
A Case of Predominance of Alicyclobacillus tolerans in Microbial Community during Bioleaching of Pentlandite-Chalcopyrite Concentrate




Abstract
:1. Introduction
2. Materials and Methods
2.1. Substrate and Conditions for Bioleaching in Bioreactor
2.2. Microbial Community Used as Inoculum for Bioleaching
2.3. Isolation and Characterization of Predominant Microorganisms during Bioleaching
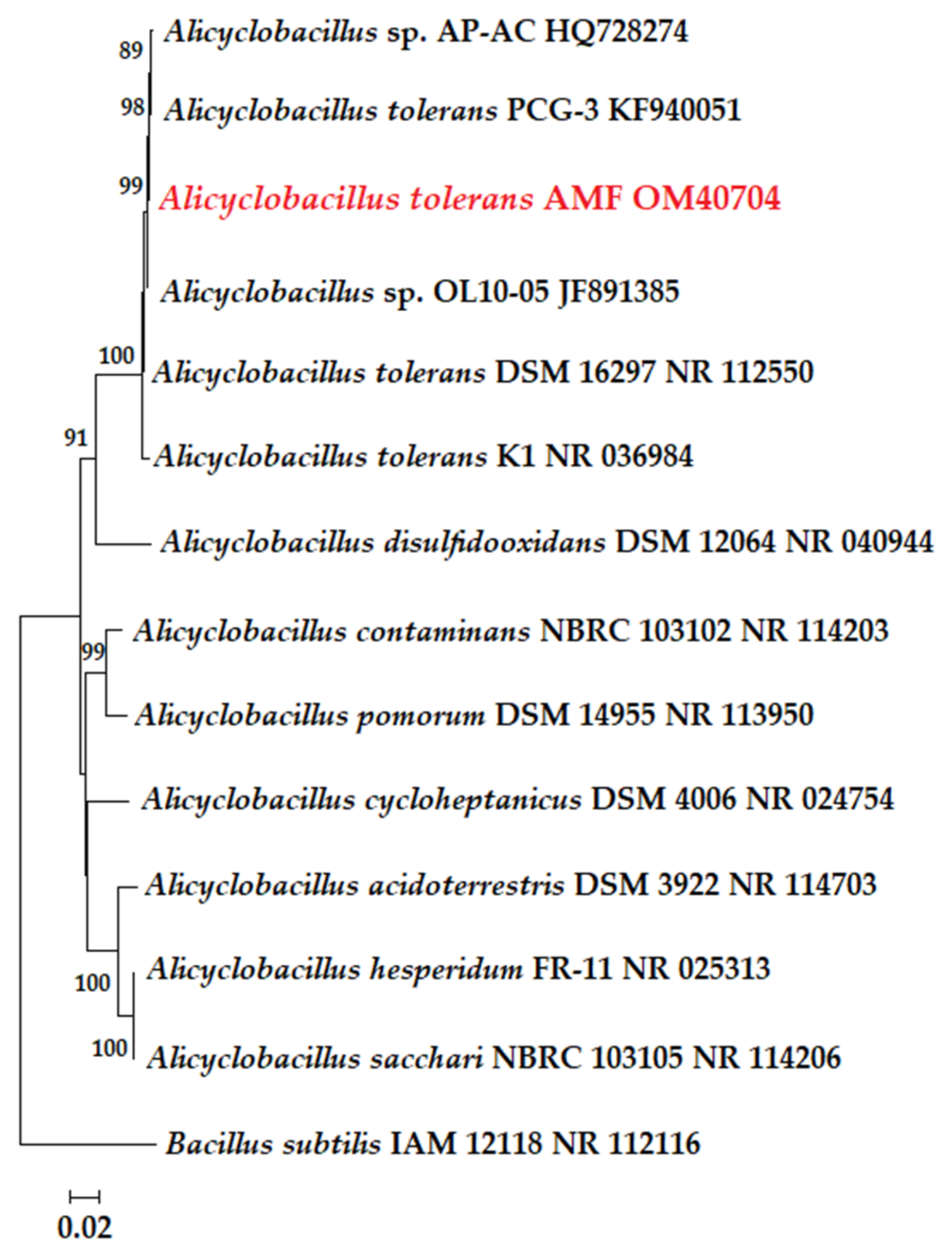
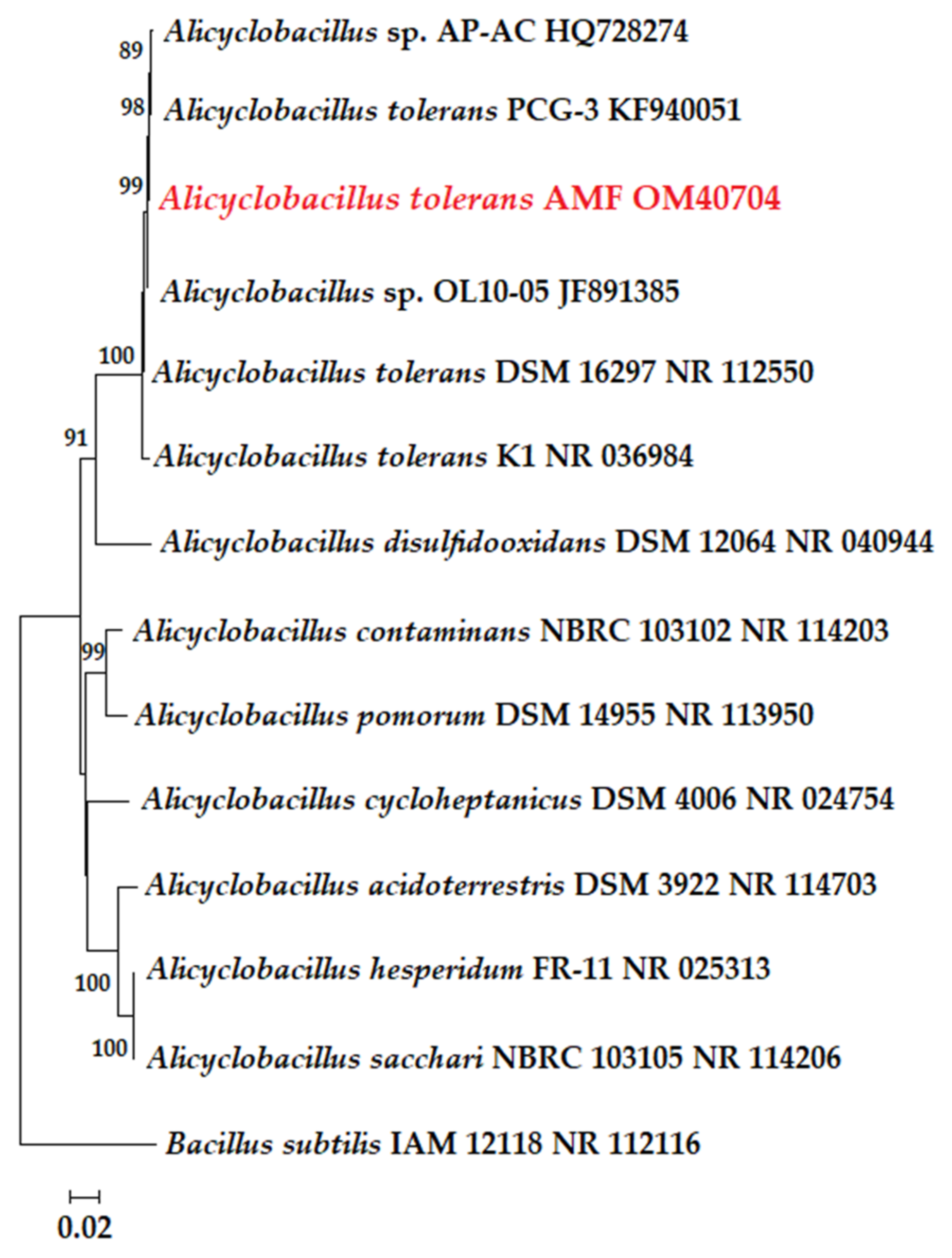
2.4. Taxonomic Research and Phylogenetic Analysis
2.5. Analytical Methods
3. Results and Discussion
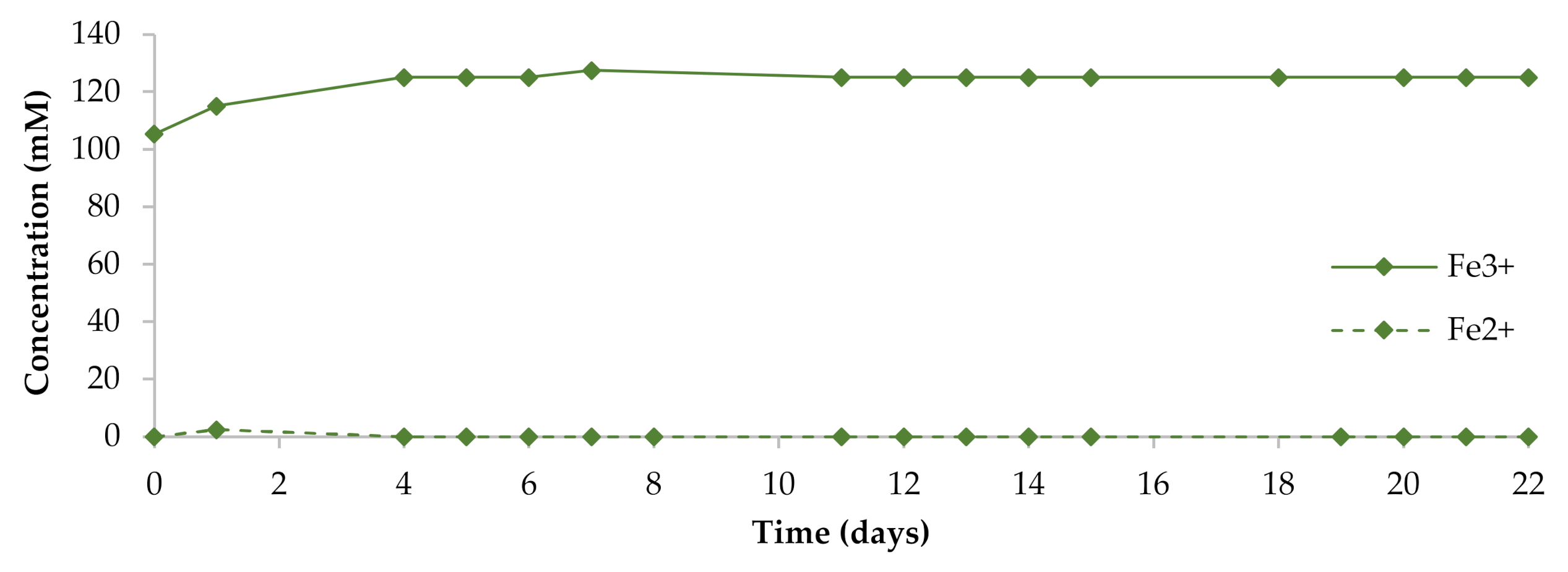
3.1. Parameters of Liquid Phase of Pulp during Bioleaching
3.2. Isolation and Identification of Strains Dominating Bioleaching Microbial Community
3.3. Characterization of Al. tolerans AMF Growth and Substrate Oxidation
3.4. Succession of Microorganisms during Violarite–Pentlandite–Chalcopyrite Concentrate Bioleaching and Role of Al. tolerans AMF in Microbial Community
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kaksonen, A.H.; Deng, X.; Bohu, T.; Zea, L.; Khaleque, H.N.; Gumulya, Y.; Boxall, N.J.; Morris, C.; Cheng, K.Y. Prospective directions for biohydrometallurgy. Hydrometallurgy 2020, 195, 105376. [Google Scholar] [CrossRef]
- Kondrat’eva, T.F.; Pivovarova, T.A.; Tsaplina, I.A.; Fomchenko, N.V.; Zhuravleva, A.E.; Murav’ev, M.I.; Melamud, V.S.; Bulayev, A.G. Diversity of the communities of acidophilic chemolithotrophic microorganisms in natural and technogenic ecosystems. Microbiology 2012, 81, 1–24. [Google Scholar] [CrossRef]
- Mahmoud, A.; Cezac, P.; Hoadley, A.F.A.; Contamine, F.; D’Hugues, P. A review of sulfide minerals microbially assisted leaching in stirred tank reactors. Int. Biodeterior. Biodegrad. 2017, 119, 118–146. [Google Scholar] [CrossRef]
