
Brunneosporopsis yunnanensis gen. et sp. nov. and Allocryptovalsa xishuangbanica sp. nov., New Terrestrial Sordariomycetes from Southwest China

 ,
,  ,
,  ,
,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Herbarium Material
2.2. Morphological Observations
2.3. DNA Isolation, PCR Amplification and Sequencing
2.4. Sequence Alignment and Phylogenetic Analysis
3. Results
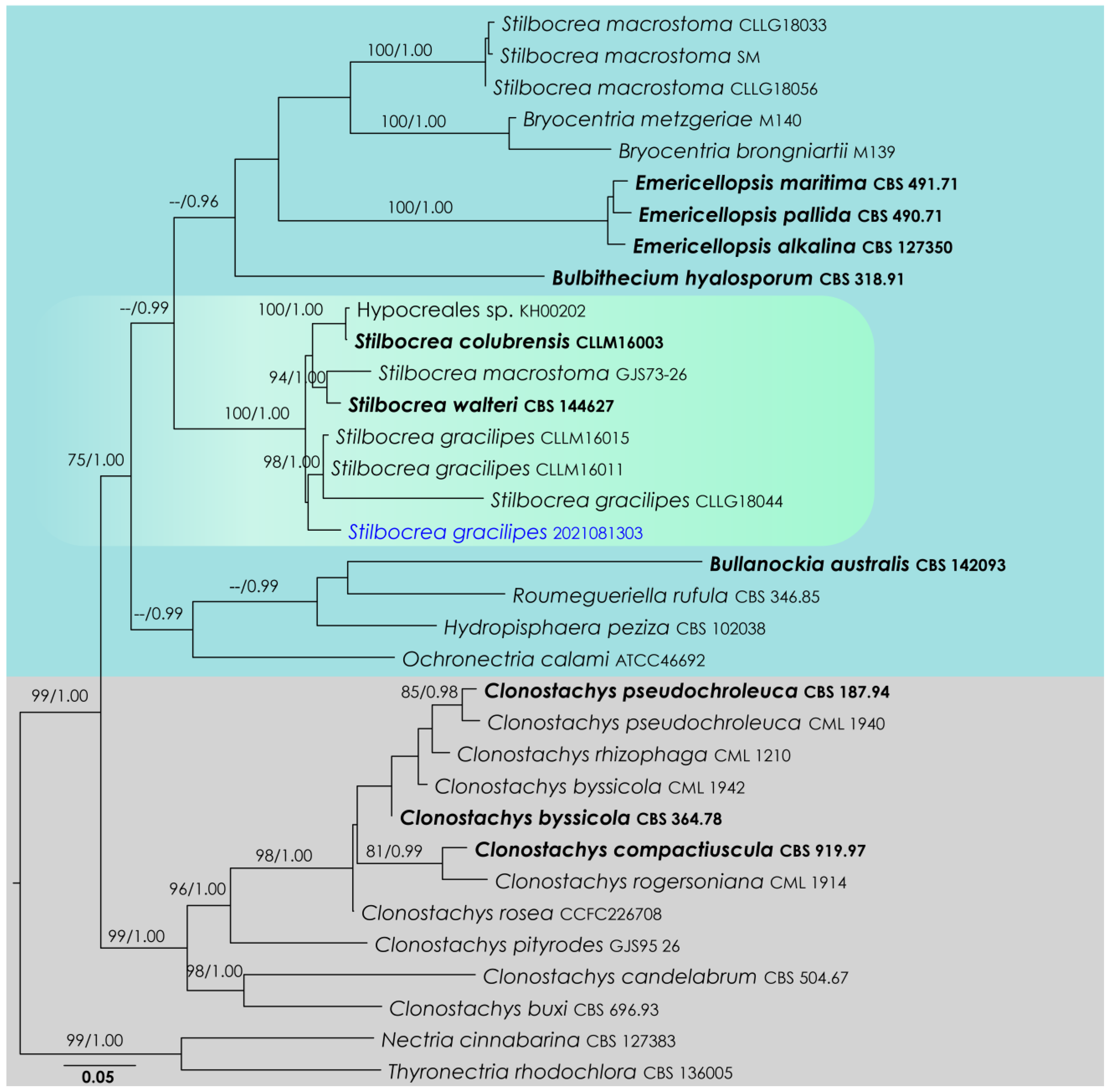
3.1. Stilbocrea Analysis
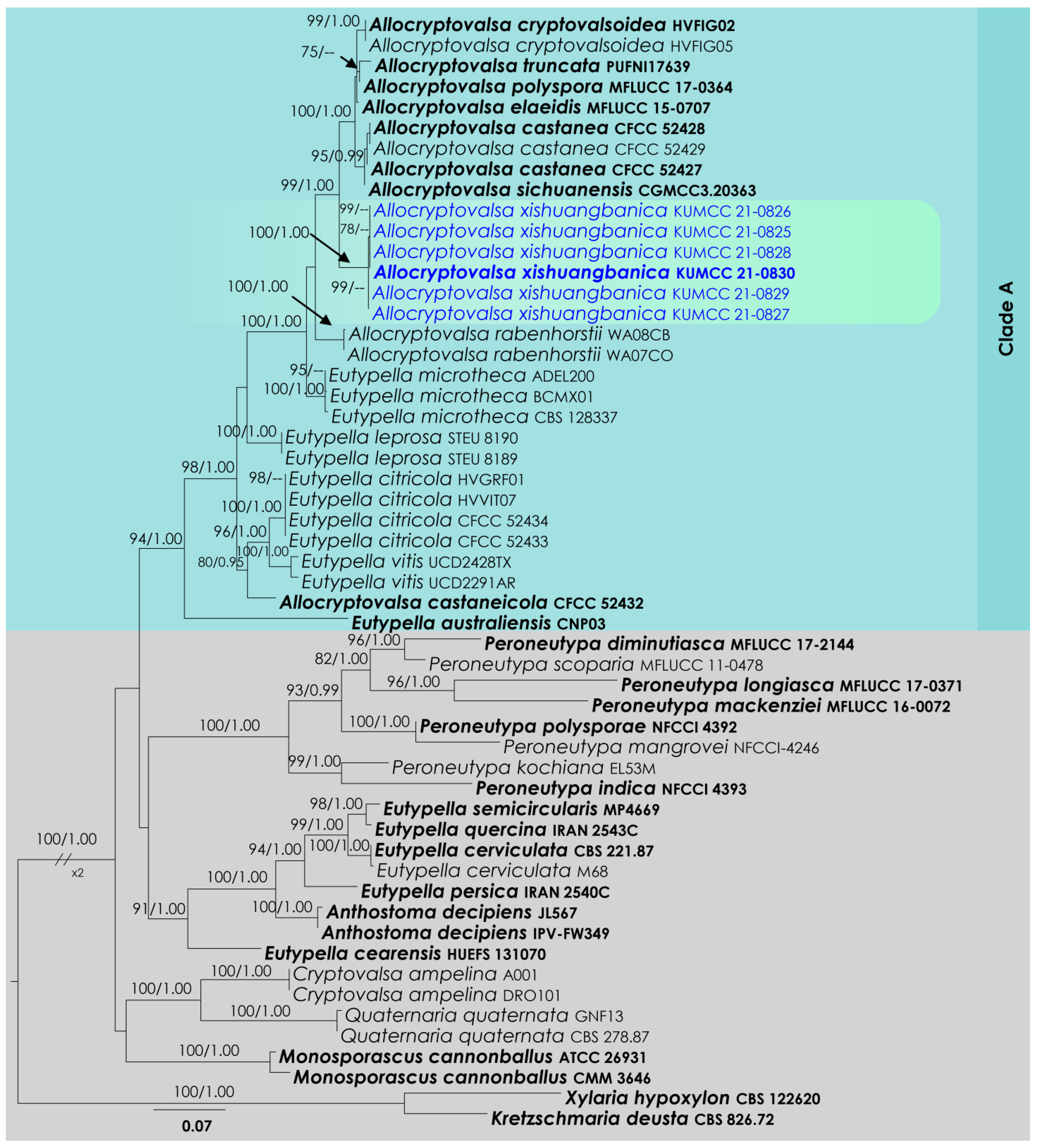
3.2. Allocryptovalsa Analysis
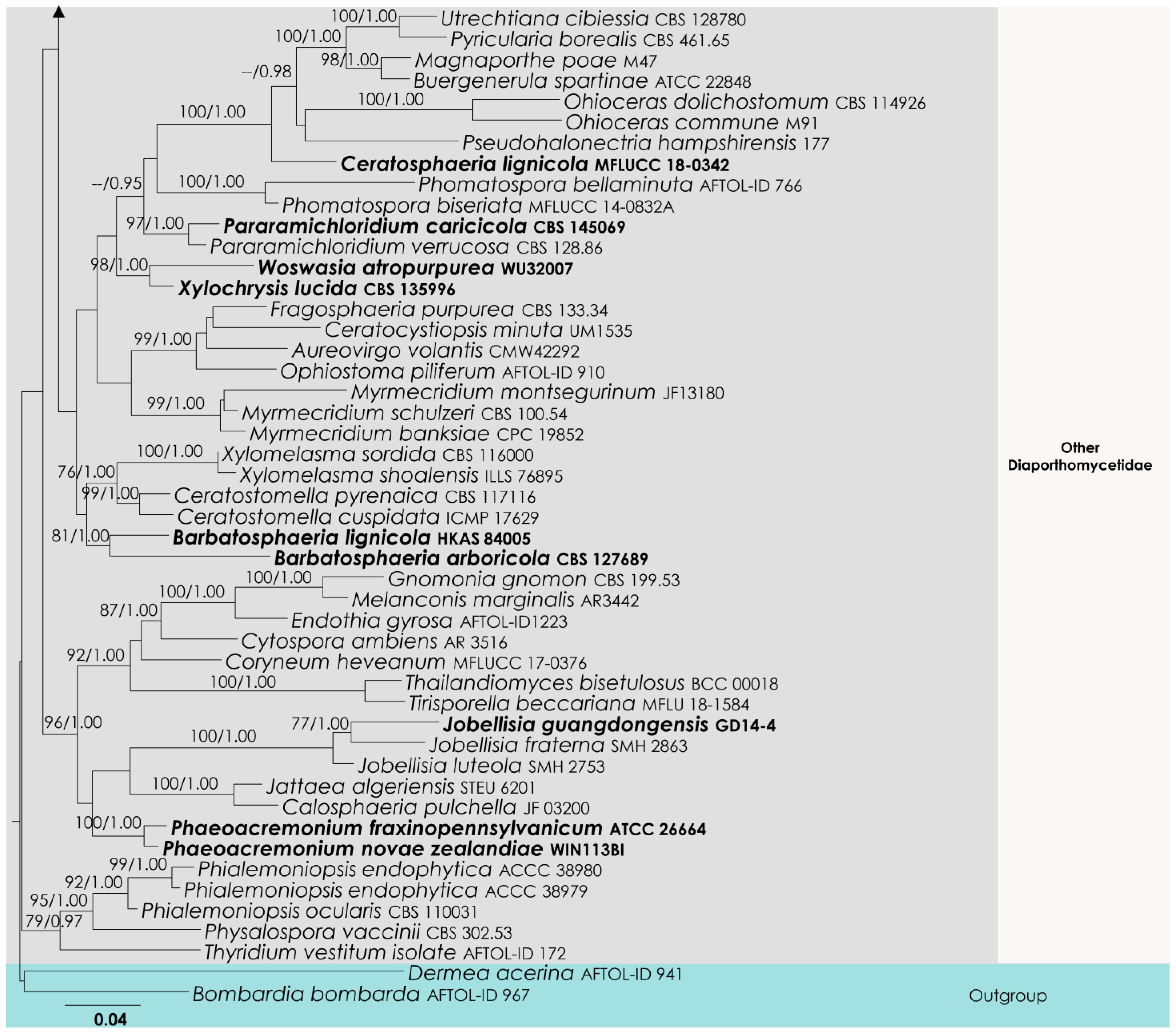
3.3. Diaporthomycetidae Analysis
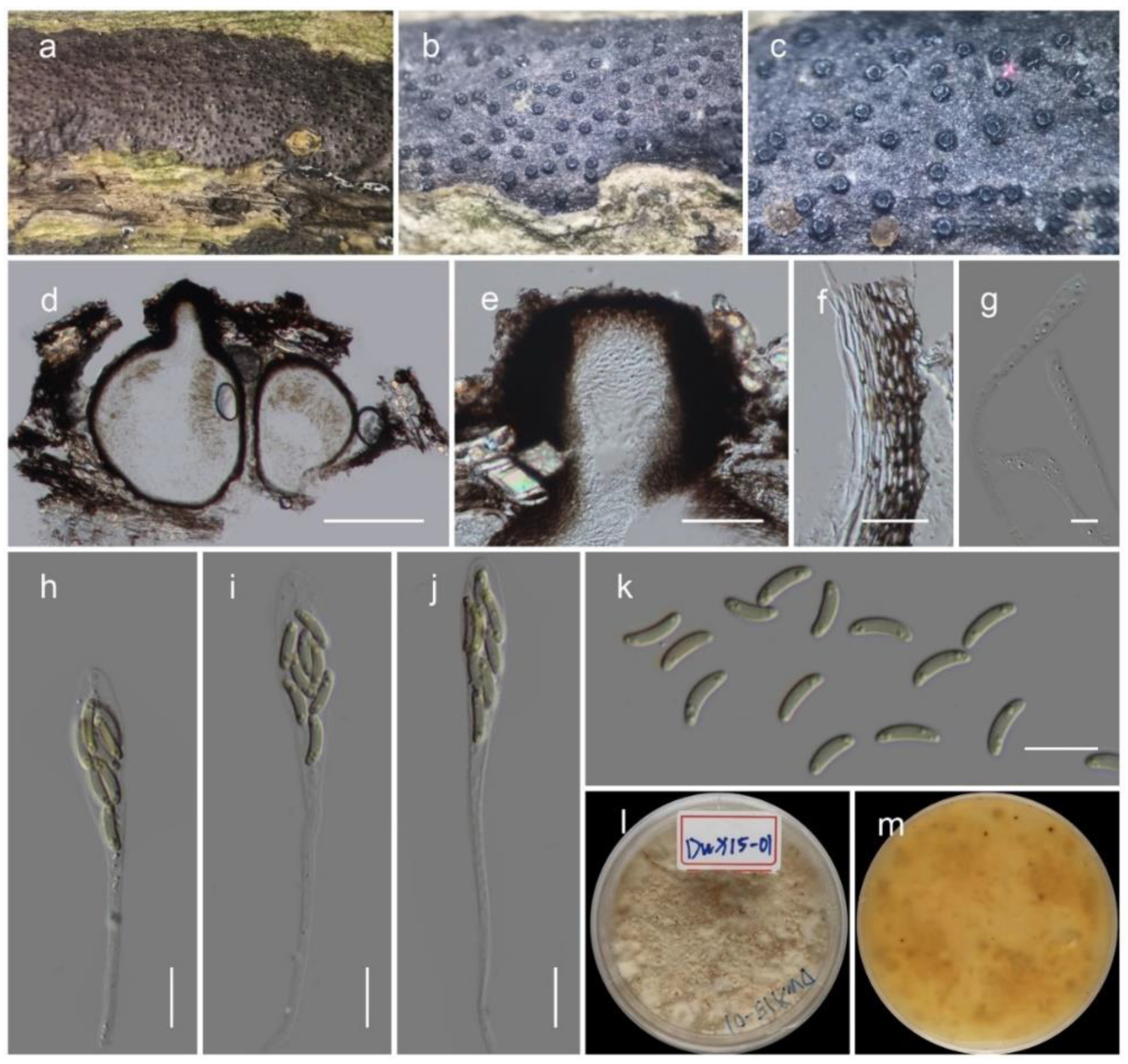
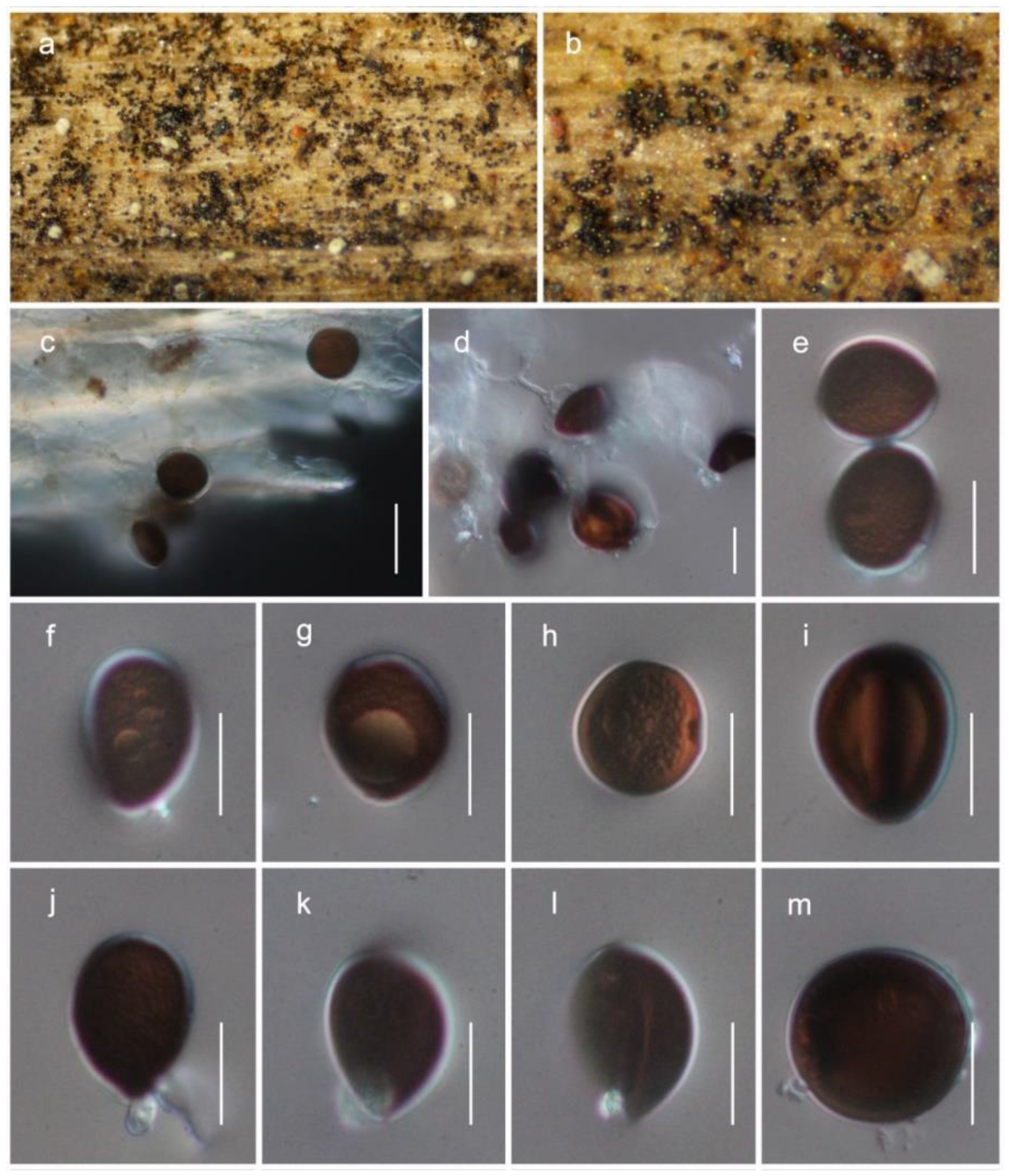
3.4. Taxonomy
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yang, J.; Maharachchikumbura, S.S.N.; Liu, J.K.; Hyde, K.D.; Jones, E.G.; Al-Sadi, A.M.; Liu, Z.Y. Pseudostanjehughesia aquitropica Gen. et Sp. Nov. and Sporidesmium Sensu Lato Species from Freshwater Habitats. Mycol. Prog. 2018, 17, 591–616. [Google Scholar] [CrossRef]
- Senanayake, I.C.; Jeewon, R.; Chomnunti, P.; Wanasinghe, D.N.; Norphanphoun, C.; Karunarathna, A.; Pem, D.; Perera, R.H.; Camporesi, E.; Eric, H.C.; et al. Taxonomic Circumscription of Diaporthales Based on Multigene Phylogeny and Morphology. Fungal Divers. 2018, 93, 241–443. [Google Scholar] [CrossRef]
