
Chronothyroidology: Chronobiological Aspects in Thyroid Function and Diseases

 ,
,  ,
, 
Abstract
1. Introduction
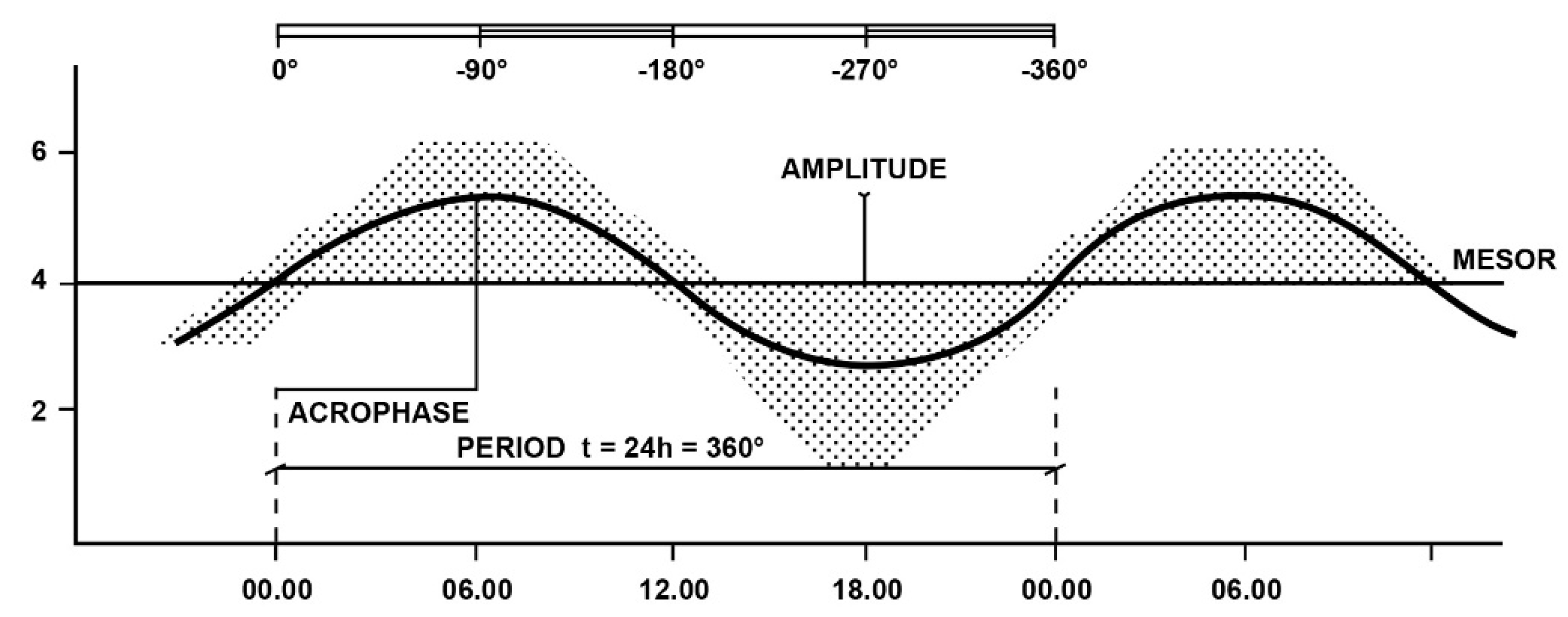
2. Elementary Glossary of Chronobiology
2.1. Main Characteristics of Biological Rhythms
2.2. Synchronizing Factors of Rhythms
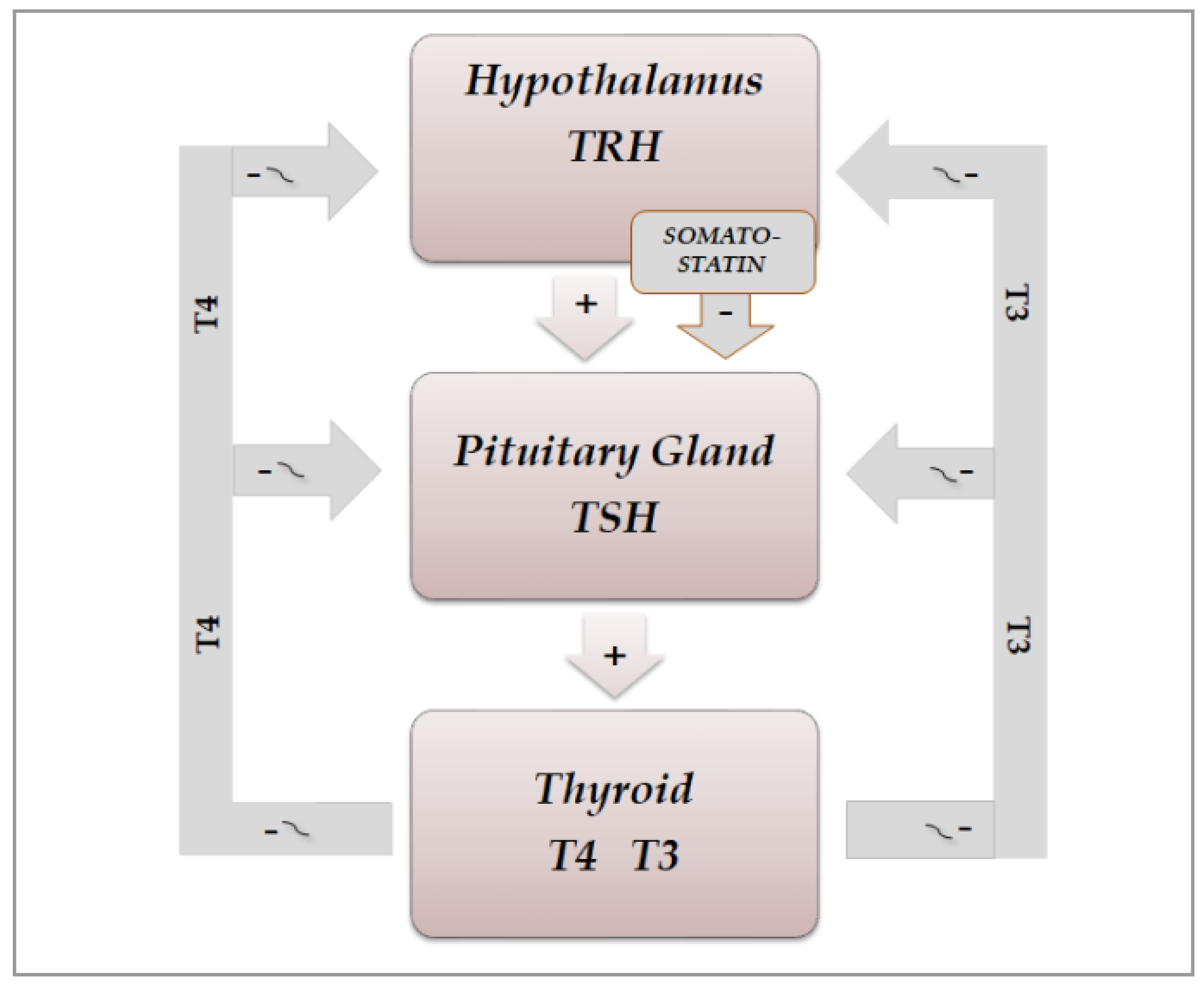
3. Chronobiology and Hypothalamic–Pituitary–Thyroid Axis
3.1. Physiological Aspects
3.2. Chrono-Organization of Hypothalamic–Pituitary–Thyroid Axis
3.2.1. Physiological Aspects
3.2.2. Pathophysiological Aspects
3.3. Light/Darkness and Thyroid Function
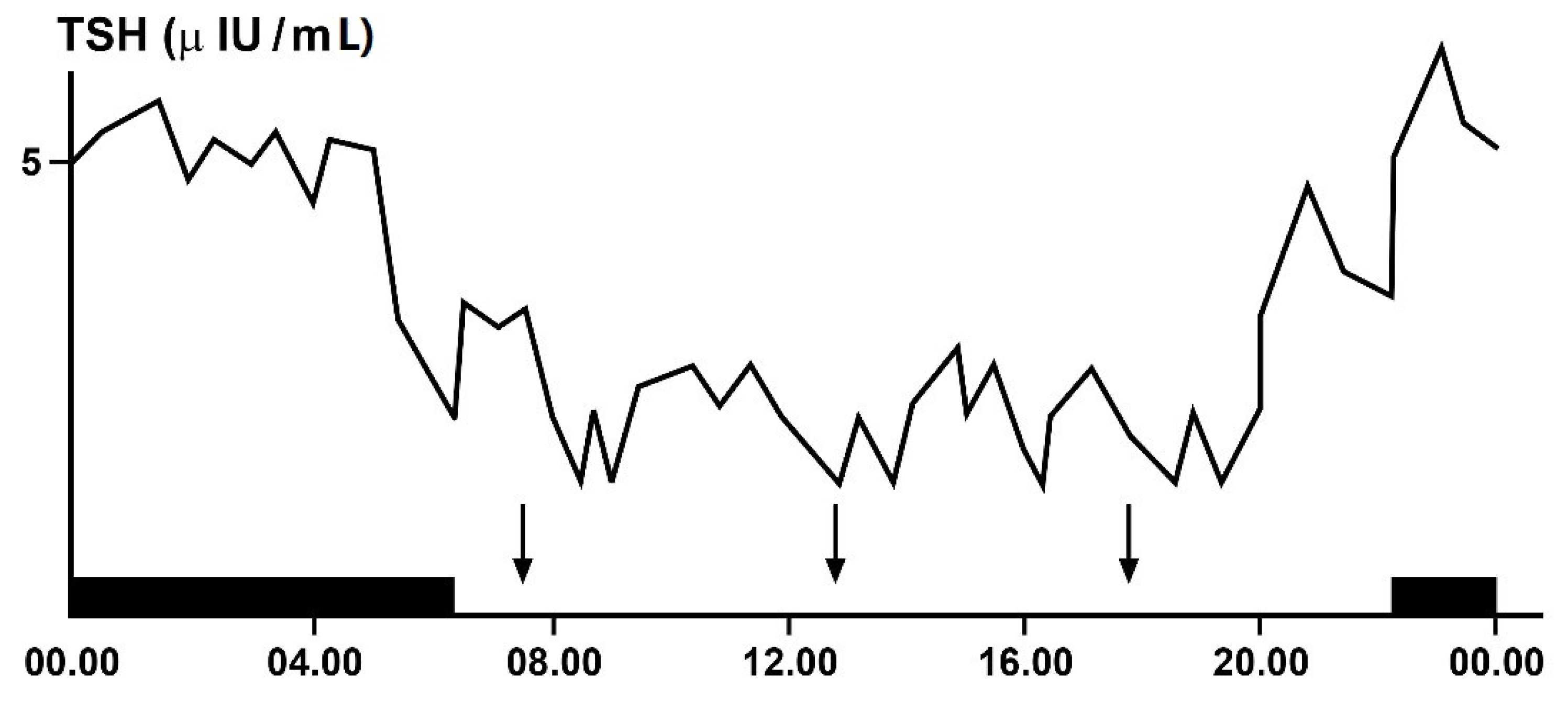
3.4. Ultradian Rhythms of TSH and Thyroid Hormones
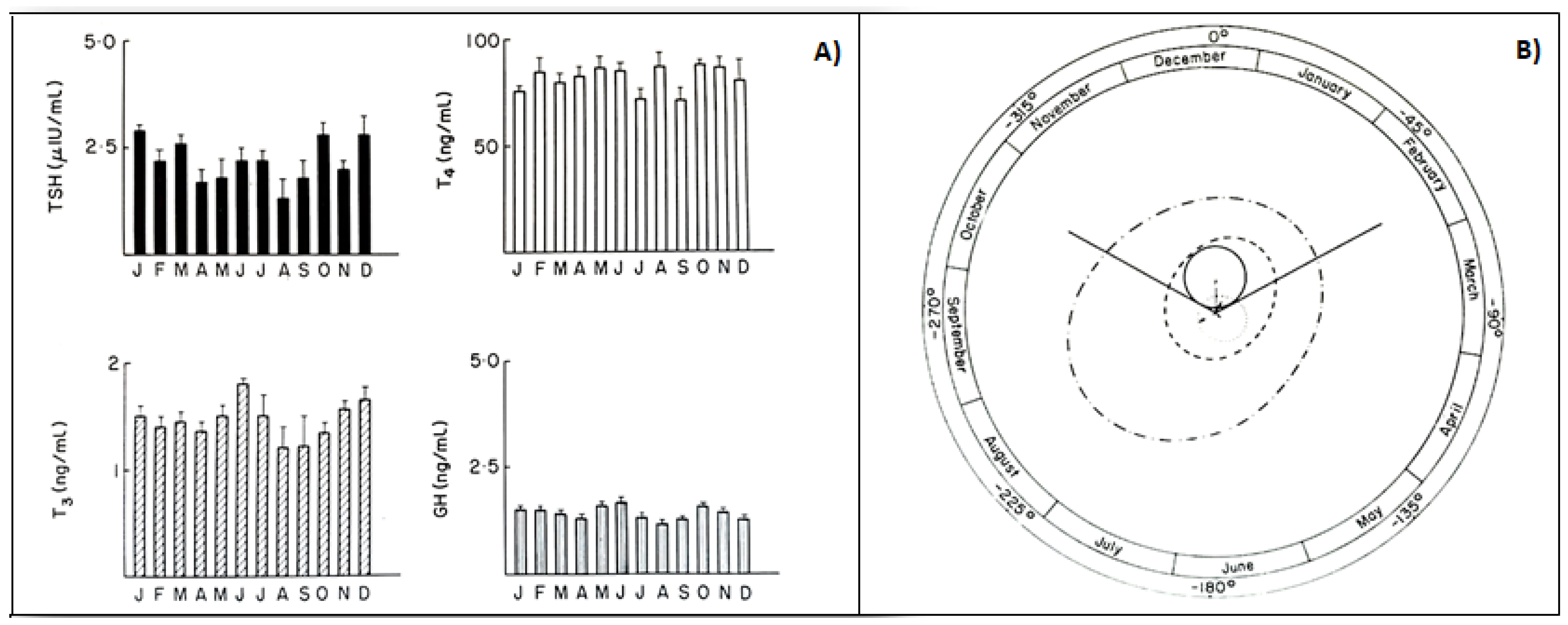
3.5. Infradian Rhythms: Seasonality of HPT Axis Secretions
4. Interrelationship between Disorders of Chrono-Organization and Thyroid Diseases
4.1. Autoimmune Thyroid Diseases and Circadian System
4.2. Thyroid Dysfunctions and Circadian Clock
4.3. Circadian Clock and Thyroid Malignancy
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Touitou, Y.; Haus, E. (Eds.) Biological rhythms from biblical to modern times. In Biologic Rhythms in Clinical and Laboratory Medicine; Springer: Berlin/Heidelberg, Germany, 1992; pp. 1–5. [Google Scholar]
- Allada, R.; Bass, J. Circadian mechanisms in Medicine. N. Engl. J. Med. 2021, 384, 550–561. [Google Scholar] [CrossRef]
- Burki, T. Nobel Prize awarded for discoveries in circadian rhythm. Lancet 2017, 390, e31. [Google Scholar] [CrossRef]
- Darwin, C.; Darwin, F. The Power of Movement in Plants; John Murray: London, UK, 1880; pp. 407–408. [Google Scholar]
- Halberg, F.; Katinas, G.S.; Chiba, Y.; Garcia-Sainz, M.; Keovatis, T.G.; Kunkel, H.; Montalbetti, N.; Reinberg, A.; Scharf, R.; Simpson, H. Chronobiologic glossary of the International Society for the study of biological rhythms. Int. J. Chronobiol. 1973, 1, 31–63. [Google Scholar] [PubMed]
- Konopka, R.J.; Benzer, S. Clock Mutants of Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 1971, 68, 2112–2116. [Google Scholar] [CrossRef]
- Hardin, P.E.; Hall, J.C.; Rosbash, M. Feedback of the Drosophila period gene product on circadian cycling of its messenger RNA levels. Nat. Cell Biol. 1990, 343, 536–540. [Google Scholar] [CrossRef]
- King, D.P.; Zhao, Y.; Sangoram, A.M.; Wilsbacher, L.D.; Tanaka, M.; Antoch, M.P.; Steeves, T.D.L.; Vitaterna, M.H.; Kornhauser, J.M.; Lowrey, P.L.; et al. Positional cloning of the mouse circadian clock gene. Cell 1997, 89, 641–653. [Google Scholar] [CrossRef]
