
Inhibition of HDACs Suppresses Cell Proliferation and Cell Migration of Gastric Cancer by Regulating E2F5 Targeting BCL2

 , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture, Treatment, and Transfection
2.2. RNA Extraction, cDNA Synthesis, and qRT-PCR
2.3. Cell Cycle Analysis
2.4. Western Blot Analysis
2.5. Wound-Healing Assay
2.6. Cell Viability Assay
2.7. Statistical Analysis
3. Results
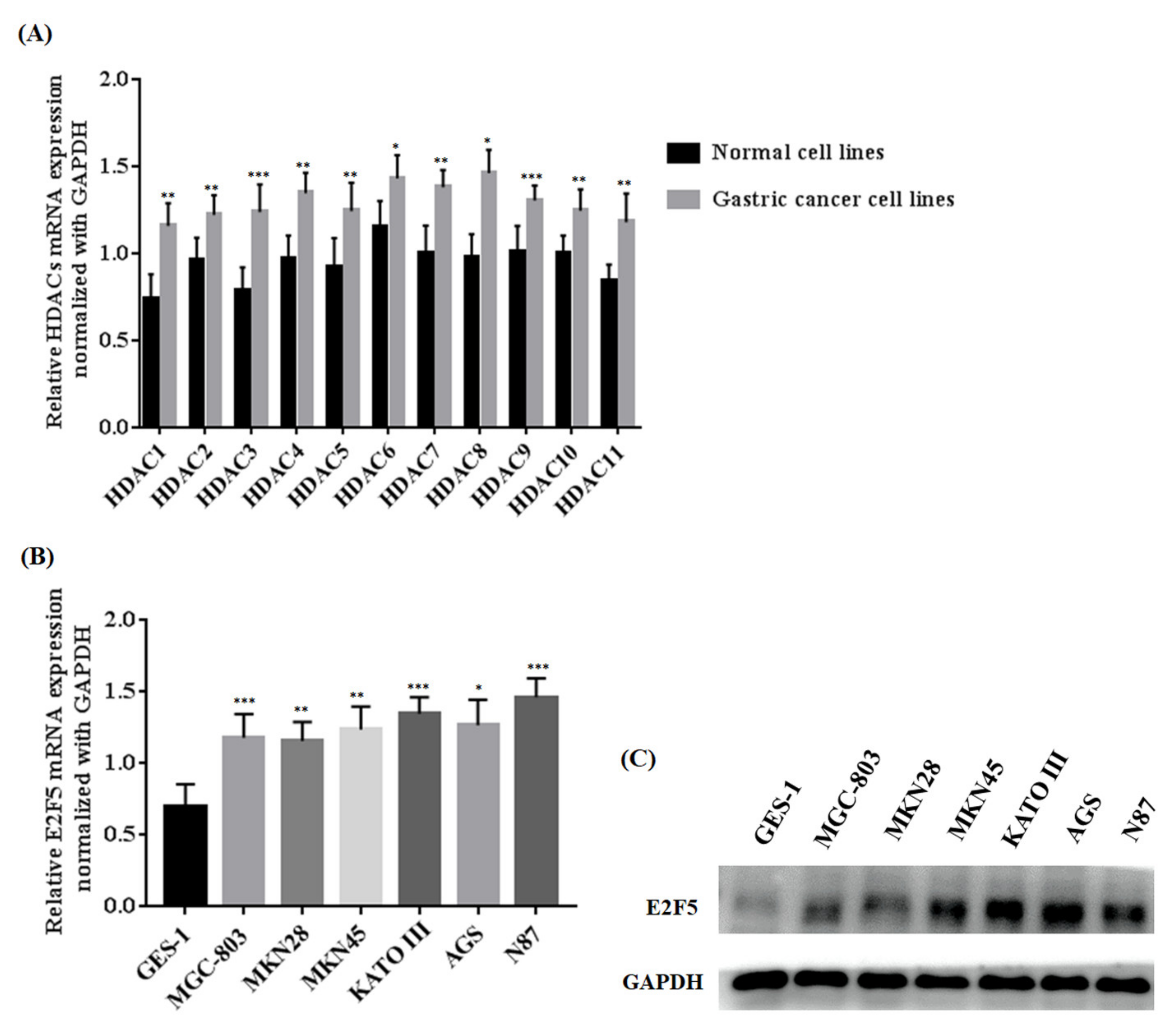
3.1. Investigation into Expression of HDACs in Gastric Cancer Cell Line and Normal Cell Line
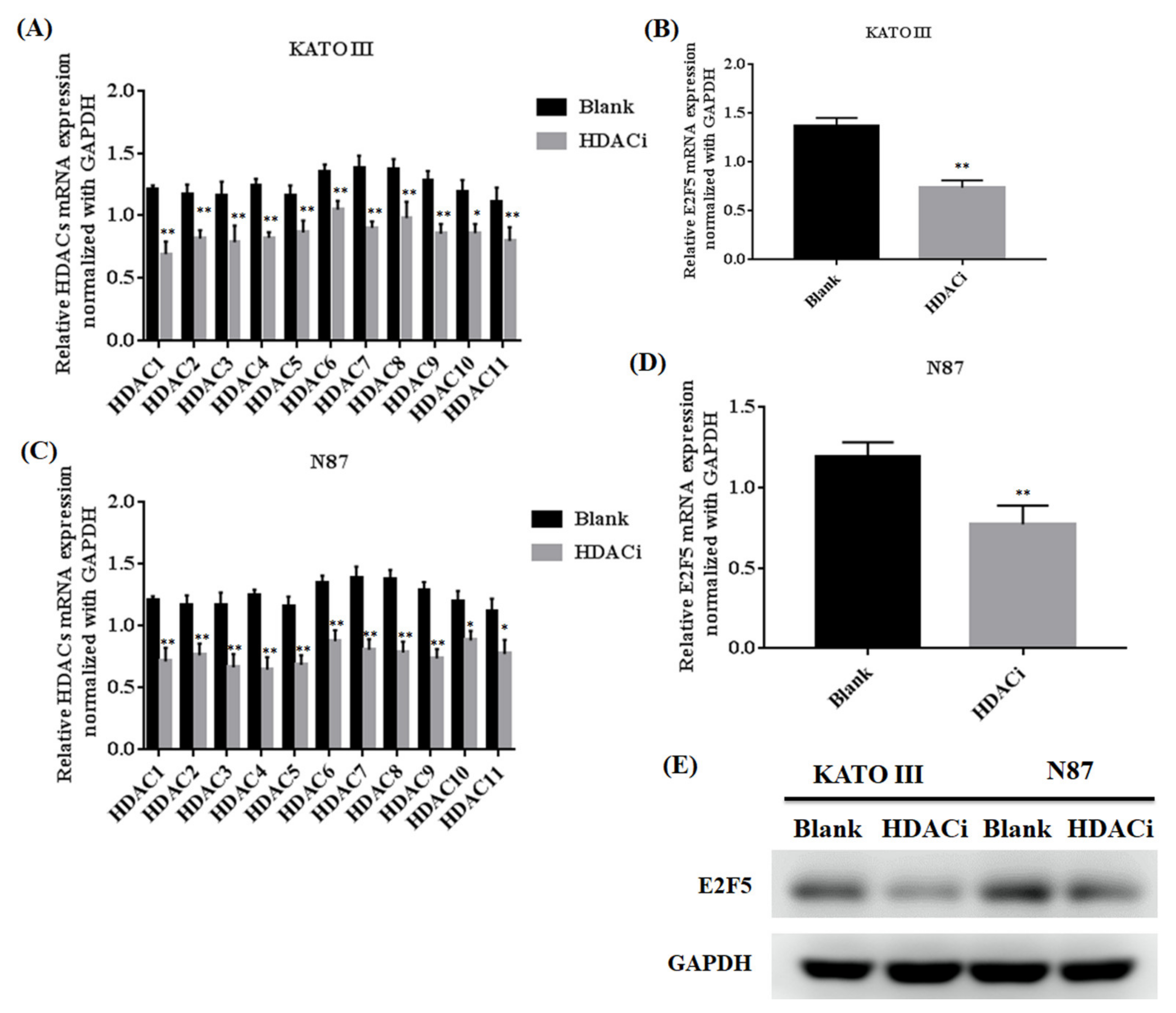
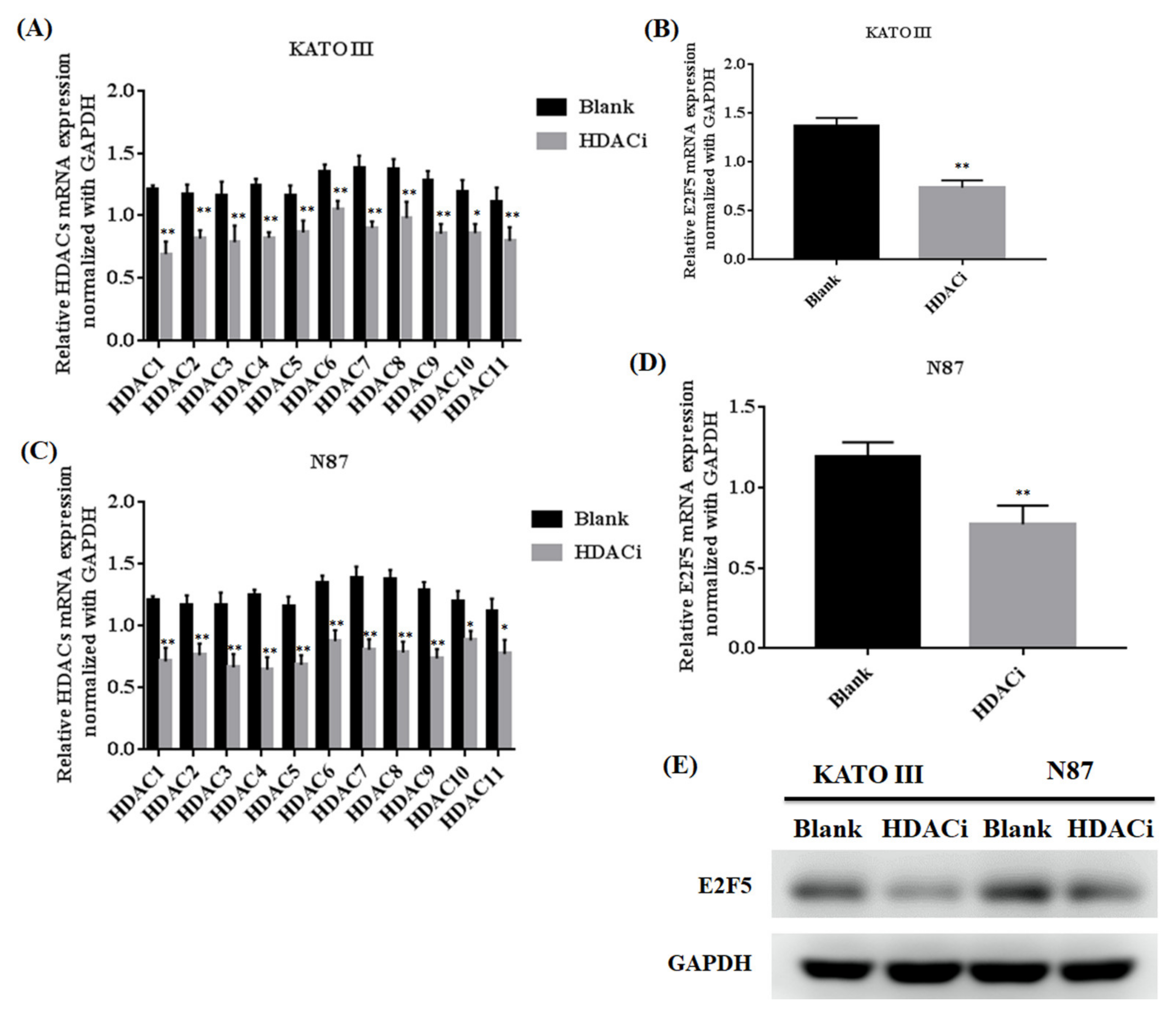
3.2. Histone Deacetylase Inhibitor (HDACi) Downregulated the Expression of HDACs and E2F5 in Gastric Cell Lines
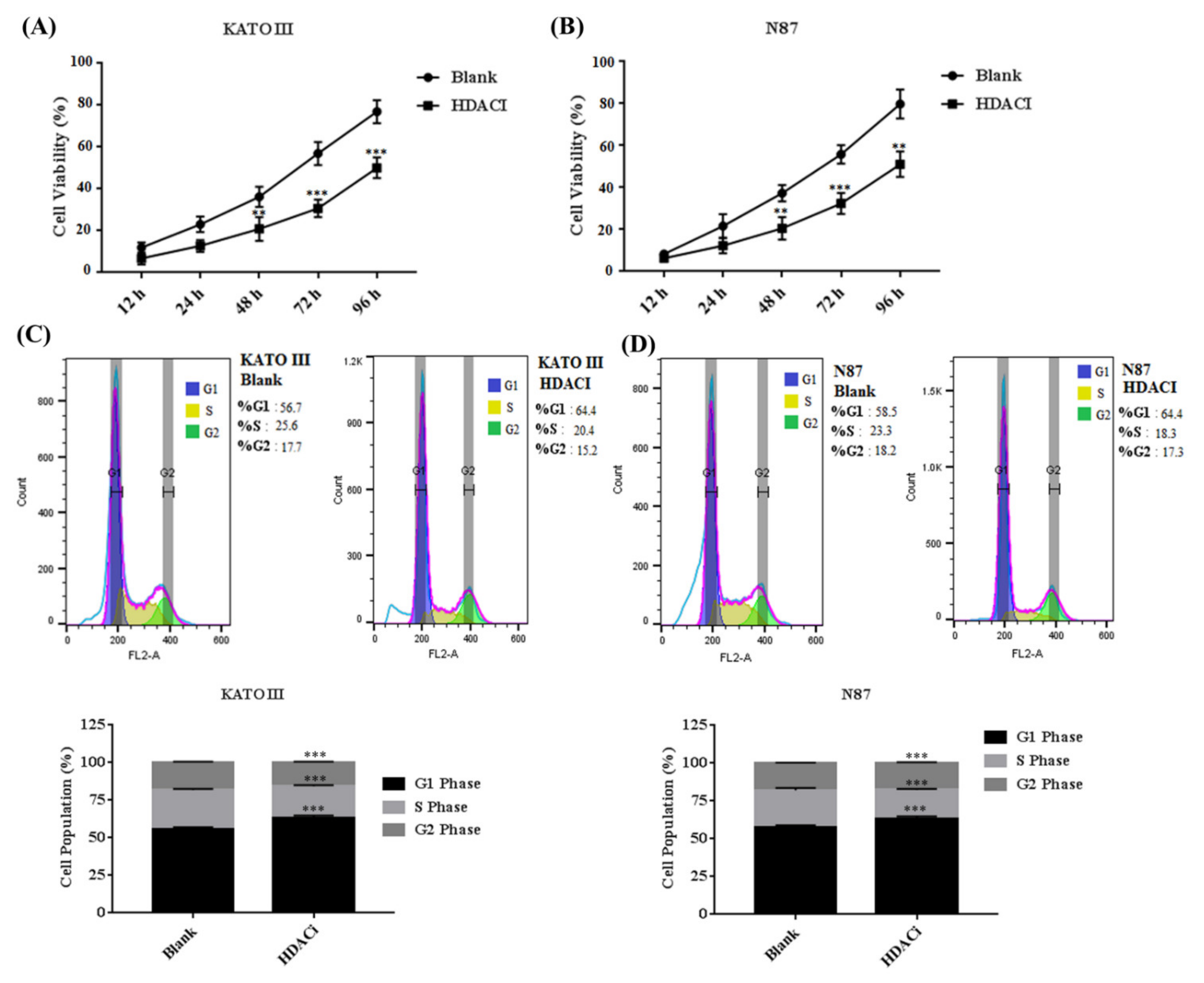
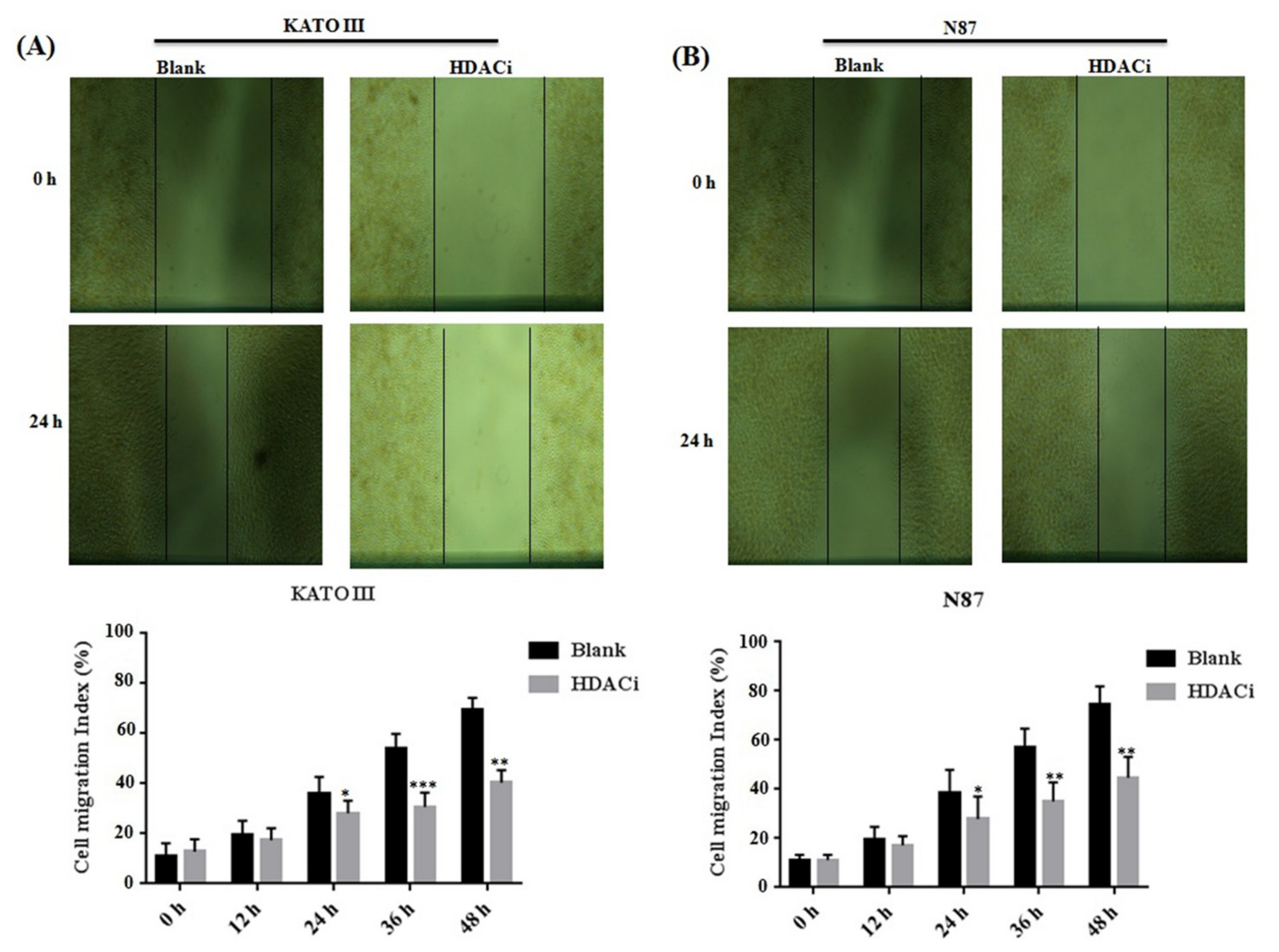
3.3. HDACi Inhibited Cell Proliferation, Cell Migration, and Blocks Cell Cycle at G1 Phase via Downregulating E2F5 in Gastric Cell Lines
3.4. Knocking Down of E2F5 Decreased the Expression of E2F5 and BCL2 in Gastric Cell Lines
