
Lipophilic and Hydrophilic Compounds from Arthrospira platensis and Its Effects on Tissue and Blood Cells—An Overview



Abstract
1. Introduction

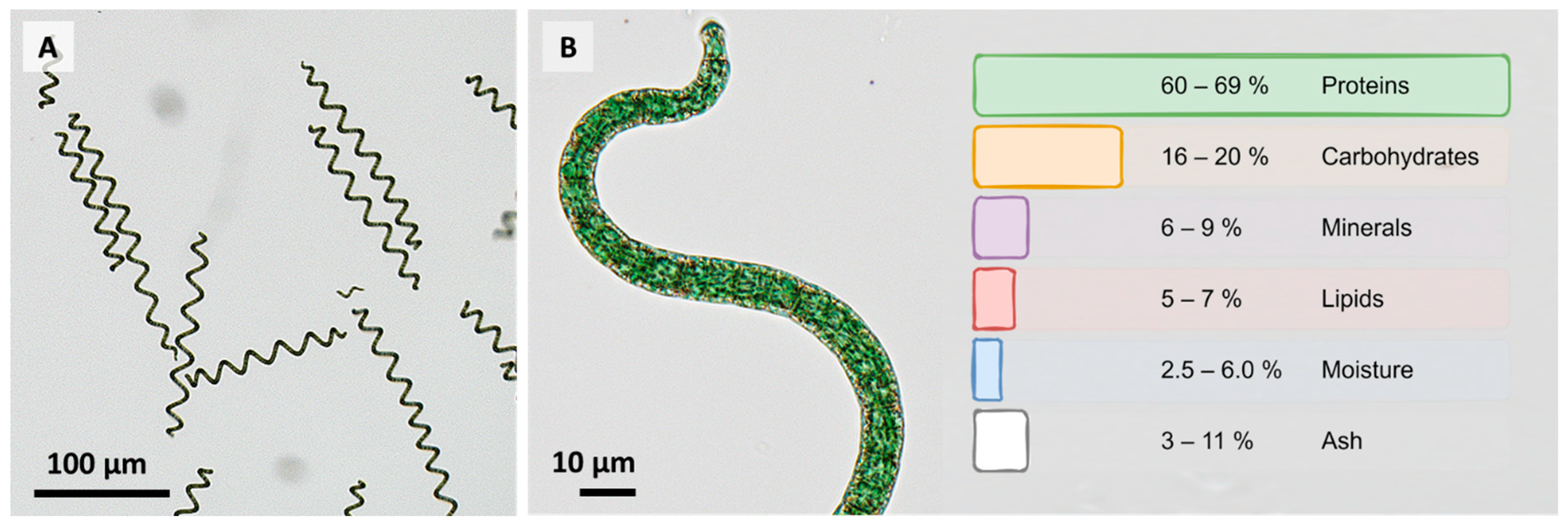{kind=link}
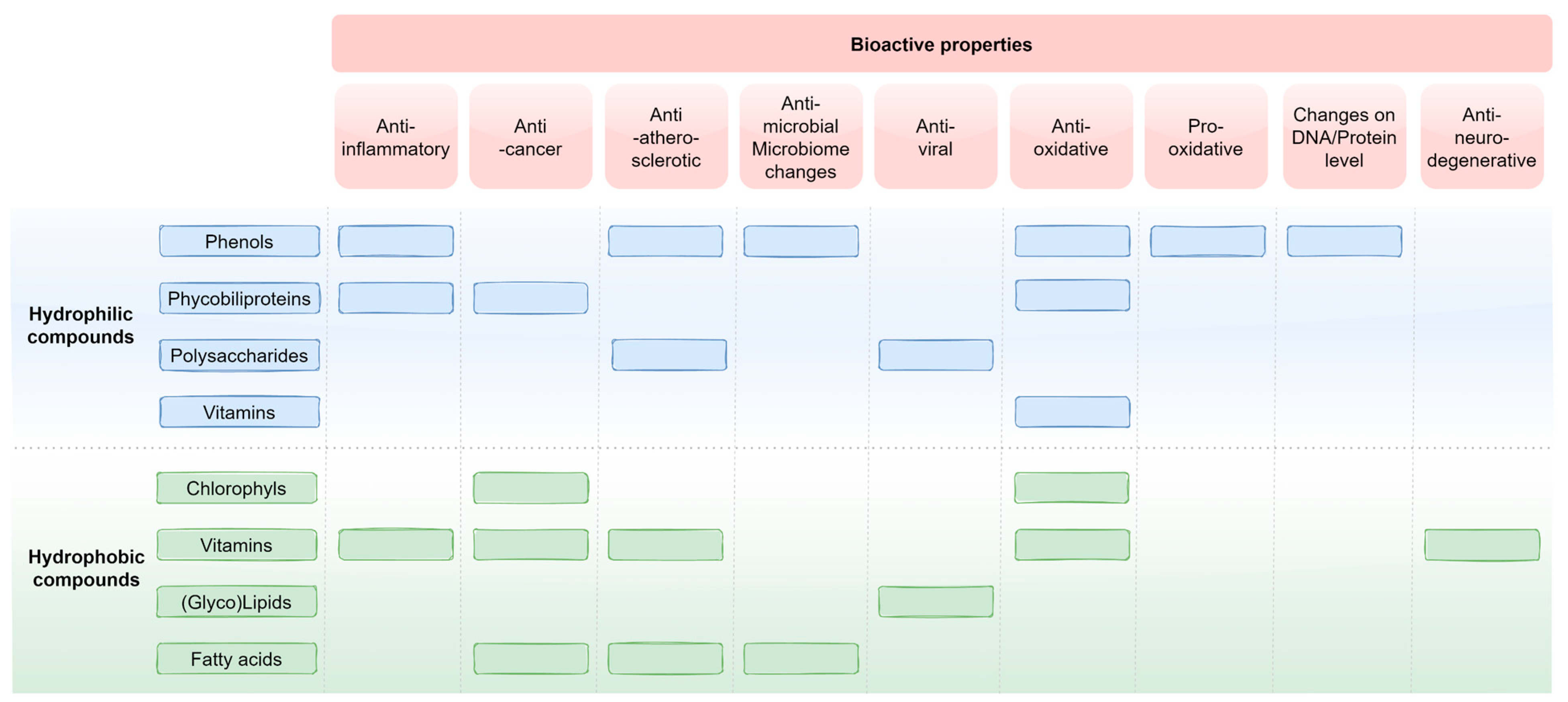{kind=link}
| Component | Amount per 100 g Dry Weight | |
|---|---|---|
| Calcium | 60–700 | mg |
| Chromium | 0.1–0.3 | mg |
| Copper | 0.20–1.2 | mg |
| Iodine | 0.142–n.a. | mg |
| Iron | 25–100 | mg |
| Manganese | 1–5 | mg |
| Magnesium | 200–400 | mg |
| Phosphorus | 700–1000 | mg |
| Potassium | 200–1830 | mg |
| Selenium | 0.003–0.010 | mg |
| Sodium | 700–1090 | mg |
| Zinc | 1–3 | mg |
2. Hydrophilic Compounds
2.1. Proteins
2.2. Phenols
| Component | Amount per 1 g Dry Biomass | |
|---|---|---|
| Apigenin | 6.00 ± 0.50 | ng |
| p-Coumaric acid | 920 ± 90 | ng |
| Catechin | n.a. | |
| Caffeic acid | n.a. | |
| Ferulic acid | 0.97 ± 0.12 | ng |
| Gallic acid | n.a. | |
| Genistein | n.a. | |
| p-Hydroxybenzoic acid | n.a. | |
| Kaempferol | n.a. | |
| Naringenin | n.a. | |
| Naringenin chalcone | n.a. | |
| Phloroglucinol | 51,000 ± 5000 | ng |
| Quercetin | n.a. | |
| Syringic acid | n.a. | |
| Vanillic acids | n.a. | |
| Superoxide dismutase enzymes | 392,000 | Units |
2.3. Phycobiliproteins
2.4. Polysaccharides
2.5. Vitamins (Vitamin C and Vitamins B1, B2, B6, and B12)
3. Lipophilic Compounds
3.1. Chlorophylls
3.2. Vitamins (Vitamin D, Vitamin E, Tocopherols and Tocotrienols, Vitamin K, and Provitamin A)
3.3. Lipids and Fatty Acids
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jung, F.; Krüger-Genge, A.; Waldeck, P.; Küpper, J.-H. Spirulina platensis, a Super Food? J. Cell. Biotechnol. 2019, 5, 43–54. [Google Scholar] [CrossRef]
- Braune, S.; Krüger-Genge, A.; Kammerer, S.; Jung, F.; Küpper, J.-H. Phycocyanin from Arthrospira platensis as Potential Anti-Cancer Drug: Review of in Vitro and in Vivo Studies. Life 2021, 11, 91. [Google Scholar] [CrossRef]
- Koníčková, R.; Vaňková, K.; Vaníková, J.; Váňová, K.; Muchová, L.; Subhanová, I.; Zadinová, M.; Zelenka, J.; Dvořák, A.; Kolář, M.; et al. Anti-Cancer Effects of Blue-Green Alga Spirulina platensis, a Natural Source of Bilirubin-like Tetrapyrrolic Compounds. Ann. Hepatol. 2014, 13, 273–283. [Google Scholar] [CrossRef]
- Mohan, A.; Misra, N.; Srivastav, D.; Umapathy, D.; Kumar, S. Spirulina, the Nature’s Wonder: A Review. Sch. J. Appl. Med. Sci. 2014, 2, 1334–1339. [Google Scholar]
- Sotiroudis, T.; Sotiroudis, G. Health Aspects of Spirulina (Arthrospira) Microalga Food Supplement. J. Serbian Chem. Soc. 2013, 78, 395–405. [Google Scholar] [CrossRef]
- Thomas, S.S. The Role of Parry Organic Spirulina in Health Management; Parry Nutraceuticals, Division of EID Parry (India): Chennai, India, 2010. [Google Scholar]
- Gershwin, M.E.; Belay, A. (Eds.) Spirulina in Human Nutrition and Health; CRC Press: Boca Raton, FL, USA, 2008; ISBN 978-1-4200-5256-5. [Google Scholar]
- Koru, E. Earth Food Spirulina (Arthrospira): Production and Quality Standarts. In Food Additive; El-Samragy, Y., Ed.; InTech: London, UK, 2012; ISBN 978-953-51-0067-6. [Google Scholar]
- Jung, F.; Jung, C.G.H.; Krüger-Genge, A.; Waldeck, P.; Küpper, J.-H. Factors Influencing the Growth of Spirulina platensis in Closed Photobioreactors under CO2–O2 Conversion. J. Cell. Biotechnol. 2019, 5, 125–134. [Google Scholar] [CrossRef]
- Aouir, A.; Amiali, M.; Bitam, A.; Benchabane, A.; Raghavan, V.G. Comparison of the Biochemical Composition of Different Arthrospira platensis Strains from Algeria, Chad and the USA. J. Food Meas. Charact. 2017, 11, 913–923. [Google Scholar] [CrossRef]
- Bhattacharya, S.; Shivaprakash, M. Evaluation of Three Spirulina Species Grown under Similar Conditions for Their Growth and Biochemicals. J. Sci. Food Agric. 2005, 85, 333–336. [Google Scholar] [CrossRef]
- Milia, M.; Corrias, F.; Addis, P.; Chini Zitelli, G.; Cicchi, B.; Torzillo, G.; Andreotti, V.; Angioni, A. Influence of Different Light Sources on the Biochemical Composition of Arthrospira Spp. Grown in Model Systems. Foods 2022, 11, 399. [Google Scholar] [CrossRef]
- Choopani, A.; Poorsoltan, M.; Fazilati, M.; Latifi, A.M.; Salavati, H. Spirulina: A Source of Gamma-Linoleic Acid and Its Applications. J. Appl. Biotechnol. Rep. 2016, 3, 483–488. [Google Scholar]
- Dibeklioglu, H.; Koru, E.; Diraman, H. Fatty Acid Profile of Spirulina platensis Used as a Food Supplement. Isr. J. Aquac.-Bamidgeh 2009, 61, 20548. [Google Scholar]
- McCance, R.A.; Widdowson, E.M. McCance and Widdowson’s the Composition of Foods, Seventh Summary ed.; Royal Society of Chemistry: Cambridge, UK, 2015; ISBN 978-1-84973-636-7. [Google Scholar]
- Becker, E.W. Micro-Algae as a Source of Protein. Biotechnol. Adv. 2007, 25, 207–210. [Google Scholar] [CrossRef] [PubMed]
- Maradona, R.M. Statement of the Head of IMSAM Observer Delegation to the United Nations Economic and Social Council at the Economic and SocialCouncil Substantive Session for 2008; Intergovernmental Institution for the Use of Micro Algae Spirulina against Malnutrition: New York, NY, USA, 2008; p. 2. [Google Scholar]
- United Nations. Report of the World Food Conference, Rome, 5–16 November 1974; United Nations: Rome, Italy, 1975; p. 64. Available online: https://digitallibrary.un.org/record/701143?ln=en (accessed on 23 August 2022).
- Smerilli, A.; Orefice, I.; Corato, F.; Gavalás Olea, A.; Ruban, A.V.; Brunet, C. Photoprotective and Antioxidant Responses to Light Spectrum and Intensity Variations in the Coastal Diatom Skeletonema Marinoi. Environ. Microbiol. 2017, 19, 611–627. [Google Scholar] [CrossRef]
- Manach, C.; Scalbert, A.; Morand, C.; Rémésy, C.; Jiménez, L. Polyphenols: Food Sources and Bioavailability. Am. J. Clin. Nutr. 2004, 79, 727–747. [Google Scholar] [CrossRef] [PubMed]
- Goiris, K.; Muylaert, K.; Voorspoels, S.; Noten, B.; De Paepe, D.; Baart, G.J.E.; De Cooman, L. Detection of Flavonoids in Microalgae from Different Evolutionary Lineages. J. Phycol. 2014, 50, 483–492. [Google Scholar] [CrossRef]
- Negara, B.F.S.P.; Sohn, J.H.; Kim, J.-S.; Choi, J.-S. Effects of Phlorotannins on Organisms: Focus on the Safety, Toxicity, and Availability of Phlorotannins. Foods 2021, 10, 452. [Google Scholar] [CrossRef] [PubMed]
- Gager, L.; Lalegerie, F.; Connan, S.; Stiger-Pouvreau, V. Marine Algal Derived Phenolic Compounds and Their Biological Activities for Medicinal and Cosmetic Applications. In Recent Advances in Micro and Macroalgal Processing; Rajauria, G., Yuan, Y.V., Eds.; Wiley: Hoboken, NJ, USA, 2021; pp. 278–334. ISBN 978-1-119-54265-0. [Google Scholar]
- Goiris, K.; Muylaert, K.; Fraeye, I.; Foubert, I.; De Brabanter, J.; De Cooman, L. Antioxidant Potential of Microalgae in Relation to Their Phenolic and Carotenoid Content. J. Appl. Phycol. 2012, 24, 1477–1486. [Google Scholar] [CrossRef]