- Panyushkina, A.; Fomchenko, N.; Babenko, V.; Muravyov, M. Effect of temperature on biobeneficiation of bulk copper–nickel concentrate with thermoacidophilic microbial communities. Metals 2021, 11, 1969. [Google Scholar] [CrossRef]
- Kaksonen, A.H.; Lakaniemi, A.-M.; Tuovinen, O.H. Acid and ferric sulfate bioleaching of uranium ores: A review. J. Clean. Prod. 2020, 264, 121586. [Google Scholar] [CrossRef]
- Li, J.; Yang, H.; Tong, L.; Sand, W. Some aspects of industrial heap bioleaching technology: From basics to practice. Miner. Process. Extr. Metall. Rev. 2021. [Google Scholar] [CrossRef]
- Fomchenko, N.V.; Muravyov, M.I. Two-step biohydrometallurgical technology of copper-zinc concentrate processing as an opportunity to reduce negative impacts on the environment. J. Environ. Manag. 2018, 226, 270–277. [Google Scholar] [CrossRef]
- Harvey, T.J.; Yen, W.T.; Paterson, J.G. Selective zinc extraction from complex copper/zinc sulphide concentrates by pressure oxidation. Miner. Eng. 1992, 5, 975–992. [Google Scholar] [CrossRef]
- Muravyov, M.; Panyushkina, A.; Bulaev, A.; Fomchenko, N. Biobeneficiation of bulk copper-zinc and copper–nickel concentrates at different temperatures. Miner. Eng. 2021, 170, 107040. [Google Scholar] [CrossRef]
- Arpalahti, A.; Lundström, M. The leaching behavior of minerals from a pyrrhotite-rich pentlandite ore during heap leaching. Miner. Eng. 2018, 119, 116–125. [Google Scholar] [CrossRef]
- Watling, H.R. The bioleaching of nickel-copper sulfides. Hydrometallurgy 2008, 91, 70–88. [Google Scholar] [CrossRef]
