
Dynamics of an Impulsive Stochastic Predator–Prey System with the Beddington–DeAngelis Functional Response


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
- All functions , , , , , , , , , , and are positive, bounded, continuous, and periodic with the same period T.
- The impulsive points satisfy with and there exists a positive integer q such that and for , 2 and .
- By the biological meanings, we assume for , 2 and .
2. Preliminaries
- (i)
- is -adapted and is continuous on and each interval , , where is the set of all -valued measurable -adapted processes satisfying almost surely for all ,, 2;
- (ii)
- For every , , and exist, and with the probability one;
- (iii)
- For all obeys the integral equationand for all obeys the following integral equation:
- (a)
- If there exist two positive constants and such that, for all ,then
- (b)
- If there exist some constants , , and λ such that, for all ,then
3. Existence of Periodic Markovian Processes
4. Extinction and Permanence in Mean
- (i)
- If and then all species of (5) are extinct, i.e.,
- (ii)
- If and then is permanent in mean and is extinct, i.e., for some positive numbers and
- (iii)
- If and then is extinct and is permanent in mean, i.e., and for some positive numbers and
- (iv)
- If and , then is permanent in mean, i.e., for some positive numbers and .
- (i)
- Integrating both sides from 0 to t yieldsWith a similar argument as above, we can obtain . Therefore, all species are extinct.
- (ii)
- If then Lemma 2 and (11) implyBy monotonicity, we can derive from (10) thatThusSince it follows from Lemma 2 that . Using (9) again, we haveLetting and using Lemma 2, we arrive atIn summary,which means is permanent in mean.
- (iii)
- Since using Lemma 2 giveswhich means is permanent in mean.
- (iv)
- Obviously, and imply that and , respectively. Therefore, species can not be extinct. It follows from (9) thatApply Lemma 2 to getThis, combined with (ii), produces , i.e., is permanent in mean.Since applying Lemma 2 to (12) yieldsMoreover, we get from (10) thatIn view of and applying Lemma 2 again yields
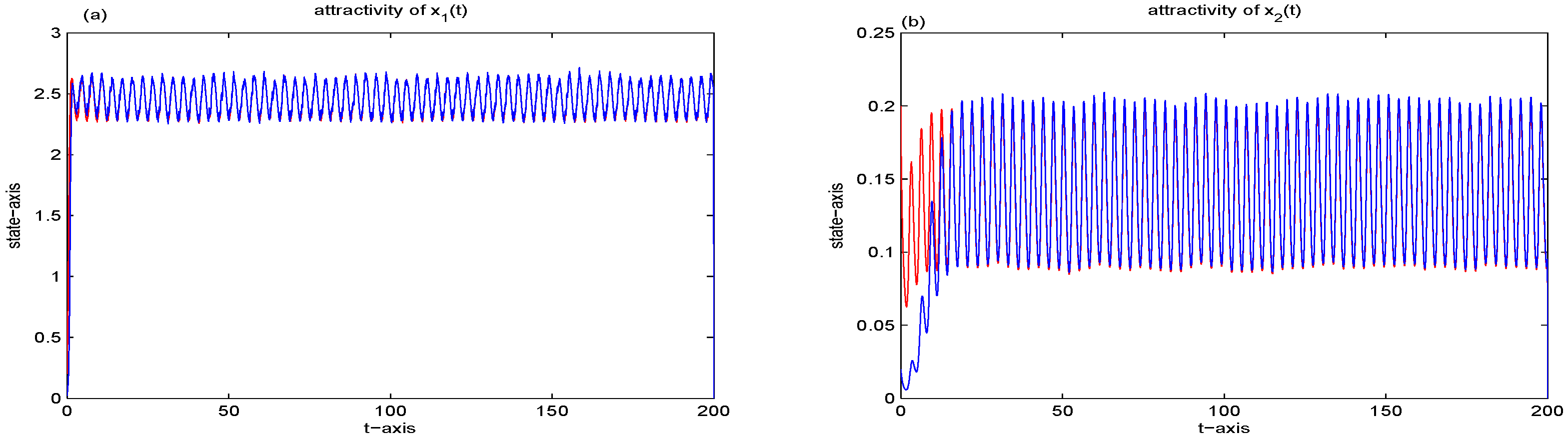
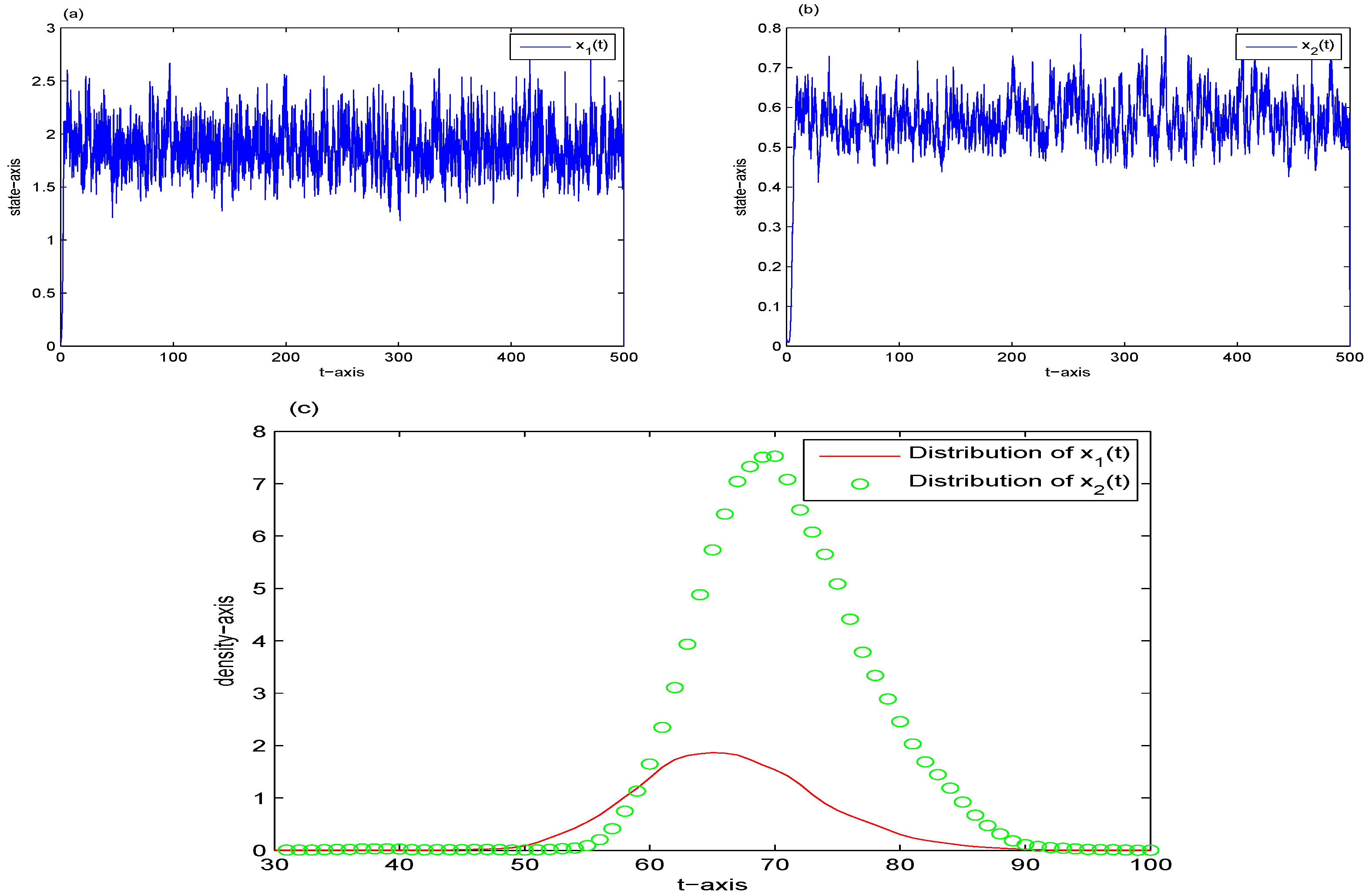
5. Stationary Distribution
- (i)
- F is uniformly elliptical in the domain U and some neighborhood thereof, where .