- Quéguineur, B.; Goya, L.; Ramos, S.; Martín, M.A.; Mateos, R.; Bravo, L. Phloroglucinol: Antioxidant Properties and Effects on Cellular Oxidative Markers in Human HepG2 Cell Line. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2012, 50, 2886–2893. [Google Scholar] [CrossRef]
- Hodnick, W.F.; MllosavljeviĆ, E.B.; Nelson, J.H.; Pardini, R.S. Electrochemistry of Flavonoids. Biochem. Pharmacol. 1988, 37, 2607–2611. [Google Scholar] [CrossRef]
- Bae, J.-S. Antithrombotic and Profibrinolytic Activities of Phloroglucinol. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2011, 49, 1572–1577. [Google Scholar] [CrossRef]
- Barwell, C.J.; Blunden, G.; Manandhar, P.D. Isolation and Characterization of Brown Algal Polyphenols as Inhibitors of α-Amylase, Lipase and Trypsin. J. Appl. Phycol. 1989, 1, 319–323. [Google Scholar] [CrossRef]
- Singh, I.P.; Sidana, J.; Bansal, P.; Foley, W.J. Phloroglucinol Compounds of Therapeutic Interest: Global Patent and Technology Status. Expert Opin. Ther. Pat. 2009, 19, 847–866. [Google Scholar] [CrossRef] [PubMed]
- Cory, H.; Passarelli, S.; Szeto, J.; Tamez, M.; Mattei, J. The Role of Polyphenols in Human Health and Food Systems: A Mini-Review. Front. Nutr. 2018, 5, 87. [Google Scholar] [CrossRef] [PubMed]
- Salah, A.M.; Dongmo, A.B.; Kamanyi, A.; Bopelet, M.; Wagner, H. Angiotensin-Conventing Enzyme-Inhibitory Effect by Ruellia Praetermissa. Pharm. Biol. 2001, 39, 16–19. [Google Scholar] [CrossRef]
- Kanner, J. Polyphenols by Generating H2O2, Affect Cell Redox Signaling, Inhibit PTPs and Activate Nrf2 Axis for Adaptation and Cell Surviving: In Vitro, In Vivo and Human Health. Antioxidants 2020, 9, 797. [Google Scholar] [CrossRef] [PubMed]
- Yagi, H.; Tan, J.; Tuan, R.S. Polyphenols Suppress Hydrogen Peroxide-Induced Oxidative Stress in Human Bone-Marrow Derived Mesenchymal Stem Cells. J. Cell. Biochem. 2013, 114, 1163–1173. [Google Scholar] [CrossRef]
- Oteiza, P.I.; Fraga, C.G.; Mills, D.A.; Taft, D.H. Flavonoids and the Gastrointestinal Tract: Local and Systemic Effects. Mol. Aspects Med. 2018, 61, 41–49. [Google Scholar] [CrossRef]
- Sroka, Z.; Cisowski, W. Hydrogen Peroxide Scavenging, Antioxidant and Anti-Radical Activity of Some Phenolic Acids. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2003, 41, 753–758. [Google Scholar] [CrossRef]
- Li, H.; Cheng, K.; Wong, C.; Fan, K.; Chen, F.; Jiang, Y. Evaluation of Antioxidant Capacity and Total Phenolic Content of Different Fractions of Selected Microalgae. Food Chem. 2007, 102, 771–776. [Google Scholar] [CrossRef]
- Saeidnia, S.; Abdollahi, M. Antioxidants: Friends or Foe in Prevention or Treatment of Cancer: The Debate of the Century. Toxicol. Appl. Pharmacol. 2013, 271, 49–63. [Google Scholar] [CrossRef]
- Duda-Chodak, A. The Inhibitory Effect of Polyphenols on Human Gut Microbiota. J. Physiol. Pharmacol. Off. J. Pol. Physiol. Soc. 2012, 63, 497–503. [Google Scholar]
- Cushnie, T.P.T.; Lamb, A.J. Antimicrobial Activity of Flavonoids. Int. J. Antimicrob. Agents 2005, 26, 343–356. [Google Scholar] [CrossRef] [PubMed]
- Lapidot, T.; Walker, M.D.; Kanner, J. Can Apple Antioxidants Inhibit Tumor Cell Proliferation? Generation of H2O2 during Interaction of Phenolic Compounds with Cell Culture Media. J. Agric. Food Chem. 2002, 50, 3156–3160. [Google Scholar] [CrossRef] [PubMed]
- Galati, G.; O’Brien, P.J. Potential Toxicity of Flavonoids and Other Dietary Phenolics: Significance for Their Chemopreventive and Anticancer Properties. Free Radic. Biol. Med. 2004, 37, 287–303. [Google Scholar] [CrossRef] [PubMed]
- Forman, H.J.; Bernardo, A.; Davies, K.J.A. What Is the Concentration of Hydrogen Peroxide in Blood and Plasma? Arch. Biochem. Biophys. 2016, 603, 48–53. [Google Scholar] [CrossRef]
- León-González, A.J.; Auger, C.; Schini-Kerth, V.B. Pro-Oxidant Activity of Polyphenols and Its Implication on Cancer Chemoprevention and Chemotherapy. Biochem. Pharmacol. 2015, 98, 371–380. [Google Scholar] [CrossRef]
- Zolotareva, E.K.; Mokrosnop, V.M.; Stepanov, S.S. Polyphenol Compounds of Macroscopic and Microscopic Algae. Int. J. Algae 2019, 21, 5–24. [Google Scholar] [CrossRef]
- Papalia, T.; Sidari, R.; Panuccio, M.R. Impact of Different Storage Methods on Bioactive Compounds in Arthrospira platensis Biomass. Molecules 2019, 24, 2810. [Google Scholar] [CrossRef]
- Sunda, W.; Kieber, D.J.; Kiene, R.P.; Huntsman, S. An Antioxidant Function for DMSP and DMS in Marine Algae. Nature 2002, 418, 317–320. [Google Scholar] [CrossRef]
- Hirata, T.; Tanaka, M.; Ooike, M.; Tsunomura, T.; Sakaguchi, M. Antioxidant Activities of Phycocyanobilin Prepared from Spirulina platensis. J. Appl. Phycol. 2000, 12, 435–439. [Google Scholar] [CrossRef]
- Patel, A.; Mishra, S.; Ghosh, P.K. Antioxidant Potential of C-Phycocyanin Isolated from Cyanobacterial Species Lyngbya, Phormidium and Spirulina Spp. Indian J. Biochem. Biophys. 2006, 43, 25–31. [Google Scholar] [PubMed]
- Dartsch, P.C. Antioxidant Potential of Selected Spirulina platensis Preparations. Phytother. Res. PTR 2008, 22, 627–633. [Google Scholar] [CrossRef] [PubMed]
- Reddy, M.C.; Subhashini, J.; Mahipal, S.V.K.; Bhat, V.B.; Srinivas Reddy, P.; Kiranmai, G.; Madyastha, K.M.; Reddanna, P. C-Phycocyanin, a Selective Cyclooxygenase-2 Inhibitor, Induces Apoptosis in Lipopolysaccharide-Stimulated RAW 264.7 Macrophages. Biochem. Biophys. Res. Commun. 2003, 304, 385–392. [Google Scholar] [CrossRef]
- Shih, C.-M.; Cheng, S.-N.; Wong, C.-S.; Kuo, Y.-L.; Chou, T.-C. Antiinflammatory and Antihyperalgesic Activity of C-Phycocyanin. Anesth. Analg. 2009, 108, 1303–1310. [Google Scholar] [CrossRef] [PubMed]
- Romay, C.; Ledón, N.; González, R. Effects of Phycocyanin Extract on Prostaglandin E2 Levels in Mouse Ear Inflammation Test. Arzneimittelforschung 2000, 50, 1106–1109. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Wong, Y.-S. In Vitro Antioxidant and Antiproliferative Activities of Selenium-Containing Phycocyanin from Selenium-Enriched Spirulina platensis. J. Agric. Food Chem. 2008, 56, 4352–4358. [Google Scholar] [CrossRef]
- Li, B.; Chu, X.; Gao, M.; Li, W. Apoptotic Mechanism of MCF-7 Breast Cells in Vivo and in Vitro Induced by Photodynamic Therapy with C-Phycocyanin. Acta Biochim. Biophys. Sin. 2010, 42, 80–89. [Google Scholar] [CrossRef]
- Roy, K.R.; Arunasree, K.M.; Reddy, N.P.; Dheeraj, B.; Reddy, G.V.; Reddanna, P. Alteration of Mitochondrial Membrane Potential by Spirulina platensis C-Phycocyanin Induces Apoptosis in the Doxorubicinresistant Human Hepatocellular-Carcinoma Cell Line HepG2. Biotechnol. Appl. Biochem. 2007, 47, 159–167. [Google Scholar] [CrossRef]