- Jones, E.G.; Pang, K.-L.; Abdel-Wahab, M.A.; Scholz, B.; Hyde, K.D.; Boekhout, T.; Ebel, R.; Rateb, M.E.; Henderson, L.; Sakayaroj, J. An Online Resource for Marine Fungi. Fungal Divers. 2019, 96, 347–433. [Google Scholar] [CrossRef]
- Hyde, K.D.; Norphanphoun, C.; Maharachchikumbura, S.S.N.; Bhat, D.J.; Jones, E.B.G.; Bundhun, D.; Chen, Y.-J.; Bao, D.-F.; Boonmee, S.; Calabon, M.S.; et al. Refined Families of Sordariomycetes. Mycosphere 2020, 11, 305–1059. [Google Scholar] [CrossRef]
- Perera, R.H.; Hyde, K.D.; Maharachchikumbura, S.S.N.; Jones, E.B.G.; McKenzie, E.H.C.; Stadler, M.; Lee, H.B.; Samarakoon, M.C.; Ekanayaka, A.H.; Camporesi, E.; et al. Fungi on Wild Seeds and Fruits. Mycosphere 2020, 11, 2108–2480. [Google Scholar] [CrossRef]
- Zhang, N.; Castlebury, L.A.; Miller, A.N.; Huhndorf, S.M.; Schoch, C.L.; Seifert, K.A.; Rossman, A.Y.; Rogers, J.D.; Kohlmeyer, J.; Volkmann-Kohlmeyer, B.; et al. An Overview of the Systematics of the Sordariomycetes Based on a Four-Gene Phylogeny. Mycologia 2006, 98, 1076–1087. [Google Scholar] [CrossRef]
- U’Ren, J.M.; Miadlikowska, J.; Zimmerman, N.B.; Lutzoni, F.; Stajich, J.E.; Arnold, A.E. Contributions of North American Endophytes to the Phylogeny, Ecology, and Taxonomy of Xylariaceae (Sordariomycetes, Ascomycota). Mol. Phylogenet. Evol. 2016, 98, 210–232. [Google Scholar] [CrossRef] [Green Version]
- Maharachchikumbura, S.S.N.; Hyde, K.D.; Jones, E.B.G.; McKenzie, E.H.C.; Huang, S.K.; Abdel-Wahab, M.A.; Daranagama, D.A.; Dayarathne, M.; D’souza, M.J.; Goonasekara, I.D.; et al. Towards a Natural Classification and Backbone Tree for Sordariomycetes. Fungal Divers. 2015, 72, 199–301. [Google Scholar] [CrossRef]
