matter. Environ Sci Technol. 2022, 56, 17420–17429. [Google Scholar] [CrossRef]
- Xu, L.; Zhu, B.J.; Li, C.N.; Yao, M.J.; Zhang, B.C.; Li, X.Z. Development of biological soil crust prompts convergent succession of prokaryotic communities. CATENA 2020, 187, 104360. [Google Scholar] [CrossRef]
- Ren, H.Y.; Wang, G.X.; Ding, W.C.; Li, H.; Shen, X.; Shen, D.B.; Jiang, X.; Qadeer, A. Response of dissolved organic matter (DOM) and microbial community to submerged macrophytes restoration in lakes: A review. Environ. Res. 2023, 231, 116185. [Google Scholar] [CrossRef]
- Yousaf, A.; Khalid, N.; Aqeel, M.; Noman, A.; Naeem, N.; Sarfraz, W.; Ejaz, U.; Qaiser, Z.; Khalid, A. Nitrogen dynamics in wetland systems and its impact on biodiversity. Nitrogen 2021, 2, 196–217. [Google Scholar] [CrossRef]
- Li, Q.C.; Wang, L.L.; Fu, Y.; Lin, D.G.; Hou, M.R.; Li, X.D.; Hu, D.D.; Wang, Z.H. Transformation of soil organic matter subjected to environmental disturbance and preservation of organic matter bound to soil minerals: A review. J. Soil Sediment 2023, 23, 1485–1500. [Google Scholar] [CrossRef]
- Crabtree, R.; Potter, C.; Mullen, R.; Sheldon, J.; Huang, S.; Harmsen, J.; Rodman, A.; Jean, C. A modeling and spatio-temporal analysis framework for monitoring environmental change using NPP as an ecosystem indicator. Remote Sens. Environ. 2009, 113, 1486–1496. [Google Scholar] [CrossRef]
- Uddin, M.J.; Sherrell, J.; Emami, A.; Khaleghian, M. Application of Artificial Intelligence and Sensor Fusion for Soil Organic Matter Prediction. Sensors 2024, 24, 2357. [Google Scholar] [CrossRef]
- Wang, J.Q.; Peñuelas, J.; Shi, X.Z.; Liu, Y.; Delgado Baquerizo, M.; Mao, J.Y.; Zhang, G.Y.; Li, C.; Pan, G. Soil microbial biodiversity supports the delivery of multiple ecosystem functions under elevated CO2 and warming. Commun. Earth Environ. 2024, 5, 615. [Google Scholar] [CrossRef]
- Furey, P.C.; Liess, A.; Lee, S. Substratum-Associated Microbiota. Water Environ. Res. 2017, 89, 1634–1675. [Google Scholar] [CrossRef]
- Feng, W.L.; Yang, J.L.; Xu, L.G.; Zhang, G.L. The spatial variations and driving factors of C, N, P stoichiometric characteristics of plant and soil in the terrestrial ecosystem. Sci. Total Environ. 2024, 951, 175543. [Google Scholar] [CrossRef]
- Hunter, P.J.; Teakle, G.R.; Bending, G.D. Root traits and microbial community interactions in relation to phosphorus availability and acquisition, with particular reference to Brassica. Front. Plant Sci. 2014, 5, 27. [Google Scholar] [CrossRef]
- Di, D.L.; Wang, S.K.; Chen, G.C.; Wang, Q.; Zhang, J.W.; Niu, X.Y.; Huang, D.Z. NH4+-N and Low Ratios of NH4+-N/NO3−-N Promote the Remediation Efficiency of Salix linearistipularis in Cd-and Pb-Contaminated Soil. Forests 2024, 15, 419. [Google Scholar] [CrossRef]
- Patel, H.V.; Zhao, R.; Eramo, A.; Blanc, S.; Fahrenfeld, N.L.; Brazil, B.; Luster-Teasley, S. Ammonium oxidation from concentrated synthetic wastewater and landfill leachate using partial nitritation in sequencing batch reactor. Water Environ. Res. 2024, 96, e11075. [Google Scholar] [CrossRef]
- Singh, H.; Halder, N.; Singh, B.; Singh, J.; Sharma, S.; Shacham-Diamand, Y. Smart farming revolution: Portable and real-time soil nitrogen and phosphorus monitoring for sustainable agriculture. Sensors 2023, 13, 5914. [Google Scholar] [CrossRef] [PubMed]