- Bingula, R.; Dupuis, C.; Pichon, C.; Berthon, J.-Y.; Filaire, M.; Pigeon, L.; Filaire, E. Study of the Effects of Betaine and/or C-Phycocyanin on the Growth of Lung Cancer A549 Cells In Vitro and In Vivo. J. Oncol. 2016, 2016, 8162952. [Google Scholar] [CrossRef]
- Li, B.; Gao, M.-H.; Chu, X.-M.; Teng, L.; Lv, C.-Y.; Yang, P.; Yin, Q.-F. The Synergistic Antitumor Effects of All-Trans Retinoic Acid and C-Phycocyanin on the Lung Cancer A549 Cells In Vitro and In Vivo. Eur. J. Pharmacol. 2015, 749, 107–114. [Google Scholar] [CrossRef]
- Wang, H.; Liu, Y.; Gao, X.; Carter, C.L.; Liu, Z.-R. The Recombinant Beta Subunit of C-Phycocyanin Inhibits Cell Proliferation and Induces Apoptosis. Cancer Lett. 2007, 247, 150–158. [Google Scholar] [CrossRef] [PubMed]
- Subhashini, J.; Mahipal, S.V.K.; Reddy, M.C.; Mallikarjuna Reddy, M.; Rachamallu, A.; Reddanna, P. Molecular Mechanisms in C-Phycocyanin Induced Apoptosis in Human Chronic Myeloid Leukemia Cell Line-K562. Biochem. Pharmacol. 2004, 68, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Gardeva, E.; Toshkova, R.; Yossifova, L.; Minkova, K.; Ivanova, N.; Gigova, L. Antitumor Activity of C-Phycocyanin from Arthronema Africanum (Cyanophyceae). Braz. Arch. Biol. Technol. 2014, 57, 675–684. [Google Scholar] [CrossRef]
- Liao, G.; Gao, B.; Gao, Y.; Yang, X.; Cheng, X.; Ou, Y. Phycocyanin Inhibits Tumorigenic Potential of Pancreatic Cancer Cells: Role of Apoptosis and Autophagy. Sci. Rep. 2016, 6, 34564. [Google Scholar] [CrossRef]
- Remirez, D.; Ledón, N.; González, R. Role of Histamine in the Inhibitory Effects of Phycocyanin in Experimental Models of Allergic Inflammatory Response. Mediat. Inflamm. 2002, 11, 81–85. [Google Scholar] [CrossRef]
- Hao, S.; Li, S.; Wang, J.; Zhao, L.; Yan, Y.; Cao, Q.; Wu, T.; Liu, L.; Wang, C. Transcriptome Analysis of Phycocyanin-Mediated Inhibitory Functions on Non-Small Cell Lung Cancer A549 Cell Growth. Mar. Drugs 2018, 16, 511. [Google Scholar] [CrossRef]
- Hao, S.; Li, S.; Wang, J.; Zhao, L.; Yan, Y.; Wu, T.; Zhang, J.; Wang, C. C-Phycocyanin Suppresses the In Vitro Proliferation and Migration of Non-Small-Cell Lung Cancer Cells through Reduction of RIPK1/NF-ΚB Activity. Mar. Drugs 2019, 17, 362. [Google Scholar] [CrossRef]
- Pan, R.; Lu, R.; Zhang, Y.; Zhu, M.; Zhu, W.; Yang, R.; Zhang, E.; Ying, J.; Xu, T.; Yi, H.; et al. Spirulina Phycocyanin Induces Differential Protein Expression and Apoptosis in SKOV-3 Cells. Int. J. Biol. Macromol. 2015, 81, 951–959. [Google Scholar] [CrossRef]
- Hao, S.; Yan, Y.; Li, S.; Zhao, L.; Zhang, C.; Liu, L.; Wang, C. The In Vitro Anti-Tumor Activity of Phycocyanin against Non-Small Cell Lung Cancer Cells. Mar. Drugs 2018, 16, 178. [Google Scholar] [CrossRef]
- Liu, Q.; Huang, Y.; Zhang, R.; Cai, T.; Cai, Y. Medical Application of Spirulina platensis Derived C-Phycocyanin. Evid.-Based Complement. Altern. Med. ECAM 2016, 2016, 7803846. [Google Scholar] [CrossRef]
- Catassi, A.; Cesario, A.; Arzani, D.; Menichini, P.; Alama, A.; Bruzzo, C.; Imperatori, A.; Rotolo, N.; Granone, P.; Russo, P. Characterization of Apoptosis Induced by Marine Natural Products in Non Small Cell Lung Cancer A549 Cells. Cell. Mol. Life Sci. CMLS 2006, 63, 2377–2386. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Rojas, B.; Hernández-Juárez, J.; Pedraza-Chaverri, J. Nutraceutical Properties of Phycocyanin. J. Funct. Foods 2014, 11, 375–392. [Google Scholar] [CrossRef]
- Jung, C.H.G.; Waldeck, P.; Sykora, S.; Braune, S.; Petrick, I.; Küpper, J.-H.; Jung, F. Influence of Different Light-Emitting Diode Colors on Growth and Phycobiliprotein Generation of Arthrospira platensis. Life 2022, 12, 895. [Google Scholar] [CrossRef] [PubMed]
- Challouf, R.; Trabelsi, L.; Dhieb, R.B.; El Abed, O.; Yahia, A.; Ghozzi, K.; Ammar, J.B.; Omran, H.; Ouda, H.B. Evaluation of Cytotoxicity and Biological Activities in Extracellular Polysaccharides. Braz. Arch. Biol. Technol. 2011, 54, 831–838. [Google Scholar] [CrossRef]
- Kaji, T.; Fujiwara, Y.; Inomata, Y.; Hamada, C.; Yamamoto, C.; Shimada, S.; Lee, J.-B.; Hayashi, T. Repair of Wounded Monolayers of Cultured Bovine Aortic Endothelial Cells Is Inhibited by Calcium Spirulan, a Novel Sulfated Polysaccharide Isolated from Spirulina platensis. Life Sci. 2002, 70, 1841–1848. [Google Scholar] [CrossRef]
- Hayakawa, Y.; Hayashi, T.; Lee, J.B.; Ozawa, T.; Sakuragawa, N. Activation of Heparin Cofactor II by Calcium Spirulan. J. Biol. Chem. 2000, 275, 11379–11382. [Google Scholar] [CrossRef]
- Hayakawa, Y.; Hayashi, T.; Hayashi, K.; Ozawa, T.; Niiya, K.; Sakuragawa, N. Calcium Spirulan as an Inducer of Tissue-Type Plasminogen Activator in Human Fetal Lung Fibroblasts. Biochim. Biophys. Acta 1997, 1355, 241–247. [Google Scholar] [CrossRef]
- Abd El Baky, H.; Hanaa El Baz, K.; EL-Latife, S. Induction of Sulfated Polysaccharides in Spirulina platensis as Response to Nitrogen Concentration and Its Biological Evaluation. J. Aquac. Res. Dev. 2014, 5, 1. [Google Scholar] [CrossRef]
- Mader, J.; Gallo, A.; Schommartz, T.; Handke, W.; Nagel, C.-H.; Günther, P.; Brune, W.; Reich, K. Calcium Spirulan Derived from Spirulina Platensis Inhibits Herpes Simplex Virus 1 Attachment to Human Keratinocytes and Protects against Herpes Labialis. J. Allergy Clin. Immunol. 2016, 137, 197–203.e3. [Google Scholar] [CrossRef]
- Hayashi, K.; Hayashi, T.; Morita, N.; Kojima, I. An Extract from Spirulina platensis Is a Selective Inhibitor of Herpes Simplex Virus Type 1 Penetration into HeLa Cells. Phytother. Res. 1993, 7, 76–80. [Google Scholar] [CrossRef]
- Hayashi, T.; Hayashi, K.; Maeda, M.; Kojima, I. Calcium Spirulan, an Inhibitor of Enveloped Virus Replication, from a Blue-Green Alga Spirulina platensis. J. Nat. Prod. 1996, 59, 83–87. [Google Scholar] [CrossRef] [PubMed]
- Hayashi, K.; Hayashi, T.; Kojima, I. A Natural Sulfated Polysaccharide, Calcium Spirulan, Isolated from Spirulina platensis: In Vitro and Ex Vivo Evaluation of Anti-Herpes Simplex Virus and Anti-Human Immunodeficiency Virus Activities. AIDS Res. Hum. Retrovir. 1996, 12, 1463–1471. [Google Scholar] [CrossRef] [PubMed]
- Luescher-Mattli, M. Algae, A Possible Source for New Drugs in the Treatment of HIV and Other Viral Diseases. Curr. Med. Chem.-Anti-Infect. Agents 2003, 2, 219–225. [Google Scholar] [CrossRef]
- Hernández-Corona, A.; Nieves, I.; Meckes, M.; Chamorro, G.; Barron, B.L. Antiviral Activity of Spirulina Maxima against Herpes Simplex Virus Type 2. Antivir. Res. 2002, 56, 279–285. [Google Scholar] [CrossRef]
- Mishima, T.; Murata, J.; Toyoshima, M.; Fujii, H.; Nakajima, M.; Hayashi, T.; Kato, T.; Saiki, I. Inhibition of Tumor Invasion and Metastasis by Calcium Spirulan (Ca-SP), a Novel Sulfated Polysaccharide Derived from a Blue-Green Alga, Spirulina platensis. Clin. Exp. Metastasis 1998, 16, 541–550. [Google Scholar] [CrossRef]