- Watling, H.R. The bioleaching of sulphide minerals with emphasis on copper sulphides: A review. Hydrometallurgy 2006, 84, 81–108. [Google Scholar] [CrossRef]
- Norris, P.R. Acidophile diversity in mineral sulfide oxidation. In Biomining; Rawlings, D.E., Johnson, D.B., Eds.; Springer: Berlin, Germany, 2007; pp. 199–216. [Google Scholar]
- Gericke, M.; Govender, Y. Bioleaching strategies for the treatment of nickel-copper sulphide concentrates. Miner. Eng. 2011, 24, 1106–1112. [Google Scholar] [CrossRef]
- Wisotzkey, J.D.; Jurtshuk, P.; Fox, G.E.; Deinhart, G.; Poralla, K. Comparative sequence analyses on the 16S rRNA (rRNA) of Bacillus acidocaldarius, Bacillus acidoterrestris, and Bacillus cycloheptanicus and proposal for creation of a new genus, Alicyclobacillus gen. nov. Int. J. Syst. Bacteriol. 1992, 42, 263–269. [Google Scholar] [CrossRef]
- Ciuffreda, E.; Bevilacqua, A.; Sinigaglia, M.; Corbo, M.R. Alicyclobacillus spp.: New insights on ecology and preserving food quality through new approaches. Microorganisms 2015, 3, 625–640. [Google Scholar] [CrossRef] [Green Version]
- Muravyov, M.; Panyushkina, A. Distinct roles of acidophiles in complete oxidation of high-sulfur ferric leach product of zinc sulfide concentrate. Microorganisms 2020, 8, 386. [Google Scholar] [CrossRef] [Green Version]
- Silverman, M.P.; Lundgren, D.G. Studies on the chemoautotrophic iron bacterium Ferrobacillus ferrooxidans. I. An improved medium and a harvesting procedure for securing high cell yields. J. Bacteriol. 1959, 77, 642–647. [Google Scholar] [CrossRef] [Green Version]