- Ma, X.; Li, L.; Zhu, T.; Ba, M.; Li, G.; Gu, Q.; Guo, Y.; Li, D. Phenylspirodrimanes with Anti-HIV Activity from the Sponge-Derived Fungus Stachybotrys chartarum MXHX73. J. Nat. Prod. 2013, 76, 2298–2306. [Google Scholar] [CrossRef]
- Ma, X.; Kang, J.; Nontachaiyapoom, S.; Wen, T.; Hyde, K.D. Nonmycorrhizal Endophytic Fungi from Orchids. Curr. Sci. 2015, 109, 36–51. [Google Scholar]
- Kuhnert, E.; Navarro-Muñoz, J.; Becker, K.; Stadler, M.; Collemare, J.; Cox, R. Secondary Metabolite Biosynthetic Diversity in the Fungal Family Hypoxylaceae and Xylaria hypoxylon. Stud. Mycol. 2021, 99, 100–118. [Google Scholar] [CrossRef] [PubMed]
- Thambugala, K.M.; Daranagama, D.A.; Phillips, A.J.L.; Kannangara, S.D.; Promputtha, I. Fungi vs. Fungi in Biocontrol: An Overview of Fungal Antagonists Applied Against Fungal Plant Pathogens. Front. Cell. Infect. Microbiol. 2020, 10, 604923. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, O.E.; Winka, K. Supraordinal Taxa of Ascomycota. Myconet 1997, 1, 1–16. [Google Scholar]
- Hongsanan, S.; Maharachchikumbura, S.S.N.; Hyde, K.D.; Samarakoon, M.C.; Jeewon, R.; Zhao, Q.; Al-Sadi, A.M.; Bahkali, A.H. An Updated Phylogeny of Sordariomycetes Based on Phylogenetic and Molecular Clock Evidence. Fungal Divers. 2017, 84, 25–41. [Google Scholar] [CrossRef]