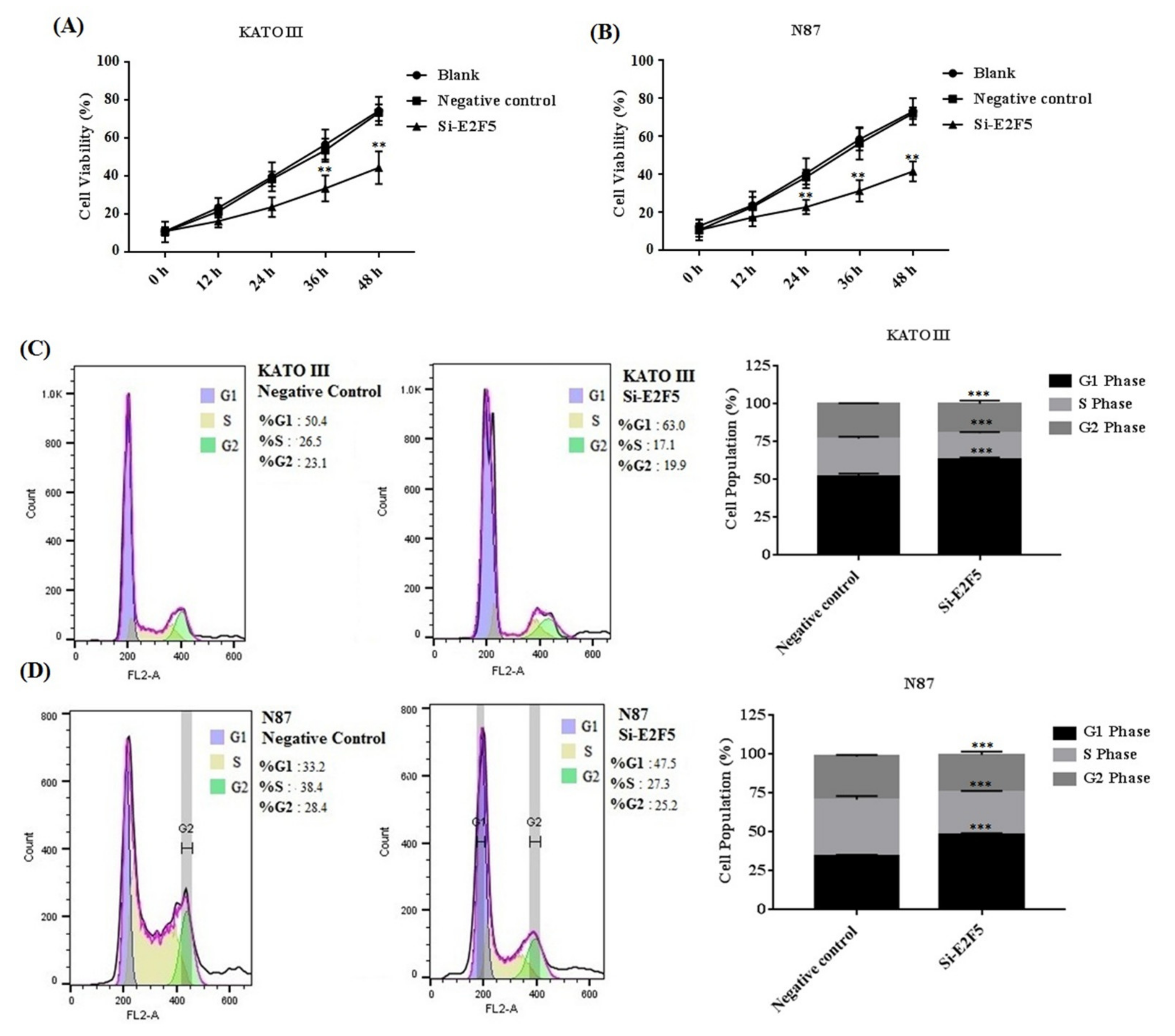
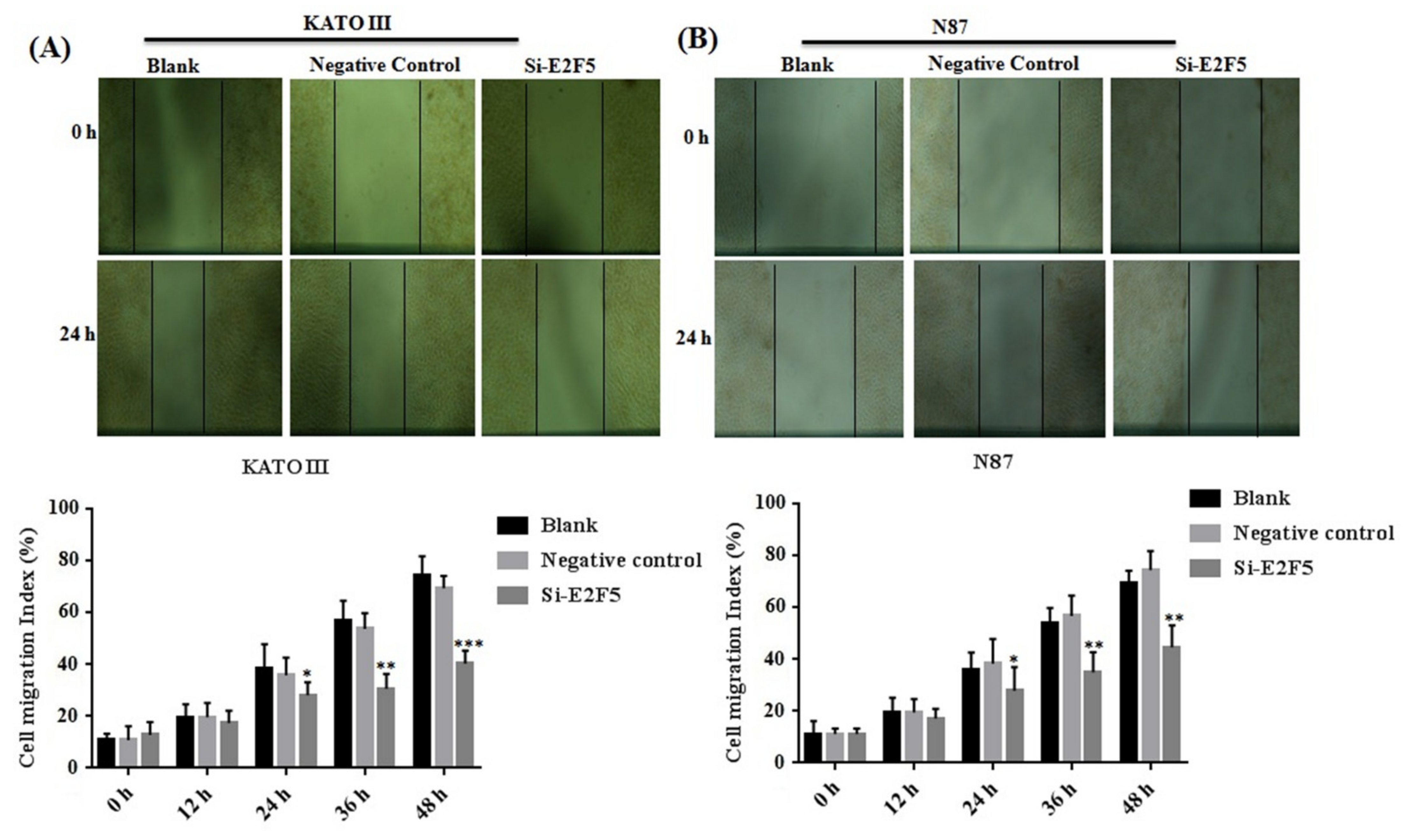
3.5. Knocking Down of E2F5 Decreased Cell Proliferation, Migration, and Block Cell Cycle at G1 Phase in Gastric Cell Lines
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wu, T.; Wu, L. The Role and Clinical Implications of the Retinoblastoma (RB)-E2F Pathway in Gastric Cancer. Front. Oncol. 2021, 11, 655630. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Yang, X.; Li, W.; Chen, Z. Comprehensive Analysis of E2F Family Members in Human Gastric Cancer. Front. Oncol. 2021, 11, 625257. [Google Scholar] [CrossRef] [PubMed]
- Lehrmann, H.; Pritchard, L.L.; Harel-Bellan, A. Histone acetyltransferases and deacetylases in the control of cell proliferation and differentiation. Adv. Cancer Res. 2002, 86, 41–65. [Google Scholar]
- Gallinari, P.; Di Marco, S.; Jones, P.; Pallaoro, M.; Steinkuhler, C. HDACs, histone deacetylation and gene transcription: From molecular biology to cancer therapeutics. Cell Res. 2007, 17, 195–211. [Google Scholar] [CrossRef]