- Grzyb, A.; Wolna-Maruwka, A.; Niewiadomska, A. The significance of microbial transformation of nitrogen compounds in the light of integrated crop management. Agronomy 2021, 7, 1415. [Google Scholar] [CrossRef]
- Albright, M.B.; Timalsina, B.; Martiny, J.B.; Dunbar, J. Comparative genomics of nitrogen cycling pathways in bacteria and archaea. Microb. Ecol. 2019, 77, 597–606. [Google Scholar] [CrossRef] [PubMed]
- Ward, L.M.; Cardona, T.; Holland-Moritz, H. Evolutionary implications of anoxygenic phototrophy in the bacterial phylum Candidatus Eremiobacterota (WPS-2). Front. Microbiol. 2019, 10, 1658. [Google Scholar] [CrossRef]
- Meena, M.; Yadav, G.; Sonigra, P.; Nagda, A.; Mehta, T.; Swapnil, P.; Marwal, A.; Kumar, S. Multifarious responses of forest soil microbial community toward climate change. Microb. Ecol. 2023, 86, 49–74. [Google Scholar] [CrossRef]
- Wang, W.M.; Wang, Z.; Ling, H.B.; Zheng, X.; Chen, C.Q.; Wang, J.P.; Cheng, Z.B. Effects of reclaimed water irrigation on soil properties and the composition and diversity of microbial communities in northwest China. Sustainability 2025, 17, 308. [Google Scholar] [CrossRef]
- Farooq, M.S.; Wang, X.; Uzair, M.; Fatima, H.; Fiaz, S.; Maqbool, Z.; Rehman, O.U.; Yousuf, M.; Khan, M.R. Recent trends in nitrogen cycle and eco-efficient nitrogen management strategies in aerobic rice system. Front. Plant Sci. 2022, 13, 960641. [Google Scholar] [CrossRef]
- Weng, X.H.; Wang, M.Y.; Sui, X.; Frey, B.; Liu, Y.N.; Zhang, R.T.; Ni, H.W.; Li, M.H. High ammonium addition changes the diversity and structure of bacterial communities in temperate wetland soils of northeastern china. Microorganisms 2023, 11, 2033. [Google Scholar] [CrossRef]
- Giagnoni, L.; Renella, G. Effects of biochar on the C use efficiency of soil microbial communities: Components and mechanisms. Environments 2022, 11, 138. [Google Scholar] [CrossRef]
- Chen, X.; Han, X.Z.; Zou, W.X.; Yan, Y.; Lu, X.C. Mechanism of biochar decomposition mediated by bacteria shifts the molecular structure of soil organic matter to high molecular aromatic components. Pedosphere 2024, 12, 7. [Google Scholar] [CrossRef]
- Mugnai, G.; Pinchuk, I.; Borruso, L.; Tiziani, R.; Sannino, C.; Canini, F.; Turchetti, B.; Mimmo, T.; Zucconi, L.; Buzzini, P. The hidden network of biocrust successional stages in the High Arctic: Revealing abiotic and biotic factors shaping microbial and metazoan communities. Sci. Total Environ. 2024, 926, 171786. [Google Scholar] [CrossRef]
- Liu, C.; Li, L.; Zhi, Y.; Chen, J.Y.; Zuo, Q.Y.; He, Q. Molecular insight into the vertical migration and degradation of dissolved organic matter in riparian soil profiles. Environ. Res. 2024, 245, 118013. [Google Scholar] [CrossRef]
- Paz-Ares, J.; Puga, M.I.; Rojas-Triana, M.; Martinez-Hevia, I.; Diaz, S.; Poza-Carrión, C.; Miñambres, M.; Leyva, A. Plant adaptation to low phosphorus availability: Core signaling, crosstalks, and applied implications. Mol. Plant. 2022, 15, 104–124. [Google Scholar] [CrossRef] [PubMed]