- Panyushkina, A.E.; Tsaplina, I.A.; Grigor’eva, N.V.; Kondrat’eva, T.F. Thermoacidophilic microbial community oxidizing the gold-bearing flotation concentrate of a pyrite-arsenopyrite ore. Microbiology 2014, 83, 539–549. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA 11: Molecular evolutionary genetics analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Tamura, K.; Nei, M.; Kumar, S. Prospects for inferring very large phylogenies by using the neighbor-joining method. Proc. Natl. Acad. Sci. USA 2004, 101, 11030–11035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, D.G.; Jacobsen, W.R. Determination of iron and iron-aluminum mixtures by titration with EDTA. Anal. Chem. 1960, 32, 215–217. [Google Scholar] [CrossRef]
- Panyushkina, A.E.; Babenko, V.V.; Nikitina, A.S.; Selezneva, O.V.; Tsaplina, I.A.; Letarova, M.A.; Kostryukova, E.S.; Letarov, A.V. Sulfobacillus thermotolerans: New insights into resistance and metabolic capacities of acidophilic chemolithotrophs. Sci. Rep. 2019, 9, 15069. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karavaiko, G.I.; Bogdanova, T.I.; Tourova, T.P.; Kondrat’eva, T.F.; Tsaplina, I.A.; Egorova, M.A.; Krasil’nikova, E.N.; Zakharchuk, L.M. Reclassification of ‘Sulfobacillus thermosulfidooxidans subsp. thermotolerans’ strain K1 as Alicyclobacillus tolerans sp. nov. and Sulfobacillus disulfidooxidans Dufresne et al. 1996 as Alicyclobacillus disulfidooxidans comb. nov., and emended description of the genus Alicyclobacillus. Int. J. Syst. Evol. Microbiol. 2005, 55, 941–947. [Google Scholar]
- Duda, V.I.; Suzina, N.E.; Severina, L.O.; Bogdanova, T.I.; Tsaplina, I.A.; Karavaiko, G.I. Ultrastructural organization of Alicyclobacillus tolerans strain K1T cells. Arch. Microbiol. 2006, 185, 63–68. [Google Scholar] [CrossRef]