- (ii)
- There is a non-negative -function and a positive constant C such that for any
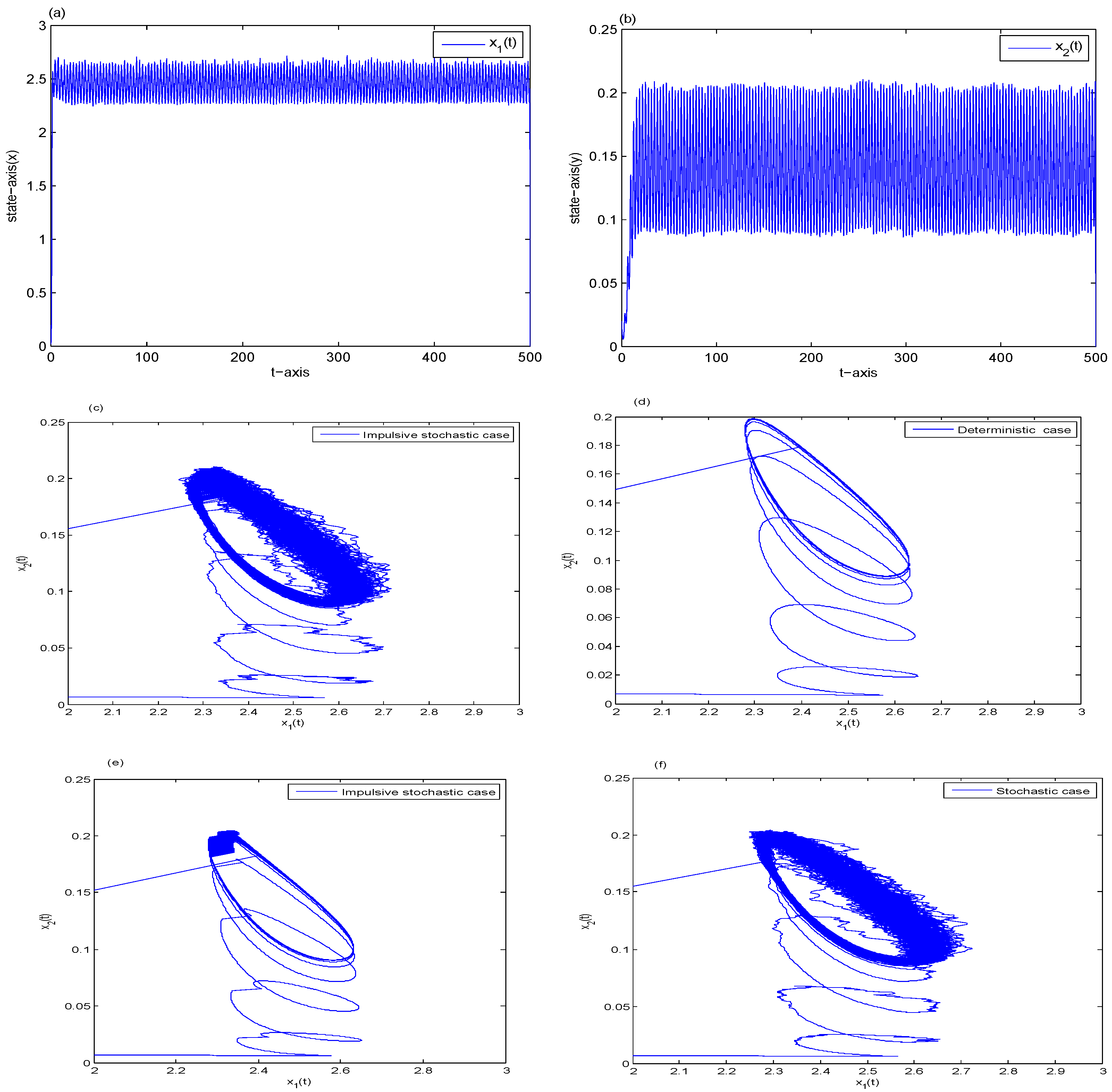
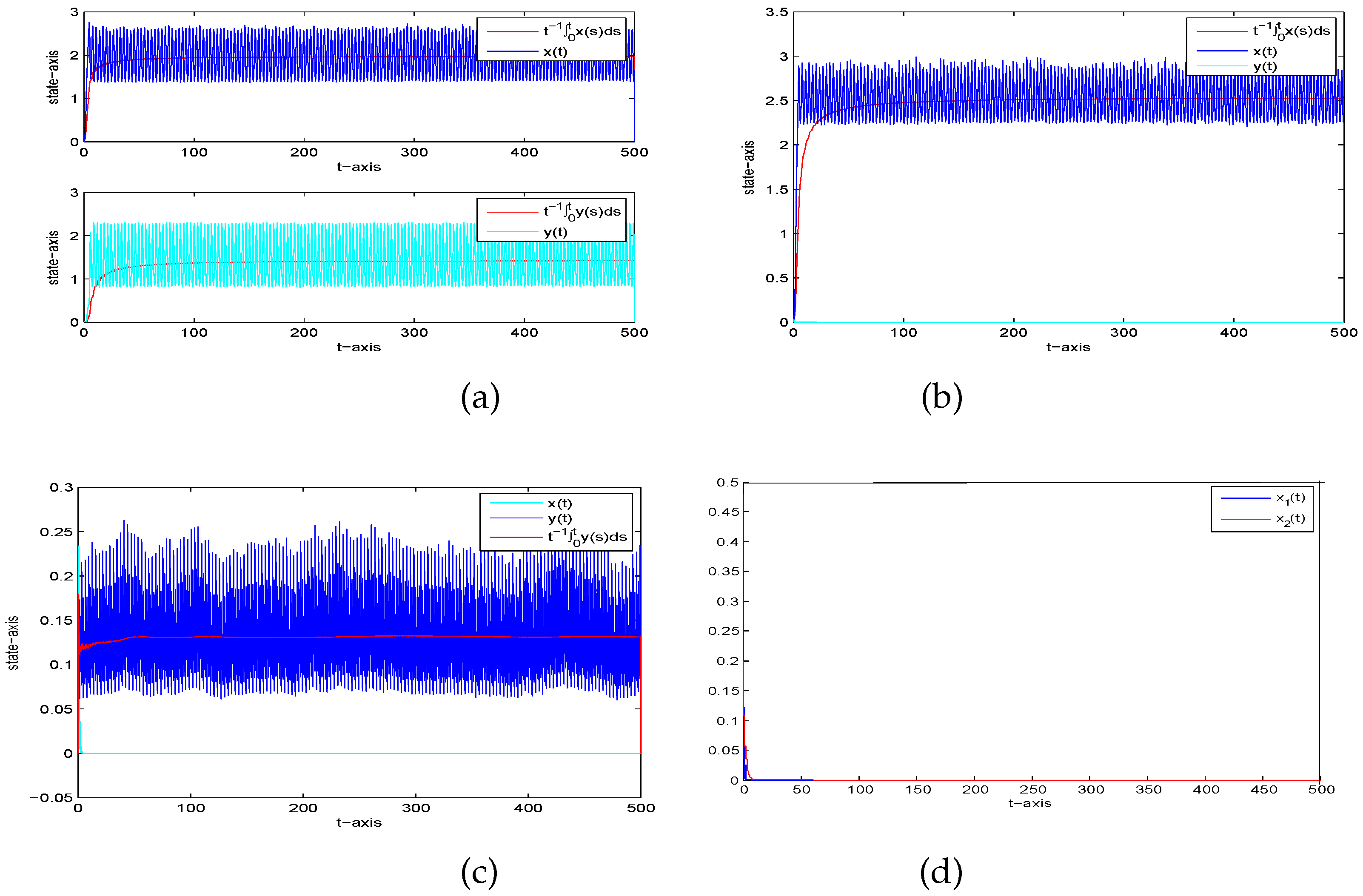
6. Examples and Simulations
- (i)
- Let , then and . Theorem 4 implies is permanent in the mean and is extinct, see Figure 3b. It shows that too much white noise results in the extinction of the predator.