- Yu, L.H.; Li, D.; Zhang, Y.F.; Wang, Y.F.; Yao, Q.; Yang, K.J. An optimal combined slow-release nitrogen fertilizer and urea can enhance the decomposition rate of straw and the yield of maize by improving soil bacterial community and structure under full straw returning system. Front. Microbiol. 2024, 15, 1358582. [Google Scholar] [CrossRef]
- Hu, Y.; Kou, T.L.; Cong, M.F.; Jia, Y.B.; Yan, H.; Huang, X.Y.; Yang, Z.L.; An, S.S.; Jia, H. Grassland degradation-induced soil organic carbon loss associated with micro-food web simplification. Soil Biol. Biochem. 2025, 201, 109659. [Google Scholar] [CrossRef]
- Tian, Y.Y.; Li, D.W.; Wang, Y.T.; Zhao, Q.Q.; Li, Z.P.; Jing, R.; Zhou, X.G. Effect of subsurface drainage combined with biochar on the bacterial community composition of coastal saline soil. Water 2023, 15, 2701. [Google Scholar] [CrossRef]
- Yahaya, S.M.; Mahmud, A.A.; Abdullahi, M.; Haruna, A. Recent advances in the chemistry of nitrogen, phosphorus and potassium as fertilizers in soil: A review. Pedosphere 2023, 33, 385–406. [Google Scholar] [CrossRef]
- Conradie, T.A.; Jacobs, K. Distribution patterns of Acidobacteriota in different fynbos soils. PLoS ONE 2021, 16, e0248913. [Google Scholar] [CrossRef]
- Elhalis, H. Expanding the horizons of saccharomyces cerevisiae: Nutrition, oenology, and bioethanol production. Sustainability 2024, 24, 11151. [Google Scholar] [CrossRef]
- Liu, G.H.; Gu, Z.; Liu, X.D.; Li, B.Y. Microenvironment heterogeneity affected by anthropogenic wildfire-perturbed soil mediates bacterial community in Pinus tabulaeformis forests. Front. Microbiol. 2024, 15, 1415726. [Google Scholar] [CrossRef]
- Waqas, M.; Liebmann, P.; Vogel, C.; Varsadiya, M.; Wang, H.T.; Shibistova, O.; Novák, M.; Urich, T.; Guggenberger, G.; Bárta, J. Different hydrological conditions after permafrost thaw result in distinct microbial community compositions. bioRxiv 2024, 12, 630578. [Google Scholar]
- Jagadesh, M.; Dash, M.; Kumari, A.; Singh, S.K.; Verma, K.K.; Kumar, P.; Bhatt, R.; Sharma, S.K. Revealing the hidden world of soil microbes: Metagenomic insights into plant, bacteria, and fungi interactions for sustainable agriculture and ecosystem restoration. Microbiol. Res. 2024, 285, 127764. [Google Scholar] [CrossRef] [PubMed]
- Islam, W.; Noman, A.; Naveed, H.; Huang, Z.Q.; Chen, H.Y.H. Role of environmental factors in shaping the soil microbiome. Environ. Sci. Pollut. Res. 2020, 27, 41225–41247. [Google Scholar] [CrossRef] [PubMed]
- Chamoli, A.; Bhambri, A.; Karn, S.K.; Raj, V. Ammonia, nitrite transformations and their fixation by different biological and chemical agents. Chem. Ecol. 2024, 40, 166–199. [Google Scholar] [CrossRef]
- Bowman, W.D.; Nemergut, D.R.; McKnight, D.M.; Miller, M.P.; Williams, M.W. A slide down a slippery slope–alpine ecosystem responses to nitrogen deposition. Plant Ecol. Divers. 2015, 8, 727–738. [Google Scholar] [CrossRef]
- Maciag, T.; Kozieł, E.; Otulak-Kozieł, K.; Jafra, S.; Czajkowski, R. Looking for resistance to soft rot disease of potatoes facing environmental hypoxia. Inter. J. Mol. Sci. 2024, 25, 3757. [Google Scholar] [CrossRef]
- Daunoras, J.; Kačergius, A.; Gudiukaitė, R. Role of soil microbiota enzymes in soil health and activity changes depending on climate change and the type of soil ecosystem. Biology 2024, 13, 85. [Google Scholar] [CrossRef]