- Patke, A.; Young, M.W.; Axelrod, S. Molecular mechanisms and physiological importance of circadian rhythms. Nat. Rev. Mol. Cell Biol. 2020, 21, 67–84. [Google Scholar] [CrossRef]
- Haus, E.; Touitou, Y. Principles of clinical chronobiology. In Biologic Rhythms in Clinical and Laboratory Medicine; Touitou, Y., Haus, E., Eds.; Springer: Berlin/Heidelberg, Germany, 1992; pp. 6–34. [Google Scholar]
- Haus, E.; Lakatua, D.J.; Sackett-Lunden, L.; Swoyer, J. Chronobiology in laboratory medicine. In Clinical Aspects of Chronobiology; Reitveld, W.T., Ed.; Bakker: Baarn, The Netherlands, 1984; pp. 13–82. [Google Scholar]
- Nelson, W.; Tong, Y.L.; Lee, J.K.; Halberg, F. Methods for cosinor rhythmometry. Chronobiologia 1979, 6, 305–323. [Google Scholar] [PubMed]
- Bellastella, G.; De Bellis, A.; Maiorino, M.I.; Paglionico, V.A.; Esposito, K.; Bellastella, A. Endocrine rhythms and sport: It is time to tak e time into account. J. Endocrinol. Investig. 2019, 42, 1137–1147. [Google Scholar] [CrossRef]
- Kalsbeek, A.; Fliers, E. Circadian and endocrine rhythms. Best. Pract. Res. Clin. Endocrinol. Metab. 2017, 31, 443–449. [Google Scholar] [CrossRef]
- Bae, S.A.; Androulakis, I.P. The synergistic role of light-feeding phase relation on entraining robust circadian rhythms in the periphery. Gene Regul. Syst. Biol. 2017, 11. [Google Scholar] [CrossRef]
- Greco, C.M.; Sassone-Corsi, P. Circadian blueprint of metabolic patways in the brain. Nat. Rev. Neurosci. 2018. [Google Scholar] [CrossRef]
- Doi, M.; Hirayama, J.; Sassone-Corsi, P. Circadian regulator CLOCK is a histone acetyltransferase. Cell 2006, 125, 497–508. [Google Scholar] [CrossRef]
- Bellastella, A.; De Bellis, A.; Bellastella, G.; Esposito, K. Opposite influence of light and blindness on pituitary-gonadal secretions. Front. Endocrinol. 2014. [Google Scholar] [CrossRef]
- Brezinski, A. Melatonin in human. N. Engl. J. Med. 1997, 16, 186–195. [Google Scholar] [CrossRef]
- Masri, S.; Kinouchi, K.; Sassone-Corsi, P. Circadian clocks, epigenetics, and cancer. Curr. Opin. Oncol. 2015, 27, 50–56. [Google Scholar] [CrossRef]
- Wergzyn, L.R.; Tamimi, R.M.; Rosner, B.A.; Brown, S.B.; Stevens, R.G.; Eliasse, A.H.; Laden, F.; Willett, W.C.; Hankinson, S.E.; Schernhammer, E.S. Rotating night shifts and risk of breast cancer in the Nurses’ Health Study. Am. J. Epidemiol. 2017, 186, 532–540. [Google Scholar]
- Czeisler, C.A.; Shanahan, T.L.; Klerman, E.B.; Martens, H.; Brotman, D.J.; Ermens, J.S.; Klein, T.; Rizzo, J.F. Suppression of melatonin secretion in some blind patients by exposure to bright light. N. Engl. Med. 1995, 332, 6–11. [Google Scholar] [CrossRef]
