Abstract
The post-impact fauna of the Dalby Limestone of Tvären Bay has been extensively examined, with the exception of the Palaeostomate bryozoan taxa present. Here, we report three palaeostomate bryozoans found in limestone boulders recovered from glacial deposits on Ringsö Island derived from Tvären Bay, Sweden. The bryozoan fauna includes Pachydictya bifurcata, Hallopora sp., and Trematoporid sp. indet. Pachydictya bifurcata has bifoliate zoaria with two layers of zooecia oriented in opposite directions along the mesotheca. Hallopora sp. is characterized by the presence of mesozooecia, tubular autozooecia with rounded apertures, thin walls, and possessing diaphragms. The zoaria of Trematoporid sp. indet are loosely arranged and slightly ambiguous, but lack further identifying features. The report of Pachydictya bifurcata herein represents an extension of its stratigraphic range within the late Ordovician. These findings enhance our understanding of the bryozoan diversity in the region, providing the first detailed report of the presence of these taxa in this geological context.
1. Introduction
Bryozoa are a colonial, aquatic, predominantly sessile, suspension-feeding phylum within the Lophotrochozoa [1,2], and have a long and extensive fossil record that stretches through much of the Phanerozoic. The first appearance of fossils interpreted as bryozoans is in the early Cambrian, with fossils reported from Australia, south China, and the western United States [3,4]. Currently, nearly 6000 extant and over 15,000 fossil bryozoan species have been recognized [5,6,7,8], with a significant increase in diversity during the Ordovician [2,9,10,11,12,13]. Given the substantial time gap between the initial appearance of bryozoans and their major radiation [3], bryozoan studies are crucial for understanding the Cambrian explosion and the Ordovician radiation. Bryozoan-rich sediments have been extensively studied in Baltoscandia [14,15] and significant advancements have been made in the understanding of Ordovician bryozoans based on material from Kinnekule, Ösergötlan, Dalarna, and Gotland, Sweden [16,17,18,19,20,21,22,23].
The Tvären region offers yet another excellent window to study the bryozoan fauna of Sweden. The Tvären region preserves a complete post-meteorite impact sedimentary succession, characterised by the fossiliferous Dalby Limestone from the Upper Ordovician. Extensive research has been conducted on the sedimentology, palaeontology and biostratigraphy within and around the Tvären crater [24,25,26,27,28,29,30,31,32,33,34,35,36]. Fossils such as chitinozoans and conodonts [27,28,29], trilobites [24,37], ostracods [24,25], graptolites [26], gastropods [38], scolecodonts [35], and brachiopods [31,36,39] from the post-impact Dalby Limestone of Tvären Bay have been examined. Chitinozoan and conodont fossil zones have been established and can be correlated globally. The distribution and ecological spatial utilisation patterns of highly diversified brachiopods have also been reported and analysed [36]. However, bryozoans from this area have not been formally reported. They were only briefly mentioned in a previous work by Thorslund [24], who stated that “several species of bryozoans have been collected; a few of them are identical with species described by Bassler [15] from the Kuckers Formation”. However, upon reviewing the species and genus list in Bassler’s work, we found that the taxa identified in this study are not included. In this study, we examined glacial boulders from the Tvären area in Sweden. The primary aim is to investigate bryozoan fossils associated with the Dalby Limestone, provide detailed descriptions of the bryozoan genera and species identified in this region, and discuss the stratigraphic distribution of these taxa.
2. Geological Setting and Stratigraphy
The boulders consist of sandstone, breccia, and fossiliferous limestone [24]. Fossiliferous limestone boulders are relatively abundant in a limited area on Ringsö Island, near the vicinity of the Tvären impact (Figure 1). These boulders are dispersed in a region that narrows significantly towards the NNW. The boundaries of the area occupied by the boulders stretch east and west, forming a sector whose apex is located Tvären (Figure 1). The direction of the boulder drift is indicated by glacial striae [24]. This evidence suggests that the boulders likely originated from Tvären itself. Calcarenitic turbidites from the Tvären-2 core are similar, in both lithology and fossil content (including rhynchonelliformean brachiopods, bryozoans, echinoderms, and ostracodes), with the studied boulders [24,32], further suggesting the origin of the boulders is Tvären. Investigations of the fossiliferous limestone boulders from Ringsö Island suggest that they originated from the Upper Ordovician Dalby Formation of the Tvären region (Figure 2) [24,26,27].
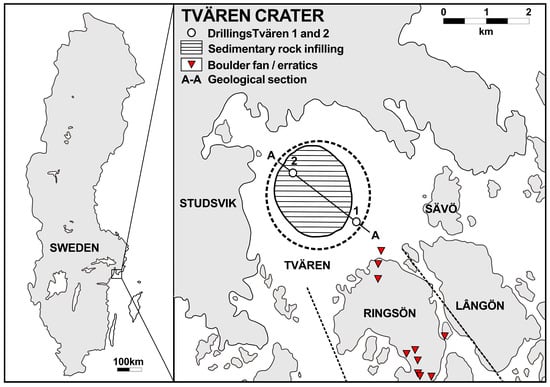
Figure 1.
Map of Sweden and the Tvären crater, showing the location of the crater in southeastern Sweden and the positions of the two drill cores. The glacial erratics from the Dalby Limestone on Ringsö Island is marked by red triangles. The two dotted lines indicate the boundaries where boulders occur.