- Li, G.; Tian, Y.; Zhu, W.G. The Roles of Histone Deacetylases and Their Inhibitors in Cancer Therapy. Front. Cell. Dev. Biol. 2020, 8, 576946. [Google Scholar] [CrossRef] [PubMed]
- Remiszewski, S.W.; Sambucetti, L.C.; Bair, K.W.; Bontempo, J.; Cesarz, D.; Chandramouli, N.; Chen, R.; Cheung, M.; Cornell-Kennon, S.; Dean, K.; et al. N-hydroxy-3-phenyl-2-propenamides as novel inhibitors of human histone deacetylase with in vivo antitumor activity: Discovery of (2E)-N-hydroxy-3-[4-[[(2-hydroxyethyl)[2-(1H-indol-3-yl)ethyl]amino]methyl]phenyl ]-2-propenamide (NVP-LAQ824). J. Med. Chem. 2003, 46, 4609–4624. [Google Scholar] [CrossRef] [PubMed]
- McClure, J.J.; Li, X.; Chou, C.J. Advances and Challenges of HDAC Inhibitors in Cancer Therapeutics. Adv. Cancer Res. 2018, 138, 183–211. [Google Scholar]
- Xie, H.; Kang, Y.; Wang, S.; Zheng, P.; Chen, Z.; Roy, S.; Zhao, C. E2f5 is a versatile transcriptional activator required for spermatogenesis and multiciliated cell differentiation in zebrafish. PLoS Genet. 2020, 16, e1008655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishimoto, T.; Shiozaki, A.; Ichikawa, D.; Fujiwara, H.; Konishi, H.; Komatsu, S.; Kubota, T.; Okamoto, K.; Nakashima, S.; Shimizu, H.; et al. E2F5 as an independent prognostic factor in esophageal squamous cell carcinoma. Anticancer Res. 2013, 33, 5415–5420. [Google Scholar] [PubMed]
- Scopa, C.D.; Vagianos, C.; Kardamakis, D.; Kourelis, T.G.; Kalofonos, H.P.; Tsamandas, A.C. bcl-2/bax ratio as a predictive marker for therapeutic response to radiotherapy in patients with rectal cancer. Appl. Immunohistochem. Mol. Morphol. 2001, 9, 329–334. [Google Scholar] [CrossRef] [PubMed]
- Jian, X.; Qu, L.; Wang, Y.; Zou, Q.; Zhao, Q.; Chen, S.; Gao, X.; Chen, H.; He, C. Trichostatin Ainduced miR30a5p regulates apoptosis and proliferation of keloid fibroblasts via targeting BCL2. Mol. Med. Rep. 2019, 19, 5251–5262. [Google Scholar] [PubMed] [Green Version]