- D’Alessandro, B.; Bellastella, A.; Esposito, V.; Colucci, C.F.; Montalbetti, N. Circadian rhythm of cortisol secretion in elderly and blind subjects. Br. Med. J. 1974, 2, 274–276. [Google Scholar] [CrossRef] [PubMed]
- Bellastella, A.; Pisano, G.; Iorio, S.; Pasquali, D.; Orio, F.; Venditto, T.; Sininsi, A.A. Endocrine secretions under abnormal light-dark cycles and in the blind. Horm. Res. 1998, 49, 153–157. [Google Scholar] [CrossRef]
- Halberg, F.; Haus, E.; Cardoso, S.S.; Scheving, L.E.; Khul, J.F.W.; Shiotsuka, R.; Rosen, G.; Pauly, J.E.; Runge, W.; Spalding, J.F.L. Toward a chronotherapy of neoplasia: Tolerance of treatment depends upon host rhythms. Experientia 1973, 29, 909–934. [Google Scholar] [CrossRef]
- Salem, A.H.; Koening, D.; Carlson, D. Pooled population pharmacokinetic analysis of phase I, II and III studies of linifanib in cancer patients. Clin. Pharm. 2014, 53, 347–359. [Google Scholar] [CrossRef]
- Vera, L.M.; Bello, C.; Paredes, J.F.; Carmona-Antonanczas, G.; Sanchez-Vazquez, F.J. Ethanol toxicity differs depending on the time of day. PLoS ONE 2018, 13, e0190406. [Google Scholar] [CrossRef]
- Fekete, C.; Lechan, R.M. Central regulation of hypothalamic-pituitary-thyroid axis under physiological and pathophysiological conditions. Endocr. Rev. 2014, 35, 159–194. [Google Scholar] [CrossRef]
- Vassart, G.; Dumont, G.E. The thyrotropin receptor and the regulation of thyrocyte function and growth. Endocr. Rev. 1992, 13, 596–611. [Google Scholar]
- Ikegami, K.; Refetoff, S.; Van Cauter, E.; Yoshimura, T. Interconnection between circadian clocks and thyroid function. Nat. Rev. Endocrinol. 2019, 15, 590–600. [Google Scholar] [CrossRef]
- Patel, Y.C. Somatostatin and its receptor family. Front. Neuroendocrnol. 1999, 20, 157–198. [Google Scholar] [CrossRef]
- Thoss, V.S.; Perez, J.; Probst, A.; Hoyer, D. Expression of five somatostatin receptor mRNAs in the human brain and pituitary. Naunyn Schmiedebergs Arch. Pharm. 1996, 354, 411–419. [Google Scholar] [CrossRef]
- Ortiga-Carvalho, T.M.; Chiamolera, M.I.; Pazos-Moura, C.C.; Wondisford, F.E. Hypothalamic-pituitary-thyroid axis. Compr. Physiol. 2016, 6, 1387–1428. [Google Scholar]
- Ikegami, K.; Yoshimura, T. The hypothalamic-pituitary-thyroid axis and biological rhythm: The discovery of TSH’s unexpected role using animal models. Best Pract. Res. Clin. Endocrinol. 2017, 31, 475–485. [Google Scholar] [CrossRef]