- Pang, Q.S.; Guo, B.J.; Ruan, J.H. Enhancement of endonuclease activity and repair DNA synthesis by polysaccharide of Spirulina platensis. Yi Chuan Xue Bao 1988, 15, 374–381. [Google Scholar]
- Mittal, A.; Kumar, P.V.; Banerjee, S.; Rao, A.R.; Kumar, A. Modulatory Potential of Spirulina Fusiformis on Carcinogen Metabolizing Enzymes in Swiss Albino Mice. Phytother. Res. PTR 1999, 13, 111–114. [Google Scholar] [CrossRef]
- Sharma, S.; Poddar, R.; Sen, P.; Andrews, J. Effect of Vitamin C on Collagen Biosynthesis and Degree of Birefringence in Polarization Sensitive Optical Coherence Tomography (PS-OCT). Afr. J. Biotechnol. 2008, 7, 2049–2054. [Google Scholar]
- Knowles, H.J.; Raval, R.R.; Harris, A.L.; Ratcliffe, P.J. Effect of Ascorbate on the Activity of Hypoxia-Inducible Factor in Cancer Cells. Cancer Res. 2003, 63, 1764–1768. [Google Scholar]
- Padayatty, S.J.; Katz, A.; Wang, Y.; Eck, P.; Kwon, O.; Lee, J.-H.; Chen, S.; Corpe, C.; Dutta, A.; Dutta, S.K.; et al. Vitamin C as an Antioxidant: Evaluation of Its Role in Disease Prevention. J. Am. Coll. Nutr. 2003, 22, 18–35. [Google Scholar] [CrossRef]
- Jóźwiak, P.; Ciesielski, P.; Zaczek, A.; Lipińska, A.; Pomorski, L.; Wieczorek, M.; Bryś, M.; Forma, E.; Krześlak, A. Expression of Hypoxia Inducible Factor 1α and 2α and Its Association with Vitamin C Level in Thyroid Lesions. J. Biomed. Sci. 2017, 24, 83. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Wang, J.; Zhang, Y.; Wang, L.; Yu, M.; Wang, F. Vitamin C Decreases VEGF Expression Levels via Hypoxia-inducible Factor-1α Dependent and Independent Pathways in Lens Epithelial Cells. Mol. Med. Rep. 2020, 22, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Nakata, Y.; Maeda, N. Vulnerable Atherosclerotic Plaque Morphology in Apolipoprotein E–Deficient Mice Unable to Make Ascorbic Acid. Circulation 2002, 105, 1485–1490. [Google Scholar] [CrossRef] [PubMed]
- D’Aniello, C.; Cermola, F.; Patriarca, E.J.; Minchiotti, G. Vitamin C in Stem Cell Biology: Impact on Extracellular Matrix Homeostasis and Epigenetics. Stem Cells Int. 2017, 2017, 8936156. [Google Scholar] [CrossRef] [PubMed]
- Carcea, M.; Sorto, M.; Batello, C.; Narducci, V.; Aguzzi, A.; Azzini, E.; Fantauzzi, P.; Finotti, E.; Gabrielli, P.; Galli, V.; et al. Nutritional Characterization of Traditional and Improved Dihé, Alimentary Blue-Green Algae from the Lake Chad Region in Africa. LWT Food Sci. Technol. 2015, 62, 753–763. [Google Scholar] [CrossRef]
- Watanabe, F.; Yabuta, Y.; Bito, T.; Teng, F. Vitamin B12-Containing Plant Food Sources for Vegetarians. Nutrients 2014, 6, 1861–1873. [Google Scholar] [CrossRef]
- Hashimoto, E.; Yabuta, Y.; Takenaka, S.; Yamaguchi, Y.; Takenaka, H.; Watanabe, F. Characterization of Corrinoid Compounds from Edible Cyanobacterium Nostochopsis Sp. J. Nutr. Sci. Vitaminol. 2012, 58, 50–53. [Google Scholar] [CrossRef]
- Watanabe, F.; Tanioka, Y.; Miyamoto, E.; Fujita, T.; Takenaka, H.; Nakano, Y. Purification and Characterization of Corrinoid-Compounds from the Dried Powder of an Edible Cyanobacterium, Nostoc Commune (Ishikurage). J. Nutr. Sci. Vitaminol. 2007, 53, 183–186. [Google Scholar] [CrossRef]
- Watanabe, F.; Miyamoto, E.; Fujita, T.; Tanioka, Y.; Nakano, Y. Characterization of a Corrinoid Compound in the Edible (Blue-Green) Alga, Suizenji-Nori. Biosci. Biotechnol. Biochem. 2006, 70, 3066–3068. [Google Scholar] [CrossRef]
- Miyamoto, E.; Tanioka, Y.; Nakao, T.; Barla, F.; Inui, H.; Fujita, T.; Watanabe, F.; Nakano, Y. Purification and Characterization of a Corrinoid-Compound in an Edible Cyanobacterium Aphanizomenon Flos-Aquae as a Nutritional Supplementary Food. J. Agric. Food Chem. 2006, 54, 9604–9607. [Google Scholar] [CrossRef]
- Watanabe, F.; Katsura, H.; Takenaka, S.; Fujita, T.; Abe, K.; Tamura, Y.; Nakatsuka, T.; Nakano, Y. Pseudovitamin B(12) Is the Predominant Cobamide of an Algal Health Food, Spirulina Tablets. J. Agric. Food Chem. 1999, 47, 4736–4741. [Google Scholar] [CrossRef] [PubMed]
- Teng, F.; Bito, T.; Takenaka, S.; Takenaka, H.; Yamaguchi, Y.; Yabuta, Y.; Watanabe, F. Characterization of Corrinoid Compounds in the Edible Cyanobacterium <I>Nostoc Flagelliforme</I> the Hair Vegetable. Food Nutr. Sci. 2014, 5, 334–340. [Google Scholar] [CrossRef]
- Craig, W.J.; Mangels, A.R.; American Dietetic Association. Position of the American Dietetic Association: Vegetarian Diets. J. Am. Diet. Assoc. 2009, 109, 1266–1282. [Google Scholar] [CrossRef]
- Richmond, A.; Hu, Q. (Eds.) Handbook of Microalgal Culture: Applied Phycology and Biotechnology; John Wiley & Sons, Ltd.: Oxford, UK, 2013; ISBN 978-1-118-56716-6. [Google Scholar]
- Scaife, M.A.; Nguyen, G.T.D.T.; Rico, J.; Lambert, D.; Helliwell, K.E.; Smith, A.G. Establishing Chlamydomonas Reinhardtii as an Industrial Biotechnology Host. Plant J. Cell Mol. Biol. 2015, 82, 532–546. [Google Scholar] [CrossRef]
- Charoonnart, P.; Worakajit, N.; Zedler, J.A.Z.; Meetam, M.; Robinson, C.; Saksmerprome, V. Generation of Microalga Chlamydomonas Reinhardtii Expressing Shrimp Antiviral DsRNA without Supplementation of Antibiotics. Sci. Rep. 2019, 9, 3164. [Google Scholar] [CrossRef]
- Barra, L.; Chandrasekaran, R.; Corato, F.; Brunet, C. The Challenge of Ecophysiological Biodiversity for Biotechnological Applications of Marine Microalgae. Mar. Drugs 2014, 12, 1641–1675. [Google Scholar] [CrossRef] [PubMed]
- Suparmi, S.; Fasitasari, M.; Martosupono, M.; Mangimbulude, J.C. Comparisons of Curative Effects of Chlorophyll from Sauropus androgynus (L) Merr Leaf Extract and Cu-Chlorophyllin on Sodium Nitrate-Induced Oxidative Stress in Rats. J. Toxicol. 2016, 2016, 8515089. [Google Scholar] [CrossRef]
- Vaňková, K.; Marková, I.; Jašprová, J.; Dvořák, A.; Subhanová, I.; Zelenka, J.; Novosádová, I.; Rasl, J.; Vomastek, T.; Sobotka, R.; et al. Chlorophyll-Mediated Changes in the Redox Status of Pancreatic Cancer Cells Are Associated with Its Anticancer Effects. Oxid. Med. Cell. Longev. 2018, 2018, 4069167. [Google Scholar] [CrossRef]
- McQuistan, T.J.; Simonich, M.T.; Pratt, M.M.; Pereira, C.B.; Hendricks, J.D.; Dashwood, R.H.; Williams, D.E.; Bailey, G.S. Cancer Chemoprevention by Dietary Chlorophylls: A 12,000-Animal Dose–Dose Matrix Biomarker and Tumor Study. Food Chem. Toxicol. 2012, 50, 341–352. [Google Scholar] [CrossRef]
- Giammanco, M.; Di Majo, D.; La Guardia, M.; Aiello, S.; Crescimannno, M.; Flandina, C.; Tumminello, F.M.; Leto, G. Vitamin D in Cancer Chemoprevention. Pharm. Biol. 2015, 53, 1399–1434. [Google Scholar] [CrossRef]