- Behera, S.K.; Manjaiah, M.; Sekar, S.; Panda, S.K.; Mavumengwana, V.; Mulaba-Bafubiandi, A.F. Optimization of microbial leaching of base metals from a South African sulfidic nickel ore concentrate by Acidithiobacillus ferrooxidans. Geomicrobiol. J. 2018, 35, 447–459. [Google Scholar] [CrossRef]
- Watling, H.R.; Perrot, F.A.; Shiers, D.W. Comparison of selected characteristics of Sulfobacillus species and review of their occurrence in acidic and bioleaching environments. Hydrometallurgy 2008, 93, 57–65. [Google Scholar] [CrossRef]
- Huynh, D.; Giebner, F.; Kaschabek, S.R.; Rivera-Araya, J.; Levican, G.; Sand, W.; Schlömann, M. Effect of sodium chloride on Leptospirillum ferriphilum DSM 14647T and Sulfobacillus thermosulfidooxidans DSM 9293T: Growth, iron oxidation activity and bioleaching of sulfidic metal ores. Miner. Eng. 2019, 138, 52–59. [Google Scholar] [CrossRef]
- Tsaplina, I.A.; Zhuravleva, A.E.; Ismailov, A.D.; Zakharchuk, L.M.; Krasil’nikova, E.N.; Bogdanova, T.I.; Karavaiko, G.I. The dependence of intracellular ATP level on the nutrition mode of the acidophilic bacteria Sulfobacillus thermotolerans and Alicyclobacillus tolerans. Microbiology 2007, 76, 654–662. [Google Scholar] [CrossRef]
- Panyushkina, A.E.; Tsaplina, I.A.; Kondrat’eva, T.F.; Belyi, A.V.; Bulaev, A.G. Physiological and morphological characteristics of acidophilic bacteria Leptospirillum ferriphilum and Acidithiobacillus thiooxidans, members of a chemolithotrophic microbial consortium. Microbiology 2018, 87, 326–338. [Google Scholar] [CrossRef]
- Bulaev, A.G.; Erofeeva, T.V.; Labyrich, M.V.; Mel’nikova, E.A. Resistance of Acidiplasma archaea to heavy metal ions. Microbiology 2017, 86, 583–589. [Google Scholar] [CrossRef]