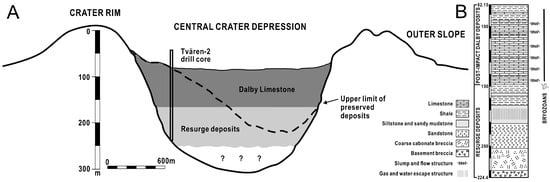
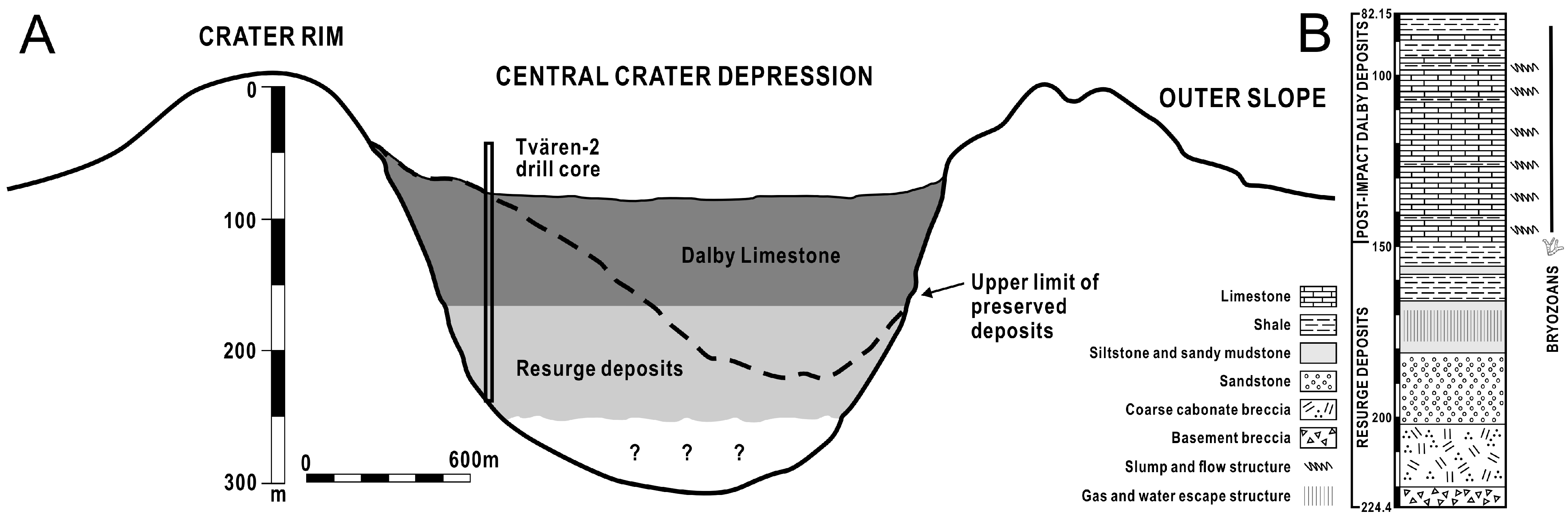
Figure 2.
(A) Cross-section of the Tvären crater based on seismic profiling and drillings as well as proposed rim heights. (B) Lithological succession of the Tvären-2 drill core with distribution of structures and bryozoans (ranges based on first through last occurrence) in the post-impact sediments.
Numerous meteorite impacts occurred in the Baltoscandian Sea during the Ordovician [30,40,41,42,43,44]. The characteristic shape of Tvären Bay on the Swedish east coast, a shallow, subcircular bowl, is presumed to be the result of one such impact [28,29,33] (Figure 1). Situated in the Stockholm Archipelago, the region is predominantly composed of Precambrian crystalline rocks and is approximately 2 km wide with water depths of 100–150 m (Figure 2). The impact formed a crater, preserving a sequence of post-impact deposits that would have otherwise eroded away. According to the Tvären-2 drill core (224.4 m), the stratigraphy at Tvären, deposited on the Proterozoic crystalline basement (224.4–219.6 m), consists of fining-upwards, resurge deposits of limestone breccia (219.6–199.05 m), sand and siltstones (199.05–161.4 m), and mixed mud and carbonate sediments (161.4–82.15 m) [28,33,35]. The carbonate sediments mainly represent the Ordovician Dalby Limestone (Figure 2), which is found across Sweden in areas such as Västergötland, Östergötland, Dalarna, Jämtland, Bothnian Bay, and the subsurface on Gotland, Gotska Sandön, Tvären Bay, and northern Öland [45]. The Dalby Limestone deposited in the crater is several times thicker than in other parts of Sweden, where its thickness is usually 15–20 m. Chitinozoans and conodonts provide excellent stratigraphic control for the Dalby Limestone [28]. Chitinozoans indicate that the post-impact sequence of the Dalby Limestone belongs to the Laufeldochitina (Cyathochitina) stentor chitinozoan Zone [28,30,35,41,44,46,47,48]. The presence of Conochitina tigrina in the uppermost Dalby Limestone limits the age of the post-impact deposits to the topmost Kukruse Stage (lower Sandbian Stage) [30,35]. The Tvären area is assigned to the Amorphognathus tvaerensis conodont Zone, and the presence of this key species, along with Baltoniodus (Prioniodus) variabilis, suggests that the post-impact sediments belong to the Baltoniodus (Prioniodus) variabilis and Baltoniodus (Prioniodus) gerda subzones of the lower Sandbian Stage [28,29,35,49].
3. Materials and Methods
The Tvären crater has been subjected to extensive subaerial and glacial erosion over long periods. All the specimens described herein were derived from limestone boulders originating from the Upper Ordovician Dalby Limestone in Tvären Bay (Figure 1). Rock samples were mechanically crushed into 3 × 3 × 3 cm blocks, then immersed in plastic buckets and dissolved using 10% industrial acetic acid. The process of collecting and drying the samples was repeated every two to three days until the limestone was completely dissolved. The residues were examined under a Zeiss binocular microscope (Stemi 305; Carl Zeiss Microscopy GmbH, Jena, Germany) to collect shelly fossils. Well-preserved fossils were selected for gold coating and were examined using scanning electron microscopy (SEM) at the State Key Laboratory of Continental Dynamics, Northwest University. The SEMs used were a Fei Quanta 450-FEG (20.0 kV, 60 Pa, Working Distance (WD) 8–10 mm; Thermo Fisher Scientific, Hillsboro, OR, USA) and a Phenom XL G2 (5–15 kV, high vacuum mode, WD 6–8 mm; Thermo Fisher Scientific, Eindhoven, The Netherlands). Semi-quantitative analysis was performed by Energy-dispersive X-ray spectroscopy (EDS; Thermo Fisher Scientific, Eindhoven, The Netherlands). In this study, SEM was chosen over traditional thin-section analysis due to the small size of the fossil material, which made thin-section preparation impractical. SEM offers several advantages, including high-resolution imaging that reveals fine surface details, the ability to observe specimens from multiple angles, and the possibility of examining fractured surfaces, providing natural cross-sections that reveal internal structures. These features enhance the accuracy of taxonomic identification. Additionally, SEM avoids the potential damage associated with thin-section preparation and allows for a more efficient analysis of multiple samples. While thin-section analysis remains the standard for continuous internal structure examination, the unique strengths of SEM make it particularly suitable for this study. Four bryozoan specimens and numerous other fossils of shallow-water benthic species were obtained from the boulders, and all specimens are deposited at the Shaanxi Key Laboratory of Early Life and Environment, Northwest University, Xi’an, under the accession numbers ELI-Tvären-Ord-01–23.