- Wijayawardene, N.N.; Hyde, K.D.; Dai, D.Q.; Sánchez-García, M.; Goto, B.T.; Saxena, R.K.; Erdoğdu, M.; Selçuk, F.; Rajeshkumar, K.C.; Aptroot, A.; et al. Outline of Fungi and Fungus-Like Taxa—2021. Mycosphere 2022, 13, 53–453. [Google Scholar]
- Wanasinghe, D.N.; Ren, G.-C.; Xu, J.-C.; Cheewangkoon, R.; Mortimer, P.E. Insight into the Taxonomic Resolution of the Pleosporalean Species Associated with Dead Woody Litter in Natural Forests from Yunnan, China. J. Fungi 2022, 8, 375. [Google Scholar] [CrossRef]
- Wanasinghe, D.N.; Phukhamsakda, C.; Hyde, K.D.; Jeewon, R.; Lee, H.B.; Jones, E.B.G.; Tibpromma, S.; Tennakoon, D.S.; Dissanayake, A.J.; Jayasiri, S.C.; et al. Fungal Diversity Notes 709–839: Taxonomic and Phylogenetic Contributions to Fungal Taxa with an Emphasis on Fungi on Rosaceae. Fungal Divers. 2018, 89, 1–236. [Google Scholar] [CrossRef]
- Wanasinghe, D.N.; Mortimer, P.E.; Xu, J. Insight into the Systematics of Microfungi Colonizing Dead Woody Twigs of Dodonaea viscosa in Honghe (China). J. Fungi 2021, 7, 180. [Google Scholar] [CrossRef]
- Voglmayr, H.; Jaklitsch, W.M. Stilbocrea walteri Sp. Nov., an Unusual Species of Bionectriaceae. Mycol. Prog. 2019, 18, 91–105. [Google Scholar] [CrossRef] [Green Version]
- White, T.J.; Bruns, T.; Lee, S.J.W.T.; Taylor, J.W. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. PCR Protocols Appl. 1990, 18, 315–322. [Google Scholar]