- Atsuko, T.; Toshio, O.; Makoto, T.; Tadashi, K. Possible Origin of Extremely High Contents of Vitamin D3 in Some Kinds of Fish Liver. Comp. Biochem. Physiol. A Physiol. 1991, 100, 483–487. [Google Scholar] [CrossRef]
- Annweiler, C.; Rolland, Y.; Schott, A.M.; Blain, H.; Vellas, B.; Herrmann, F.R.; Beauchet, O. Higher Vitamin D Dietary Intake Is Associated with Lower Risk of Alzheimer’s Disease: A 7-Year Follow-Up. J. Gerontol. A Biol. Sci. Med. Sci. 2012, 67, 1205–1211. [Google Scholar] [CrossRef] [PubMed]
- Vanhevel, J.; Verlinden, L.; Doms, S.; Wildiers, H.; Verstuyf, A. The Role of Vitamin D in Breast Cancer Risk and Progression. Endocr. Relat. Cancer 2022, 29, R33–R55. [Google Scholar] [CrossRef] [PubMed]
- Lappe, J.M.; Travers-Gustafson, D.; Davies, K.M.; Recker, R.R.; Heaney, R.P. Vitamin D and Calcium Supplementation Reduces Cancer Risk: Results of a Randomized Trial. Am. J. Clin. Nutr. 2007, 85, 1586–1591. [Google Scholar] [CrossRef] [PubMed]
- Feldman, D.; Krishnan, A.V.; Swami, S.; Giovannucci, E.; Feldman, B.J. The Role of Vitamin D in Reducing Cancer Risk and Progression. Nat. Rev. Cancer 2014, 14, 342–357. [Google Scholar] [CrossRef]
- Ljubic, A.; Thulesen, E.T.; Jacobsen, C.; Jakobsen, J. UVB Exposure Stimulates Production of Vitamin D3 in Selected Microalgae. Algal Res. 2021, 59, 102472. [Google Scholar] [CrossRef]
- Ljubic, A.; Jacobsen, C.; Holdt, S.L.; Jakobsen, J. Microalgae Nannochloropsis Oceanica as a Future New Natural Source of Vitamin D3. Food Chem. 2020, 320, 126627. [Google Scholar] [CrossRef]
- Kawa, S.; Nikaido, T.; Aoki, Y.; Zhai, Y.; Kumagai, T.; Furihata, K.; Fujii, S.; Kiyosawa, K. Vitamin D Analogues Up-Regulate P21 and P27 during Growth Inhibition of Pancreatic Cancer Cell Lines. Br. J. Cancer 1997, 76, 884–889. [Google Scholar] [CrossRef]
- Bhoora, S.; Punchoo, R. Policing Cancer: Vitamin D Arrests the Cell Cycle. Int. J. Mol. Sci. 2020, 21, 9296. [Google Scholar] [CrossRef]
- Trummer, C.; Schwetz, V.; Pandis, M.; Grübler, M.R.; Verheyen, N.; Gaksch, M.; Zittermann, A.; März, W.; Aberer, F.; Lang, A.; et al. Effects of Vitamin D Supplementation on IGF-1 and Calcitriol: A Randomized-Controlled Trial. Nutrients 2017, 9, 623. [Google Scholar] [CrossRef]
- Ciulei, G.; Orasan, O.H.; Coste, S.C.; Cozma, A.; Negrean, V.; Procopciuc, L.M. Vitamin D and the Insulin-like Growth Factor System: Implications for Colorectal Neoplasia. Eur. J. Clin. Investig. 2020, 50, e13265. [Google Scholar] [CrossRef] [PubMed]
- González-Sancho, J.M.; Larriba, M.J.; Muñoz, A. Wnt and Vitamin D at the Crossroads in Solid Cancer. Cancers 2020, 12, 3434. [Google Scholar] [CrossRef] [PubMed]
- Larriba, M.J.; González-Sancho, J.M.; Barbáchano, A.; Niell, N.; Ferrer-Mayorga, G.; Muñoz, A. Vitamin D Is a Multilevel Repressor of Wnt/β-Catenin Signaling in Cancer Cells. Cancers 2013, 5, 1242–1260. [Google Scholar] [CrossRef]
- Lu, R.; Zhang, Y.-G.; Xia, Y.; Sun, J. Imbalance of Autophagy and Apoptosis in Intestinal Epithelium Lacking the Vitamin D Receptor. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2019, 33, 11845–11856. [Google Scholar] [CrossRef]
- Coulombier, N.; Jauffrais, T.; Lebouvier, N. Antioxidant Compounds from Microalgae: A Review. Mar. Drugs 2021, 19, 549. [Google Scholar] [CrossRef]
- Gómez-Coronado, D.J.M.; Ibañez, E.; Rupérez, F.J.; Barbas, C. Tocopherol Measurement in Edible Products of Vegetable Origin. J. Chromatogr. A 2004, 1054, 227–233. [Google Scholar] [CrossRef] [PubMed]
- Meydani, M. Vitamin E and Atherosclerosis: Beyond Prevention of LDL Oxidation. J. Nutr. 2001, 131, 366S–368S. [Google Scholar] [CrossRef]
- Niki, E. Do Free Radicals Play Causal Role in Atherosclerosis? Low Density Lipoprotein Oxidation and Vitamin E Revisited. J. Clin. Biochem. Nutr. 2011, 48, 3–7. [Google Scholar] [CrossRef]
- Bowry, V.W.; Ingold, K.U.; Stocker, R. Vitamin E in Human Low-Density Lipoprotein. When and How This Antioxidant Becomes a pro-Oxidant. Biochem. J. 1992, 288 Pt 2, 341–344. [Google Scholar] [CrossRef]
- Zhang, P.-Y.; Xu, X.; Li, X.-C. Cardiovascular Diseases: Oxidative Damage and Antioxidant Protection. Eur. Rev. Med. Pharmacol. Sci. 2014, 18, 3091–3096. [Google Scholar]
- Husain, K.; Francois, R.A.; Yamauchi, T.; Perez, M.; Sebti, S.M.; Malafa, M.P. Vitamin E δ-Tocotrienol Augments the Antitumor Activity of Gemcitabine and Suppresses Constitutive NF-ΚB Activation in Pancreatic Cancer. Mol. Cancer Ther. 2011, 10, 2363–2372. [Google Scholar] [CrossRef] [PubMed]
- Husain, K.; Francois, R.A.; Hutchinson, S.Z.; Neuger, A.M.; Lush, R.; Coppola, D.; Sebti, S.; Malafa, M.P. Vitamin E Delta-Tocotrienol Levels in Tumor and Pancreatic Tissue of Mice after Oral Administration. Pharmacology 2009, 83, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Ni, J.; Wen, X.; Yao, J.; Chang, H.-C.; Yin, Y.; Zhang, M.; Xie, S.; Chen, M.; Simons, B.; Chang, P.; et al. Tocopherol-Associated Protein Suppresses Prostate Cancer Cell Growth by Inhibition of the Phosphoinositide 3-Kinase Pathway. Cancer Res. 2005, 65, 9807–9816. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Siti, H.N.; Kamisah, Y.; Kamsiah, J. The Role of Oxidative Stress, Antioxidants and Vascular Inflammation in Cardiovascular Disease (a Review). Vascul. Pharmacol. 2015, 71, 40–56. [Google Scholar] [CrossRef] [PubMed]
- Ashor, A.W.; Siervo, M.; Lara, J.; Oggioni, C.; Afshar, S.; Mathers, J.C. Effect of Vitamin C and Vitamin E Supplementation on Endothelial Function: A Systematic Review and Meta-Analysis of Randomised Controlled Trials. Br. J. Nutr. 2015, 113, 1182–1194. [Google Scholar] [CrossRef]
- Tarento, T.D.C.; McClure, D.D.; Vasiljevski, E.; Schindeler, A.; Dehghani, F.; Kavanagh, J.M. Microalgae as a Source of Vitamin K1. Algal Res. 2018, 36, 77–87. [Google Scholar] [CrossRef]
- Simes, D.C.; Viegas, C.S.B.; Araújo, N.; Marreiros, C. Vitamin K as a Diet Supplement with Impact in Human Health: Current Evidence in Age-Related Diseases. Nutrients 2020, 12, 138. [Google Scholar] [CrossRef]
- Halder, M.; Petsophonsakul, P.; Akbulut, A.C.; Pavlic, A.; Bohan, F.; Anderson, E.; Maresz, K.; Kramann, R.; Schurgers, L. Vitamin K: Double Bonds beyond Coagulation Insights into Differences between Vitamin K1 and K2 in Health and Disease. Int. J. Mol. Sci. 2019, 20, 896. [Google Scholar] [CrossRef]
- Katsuura, S.; Imamura, T.; Bando, N.; Yamanishi, R. Beta-Carotene and Beta-Cryptoxanthin but Not Lutein Evoke Redox and Immune Changes in RAW264 Murine Macrophages. Mol. Nutr. Food Res. 2009, 53, 1396–1405. [Google Scholar] [CrossRef]
- Bai, S.-K.; Lee, S.-J.; Na, H.-J.; Ha, K.-S.; Han, J.-A.; Lee, H.; Kwon, Y.-G.; Chung, C.-K.; Kim, Y.-M. Beta-Carotene Inhibits Inflammatory Gene Expression in Lipopolysaccharide-Stimulated Macrophages by Suppressing Redox-Based NF-KappaB Activation. Exp. Mol. Med. 2005, 37, 323–334. [Google Scholar] [CrossRef]