4. Systematic Palaeontology
Phylum Bryozoa Ehrenberg, 1831
Class Stenolaemata Borg, 1926
Order Cryptostomata Vine, 1884
Suborder Ptilodictyina Astrova and Morozova, 1956
Family Rhinidictyidae Ulrich, 1893
Genus Pachydictya Ulrich, 1882
Type species. Pachydictya robusta Ulrich, 1882, from Ordovician Caradoc of Knoxville, Tennessee, USA.
Diagnosis. Broad bifoliate fronds with maculae (or monticules); less commonly narrow bifurcating stipes. Zooecia erect, with well-spaced diaphragms but no hemisepta; apertures oval, crowded. Vesicles between zooecia, filled by calcareous tissue near surface and traversed by tubuli.
Pachydictya bifurcata (Hall, 1883)

Figure 3.
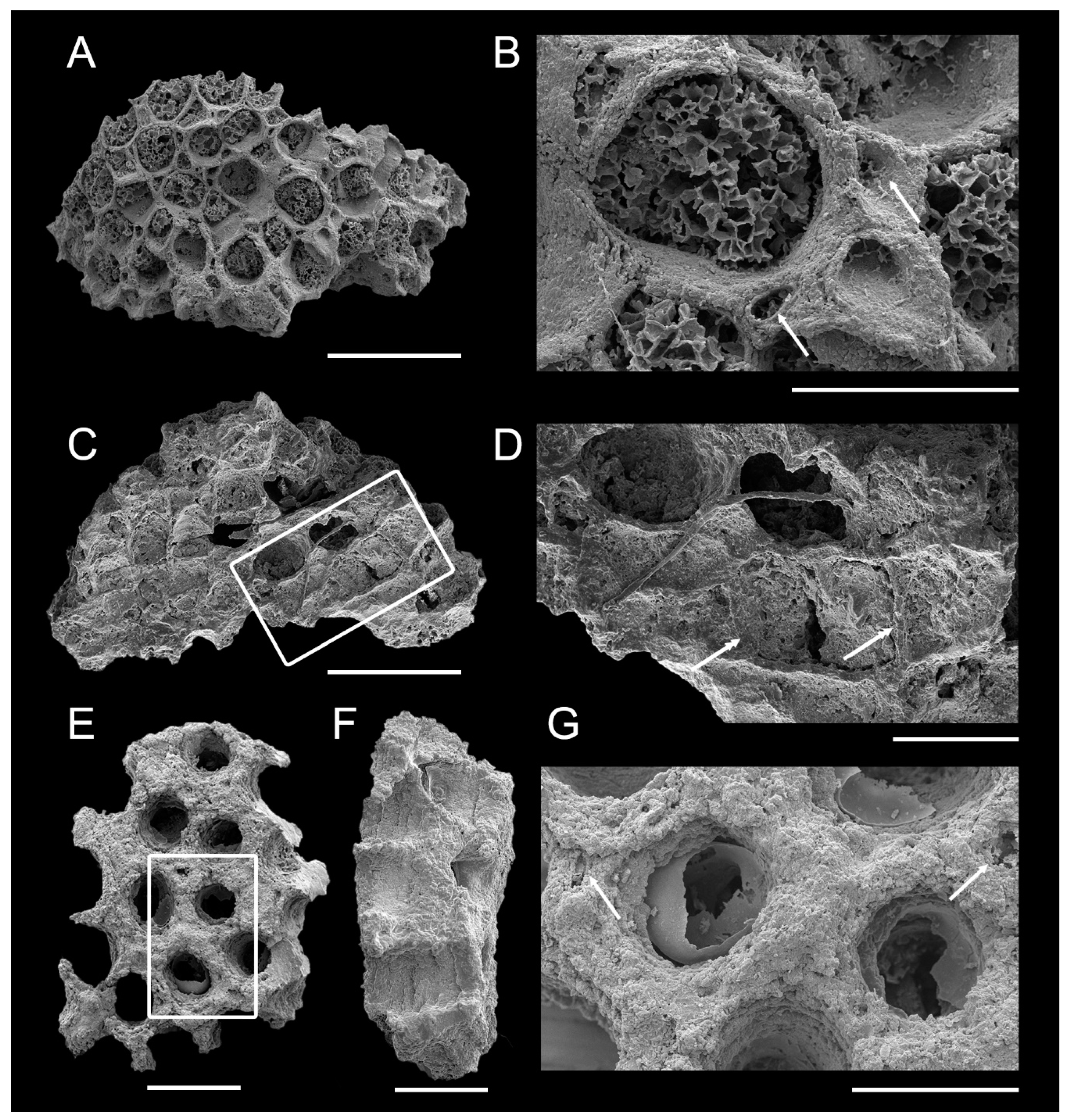
Pachydictya bifurcata from the post-impact Dalby Limestone of erratic boulders on Ringsö Island, Sweden. (A–F) ELI-Tvären-Ord-16, (G–I) ELI-Tvären-Ord-17. (A) Fragment of branch, showing many rows of zooecia; box indicates the position of (D). (B) The lateral view, showing the straight mesotheca and the angle between the zooecia and mesotheca. (C) Oblique lateral view, showing the position of (E,F). (D) Partial enlargement of the zooecia. (E,F) Partial enlargement of mesotheca and vertical wall, showing the vesicles in the zooecial interspace (single arrow). (G) Fragment of branch, showing many rows of zooecia, and the close-up views in (H,I). (H,I) Partial enlargement of the zooecia, showing the tubuli (double arrow). Scale bars 1 mm (A,C,G), 800 μm (B), 300 μm (D–F), 200 μm (H,I).