- Bian, X.; Liang, Z.; Feng, A.; Salgado, E.; Shim, H. HDAC inhibitor suppresses proliferation and invasion of breast cancer cells through regulation of miR-200c targeting CRKL. Biochem. Pharmacol. 2018, 147, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Schizas, D.; Mastoraki, A.; Naar, L.; Tsilimigras, D.I.; Katsaros, I.; Fragkiadaki, V.; Karachaliou, G.S.; Arkadopoulos, N.; Liakakos, T.; Moris, D. Histone Deacetylases (HDACs) in Gastric Cancer: An Update of their Emerging Prognostic and Therapeutic Role. Curr. Med. Chem. 2020, 27, 6099–6111. [Google Scholar] [CrossRef]
- Amnekar, R.V.; Khan, S.A.; Rashid, M.; Khade, B.; Thorat, R.; Gera, P.; Shrikhande, S.V.; Smoot, D.T.; Ashktorab, H.; Gupta, S. Histone deacetylase inhibitor pre-treatment enhances the efficacy of DNA-interacting chemotherapeutic drugs in gastric cancer. World J. Gastroenterol. 2020, 26, 598–613. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Gong, X. Let-7c Inhibits the Proliferation, Invasion, and Migration of Glioma Cells via Targeting E2F5. Oncol. Res. 2018, 26, 1103–1111. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.S.; Gonugunta, V.K.; Bandyopadhyay, A.; Rao, M.K.; Goodall, G.J.; Sun, L.Z.; Tekmal, R.R.; Vadlamudi, R.K. Significance of PELP1/HDAC2/miR-200 regulatory network in EMT and metastasis of breast cancer. Oncogene 2014, 33, 3707–3716. [Google Scholar] [CrossRef] [Green Version]
- Mekala, J.R.; Naushad, S.M.; Ponnusamy, L.; Arivazhagan, G.; Sakthiprasad, V.; Pal-Bhadra, M. Epigenetic regulation of miR-200 as the potential strategy for the therapy against triple-negative breast cancer. Gene 2018, 641, 248–258. [Google Scholar] [CrossRef] [PubMed]
- Stella di Stadio, C.; Faraonio, R.; Federico, A.; Altieri, F.; Rippa, E.; Arcari, P. GKN1 expression in gastric cancer cells is negatively regulated by miR-544a. Biochimie 2019, 167, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Liang, G.; Yang, S.; Sui, J.; Yao, W.; Shen, X.; Zhang, Y.; Peng, H.; Hong, W.; Xu, S.; et al. Dysregulated lncRNA-UCA1 contributes to the progression of gastric cancer through regulation of the PI3K-Akt-mTOR signaling pathway. Oncotarget 2017, 8, 93476–93491. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Fang, H.; Hou, Z.; Sang, L.; Yang, X. An in-line capillary electrophoresis assay for the high-throughput screening of histone deacetylase inhibitors. J. Chromatogr. A. 2019, 1591, 171–177. [Google Scholar] [CrossRef]
- Yao, S.; Yan, W. Overexpression of Mst1 reduces gastric cancer cell viability by repressing the AMPK-Sirt3 pathway and activating mitochondrial fission. Onco. Targets Ther. 2018, 11, 8465–8479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Wang, X.; Lu, Y.; Han, S.; Zhang, F.; Zhai, H.; Lei, T.; Liang, J.; Wang, J.; Wu, K.; et al. Expression of 15-PGDH is downregulated by COX-2 in gastric cancer. Carcinogenesis 2008, 29, 1219–1227. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ali, A.; Ali, A.; Khan, S.; Ibrahim, M.; Alshehri, M.A.; Thirupathi, A. Inhibition of HDACs Suppresses Cell Proliferation and Cell Migration of Gastric Cancer by Regulating E2F5 Targeting BCL2. Life 2021, 11, 1425. https://doi.org/10.3390/life11121425
Ali A, Ali A, Khan S, Ibrahim M, Alshehri MA, Thirupathi A. Inhibition of HDACs Suppresses Cell Proliferation and Cell Migration of Gastric Cancer by Regulating E2F5 Targeting BCL2. Life. 2021; 11(12):1425. https://doi.org/10.3390/life11121425
Chicago/Turabian StyleAli, Arshad, Ayaz Ali, Shaker Khan, Muhammad Ibrahim, Mohammed Ali Alshehri, and Anand Thirupathi. 2021. "Inhibition of HDACs Suppresses Cell Proliferation and Cell Migration of Gastric Cancer by Regulating E2F5 Targeting BCL2" Life 11, no. 12: 1425. https://doi.org/10.3390/life11121425
APA StyleAli, A., Ali, A., Khan, S., Ibrahim, M., Alshehri, M. A., & Thirupathi, A. (2021). Inhibition of HDACs Suppresses Cell Proliferation and Cell Migration of Gastric Cancer by Regulating E2F5 Targeting BCL2. Life, 11(12), 1425. https://doi.org/10.3390/life11121425