- Banerjee, S.; Zhao, C.; Garland, G.; Edlinger, A.; García-Palacios, P.; Romdhane, S.; van der Heijden, M.G. Biotic homogenization, lower soil fungal diversity and fewer rare taxa in arable soils across Europe. Nat. Commun. 2024, 15, 327. [Google Scholar] [CrossRef]
- Mustafa, G.; Hussain, S.; Liu, Y.H.; Ali, I.; Liu, J.Y.; Bano, H. Microbiology of wetlands and the carbon cycle in coastal wetland mediated by microorganisms. Sci. Total Environ. 2024, 954, 175734. [Google Scholar] [CrossRef]
- Yao, Y.F.; Zhu, R.Y.; Li, X.D.; Hu, G.Q.; Dong, Y.J.; Liu, Z.X. Long-term adoption of plow tillage and green manure improves soil physicochemical properties and optimizes microbial communities under a continuous peanut monoculture system. Front. Microbiol. 2025, 15, 1513528. [Google Scholar] [CrossRef]
- Shaaban, M. Microbial pathways of nitrous oxide emissions and mitigation approaches in drylands. J. Environ. Manag. 2024, 354, 120393. [Google Scholar] [CrossRef] [PubMed]
- Kaviraj, M.; Kumar, U.; Chatterjee, S.; Parija, S.; Padbhushan, R.; Nayak, A.K.; Gupta, V.V. Dissimilatory nitrate reduction to ammonium (DNRA): A unique biogeochemical cycle to improve nitrogen (N) use efficiency and reduce N-loss in rice paddy. Rhizosphere 2024, 30, 100875. [Google Scholar] [CrossRef]
- Xing, Y.Y.; Wang, X.K. Impact of agricultural activities on climate change: A review of greenhouse gas emission patterns in field crop systems. Plants 2024, 13, 2285. [Google Scholar] [CrossRef]
- Stoicescu, S.T.; Hoikkala, L.; Fleming, V.; Lips, U. Continuing long-term expansion of low-oxygen conditions in the Eastern Gulf of Finland. Oceanologia 2024, 66, 139–152. [Google Scholar] [CrossRef]
- Pascual, R.; Piana, L.; Bhat, S.U.; Castro, P.F.; Corbera, J.; Cummings, D.; Delgado, C.; Eades, E.; Fensham, R.J.; Fernández-Martínez, M.; et al. The cultural ecohydrogeology of mediterranean-climate springs: A global review with case studies. Environments 2024, 11, 110. [Google Scholar] [CrossRef]
- Artime, O.; Grassia, M.; De Domenico, M.; Gleeson, J.P.; Makse, H.A.; Mangioni, G.; Perc, M.; Radicchi, F. Robustness and resilience of complex networks. Nat. Rev. Phys. 2024, 6, 114–131. [Google Scholar] [CrossRef]
- Mikiciuk, G.; Miller, T.; Kisiel, A.; Cembrowska-Lech, D.; Mikiciuk, M.; Łobodzińska, A.; Bokszczanin, K. Harnessing beneficial microbes for drought tolerance: A review of ecological and agricultural innovations. Agriculture 2024, 14, 2228. [Google Scholar] [CrossRef]
- Wu, D.; Yu, C.Q.; Zhao, Y.N.; Guo, J.L. Changes in vulnerability of global container shipping networks before and after the COVID-19 pandemic. J. Transp. Geogr. 2024, 114, 103783. [Google Scholar] [CrossRef]
- Gao, W.; Larjavaara, M. Wind disturbance in forests: A bibliometric analysis and systematic review. Forest Ecol. Manag. 2024, 564, 122001. [Google Scholar] [CrossRef]
- Xu, N.; Duan, H.R. Ecological functional zoning in urban fringe areas based on the trade-offs between ecological–social values in ecosystem services: A case study of Jiangning district, Nanjing. Land 2024, 13, 1957. [Google Scholar] [CrossRef]