- Van Cauter, E.; Spiegel, K. Circadian and sleep control of hormonal secretions. In Regulation of Sleep and Circadian Rhythms; Turek, F.W., Zee, P.C., Eds.; Marcel Dekker, Inc.: New York, NY, USA, 1999; pp. 397–426. [Google Scholar]
- Spiegel, K.; Leprout, R.; Van Cauter, E. Impact of sleep debt on metabolic and endocrine functions. Lancet 1999, 354, 1435–1439. [Google Scholar] [CrossRef]
- Kalsbeek, A.; Fliers, E.; Franke, A.N.; Wortel, J.; Buijs, R.M. Functional connections between the suprachiasmatic nucleus and the thyroid gland as revealed by lesioning and viral tracing techniques in the rat. Endocrinology 2000, 141, 3832–3841. [Google Scholar] [CrossRef]
- Leso, V.; Vetrani, I.; Sicignano, A.; Romano, R.; Iavicoli, I. The impact of shift-work and night shift-work on thyroid: A systematic review. Int. J. Envir. Res. Public Health 2020, 17, 1527. [Google Scholar] [CrossRef]
- Roelfsema, F.; Boelen, A.; Kalsbeek, A.; Fliers, E. Regulatory aspects of the human hypothalamus-pituitary-thyroid axis. Best Pract. Res. Clin. Endocrinol. 2017, 31, 487–503. [Google Scholar] [CrossRef]
- Bartalena, L.; Martino, E.; Petrini, L.; Velluzzi, F.; Loviselli, A.; Grasso, L.; Mammoli, C.; Pinchera, A. The nocturnal serum thyrotropin surge is abolished in patients with adrenocorticotropin (ACTH)-dependent or ACTH-independent Cushing’s syndrome. J. Clin. Endocrinol. Metab. 1991, 72, 1195–1199. [Google Scholar] [CrossRef]
- Bellastella, G.; Maiorino, M.I.; Scappaticcio, L.; Casciano, O.; Petrizzo, M.; Caputo, M.; Paglionico, V.A.; Giugliano, D.; Esposito, K. TSH oscillations in young patients with type 1 diabetes may be due to glycemic variabilità. J. Endocrinol. Investig. 2017. [Google Scholar] [CrossRef]
- Selmaoui, B.; Lambrozo, J.; Touitou, Y. Endocrine functions in young men exposed for one night to a 50-Hz magnetic field. A circadian study of pituitary, thyroid and adrenocortical hormones. Life Sci. 1997, 61, 473–486. [Google Scholar] [CrossRef]
- Kunt, H.; Sentuk, I.; Gonul, Y.; Korkmaz, M.; Ahsen, A.; Hazman, O.; Bal, A.; Genc, A.; Songur, A. Effect of electromagnetic radiation exposure on bone mineral density, thyroid, and oxidative stress index in electrical workers. OncoTargets Ther. 2016, 9, 745–754. [Google Scholar]
- Singh, D.V.; Turner, C.W. Effect of light and darkness upon thyroid secretion rate and on the endocrine glands of female rats. Proc. Soc. Exp. Biol. Med. 1969, 131, 1296–1301. [Google Scholar] [CrossRef]