1860 Stictopora scalpellum Londsdale, Eichwald, p. 390, pl. 24, figs. 15a–c.
1883 Stictopora bifurcata sp. n., Hall, p. 267, pl. 13, figs. 3–4.
1911 Pachydictya bifurcata (Hall), Bassler, p. 143-5, textfig. 67.
1962 Pachydictya bifurcata (Hall), Kiepura, p. 407. 8, pl. IX, figs. 2–3.
1975 Metadichtya bifurcata (Hall), Kopajevich, p. 62.
1984 Pachydichtya bifurcata (Hall 1883), Spjeldnaes, N., pl. 7, figs. 1, 6, textfig. 5A–B.
Material. Two specimens (ELI-Tvären-Ord-16 and ELI-Tvären-Ord-17) recovered from the Dalby limestone in Tvären Bay, Sweden.
Description. Specimens white, translucent, composed of calcareous or siliceous material. Zoaria erect, planate. Branching not observed due to incomplete preservation. Fragments 2.017 mm wide, semi-elliptical in cross-section, with thin, sharp, non-poriferous margins (Figure 3A–C,G). Mesotheca generally straight (Figure 3B). Autozooecial ranges aligned or alternating across mesotheca. Zooecia arranged in seven longitudinal rows (Figure 3A,G). Autozooecia oval to sub-hexagonal in aperture, arranged diagonally in intersecting series, broadest in middle, narrowing proximally, distal edges rounded (Figure 3). Autozooecia partly separated by zooecial interspace, laterally without continuous longitudinal walls (Figure 3E–F). Zooecial interspace filled with vesicles (Figure 3E–F). Tubuli present (Figure 3H–I). Zooecia uniform in size, average width 0.253 mm, length 0.418 mm. Endozones and exozones of colony not clearly defined. In exozones, zooecia form angles 54°–66° with mesotheca (Figure 3B). Living chambers in material 2 filled with aluminosilicate minerals (Figure 3G–I). Exilazooecia absent. Superior hemisepta, basal diaphragms not visible. Measurements and counts in Table 1.

Table 1.
Summary of measurements of Pachydictya bifurcata (Hall, 1883). ELI-Tvären-Ord-16–17.
Remarks. Despite the poor preservation, specimens share median tubuli between the longitudinal row of zooecial tubes and between the median laminae typical of the Family Rhinidictyidae [50,51,52]. These specimens exhibit the characteristics of the genus Pachydictya, including the relevant autozooecial budding patterns, aperture shape, and wall laminar construction. The zooecia are more or less longitudinally arranged in seven to eight rows, without non-porous margins. Based on these features, the specimens are therefore identified as Pachydictya bifurcata. It is similar to the type species Pachydictya robusta in terms of the angle between the zooecia and mesotheca. However, the type species has a larger branch width, approximately 11–13 mm, and develops a non-zooecial margin (non-poriferous margin) that is about 0.6–0.9 mm wide. The type species also has a branch with 24–28 rows of zooecia. Compared with other species such as P. everetti, P. foliata, P. magnipora, P. hexagonalis, and P. gigantea, the Pachydictya bifurcata zoarium is relatively narrow in width, lacks maculae or clusters of large zooecia, and has subparallel margins.
Pachydictya barkholmensis is also very similar to Pachydictya bifurcata, sharing comparable zooecial dimensions and similar internal structures. It was previously proposed that Pachydictya barkholmensis is considered synonymous with Pachydictya bifurcata [15] and it is clear that the two do possess similar zooecia. However, 787 specimens from the Ojl Myr fauna show that Pachydictya barkholmensis only has four to six rows of zooecia, compared to seven to eight rows in Pachydictya bifurcata [21]. The range of zooecial rows in the two species are distinct and do not overlap. This suggests that Pachydictya barkholmensis is not a junior synonym of Pachydictya bifurcata. Specimens in this study also do not overlap in terms of the number of zooecial rows. Pachydictya bifurcata differs from Pachydictya flabellum and Pachydictya elegans due to its larger zooecia, with the latter two typically approximately 0.285 mm transversely and 0.333–0.4 mm longitudinally. Pachydictya flabellum also has more surface ornamentation and Pachydictya elegans possesses poorly developed “rings” or hexagonal areas because of the well-defined peristome around the apertures [15]. The apertures of Pachydictya elegans are also much more rounded than Pachydictya bifurcata. Pachydictya cyclostomoides possesses larger apertures compared to Pachydictya bifurcata, usually about 0.666 mm longitudinally, and also has distinct maculae [15].
Occurrence. Lower Sandbian Stage of Upper Ordovician in the Dalby Limestone at Tvären Bay, the Porkuni (Hirnantian) Stage of Ojl Myr (formerly Öjle Myr), island of Gotland, Sweden and Estonia; the Silurian limestone at Dayton and other localities in Ohio and Indiana of USA. [15,21,53].
Order Trepostomatida Ulrich, 1882.
Family Halloporidae Bassler, 1911.
Genus Hallopora Bassler, 1911.
Type species. Callopora elegantula Hall, 1852, lower Silurian (Niagaran); USA.
Diagnosis. Revised definition from Bassler [15]: Ramose or encrusting zoaria having a columnar structure or form; zooecia rarely septate, the zooecial apertures circular or petaloid but not contiguous; and intermediate spaces occupied by angular transversely septate mesozooecia. Zooecial apertures commonly closed by ornamented perforate covers which become diaphragms when left behind during growth.
Remarks. The name of this genus had often been disputed in the past. Calopora is considered a misspelling, Callopora now refers to a genus of cheilostome bryozoan, and the genus name Hallopora is now preferred [15,54,55,56,57].
Hallopora sp.