- Vilgalys, R.; Hester, M. Rapid Genetic Identification and Mapping of Enzymatically Amplified Ribosomal DNA from Several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woudenberg, J.H.C.; Aveskamp, M.M.; de Gruyter, J.; Spiers, A.G.; Crous, P.W. Multiple Didymella teleomorphs are linked to the Phoma clematidina morphotype. Persoonia 2009, 22, 56–62. [Google Scholar] [CrossRef] [Green Version]
- Rehner, S. Primers for Elongation Factor 1-α (EF1-α). Available online: http://ocid.NACSE.ORG/research/deephyphae/EF1primer.pdf (accessed on 30 September 2021).
- Rehner, S.A.; Buckley, E.A. Beauveria Phylogeny Inferred from Nuclear ITS and EF1-α Sequences: Evidence for Cryptic Diversification and Links to Cordyceps Teleomorphs. Mycologia 2005, 97, 84–98. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.L.; Whelen, S.; Hall, B.D. Phylogenetic Relationships among Ascomycetes: Evidence from an RNA Polymerase II Subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef] [PubMed]
- Sung, G.-H.; Sung, J.-M.; Hywel-Jones, N.L.; Spatafora, J.W. A Multigene Phylogeny of Clavicipitaceae (Ascomycota, Fungi): Identification of Localized Incongruence Using a Combinational Bootstrap. Mol. Phylogenet. Evol. 2007, 44, 1204–1223. [Google Scholar] [CrossRef] [PubMed]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A.; Chen, W. Nuclear Ribosomal Internal Transcribed Spacer (ITS) Region as a