- Schafer, F.Q.; Wang, H.P.; Kelley, E.E.; Cueno, K.L.; Martin, S.M.; Buettner, G.R. Comparing Beta-Carotene, Vitamin E and Nitric Oxide as Membrane Antioxidants. Biol. Chem. 2002, 383, 671–681. [Google Scholar] [CrossRef] [PubMed]
- Kirsh, V.A.; Hayes, R.B.; Mayne, S.T.; Chatterjee, N.; Subar, A.F.; Dixon, L.B.; Albanes, D.; Andriole, G.L.; Urban, D.A.; Peters, U.; et al. Supplemental and Dietary Vitamin E, Beta-Carotene, and Vitamin C Intakes and Prostate Cancer Risk. J. Natl. Cancer Inst. 2006, 98, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Vitamin A Supplementation: Who, When and How. Community Eye Health 2013, 26, 71.
- World Health Organization; Food and Agriculture Organization of the United Nations (Eds.) Vitamin and Mineral Requirements in Human Nutrition, 2nd ed.; World Health Organization: Geneva, Switzerland; FAO: Rome, Italy, 2004; ISBN 978-92-4-154612-6. [Google Scholar]
- Yates, C.M.; Calder, P.C.; Ed Rainger, G. Pharmacology and Therapeutics of Omega-3 Polyunsaturated Fatty Acids in Chronic Inflammatory Disease. Pharmacol. Ther. 2014, 141, 272–282. [Google Scholar] [CrossRef] [PubMed]
- Ramadan, M.F.; Asker, M.M.S.; Ibrahim, Z.K. Functional Bioactive Compounds and Biological Activities of Spirulina platensis Lipids. Czech J. Food Sci. 2008, 26, 211–222. [Google Scholar] [CrossRef]
- Xue, C.; Hu, Y.; Saito, H.; Zhang, Z.; Li, Z.; Cai, Y.; Ou, C.; Lin, H.; Imbs, A.B. Molecular Species Composition of Glycolipids from Sprirulina Platensis. Food Chem. 2002, 77, 9–13. [Google Scholar] [CrossRef]
- Colombo, D.; Gagliardi, C.; Vetro, M.; Ronchetti, F.; Takasaki, M.; Konoshima, T.; Suzuki, N.; Tokuda, H. New 6-Amino-6-Deoxy-Glycoglycerolipids Derived from 2-O-β-d-Glucopyranosylglycerol: Insights into the Structure–Activity Relationship of Glycoglycerolipids as Anti-Tumor Promoters. Carbohydr. Res. 2013, 373, 64–74. [Google Scholar] [CrossRef]
- Chirasuwan, N.; Chaiklahan, R.; Kittakoop, P.; Chanasattru, W.; Ruengjitchatchawalya, M.; Tanticharoen, M.; Bunnag, B. Anti HSV1 Activity of Sulphoquinovosyl Diacylglycerol Isolated from Arthrospira platensis. Sci. Asia 2009, 35, 137–141. [Google Scholar] [CrossRef]
- Ayehunie, S.; Belay, A.; Baba, T.W.; Ruprecht, R.M. Inhibition of HIV-1 Replication by an Aqueous Extract of Spirulina platensis (Arthrospira platensis). J. Acquir. Immune Defic. Syndr. Hum. Retrovirol. Off. Publ. Int. Retrovirol. Assoc. 1998, 18, 7–12. [Google Scholar] [CrossRef]
- Gustafson, K.R.; Cardellina, J.H.; Fuller, R.W.; Weislow, O.S.; Kiser, R.F.; Snader, K.M.; Patterson, G.M.L.; Boyd, M.R. AIDS-Antiviral Sulfolipids From Cyanobacteria (Blue-Green Algae). JNCI J. Natl. Cancer Inst. 1989, 81, 1254–1258. [Google Scholar] [CrossRef]
- Reshef, V.; Mizrachi, E.; Maretzki, T.; Silberstein, C.; Loya, S.; Hizi, A.; Carmeli, S. New Acylated Sulfoglycolipids and Digalactolipids and Related Known Glycolipids from Cyanobacteria with a Potential to Inhibit the Reverse Transcriptase of HIV-1. J. Nat. Prod. 1997, 60, 1251–1260. [Google Scholar] [CrossRef] [PubMed]
- Bruno, A.; Rossi, C.; Marcolongo, G.; Di Lena, A.; Venzo, A.; Berrie, C.P.; Corda, D. Selective in Vivo Anti-Inflammatory Action of the Galactolipid Monogalactosyldiacylglycerol. Eur. J. Pharmacol. 2005, 524, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Meireles, L.A.; Guedes, A.C.; Malcata, F.X. Lipid Class Composition of the Microalga Pavlova Lutheri : Eicosapentaenoic and Docosahexaenoic Acids. J. Agric. Food Chem. 2003, 51, 2237–2241. [Google Scholar] [CrossRef] [PubMed]
- Banskota, A.H.; Gallant, P.; Stefanova, R.; Melanson, R.; O’Leary, S.J.B. Monogalactosyldiacylglycerols, Potent Nitric Oxide Inhibitors from the Marine Microalga Tetraselmis Chui. Nat. Prod. Res. 2013, 27, 1084–1090. [Google Scholar] [CrossRef]
- Banskota, A.; Stefanova, R.; Sperker, S.; Lall, S.; Craigie, J.; Hafting, J.; Critchley, A. Polar Lipids from the Marine Macroalga Palmaria Palmata Inhibit Lipopolysaccharide-Induced Nitric Oxide Production in RAW264.7 Macrophage Cells. Phytochemistry 2014, 101, 101–108. [Google Scholar] [CrossRef]
- Hossain, Z.; Kurihara, H.; Hosokawa, M.; Takahashi, K. Growth Inhibition and Induction of Differentiation and Apoptosis Mediated by Sodium Butyrate in Caco-2 Cells with Algal Glycolipids. In Vitro Cell. Dev. Biol. Anim. 2005, 41, 154–159. [Google Scholar] [CrossRef]
- Ricigliano, V.A.; Simone-Finstrom, M. Nutritional and Prebiotic Efficacy of the Microalga Arthrospira platensis (Spirulina) in Honey Bees. Apidologie 2020, 51, 898–910. [Google Scholar] [CrossRef]
- Fithriani, D.; Sinurat, E. Utilization of Spirulina as Functional Food: Phytosterol and Amino Acid Profiles Study. IOP Conf. Ser. Earth Environ. Sci. 2019, 278, 012028. [Google Scholar] [CrossRef]
- El Baroty, G.S.; Abd El Baky, H.H.; Saleh, M.A. Egyptian Arthrospira Phytosterols: Production, Identification, Antioxidant and Antiproliferative Activities. Not. Bot. Horti Agrobot. Cluj-Napoca 2020, 48, 666–680. [Google Scholar] [CrossRef]
- Lope, V.; Guerrero-Zotano, Á.; Casas, A.; Baena-Cañada, J.M.; Bermejo, B.; Pérez-Gómez, B.; Criado-Navarro, I.; Antolín, S.; Sánchez-Rovira, P.; Ramos-Vázquez, M.; et al. Serum Phospholipids Fatty Acids and Breast Cancer Risk by Pathological Subtype. Nutrients 2020, 12, 3132. [Google Scholar] [CrossRef]
- Zhou, Y.; Wang, T.; Zhai, S.; Li, W.; Meng, Q. Linoleic Acid and Breast Cancer Risk: A Meta-Analysis. Public Health Nutr. 2016, 19, 1457–1463. [Google Scholar] [CrossRef] [PubMed]
- Ogata, R.; Mori, S.; Kishi, S.; Sasaki, R.; Iwata, N.; Ohmori, H.; Sasaki, T.; Nishiguchi, Y.; Nakashima, C.; Goto, K.; et al. Linoleic Acid Upregulates Microrna-494 to Induce Quiescence in Colorectal Cancer. Int. J. Mol. Sci. 2021, 23, 225. [Google Scholar] [CrossRef] [PubMed]
- Ohmori, H.; Sasahira, T.; Fujii, K.; Yi, L.; Shimomoto, T.; Kuniyasu, H. Linoleic-Acid-Induced Growth Suppression Induces Quiescent Cancer Cell Nests in Nude Mice. Pathobiol. J. Immunopathol. Mol. Cell. Biol. 2008, 75, 226–232. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Guasch-Ferré, M.; Li, Y.; Hu, F.B. Dietary Intake and Biomarkers of Linoleic Acid and Mortality: Systematic Review and Meta-Analysis of Prospective Cohort Studies. Am. J. Clin. Nutr. 2020, 112, 150–167. [Google Scholar] [CrossRef]
- Farvid, M.S.; Ding, M.; Pan, A.; Sun, Q.; Chiuve, S.E.; Steffen, L.M.; Willett, W.C.; Hu, F.B. Dietary Linoleic Acid and Risk of Coronary Heart Disease: A Systematic Review and Meta-Analysis of Prospective Cohort Studies. Circulation 2014, 130, 1568–1578. [Google Scholar] [CrossRef]