Figure 4A–D
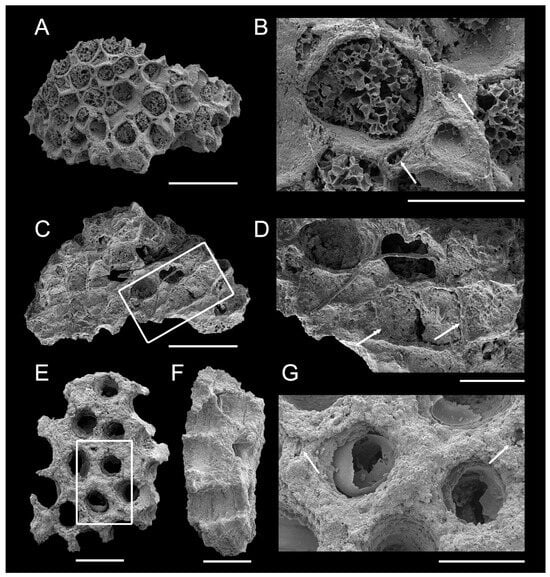
Figure 4.
Trepostomes from the post-impact Dalby limestone of erratic boulders on Ringsö Island, Sweden. (A–D) Hallopora sp. ELI-Tvären-Ord-18. (A) Colony surface, showing the apertures. (B) Partial enlargement of (A), showing the aperture and mesozooecia (single arrow). (C) Longitudinal split of (A), showing long tubular zooecia with diaphragms; box indicates position of (D). (D) Partial enlargement of (C), showing the thin, straight diaphragms (double arrow). (E–G) Trematoporid sp. indet, ELI-Tvären-Ord-19. (E) Colony surface, showing the apertures; box indicates position of (G). (F) Longitudinal split, showing the longitudinal section and vertical walls of three zooecia. (G) Partial enlargement of (E), showing possibly exilazooecia (single arrow). Scale bars 800 μm (A,C), 300 μm (B,D–G).
Material. One specimen (ELI-Tvären-Ord-18) recovered from the Dalby Limestone in Tvären Bay, Sweden.
Description. Colony partially preserved, 2 mm maximum length. Branching not observed. Over 20 autozooeica, long tubular in longitudinal section, subcircular in cross-section of the fracture, diameter ~0.3 mm. Diaphragms common, flat, straight, ~0.2 mm apart. Zooecial walls thin, ~0.02 mm, slightly thickened at apertures, ~0.04–0.06 mm. Zooecial apertures oblique, ovate, partly separated by numerous mesozooecia, internally filled with secondary mineral material. Two to seven mesozooecia present surrounding each zoocium. Mesozooecia 0.04–0.125 mm in diameter, polygonal-oval. Acanthostyles lacking.
Remarks. The material here is identified as Hallopora, belonging to the Halloporidae, based on the characteristics of the zooecial tubes, which are generally subcircular with diaphragms but lacking acanthostyles. Halloporidae can be distinguished from other families of Trepostomata, like the Amplexoporidae and Aisenvergiidae, based on the nature of the zooecia. In Halloporidae, the zooecia are simple prismatic tubes, and exilazooecia are present instead of the mesozooecia [58,59]. The overall morphology of the colony and the structure of the zooecia in the endozone and exozone remains unknown however, making it difficult to assign these specimens to a specific species. Over 70 species of Hallopora have been reported from the Ordovician to the Silurian [60], although some have been subsequently consolidated [16,61]. In this study, Ordovician species widely found in Europe were selected for comparison. Hallopora sp. is similar to Hallopora dumalis from Estonia [15] and Poland [62], particularly in the number and spacing of diaphragms. However, the apertural diameters of zooecia in Hallopora sp. differs slightly from the Polish specimens: seven to eight apertures occur in a 2 mm length in our and the Estonian specimens [15], whereas five to six apertures occur in the same length of Polish specimens [62]. The type species Hallopora elegantula differs from Hallopora sp. in having more mesozooecia [15,21]. Additionally, ornamented and centrally perforated opercula, which are present in Hallopora elegantula and other species [15,21,50,63,64], are not preserved in the specimens examined here. Hallopora tolli has more mesozooecia and larger zooecial openings than Hallopora sp., with densely packed diaphragms, spaced 0.08–0.16 mm apart [15,61]. Such dense diaphragms are also present in Hallopora mckinneyi and Hallopora multipora [16,61]. Mesozooecia in Hallopora multilabulata and Hallopora goodhuensis are fewer in number than in Hallopora sp. [15,65]. Overall, the specimens examined closely match the characteristics of Hallopora dumalis [62] and are clearly distinct from other species. However, more specimens are needed to determine the changes in zooecial features between the endozone and exozone, so this specimen is temporarily classified as Hallopora sp.
Occurrence. lower Sandbian Stage of Upper Ordovician in the Dalby limestone at Tvären Bay.
Family Trematoporidae Miller, 1889
Trematoporid sp. indet
Figure 4E–G
Description. Colony surface flat. Autozooecia apertures oval or circular, ~0.3 mm. Autozooecia loosely arranged, 6–7 per 2 mm. Zooecial walls thickened, ~0.03 mm. Adjacent zooecial walls not fused; layered structure. Exilazooecia or acanthostyles present. No diaphragms in autozooecia or exilazooecia.
Remarks. Apertures of autozooecia are oval or circular, with layered walls and developing immature zooecia (possibly exilazooecia), a feature typical of the suborder Amplexoporina. The thick walls suggest fragmentary colony might be the exozone. The structure of the colony here is generally loose and slightly ambiguous, similar to Trematoporids but quite different from other families.