Universal DNA Barcode Marker for Fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. The CIPRES Science Gateway: Enabling High-Impact Science for Phylogenetics Researchers with Limited Resources. In Proceedings of the 1st Conference of the Extreme Science and Engineering Discovery Environment: Bridging from the Extreme to the Campus and Beyond, Chicago, IL, USA, 16–20 July 2012; Association for Computing Machinery: New York, NY, USA, 2012; pp. 1–8. [Google Scholar]
- Maharachchikumbura, S.S.N.; Wanasinghe, D.N.; Cheewangkoon, R.; Al-Sadi, A.M. Uncovering the Hidden Taxonomic Diversity of Fungi in Oman. Fungal Divers. 2021, 106, 229–268. [Google Scholar] [CrossRef]
- Nylander, J.A.A. MrModeltest 2.0. Program Distributed by the Author; Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Hyde, K.D.; Bao, D.F.; Hongsanan, S.; Chethana, K.W.; Yang, J.; Suwannarach, N. Evolution of Freshwater Diaporthomycetidae (Sordariomycetes) Provides Evidence for Five New Orders and Six New Families. Fungal Divers. 2021, 107, 71–105. [Google Scholar] [CrossRef]
- Zhang, H.; Dong, W.; Hyde, K.D.; Maharachchikumbura, S.S.N.; Hongsanan, S.; Bhat, D.J.; Al-Sadi, A.M.; Zhang, D. Towards a Natural Classification of Annulatascaceae-like Taxa: Introducing Atractosporales Ord. Nov. And Six New Families. Fungal Divers. 2017, 85, 75–110. [Google Scholar] [CrossRef]