- Martino, E.; Seo, H.; Lernmark, A.; Refetoff, S. Ontogenetic pattern of thyrotropin-releasing hormone-like material in rat hypothalamus, pancreas and retina. Proc. Nat. Acad. Sci. USA 1980, 77, 4345–4350. [Google Scholar] [CrossRef]
- Petterborg, L.J.; Vaughan, M.K.; Johnson, L.Y.; Champney, T.H.; Reiter, R.J. Modification of testicular and thyroid function by chronic exposure to short photoperiod: A comparison in four rodent species. Comp. Biochem. Physiol. 1984, 78, 31–37. [Google Scholar] [CrossRef]
- Sharp, P.J.; Klandorf, H.; Lea, R.W. Influence of lighting cyclesz on daily rhythms in concentrations of plasma triiodotyronine and thyroxine in intact and pinealectomized immature broiler hens (Gallus domesticus). J. Endocrinol. 1984, 193, 337–341. [Google Scholar] [CrossRef]
- Singh, D.V.; Narang, G.D.; Turner, C.W. Effect of melatonin and its withdrawal on thyroid hormone secretion rate of female rats. J. Endocrinol. 1969, 43, 489–494. [Google Scholar] [CrossRef]
- Bellastella, A.; Amato, G.; Bizzarro, A.; Carella, C.; Criscuolo, T.; Iorio, S.; Pisano, G.; Sinisi, A.A.; De Bellis, A. Light, blindness and endocrine scretions. J. Endocrinol. Investig. 1999, 22, 874–885. [Google Scholar] [CrossRef] [PubMed]
- Juvone, P.M. Development of melatonin synthesis in chicken retina. Regulation of serotonin-acetyltransferase activity by light circadian oscillators and cyclic AMP. J. Neurochem. 1990, 54, 1562–1566. [Google Scholar]
- Pang, S.F.; Lee, P.P.; Tang, C.L. Sensory receptors as a special class of hormonal cells. Neuroendocrinology 1991, 53, 2–11. [Google Scholar] [CrossRef]
- Tosini, G.; Menaker, M. Circadian rhythms in cultured mammalian retina. Science 1996, 272, 419–421. [Google Scholar] [CrossRef] [PubMed]
- Bellastella, A.; Criscuolo, T.; Sinisi, A.A.; Iorio, S.; Mazzuca, A.; Parlato, F.; Perrone, L.; Faggiano, M. Plasma Thyrotropin, thyroxine, triiodothyronine, free thyroxine, free triiodothyronine and cortisol levels in blind prepubertal boys. J. Endocrinol. Investig. 1988, 11, 171–174. [Google Scholar] [CrossRef]
- Wartofsky, L.; Burman, K.D. Alteration in patients with sistemi illness: The “euthyroid sick syndrome”. Endocr. Rev. 1982, 3, 164–172. [Google Scholar] [CrossRef]
- Refetoff, S.; Dewind, L.T.; De Groot, L.J. Familial syndrome combining deaf-mutism, stippled epiphyses, goiter and abnormally high P.B.I.: Possible refractoriness to thyroid hormone. J. Clin. Endocrinol. Metab. 1967, 27, 279–283. [Google Scholar] [CrossRef]
- Refetoff, S.; Weiss, R.E.; Usala, S.J. The syndromes of resistance to thyroid hormone. Endocr. Rev. 1993, 14, 348–399. [Google Scholar]