5. Stratigraphic and Geographical Distribution
The presence of graptolites, conodonts, and other shallow-water biota in the limestone boulders suggests that the boulders originated from the Upper Ordovician Dalby Limestone of the Tvären region [27,28,29,38]. Bassler documented a diverse assemblage of bryozoans from contemporary Baltic strata, predominantly cyclostomes, cryptostomes, and trepostomes, including species such as Pachydictya elegans, Pachydictya cyclostomoides, Hallopora dumalis, and Hallopora tolli [63]. While the bryozoans identified from the boulders in this study includes some of these genera [24], comparative analysis reveals notable differences between Pachydictya bifurcata described herein and the previously reported Pachydictya species. Additionally, the Hallopora specimen in this study shows similarities to Hallopora dumalis, but the absence of definitive diagnostic features precludes a more precise identification. These findings contribute to an increased understanding of the bryozoan diversity in the region. The genus Pachydictya is globally distributed [60], but Pachydictya bifurcata has only been reported from the Baltica and North America regions [15,21,62]. In the Baltica region, Pachydictya bifurcata occurrences are predominantly concentrated within the Harju Series (the Lyckholm and Borkholm limestone, now the Kõrgessaare Formation) of Estonia [15], spanning the Katian to Hirnantian stages. It is also documented in the Porkuni (Hirnantian) Stage of Öjle Myr, Gotland, Sweden [21], as well as in boulders from Poland, where it is among the most abundant bryozoans [62]. Comparative analysis reveals a strong similarity between the bryozoan assemblages of the boulders in Poland and Gotland and the material found in the Kõrgessaare Formation, Estonia [15,21,62]. This resemblance is also the case for other fauna, with comparable assemblages of ostracods, brachiopods, and graptolites, suggesting these strata are correlated [62]. In North America, Pachydictya bifurcata has only been identified in Silurian deposits [15,21]. The discovery of Pachydictya bifurcata in the Lower Sandbian Stage of the post-impact Dalby limestone at Tvären Bay significantly extends the known temporal range of this species within the Baltica region.
6. Summary
We present the first formal report of three palaeostomate bryozoans from limestone boulders on Ringsö Island, Sweden. The bryozoan assemblage includes Pachydictya bifurcata, Hallopora sp., and an indeterminate Trematoporid species. Pachydictya bifurcata is characterized by bifoliate zoaria with two layers of autozooecia oriented in opposite directions along the mesotheca. Its presence here marks a significant stratigraphic extension of this species in Baltica, and it is now documented from the Lower Sandbian Stage of the Upper Ordovician. Hallopora sp. features mesozooecia, tubular autozooecia with rounded apertures, thin walls, and diaphragms, while the zoaria of the indeterminate Trematoporid species are loose and lack diagnostic features for more precise identification. This study enriches the bryozoan diversity previously known from the Tvären region and provides the first detailed report of their presence in this geological context.
Author Contributions
Conceptualization and supervision, Z.Z.; sample collection, L.E.H.; specimen extraction and dissolution, Y.L. and Z.Z.; writing—original draft preparation, B.S.; writing—review and editing, B.S., Y.L., Z.Z., L.E.H., L.C.S. and J.M.; visualization, B.S.; project administration, Z.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by National Key Research and Development Program of China [2023YFF0803601], the National Natural Science Foundation of China [42302010], the Department of Science and Technology of Shaanxi Province [2022TD-11], the Overseas Expertise Introduction Centre for Discipline Innovation [111 Center: D17013], the Swedish Research Council [VR Project no. 2018-03390], and the China Scholarship Council [no. CSC202306970008].
Data Availability Statement
The specimens are deposited in the State Key Laboratory of Continental Dynamics, Shaanxi Key Laboratory of Early Life and Environments, Department of Geology, Northwest University, Xi’an 710069, China.
Acknowledgments
This paper is a contribution to IGCP 735. We are grateful to Juanping Zhai, Ruisen Fang (Northwest University, China) for their kindly help in the experimental treatment of samples. We thank the editor and three anonymous reviewers for their critical and helpful comments and efforts to improve our manuscript.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Schwaha, T. Morphology of bryozoans. In Phylum Bryozoa; Schwaha, T., Ed.; De Gruyter: Boston, MA, USA, 2020; pp. 57–100. [Google Scholar]
- Taylor, P.D. Bryozoan Paleobiology; John Wiley & Sons: London, UK, 2020; pp. 1–336. [Google Scholar]
- Zhang, Z.; Zhang, Z.; Ma, J.; Taylor, P.D.; Strotz, L.C.; Jacquet, S.M.; Skovsted, C.B.; Chen, F.; Han, J.; Brock, G.A. Fossil Evidence Unveils an Early Cambrian Origin for Bryozoa. Nature 2021, 599, 251–255. [Google Scholar] [CrossRef] [PubMed]
- Pruss, S.B.; Leeser, L.; Smith, E.F.; Zhuravlev, A.Y.; Taylor, P.D. The oldest mineralized bryozoan? A possible palaeostomate in the lower Cambrian of Nevada, USA. Sci. Adv. 2022, 8, eabm8465. [Google Scholar] [CrossRef] [PubMed]