- Carta, G.; Murru, E.; Banni, S.; Manca, C. Palmitic Acid: Physiological Role, Metabolism and Nutritional Implications. Front. Physiol. 2017, 8, 902. [Google Scholar] [CrossRef]
- German, J.B. Dietary Lipids from an Evolutionary Perspective: Sources, Structures and Functions. Matern. Child. Nutr. 2011, 7 (Suppl. 2), 2–16. [Google Scholar] [CrossRef]
- Fatima, S.; Hu, X.; Gong, R.-H.; Huang, C.; Chen, M.; Wong, H.L.X.; Bian, Z.; Kwan, H.Y. Palmitic Acid Is an Intracellular Signaling Molecule Involved in Disease Development. Cell. Mol. Life Sci. 2019, 76, 2547–2557. [Google Scholar] [CrossRef]
- Kien, C.L.; Bunn, J.Y.; Ugrasbul, F. Increasing Dietary Palmitic Acid Decreases Fat Oxidation and Daily Energy Expenditure. Am. J. Clin. Nutr. 2005, 82, 320–326. [Google Scholar] [CrossRef]
- Kabagambe, E.K.; Tsai, M.Y.; Hopkins, P.N.; Ordovas, J.M.; Peacock, J.M.; Borecki, I.B.; Arnett, D.K. Erythrocyte Fatty Acid Composition and the Metabolic Syndrome: A National Heart, Lung, and Blood Institute GOLDN Study. Clin. Chem. 2008, 54, 154–162. [Google Scholar] [CrossRef]
- Yu, Y.; Cai, Z.; Zheng, J.; Chen, J.; Zhang, X.; Huang, X.-F.; Li, D. Serum Levels of Polyunsaturated Fatty Acids Are Low in Chinese Men with Metabolic Syndrome, Whereas Serum Levels of Saturated Fatty Acids, Zinc, and Magnesium Are High. Nutr. Res. 2012, 32, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Palomer, X.; Pizarro-Delgado, J.; Barroso, E.; Vázquez-Carrera, M. Palmitic and Oleic Acid: The Yin and Yang of Fatty Acids in Type 2 Diabetes Mellitus. Trends Endocrinol. Metab. TEM 2018, 29, 178–190. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.L.; Zhang, L.; Youker, K.; Zhang, M.-X.; Wang, J.; LeMaire, S.A.; Coselli, J.S.; Shen, Y.H. Free Fatty Acids Inhibit Insulin Signaling-Stimulated Endothelial Nitric Oxide Synthase Activation through Upregulating PTEN or Inhibiting Akt Kinase. Diabetes 2006, 55, 2301–2310. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-P.; Kuo, W.-W.; Baskaran, R.; Day, C.-H.; Chen, R.-J.; Wen, S.-Y.; Ho, T.-J.; Padma, V.V.; Kuo, C.-H.; Huang, C.-Y. Acute Hypoxic Preconditioning Prevents Palmitic Acid-Induced Cardiomyocyte Apoptosis via Switching Metabolic GLUT4-Glucose Pathway Back to CD36-Fatty Acid Dependent. J. Cell. Biochem. 2018, 119, 3363–3372. [Google Scholar] [CrossRef]
- Chen, Y.-P.; Tsai, C.-W.; Shen, C.-Y.; Day, C.-H.; Yeh, Y.-L.; Chen, R.-J.; Ho, T.-J.; Padma, V.V.; Kuo, W.-W.; Huang, C.-Y. Palmitic Acid Interferes with Energy Metabolism Balance by Adversely Switching the SIRT1-CD36-Fatty Acid Pathway to the PKC Zeta-GLUT4-Glucose Pathway in Cardiomyoblasts. J. Nutr. Biochem. 2016, 31, 137–149. [Google Scholar] [CrossRef]
- Huang, J.-P.; Huang, S.-S.; Deng, J.-Y.; Hung, L.-M. Impairment of Insulin-Stimulated Akt/GLUT4 Signaling Is Associated with Cardiac Contractile Dysfunction and Aggravates I/R Injury in STZ-Diabetic Rats. J. Biomed. Sci. 2009, 16, 77. [Google Scholar] [CrossRef]
- Bairwa, S.C.; Parajuli, N.; Dyck, J.R.B. The Role of AMPK in Cardiomyocyte Health and Survival. Biochim. Biophys. Acta 2016, 1862, 2199–2210. [Google Scholar] [CrossRef]
- Yuan, L.; Mao, Y.; Luo, W.; Wu, W.; Xu, H.; Wang, X.L.; Shen, Y.H. Palmitic Acid Dysregulates the Hippo-YAP Pathway and Inhibits Angiogenesis by Inducing Mitochondrial Damage and Activating the Cytosolic DNA Sensor CGAS-STING-IRF3 Signaling Mechanism. J. Biol. Chem. 2017, 292, 15002–15015. [Google Scholar] [CrossRef]
- Knowles, C.J.; Cebova, M.; Pinz, I.M. Palmitate Diet-Induced Loss of Cardiac Caveolin-3: A Novel Mechanism for Lipid-Induced Contractile Dysfunction. PLoS ONE 2013, 8, e61369. [Google Scholar] [CrossRef]
- Tian, D.; Qiu, Y.; Zhan, Y.; Li, X.; Zhi, X.; Wang, X.; Yin, L.; Ning, Y. Overexpression of Steroidogenic Acute Regulatory Protein in Rat Aortic Endothelial Cells Attenuates Palmitic Acid-Induced Inflammation and Reduction in Nitric Oxide Bioavailability. Cardiovasc. Diabetol. 2012, 11, 144. [Google Scholar] [CrossRef]
- Mao, Y.; Luo, W.; Zhang, L.; Wu, W.; Yuan, L.; Xu, H.; Song, J.; Fujiwara, K.; Abe, J.-I.; LeMaire, S.A.; et al. STING-IRF3 Triggers Endothelial Inflammation in Response to Free Fatty Acid-Induced Mitochondrial Damage in Diet-Induced Obesity. Arterioscler. Thromb. Vasc. Biol. 2017, 37, 920–929. [Google Scholar] [CrossRef] [PubMed]
- Trombetta, A.; Togliatto, G.; Rosso, A.; Dentelli, P.; Olgasi, C.; Cotogni, P.; Brizzi, M.F. Increase of Palmitic Acid Concentration Impairs Endothelial Progenitor Cell and Bone Marrow-Derived Progenitor Cell Bioavailability: Role of the STAT5/PPARγ Transcriptional Complex. Diabetes 2013, 62, 1245–1257. [Google Scholar] [CrossRef]
- Briggs, M.A.; Petersen, K.S.; Kris-Etherton, P.M. Saturated Fatty Acids and Cardiovascular Disease: Replacements for Saturated Fat to Reduce Cardiovascular Risk. Healthcare 2017, 5, 29. [Google Scholar] [CrossRef] [PubMed]
- Feingold, K.R. The Effect of Diet on Cardiovascular Disease and Lipid and Lipoprotein Levels. In Endotext; Feingold, K.R., Anawalt, B., Boyce, A., Chrousos, G., de Herder, W.W., Dhatariya, K., Dungan, K., Hershman, J.M., Hofland, J., Kalra, S., et al., Eds.; MDText.com, Inc.: South Dartmouth, MA, USA, 2000. [Google Scholar]
- Zhu, S.; Jiao, W.; Xu, Y.; Hou, L.; Li, H.; Shao, J.; Zhang, X.; Wang, R.; Kong, D. Palmitic Acid Inhibits Prostate Cancer Cell Proliferation and Metastasis by Suppressing the PI3K/Akt Pathway. Life Sci. 2021, 286, 120046. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.-S.; Huang, S.-M.; Wang, Y.-C. Palmitate Enhances the Efficacy of Cisplatin and Doxorubicin against Human Endometrial Carcinoma Cells. Int. J. Mol. Sci. 2021, 23, 80. [Google Scholar] [CrossRef] [PubMed]
- Baumann, J.; Wong, J.; Sun, Y.; Conklin, D.S. Palmitate-Induced ER Stress Increases Trastuzumab Sensitivity in HER2/Neu-Positive Breast Cancer Cells. BMC Cancer 2016, 16, 551. [Google Scholar] [CrossRef]
- Pascual, G.; Domínguez, D.; Elosúa-Bayes, M.; Beckedorff, F.; Laudanna, C.; Bigas, C.; Douillet, D.; Greco, C.; Symeonidi, A.; Hernández, I.; et al. Dietary Palmitic Acid Promotes a Prometastatic Memory via Schwann Cells. Nature 2021, 599, 485–490. [Google Scholar] [CrossRef]
- Pascual, G.; Avgustinova, A.; Mejetta, S.; Martín, M.; Castellanos, A.; Attolini, C.S.-O.; Berenguer, A.; Prats, N.; Toll, A.; Hueto, J.A.; et al. Targeting Metastasis-Initiating Cells through the Fatty Acid Receptor CD36. Nature 2017, 541, 41–45. [Google Scholar] [CrossRef]
- Norgren, J.; Sindi, S.; Sandebring-Matton, A.; Kåreholt, I.; Daniilidou, M.; Akenine, U.; Nordin, K.; Rosenborg, S.; Ngandu, T.; Kivipelto, M. Ketosis After Intake of Coconut Oil and Caprylic Acid—With and Without Glucose: A Cross-Over Study in Healthy Older Adults. Front. Nutr. 2020, 7, 40. [Google Scholar] [CrossRef]