- Luo, Z.L.; Hyde, K.D.; Liu, J.K.; Maharachchikumbura, S.S.N.; Jeewon, R.; Bao, D.F.; Bhat, D.J.; Lin, C.G.; Li, W.L.; Yang, J.; et al. Freshwater Sordariomycetes. Fungal Divers. 2019, 99, 451–660. [Google Scholar] [CrossRef] [Green Version]
- Seifert, K.A. A Monograph of Stilbella and Some Allied Hyphomycetes. Stud. Mycol. 1985, 27, 1–235. [Google Scholar]
- Guu, J.R.; Ju, Y.M.; Hsieh, H.J. Bionectriaceous Fungi Collected from Forests in Taiwan. Bot. Stud. 2010, 1, 51. [Google Scholar]
- Lechat, C.; Fournier, J. New Insights into Stilbocrea (Hypocreales, Bionectriaceae): Recognition of S. colubrensis, a New Species from Martinique (French West Indies), and Observations on Lifestyle and Synnematous Asexual Morph. Ascomycete 2019, 11, 183–190. [Google Scholar]
- Trouillas, F.P.; Pitt, W.M.; Sosnowski, M.R.; Huang, R.; Peduto, F.; Loschiavo, A.; Savocchia, S.; Scott, E.S.; Gubler, W.D. Taxonomy and DNA Phylogeny of Diatrypaceae Associated with Vitis vinifera and Other Woody Plants in Australia. Fungal Divers. 2011, 49, 203–223. [Google Scholar] [CrossRef] [Green Version]
- Zhu, H.; Pan, M.; Wijayawardene, N.N.; Jiang, N.; Ma, R.; Dai, D.; Tian, C.; Fan, X. The Hidden Diversity of Diatrypaceous Fungi in China. Front. Microbiol. 2021, 12, 706. [Google Scholar] [CrossRef]