- Horowitz, A.S.; Pachut, J.F. The fossil record of bryozoan species diversity. In Proceedings of the 11th International Bryozoology Association Conference, Smithsonian Tropical Research Institute, Panama City, Panama, 26–31 January 1998; Herrera Cubilla, A., Jackson, J.B.C., Eds.; Smithsonian Tropical Research Institute: Panama City, Panama, 2000; pp. 245–248. [Google Scholar]
- Gordon, D.P.; Taylor, P.D.; Bigey, F.P. Phylum Bryozoa. In New Zealand Inventory of Biodiversity; Gordon, D.P., Ed.; Canterbury University Press: Canterbury, UK, 2009; Volume 1, pp. 271–297. [Google Scholar]
- Bock, P.E.; Gordon, D.P. Phylum Bryozoa Ehrenberg, 1831. Zootaxa 2013, 3703, 67–74. [Google Scholar]
- Gordon, D.P.; Costello, M.J. Bryozoa—Not a minor phylum. New Zealand Sci. Rev. 2016, 73, 63–66. [Google Scholar] [CrossRef]
- Ma, J.; Buttler, C.J.; Taylor, P.D. Cladistic analysis of the ‘trepostome’ Suborder Esthonioporina and the systematics of Palaeozoic bryozoans. Studi Trentini Di Sci. Nat. 2014, 94, 153–161. [Google Scholar]
- Ernst, A. Fossil record and evolution of Bryozoa. In Phylum Bryozoa; Schwaha, T., Ed.; De Gruyter: Boston, MA, USA, 2020; pp. 11–55. [Google Scholar]
- Ernst, A. Diversity dynamics of Ordovician bryozoa. Lethaia 2018, 51, 198–206. [Google Scholar] [CrossRef]
- Ernst, A. Fossilized soft tissues in Palaeozoic bryozoans. Pap. Palaeontol. 2023, 9, e1483. [Google Scholar] [CrossRef]
- Hageman, S.J.; Ernst, A. The last phylum: Occupation of Bryozoa morpho-ecospace (colony growth habits) during the early phase of the Great Ordovician Biodiversification Event. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2019, 534, 109270. [Google Scholar] [CrossRef]
- Eichwald, E. Zoologia Specialis Quam Expositis Animalibus Tum Vivis, Fom Fossilibus Potissimum Rossiae in Universum, et Poloniae in Species, Volume. 1. Bryozoa; Typis Josephi Zawadzki, Vilnae: Vilnius, Lithuania, 1829; pp. 1–334. [Google Scholar]
- Bassler, R.S. The early Palaeozoic Bryozoa of the Baltic Provinces. United States Natl. Mus. Bull. 1911, 77, 1–382. [Google Scholar]
- Brood, K. Upper Ordovician Bryozoa from Dalmantina beds of Boreahult, Ösergötlan, Sweden. Geol. Palaeontol. 1978, 12, 53–72. [Google Scholar]
- Brood, K. Bryozoa from the Upper Ordovician Dalmantina beds of Kinnekule, Sweden. Geol. Föreningensis I Stockh. Förhandlingar 1980, 102, 27–35. [Google Scholar] [CrossRef]
- Brood, K. Hirnantian (Upper Ordovician) Bryozoa from Baltoscandia. In Recent and Fossil Bryozoa. International Conference on Bryozoa; Larwood, G.P., Nielsen, C., Eds.; Olsen & Olsen: Fredensborg, Denmark, 1981; pp. 19–27. [Google Scholar]
- Brood, K. Ashgill Bryosoa from a fissure filling at Solberga, Dalarna. Geol. Föreningensis I Stockh. Förhandlingar 1982, 104, 167–181. [Google Scholar] [CrossRef]
- Brood, K. Bryozoa from the Klingkalk at Jalltjarn in Dalarna, Sweden. Stockh. Contrib. Geol. 1982, 37, 43–48. [Google Scholar]
- Spjeldnaes, N. Upper Ordovician bryozoans from Öjle Myr, Gotland, Sweden. Bull. Geol. Inst. Univ. Upps. 1984, 10, 1–66. [Google Scholar]
- Ernst, A. Inconobotopora (Cystoporata, Bryozoa) from the Kullsberg Limestone (early Katian, Late Ordovician) of Siljan District, Dalarna, central Sweden. GFF 2012, 134, 19–25. [Google Scholar] [CrossRef]
- Ernst, A.; Tolokonnikova, Z. Unusual cystoporate? bryozoan from the Upper Ordovician of Siljan District, Dalarna, central Sweden. GFF 2022, 144, 210–219. [Google Scholar] [CrossRef]
- Thorslund, P. On the Chasmops Series of Jemtland and Södermanland (Tvären). Sver. Geol. Undersökning C 1940, 436, 1–191. [Google Scholar]
- Jaanusson, V. Middle Ordovician Ostracodes of Central and Southern Sweden. Bull. Geol. Inst. Univ. Upps. 1957, 37, 173–442. [Google Scholar]
- Strachan, I. Graptolites from the Ludibundus beds (Middle Ordovician) of Tvären, Sweden. Bull. Geol. Inst. Univ. Upps. 1959, 38, 1–68. [Google Scholar]
- Bergström, S.M. Conodonts from the Ludibundus Limestone (Middle Ordovician) of the Tvären area (S.E. Sweden). Ark. För Mineral. Och Geol. 1962, 3, 1–61. [Google Scholar]
- Lindström, M.; Flodén, T.; Grahn, Y.; Kathol, B. Post-impact deposits in Tvären, a marine Ordovician crater south of Stockholm, Sweden. Geol. Mag. 1994, 131, 91–103. [Google Scholar] [CrossRef]
- Ormö, J. The pre-impact Ordovician stratigraphy of the Tvären Bay impact structure, SE Sweden. GFF 1994, 116, 139–144. [Google Scholar] [CrossRef]
- Grahn, Y.; Nõlvak, J.; Paris, F. Precise chitinozoan dating of Ordovician impact events in Baltoscandia. J. Micropaleontol. 1996, 15, 21–35. [Google Scholar] [CrossRef]
- Frisk, Å.M.; Holmer, L.E. Linguliform and craniiform brachiopods from the Ordovician Tvären Crater, Sweden. In Abstracts: Fifth International Brachiopod Congress, Copenhagen, Denmark, 2005; Harper, D.A.T., Long, S.L., McCorry, M., Eds.; Geological Museum and GEUS: Copenhagen, Denmark, 2005. [Google Scholar]
- Frisk, Å.M.; Ormö, J. Facies distribution of post-impact sediments in the Ordovician Lockne and Tvären impact craters: Indications for unique impact-generated environments. Meteorit. Planet. Sci. 2007, 42, 1971–1984. [Google Scholar] [CrossRef]
- Ormö, J.; Sturkell, E.; Lindström, M. Sedimentological analysis of resurge deposits at the Lockne and Tvären craters: Clues to flow dynamics. Meteorit. Planet. Sci. 2007, 42, 1929–1943. [Google Scholar] [CrossRef]
- Henkel, H.; Ekneligoda, T.C.; Aaro, S. The extent of impact induced fracturing from gravity modeling of the Granby and Tvären simple craters. Tectonophysics 2010, 485, 290–305. [Google Scholar] [CrossRef]