- Francis, C.A.; Roberts, K.J.; Beman, J.M.; Santoro, A.E.; Oakley, B.B. Ubiquity and diversity of ammonia-oxidizing archaea in water columns and sediments of the ocean. Proc. Natl. Acad. Sci. USA 2005, 102, 14683–14688. [Google Scholar] [CrossRef] [PubMed]
- Rotthauwe, J.H.; Witzel, K.P.; Liesack, W. The ammonia monooxygenase structural gene amoA as a functional marker: Molecular fine-scale analysis of natural ammonia-oxidizing populations. Appl. Environ. Microb. 1997, 63, 4704–4712. [Google Scholar] [CrossRef]
- Braker, G.; Fesefeldt, A.; Witzel, K.P. Development of PCR primer systems for amplification of nitrite reductase genes (nirK and nirS) to detect denitrifying bacteria in environmental samples. Appl. Environ. Microb. 1998, 64, 3769–3775. [Google Scholar] [CrossRef] [PubMed]
- Throbäck, I.N.; Enwall, K.; Jarvis, Å.; Hallin, S. Reassessing PCR primers targeting nirS, nirK and nosZ genes for community surveys of denitrifying bacteria with DGGE. FEMS Microbiol. Ecol. 2004, 49, 401–417. [Google Scholar] [CrossRef]
- Govindarajulu, M.; Pfeffer, P.E.; Jin, H.; Abubaker, J.; Douds, D.D.; Allen, J.W.; Shachar-Hill, Y. Nitrogen transfer in the arbuscular mycorrhizal symbiosis. Nature 2005, 435, 819–823. [Google Scholar] [CrossRef] [PubMed]
- Rosch, C.; Bothe, H. Improved assessment of denitrifying, N2-fixing, and total-community bacteria by terminal restriction fragment length polymorphism analysis using multiple restriction enzymes. Appl. Environ. Microb. 2005, 71, 2026–2035. [Google Scholar] [CrossRef]
- Bru, D.; Sarr, A.; Philippot, L. Relative abundances of proteobacterial membrane-bound and periplasmic nitrate reductases in selected environments. Appl. Environ. Microb. 2007, 64, 5971–5974. [Google Scholar] [CrossRef]
- Song, W.S.; Kim, J.H.; Namgung, B.; Cho, H.Y.; Shin, H.; Oh, H.B.; Ha, N.C.; Yoon, S.I. Complementary hydrophobic interaction of the redox enzyme maturation protein NarJ with the signal peptide of the respiratory nitrate reductase NarG. Int. J. Biol. Macromol. 2024, 262, 129620. [Google Scholar] [CrossRef]
- Hamady, A.B.; Abd El-Fadeal, N.M.; Imbaby, S.; Nassar, H.M.; Sakr, M.G.; Marei, Y.E. Expression of norA, norB and norC efflux pump genes mediating fluoroquinolones resistance in MRSA isolates. J. Infect. Dev. Countr. 2024, 18, 399–406. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ding, J.; Yu, S. Impacts of Land Use on Soil Nitrogen-Cycling Microbial Communities: Insights from Community Structure, Functional Gene Abundance, and Network Complexity. Life 2025, 15, 466. https://doi.org/10.3390/life15030466
Ding J, Yu S. Impacts of Land Use on Soil Nitrogen-Cycling Microbial Communities: Insights from Community Structure, Functional Gene Abundance, and Network Complexity. Life. 2025; 15(3):466. https://doi.org/10.3390/life15030466
Chicago/Turabian StyleDing, Junnan, and Shaopeng Yu. 2025. "Impacts of Land Use on Soil Nitrogen-Cycling Microbial Communities: Insights from Community Structure, Functional Gene Abundance, and Network Complexity" Life 15, no. 3: 466. https://doi.org/10.3390/life15030466
APA StyleDing, J., & Yu, S. (2025). Impacts of Land Use on Soil Nitrogen-Cycling Microbial Communities: Insights from Community Structure, Functional Gene Abundance, and Network Complexity. Life, 15(3), 466. https://doi.org/10.3390/life15030466