- Senwanna, C.; Phookamsak, R.; Doilom, M.; Hyde, K.D.; Cheewangkoon, R. Novel Taxa of Diatrypaceae from Para Rubber (Hevea brasiliensis) in Northern Thailand; Introducing a Novel Genus Allocryptovalsa. Mycosphere 2017, 8, 1835–1855. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maharachchikumbura, S.S.N.; Wanasinghe, D.N.; Elgorban, A.M.; Al-Rejaie, S.S.; Kazerooni, E.A.; Cheewangkoon, R. Brunneosporopsis yunnanensis gen. et sp. nov. and Allocryptovalsa xishuangbanica sp. nov., New Terrestrial Sordariomycetes from Southwest China. Life 2022, 12, 635. https://doi.org/10.3390/life12050635
Maharachchikumbura SSN, Wanasinghe DN, Elgorban AM, Al-Rejaie SS, Kazerooni EA, Cheewangkoon R. Brunneosporopsis yunnanensis gen. et sp. nov. and Allocryptovalsa xishuangbanica sp. nov., New Terrestrial Sordariomycetes from Southwest China. Life. 2022; 12(5):635. https://doi.org/10.3390/life12050635
Chicago/Turabian StyleMaharachchikumbura, Sajeewa S. N., Dhanushka N. Wanasinghe, Abdallah M. Elgorban, Salim S. Al-Rejaie, Elham A. Kazerooni, and Ratchadawan Cheewangkoon. 2022. "Brunneosporopsis yunnanensis gen. et sp. nov. and Allocryptovalsa xishuangbanica sp. nov., New Terrestrial Sordariomycetes from Southwest China" Life 12, no. 5: 635. https://doi.org/10.3390/life12050635
APA StyleMaharachchikumbura, S. S. N., Wanasinghe, D. N., Elgorban, A. M., Al-Rejaie, S. S., Kazerooni, E. A., & Cheewangkoon, R. (2022). Brunneosporopsis yunnanensis gen. et sp. nov. and Allocryptovalsa xishuangbanica sp. nov., New Terrestrial Sordariomycetes from Southwest China. Life, 12(5), 635. https://doi.org/10.3390/life12050635