- Eriksson, M.E.; Frisk, Å.M. Polychaete palaeoecology in an early Late Ordovician marine astrobleme of Sweden. Geol. Mag. 2011, 148, 269–287. [Google Scholar] [CrossRef]
- Frisk, Å.M.; Harper, D.A. Late Ordovician brachiopod distribution and ecospace partitioning in the Tvären crater system, Sweden. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2013, 369, 114–124. [Google Scholar] [CrossRef]
- Jaanusson, V. Unterordovicische Illaeniden aus Skandinavien. Bull. Geol. Inst. Univ. Upps. 1957, 37, 79–165. [Google Scholar]
- Frisk, Å.M.; Ebbestad, J.O.R. Paragastropods, tergomya, and gastropoda (Mollusca) from the Upper Ordovician Dalby Limestone, Sweden. GFF 2007, 129, 83–99. [Google Scholar] [CrossRef]
- Frisk, Å.M.; Harper, D.A.T. Palaeoenvironmental aspects of Late Ordovician Sericoidea shell concentrations in an impact crater, Tvären, Sweden. Lethaia 2011, 44, 383–396. [Google Scholar] [CrossRef]
- Lindström, M.; Sturkell, E.F.F. Geology of the Early Palaeozoic Lockne impact structure, central Sweden. Tectonophysics 1992, 216, 169–185. [Google Scholar] [CrossRef]
- Grahn, Y.; Nõlvak, J. Chitinozoan dating of Ordovician impact events in Sweden and Estonia. A preliminary note. Geol. Föreningens I Stockh. Förhandlingar 1993, 115, 263–264. [Google Scholar] [CrossRef]
- Lindström, M.; Shuvalov, V.; Ivanov, B. Lockne crater as a result of marine-target oblique impact. Planet. Space Sci. 2005, 53, 803–815. [Google Scholar] [CrossRef]
- Lindström, M.; Ormö, J.; Sturkell, E.F.F.; von Dalwigk, I. The Lockne Crater: Revision and reassessment of structure and impact stratigraphy. In Impact Tectonics. Impact Studies; Koeberl, C., Henkel, H., Eds.; Springer: Berlin/Heidelberg, Germany, 2005; pp. 357–388. [Google Scholar]
- Grahn, Y. Chitinozoan biostratigraphy of the Early Caradocian Lockne impact structure, Jämtland, Sweden. Meteorit. Planet. Sci. 1997, 32, 745–747. [Google Scholar] [CrossRef]
- Lindström, M. On the Lower Chasmops beds in the Fågelsång district (Scania). Geol. Föreningens I Stockh. Förhandlingar 1953, 75, 125–148. [Google Scholar] [CrossRef]
- Grahn, Y.; Nõlvak, J. Swedish Ordovician Chitinozoa and biostratigraphy: A review and new data. Palaeontogr. Abt. B 2010, 283, 1–67. [Google Scholar] [CrossRef]
- Nõlvak, J.; Grahn, Y. Ordovician chitinozoan zones from Baltoscandia. Rev. Palaeobot. Palynol. 1993, 79, 245–269. [Google Scholar] [CrossRef]
- Nõlvak, J.; Grahn, Y.; Sturkell, E.F.F. Chitinozoan biostratigraphy of the Middle Ordovician Dalby Limestone in the Fjäcka section, Siljan district, Sweden. Proc. Est. Acad. Sci. Geol. 1999, 48, 75–85. [Google Scholar]
- Bergström, S.M. Conodont biostratigraphy of the Middle and Upper Ordovician of Europe and eastern North America. Geol. Soc. Am. Mem. 1970, 127, 83–162. [Google Scholar]
- Ulrich, E.O. American Paleozoic Bryozoa. Cincinnati Soc. Nat. Hist. 1882, 5, 121–175. [Google Scholar]
- Ulrich, E.O. New Lower Silurian Bryozoa. J. Cincinnati Soc. Nat. Hist. 1890, 12, 173–198. [Google Scholar]
- Ulrich, E.O. On Lower Silurian Bryozoa of Minnesota. Geol. Nat. Hist. Surv. Minn. 1893, 3, 96–332. [Google Scholar]
- Wiman, C. Über die Borkholmer Schicht im Mittelbaltischen Silurgebiet. Bull. Geol. Inst. Univ. Upps. 1901, 5, 149–222. [Google Scholar]
- Fritz, M.A. Redescription of type specimens of bryozoan Hallopora from upper Ordovician of Toronto region, Ontario. Proc. Geol. Assoc. Can. 1970, 21, 15–23. [Google Scholar]
- Ross, J.R. Discussion of ‘On the validity of the name Hallopora for a genus of trepostome bryozoan’ by RJ Singh. J. Paleontol. 1970, 44, 785. [Google Scholar]
- Singh, R.J. On the validity of the name Hallopora for a genus of trepostome bryozoan. J. Paleontol. 1970, 44, 784–785. [Google Scholar]
- Corneliussen, E.F.; Perry, T.G. The ectoproct Batostoma? cornula (Cumings & Galloway) and its enigmatic intrazooecial spines [Fort Atkinson Limestone (Cincinnatian), Wilmington, Illinois]. J. Paleontol. 1970, 44, 997–1008. [Google Scholar]
- Astrova, G.G. Morphologiya, istoriya razvitiya i sistema ordoviksikh i siluriyskikh mshanok (Morphology, history of development, and systematics of Ordovician and Silurian Bryozoa). Trans. Paleontol. Instituta Akad. Nauk SSSR 1965, 106, 1–432. [Google Scholar]
- Astrova, G.G. The history of development, system, and phylogeny of the Bryozoa: Order Trepostomata. Trans. Paleontol Inst Akad Nauk SSSR 1978, 169, 1–240. [Google Scholar]
- The Bryozoa Home Page. Available online: http://bryozoa.net/trepostomatida/halloporidae/hallopora.html (accessed on 13 August 2024).
- Pushkin, V.I. Middle Ordovician bryozoa from the Podlasie-Brest Depression (Belarussian part): Suborders Ceramoporina, Esthonioporina, Amplexoporina, Halloporina. Geol. Quart. 2002, 46, 411–434. [Google Scholar]
- Kiepura, M. Bryozoa from the Ordovician erratic boulders of Poland. Acta Palaeontol. Pol. 1962, 7, 347–428. [Google Scholar]
- Bassler, R.S. Bryozoa. Treatise on Invertebrate Paleontology; University of Kansas Press and Geological Society of America: Lawrence, KS, USA, 1953. [Google Scholar]
- Bassler, R.S. The Bryozoan Fauna of the Rochester Shale; United States Geological Survey Bulletin: Washington, DC, USA, 1906; Volume 292, pp. 1–65.
- Ulrich, E.O. Report on the Lower Silurian Bryozoa with preliminary descriptions of some of the new species. Minn. Geol. Nat. Hist. Surv. Annu. Rep. 1886, 14, 57–103. [Google Scholar]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).