- Bellastella, A.; Sinisi, A.A.; Criscuolo, T.; De Bellis, A.; Carella, C.; Iorio, S.; Sinisi, A.M.; Parlato, F.; Venditto, T.; Pisano, G. Melatonin and the pituitary-thyroid axis status in blind adults: A possible resetting after puberty. Clin. Endocrinol. 1995, 43, 707–711. [Google Scholar] [CrossRef] [PubMed]
- Fliers, E.; Wiersinga, W.M.; Swaab, D.F. Physiological and pathophysiological aspects of thyrotropin-releasing hormone gene expression in the human hypothalamus. Thyroid 1998, 8, 921–928. [Google Scholar] [CrossRef]
- Haus, E. Chronobiology in the endocrine system. Adv. Drug Deliv. Rev. 2007, 59, 985–1014. [Google Scholar] [CrossRef]
- Weeke, J.; Gundersen, H.J. Circadian and 30 min variations in serum TSH and thyroid hormones in normal subjects. Eur. J. Endocrinol. 1999, 89, 659–672. [Google Scholar] [CrossRef]
- Romijn, J.A.; Adriaanse, R.; Brabant, G.; Prank, K.; Endert, E.; Wiersinga, W.M. Pulsatile Secretion of Thyrotropin during Fasting: A Decrease of Thyrotropin Pulse Amplitude. J. Clin. Endocrinol. Metab. 1990, 70, 1631–1636. [Google Scholar] [CrossRef]
- Smals, A.; Ross, H.; Kloppenborg, P. Seasonal variation in serum t3 and t4 levels in man. J. Clin. Endocrinol. Metab. 1977, 44, 998–1001. [Google Scholar] [CrossRef]
- Palinkas, L.; Reed, H.; Reedy, K.R.; Van Do, N.; Case, H.; Finney, N.S. Circannual pattern of hypothalamic–pituitary–thyroid (HPT) function and mood during extended antarctic residence. Psychoneuroendocrinology 2001, 26, 421–431. [Google Scholar] [CrossRef]
- De Miera, C.S.; Bothorel, B.; Jaeger, C.; Simonneaux, V.; Hazlerigg, D. Maternal photoperiod programs hypothalamic thyroid status via the fetal pituitary gland. Proc. Natl. Acad. Sci. USA 2017, 114, 8408–8413. [Google Scholar] [CrossRef]
- Peschke, E.; Peschke, D.; Peil, J.; Mess, B. Circannual Oscillations of Thyroxine and Cholesterol Levels After Pinealectomy and Ganglionectomy Related to the Weight Changes of the Pineal, Pituitary, and Thyroid Glands. J. Pineal Res. 1988, 5, 191–202. [Google Scholar] [CrossRef]
- Nicolau, G.Y.; Dumitriu, L.; Plinga, L.; Petrescu, E.; Sacckett-Lundeen, L.; Lakatua, D.J.; Haus, E. Circadian and circannual variations of thyroid function in children +/−1.5 years of age with and without endemic goiter. Prog. Clin. Biol. Res. 1987, 2278, 229–247. [Google Scholar]
- Bellastella, A.; Criscuolo, T.; Mango, A.; Perrone, A.M.L.; Sinisi, A.A.; Faggiano, M. Circannual rhythms of plasma growth hormone, thyrotropin and thyroid hormones in prepuberty. Clin. Endocrinol. 1984, 20, 531–537. [Google Scholar] [CrossRef]