- (ii)
- If then and . Theorem 4 implies is extinct and is permanent in the mean, as illustrated in Figure 3c, which shows that too large a pulse leads to the extinction of the prey.
- (iii)
- If , then and . Theorem 4 shows that both prey and predator are extinct (Figure 3d). This indicates that the white noise has a huge influence on the system permanence, and too much noise will make all species extinct.
7. Discussion and Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cantrell, R.S.; Cosner, C. On the dynamics of predator-prey models with the Bedding-DeAngelis functional response. J. Math. Anal. Appl. 2001, 257, 206–222. [Google Scholar] [CrossRef]
- Fan, M.; Kuang, Y. Dynamics of a non-autonomous predator-prey system with the Beddington-DeAngelis functional response. J. Math. Anal. Appl. 2004, 295, 15–39. [Google Scholar] [CrossRef] [Green Version]
- Wang, H. Global analysis of the predator-prey system with Beddington-DeAngelis functional response. J. Math. Anal. Appl. 2003, 281, 395–401. [Google Scholar] [CrossRef] [Green Version]
- Friedman, A. Stochastic Differential Equations and Their Applications; Academic Press: New York, NY, USA, 1976. [Google Scholar]
- Du, N.H.; Sam, V.H. Dynamics of a stochastic Lotka-Volterra model perturbed by white noise. J. Math. Anal. Appl. 2006, 324, 82–97. [Google Scholar] [CrossRef] [Green Version]
- Mao, X.; Marion, G.; Renshaw, E. Environmental Brownian noise suppresses explosions in populations dynamics. Stoch. Process. Appl. 2002, 97, 95–110. [Google Scholar] [CrossRef]
- Li, X.; Mao, X. Population dynamical behavior of non-autonomous Lotka-Volterra competitive system with random perturbation. Discret. Contin. Dyn. Syst. 2009, 24, 523–545. [Google Scholar] [CrossRef] [Green Version]
- Zuo, W.; Jiang, D.; Sun, X.; Hayat, T.; Alsaedib, A. Long-time behaviors of a stochastic cooperative Lotka-Volterra system with distributed delay. Physica A 2018, 506, 542–559. [Google Scholar] [CrossRef]
- Yagi, A.; Ton, T.V. Dynamic of a stochastic predator-prey population. Appl. Math. Comput. 2011, 218, 3100–3109. [Google Scholar] [CrossRef]
- Shao, Y.; Li, Y. Dynamical analysis of a stage structured predator-prey system with impulsive diffusion and generic functional response. Appl. Math. Comput. 2013, 220, 472–481. [Google Scholar] [CrossRef]
- Wu, R.H.; Zou, X.L.; Wang, K. Asymptotic behavior of a stochastic non-autonomous predator-prey model with impulsive perturbations. Commun. Nonlinear Sci. Numer. Simul. 2015, 20, 965–974. [Google Scholar] [CrossRef]
- Liu, M.; Wang, K. On a stochastic logistic equation with impulsive perturbations. Comput. Math. Appl. 2012, 63, 871–886. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.W.; Tan, D.J. Dynamics of a stochastic predator-prey system in a polluted environment with pulse toxicant input and impulsive perturbations. Appl. Math. Model. 2015, 39, 6319–6331. [Google Scholar] [CrossRef]
- Chen, L.J.; Chen, F.D. Dynamic behaviors of the periodic predator-prey system with distributed time delays and impulsive effect. Nonlinear Anal. Real World Appl. 2011, 12, 2467–2473. [Google Scholar] [CrossRef]
- Zhang, S.Q.; Meng, X.Z.; Feng, T.; Zhang, T.H. Dynamics analysis and numerical simulations of a stochastic non-autonomous predator-prey system with impulsive effects. Nonlinear Anal. Hybrid Syst. 2017, 26, 19–37. [Google Scholar] [CrossRef]
- Jiang, D.; Zuo, W.J.; Hayat, T.; Alsaedi, A. Stationary distribution and periodic solutions for stochastic Holling-Leslie predator-prey systems. Physica A 2016, 460, 16–28. [Google Scholar] [CrossRef]