- Altinoz, M.A.; Ozpinar, A.; Seyfried, T.N. Caprylic (Octanoic) Acid as a Potential Fatty Acid Chemotherapeutic for Glioblastoma. Prostaglandins Leukot. Essent. Fatty Acids 2020, 159, 102142. [Google Scholar] [CrossRef]
- Tindall, A.M.; McLimans, C.J.; Petersen, K.S.; Kris-Etherton, P.M.; Lamendella, R. Walnuts and Vegetable Oils Containing Oleic Acid Differentially Affect the Gut Microbiota and Associations with Cardiovascular Risk Factors: Follow-up of a Randomized, Controlled, Feeding Trial in Adults at Risk for Cardiovascular Disease. J. Nutr. 2020, 150, 806–817. [Google Scholar] [CrossRef] [PubMed]
- Sales-Campos, H.; de Souza, P.R.; Peghini, B.C.; da Silva, J.S.; Cardoso, C.R. An Overview of the Modulatory Effects of Oleic Acid in Health and Disease. Mini Rev. Med. Chem. 2013, 13, 201–210. [Google Scholar] [PubMed]
- Malodobra-Mazur, M.; Cierzniak, A.; Dobosz, T. Oleic Acid Influences the Adipogenesis of 3T3-L1 Cells via DNA Methylation and May Predispose to Obesity and Obesity-Related Disorders. Lipids Health Dis. 2019, 18, 230. [Google Scholar] [CrossRef] [PubMed]
- Banim, P.J.; Luben, R.; Khaw, K.-T.; Hart, A.R. Dietary Oleic Acid Is Inversely Associated with Pancreatic Cancer—Data from Food Diaries in a Cohort Study. Pancreatol. Off. J. Int. Assoc. Pancreatol. IAP Al 2018, 18, 655–660. [Google Scholar] [CrossRef]
- Calder, P.C. Functional Roles of Fatty Acids and Their Effects on Human Health. J. Parenter. Enter. Nutr. 2015, 39, 18S–32S. [Google Scholar] [CrossRef]
- Choi, C.W.; Kim, S.C.; Hwang, S.S.; Choi, B.K.; Ahn, H.J.; Lee, M.Y.; Park, S.H.; Kim, S.K. Antioxidant Activity and Free Radical Scavenging Capacity between Korean Medicinal Plants and Flavonoids by Assay-Guided Comparison. Plant Sci. 2002, 163, 1161–1168. [Google Scholar] [CrossRef]
- Chu, W.-L.; Lim, Y.-W.; Radhakrishnan, A.K.; Lim, P.-E. Protective Effect of Aqueous Extract from Spirulina platensis against Cell Death Induced by Free Radicals. BMC Complement. Altern. Med. 2010, 10, 53. [Google Scholar] [CrossRef]
- Junpeng, J.; Xupeng, C.; Miao, Y.; Song, X. Monogalactosyldiacylglycerols with High PUFA Content from Microalgae for Value-Added Products. Appl. Biochem. Biotechnol. 2020, 190, 1212–1223. [Google Scholar] [CrossRef]
- Jung, C.H.G.; Braune, S.; Waldeck, P.; Küpper, J.-H.; Petrick, I.; Jung, F. Morphology and Growth of Arthrospira platensis during Cultivation in a Flat-Type Bioreactor. Life 2021, 11, 536. [Google Scholar] [CrossRef]
- Angelis, S.; Novak, A.C.; Sydney, E.B.; Soccol, V.T.; Carvalho, J.C.; Pandey, A.; Noseda, M.D.; Tholozan, J.L.; Lorquin, J.; Soccol, C.R. Co-Culture of Microalgae, Cyanobacteria, and Macromycetes for Exopolysaccharides Production: Process Preliminary Optimization and Partial Characterization. Appl. Biochem. Biotechnol. 2012, 167, 1092–1106. [Google Scholar] [CrossRef]


| Component Class | Component | Amount per 100 g Dry Weight | |
|---|---|---|---|
| Nonessential amino acids | Alanine | 4.0–5.0 | g |
| Arginine | 3.0–5.0 | g | |
| Aspartic acid | 1.5–5.9 | g | |
| Cystine | 0.5–0.75 | g | |
| Glutamic acid | 6.0–9.1 | g | |
| Glycine | 2.0–4.0 | g | |
| Proline | 2.0–3.0 | g | |
| Serine | 2.7–4.5 | g | |
| Tyrosine | 1.0–3.0 | g | |
| Essential amino acids | Histidine | 0.5–1.5 | g |
| Isoleucine | 3.0–4.0 | g | |
| Leucine | 3.0–6.0 | g | |
| Lysine | 2.9–6.0 | g | |
| Methionine | 1.0–6.0 | g | |
| Phenylalanine | 2.5–3.5 | g | |
| Threonine | 1.5–3.0 | g | |
| Tryptophan | 0.9–2.0 | g | |
| Valine | 1.0–3.5 | g | |
| Non-proteinogenic amino acids | Theanine | 2.97–n.a. | g |
| Component | Amount per 100 G Dry Weight | |
|---|---|---|
| Galactose | 3 | g |
| Glucose | 54.4 | g |
| Mannose | 9.3 | g |
| Rhamnose | 22.3 | g |
| Xylose | 7 | g |
| Component | Amount per 100 g Dry Weight | ||
|---|---|---|---|
| Biotin | 0.005–n.a. | mg | |
| Folic acid | 0.05–0.30 | mg | |
| Inositol | 70–90 | mg | |
| Vitamin A * | (Provitamin A as beta-Carotene) | 150–250 | mg |
| Vitamin B1 | (Thiamine) | 1.5–4.0 | mg |
| Vitamin B2 | (Riboflavin) | 3–5 | mg |
| Vitamin B3 | (Niacin) | 10–25 | mg |
| Vitamin B6 | (Pyridoxine) | 0.5–0.7 | mg |
| Vitamin B12 | (Cobalamin) | 0.05–2.0 | mg |
| Vitamin E | (Tocopherol) | 5–20 | mg |
| Vitamin K | 0.90–1.05 | mg | |
| Neutral Lipids | Glycolipids | Phospholipids |
|---|---|---|
| Free fatty acids | Cerebrosides | Phosphatidylcholine |
| Free sterols | Digalactosyl diacylglycerol | Phosphatidylethanolamine |
| Diacylglycerols | Esterified steryl glucoside | Phosphatidylinositol |
| Monoacylglycerols | Monogalactosyl diacylglycerol | Phosphatidylserine |
| Sterol esters | Steryl glucoside | |
| Triacylglycerols | Sulphoquinovosyl diacylglycerol |
| Component Class | Component | Amount per 100 g Dry Weight | |
|---|---|---|---|
| Saturated fatty acids (1.95 g per 100 g dry weight) | in total | 33.68–66.75 | % 1 |
| Pentadecenoic | 1.26–3.16 | % 1 | |
| Pentadecanoic acid | 0.70–1.53 | % 1 | |
| Caprylic acid | 3.65–3.73 | % 1 | |
| Palmitic acid | 18.00–46.07 | % 1 | |
| Stearic acid | 0.95–1.41 | % 1 | |
| Polyunsaturated fatty acids (1.93 g per 100 g dry weight) | in total | 28.2–47.8 | % 1 |
| Linoleic acid | 16.18–17.43 | % 1 | |
| γ-Linolenic acid | 8.87–21.73 | % 1 | |
| Hexadecadienoic acid | 2.43–3.38 | % 1 | |
| Monounsaturated fatty acids (0.26 g per 100 g dry weight) | in total | n.a. | |
| Palmitoleic | 1.00–1.50 | % 1 | |
| Oleic acid | 1.97–5.23 | % 1 | |
| Trans fatty acids | in total | 0.3–0.5 | % 1 |
| Cholesterol | in total | <0.1 | mg |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jung, F.; Braune, S.; Jung, C.H.G.; Krüger-Genge, A.; Waldeck, P.; Petrick, I.; Küpper, J.-H. Lipophilic and Hydrophilic Compounds from Arthrospira platensis and Its Effects on Tissue and Blood Cells—An Overview. Life 2022, 12, 1497. https://doi.org/10.3390/life12101497
Jung F, Braune S, Jung CHG, Krüger-Genge A, Waldeck P, Petrick I, Küpper J-H. Lipophilic and Hydrophilic Compounds from Arthrospira platensis and Its Effects on Tissue and Blood Cells—An Overview. Life. 2022; 12(10):1497. https://doi.org/10.3390/life12101497
Chicago/Turabian StyleJung, Friedrich, Steffen Braune, Conrad H. G. Jung, Anne Krüger-Genge, Peter Waldeck, Ingolf Petrick, and Jan-Heiner Küpper. 2022. "Lipophilic and Hydrophilic Compounds from Arthrospira platensis and Its Effects on Tissue and Blood Cells—An Overview" Life 12, no. 10: 1497. https://doi.org/10.3390/life12101497
APA StyleJung, F., Braune, S., Jung, C. H. G., Krüger-Genge, A., Waldeck, P., Petrick, I., & Küpper, J.-H. (2022). Lipophilic and Hydrophilic Compounds from Arthrospira platensis and Its Effects on Tissue and Blood Cells—An Overview. Life, 12(10), 1497. https://doi.org/10.3390/life12101497