- Bellastella, A.; Criscuolo, T.; Sinisi, A.A.; Rinaldi, A.; Faggiano, M. Circannual Variations of Plasma Thyrotropin in Klinefelter’s Syndrome. Neuroendocrinology 1986, 43, 132–134. [Google Scholar] [CrossRef]
- Yoshihara, A.; Noh, J.Y.; Watanabe, N.; Iwaku, K.; Kunii, Y.; Ohye, H.; Suzuki, M.; Matsumoto, M.; Suzuki, N.; Sugino, K.; et al. Seasonal Changes in Serum Thyrotropin Concentrations Observed from Big Data Obtained During Six Consecutive Years from 2010 to 2015 at a Single Hospital in Japan. Thyroid 2018, 28, 429–436. [Google Scholar] [CrossRef]
- Lange, T.; Dimitrov, S.; Born, J. Effects of sleep and circadian rhythm on the human immune system. Ann. N. Y. Acad. Sci. 2010, 1193, 48–59. [Google Scholar] [CrossRef] [PubMed]
- Bollinger, T.; Bollinger, A.; Oster, H.; Solbach, W. Sleep, Immunity, and Circadian Clocks: A Mechanistic Model. Gerontology 2010, 56, 574–580. [Google Scholar] [CrossRef]
- Hergenhan, S.; Holtkamp, S.; Scheiermann, C. Molecular Interactions Between Components of the Circadian Clock and the Immune System. J. Mol. Biol. 2020, 432, 3700–3713. [Google Scholar] [CrossRef] [PubMed]
- Bargi-Souza, P.; Peliciari-Garcia, R.A.; Nunes, M.T. Disruption of the Pituitary Circadian Clock Induced by Hypothyroidism and Hyperthyroidism: Consequences on Daily Pituitary Hormone Expression Profiles. Thyroid 2019, 29, 502–512. [Google Scholar] [CrossRef]
- Custro, N.; Scafidi, V.; Notarbartolo, A. Pituitary resistance to thyroid hormone action with preserved circadian rhythm of thyrotropin in a postmenopausal woman. J. Endocrinol. Investig. 1992, 15, 121–126. [Google Scholar] [CrossRef]
- Roelfsema, F.; Pereira, A.M.; Keena, D.M.; Veldhuis, J.D.; Romjin, J.A. Thyrotrpin secretion by thyrotropinomas is characterized by increase pulse frequency, delayed and disorderliness. J. Clin. Endocrinol. Metab. 2008, 93, 4052–4057. [Google Scholar] [CrossRef]
- Mannic, T.; Meyer, P.; Triponez, F.; Pusztaszeri, M.; Le Martelot, G.; Mariani, O.; Schmitter, D.; Sage, D.; Philippe, J.; Dibner, C. Circadian Clock Characteristics Are Altered in Human Thyroid Malignant Nodules. J. Clin. Endocrinol. Metab. 2013, 98, 4446–4456. [Google Scholar] [CrossRef]
- Jensen, L.D. The circadian clock and hypoxia in tumor cell de-differentiation and metastasis. Biochim. Biophys. Acta Gen. Subj. 2015, 1850, 1633–1641. [Google Scholar] [CrossRef]