- Zhao, Y.; Yuan, S.; Zhang, T.H. Stochastic periodic solution of a non-autonomous toxic-producing phytoplankton allelopathy model with environmental fluctuation. Commun. Nonlinear Sci. Numer. Simul. 2017, 44, 266–276. [Google Scholar] [CrossRef]
- Jiang, D.; Zhang, Q.M.; Hayat, T.; Alsaedi, A. Periodic solution for a stochastic non-autonomous competitive Lotka-Volterra model in a polluted environment. Physica A 2017, 471, 276–287. [Google Scholar] [CrossRef]
- Shao, Y. Globally asymptotical stability and periodicity for a nonautonomous two-species system with diffusion and impulses. Appl. Math. Model. 2012, 36, 288–300. [Google Scholar] [CrossRef]
- Li, D.; Xu, D. Periodic solutions of stochastic delay differential equations and applications to Logistic equation and neural networks. J. Korean Math. Soc. 2013, 50, 1165–1181. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Gopalsamy, K. On the periodic solution of N-dimensional stochastic population models. Stoch. Anal. Appl. 2000, 18, 323–331. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, S.H.; Gao, S.J.; Wei, X. Stochastic periodic solution for a perturbed non-autonomous predator-prey model with generalized nonlinear harvesting and impulses. Physica A 2017, 486, 347–366. [Google Scholar] [CrossRef]
- Zuo, W.J.; Jiang, D. Periodic solutions for a stochastic non-autonomous Holling-Tanner predator-prey system with impulses. Nonlin. Anal. Hybrid Syst. 2016, 22, 191–201. [Google Scholar] [CrossRef]
- Liu, M.; Wang, K. Asymptotic behavior of a stochastic nonautonomous Lotka-Volterra competitive system with impulsive perturbations. Math. Comput. Model. 2013, 57, 909–925. [Google Scholar] [CrossRef]
- Khas’minskii, R.Z. Stochastic Stability of Differential Equations; Sijthoff Noordhoff: Alphen aan den Rijn, The Netherlands; Springer Science & Business Media: Dordrecht, The Netherlands, 1980. [Google Scholar]
- Ji, C.Y.; Jiang, D. Dynamics of a stochastic density dependent predator-prey system with Beddington-DeAngelis functional response. J. Math. Anal. Appl. 2011, 381, 441–453. [Google Scholar] [CrossRef] [Green Version]
- Qiu, H.; Liu, M.; Wang, K.; Wang, Y. Dynamics of a stochastic predator-prey system with Beddington-DeAngelis functional response. Appl. Math. Comput. 2012, 219, 2303–2312. [Google Scholar] [CrossRef]
- Karatzas, I.; Shreve, S.E. Brownian Motion and Stochastic Calculus; Springer: Berlin/Heidelberg, Germany, 1991. [Google Scholar]
- Barbalat, I. Systems dequations differentielles d’osci d’oscillations nonlineaires. Rev. Roum. Math. Pures Appl. 1959, 4, 267–270. [Google Scholar]
- Higham, D.J. An algorithmic introduction to numerical simulation of stochastic differential equations. SIAM Rev. 2001, 43, 525–546. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shao, Y. Dynamics of an Impulsive Stochastic Predator–Prey System with the Beddington–DeAngelis Functional Response. Axioms 2021, 10, 323. https://doi.org/10.3390/axioms10040323
Shao Y. Dynamics of an Impulsive Stochastic Predator–Prey System with the Beddington–DeAngelis Functional Response. Axioms. 2021; 10(4):323. https://doi.org/10.3390/axioms10040323
Chicago/Turabian StyleShao, Yuanfu. 2021. "Dynamics of an Impulsive Stochastic Predator–Prey System with the Beddington–DeAngelis Functional Response" Axioms 10, no. 4: 323. https://doi.org/10.3390/axioms10040323
APA StyleShao, Y. (2021). Dynamics of an Impulsive Stochastic Predator–Prey System with the Beddington–DeAngelis Functional Response. Axioms, 10(4), 323. https://doi.org/10.3390/axioms10040323