- Philippe, J.; Dibner, C. Thyroid circadian timing: Roles in physiology and thyroid malignancies. J. Biol. Rhythms 2015, 10, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Angelousi, A.; Kassi, E.; Ansari-Nasiri, N.; Randeva, H.; Kaltsas, G. Clock genes and cancer development in particolar in endocrine tissues. Endocr. Relat. Cancer 2019, 26, R305–R317. [Google Scholar] [CrossRef] [PubMed]
- Malaguarnera, R.; Ledda, C.; Filippello, A.; Frasca, F.; Francavilla, V.C.; Ramaci, T.; Parisi, M.C.; Rapisarda, V.; Piro, S. Thyroid cancer and circadian clock disruption. Cancer 2020, 12, 3109. [Google Scholar] [CrossRef]
- Yang, J.; Wang, B.; Chen, H.; Chen, X.; Li, J.; Chen, Y.; Yuan, D.; Zheng, S. Thyrotroph embryonic factor is downregulated in bladder cancer and suppresses proliferation and tuumorigenesis via the AKT/FOXOs signaling pathway. Cell Prolif. 2019, 52, e12560. [Google Scholar] [CrossRef]
- Touitou, Y.; Reinberg, A.; Touitou, D. Association between light at night, melatonin secretion, sleep deprivation, and the internal clock: Health impacts and mechanisms of circadian disruption. Life Sci. 2017, 173, 94–106. [Google Scholar] [CrossRef]
- Wang, Q.; Yin, Y.; Zhang, W. Ghrelin restores the disruption of the circadian clock in steatotic liver. Int. J. Mol. Sci. 2018, 19, 3134. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Light/darkness cycle |
| Sleep/wake alternations |
| Periodic food intake |
| Social environment |
| Physical and mental work |
| Appropriate energy variability |
| Individual chronotype |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bellastella, G.; Maiorino, M.I.; Scappaticcio, L.; De Bellis, A.; Mercadante, S.; Esposito, K.; Bellastella, A. Chronothyroidology: Chronobiological Aspects in Thyroid Function and Diseases. Life 2021, 11, 426. https://doi.org/10.3390/life11050426
Bellastella G, Maiorino MI, Scappaticcio L, De Bellis A, Mercadante S, Esposito K, Bellastella A. Chronothyroidology: Chronobiological Aspects in Thyroid Function and Diseases. Life. 2021; 11(5):426. https://doi.org/10.3390/life11050426
Chicago/Turabian StyleBellastella, Giuseppe, Maria Ida Maiorino, Lorenzo Scappaticcio, Annamaria De Bellis, Silvia Mercadante, Katherine Esposito, and Antonio Bellastella. 2021. "Chronothyroidology: Chronobiological Aspects in Thyroid Function and Diseases" Life 11, no. 5: 426. https://doi.org/10.3390/life11050426
APA StyleBellastella, G., Maiorino, M. I., Scappaticcio, L., De Bellis, A., Mercadante, S., Esposito, K., & Bellastella, A. (2021). Chronothyroidology: Chronobiological Aspects in Thyroid Function and Diseases. Life, 11(5), 426. https://doi.org/10.3390/life11050426