- Golyshina, O.V.; Pivovarova, T.A.; Karavaiko, G.I.; Kondrat’eva, T.F.; Moore, E.R.B.; Abraham, W.-R.; Lünsdorf, H.; Timmis, K.N.; Yakimov, M.M.; Golyshin, P.N. Ferroplasma acidiphilum gen. nov., sp. nov., an acidophilic, autotrophic, ferrous-iron-oxidizing, cell-wall-lacking, mesophilic member of the Ferroplasmaceae fam. nov., comprising a distinct lineage of the Archaea. Int. J. Syst. Evol. Microbiol. 2000, 50, 997–1006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehrotra, A.; Sreekrishnan, T.R. Heavy metal bioleaching and sludge stabilization in a single-stage reactor using indigenous acidophilic heterotrophs. Environ. Technol. 2017, 38, 2709–2724. [Google Scholar] [CrossRef] [PubMed]
- Korehi, H.; Blöthe, M.; Sitnikova, M.A.; Dold, B.; Schippers, A. Metal mobilization by iron- and sulfur-oxidizing bacteria in a multiple extreme mine tailings in the Atacama Desert, Chile. Environ. Sci. Technol. 2013, 47, 2189–2196. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Cp 1 | Pn 2 | Vo 3 | Py 4 | Po 5 | S | Fe | Si | Cu | Ni | Ca |
|---|---|---|---|---|---|---|---|---|---|---|
| 31 | 12 | 8 | 4 | 6 | 26.5 | 21.8 | 18.0 | 10.8 | 7.2 | 0.96 |
| Parameter | Day 0 | Day 22 |
|---|---|---|
| pH | 1.15 | 1.10 |
| Redox potential (mV) | 910 | 930 |
| Cu, mM | 0 | 1.33 |
| Ni, mM | 0 | 11.9 |
| Cell number, cells/mL | 8.0 × 108 | 25.0 × 108 |
| Characteristic | AMF | K1 |
|---|---|---|
| Source of Isolation | Stirred-tank bioleaching of sulfide copper–nickel concentrate | Oxidizable lead–zinc ores of the Kurgashinkan deposit (Uzbekistan) |
| Temperature, °C | 35–50 | 37–42 (temperature optimum) |
| pH | 1.0–2.2 | 1.5–5.0 (optimum, 2.5–2.7) |
| Morphology | Rods | Rods |
| Cell size, µm | 2.5–4.0 × 1.0–1.2 | 3.0–6.0 × 0.9–1.0 |
| Motility | No | No |
| Sporulation | Terminal or subterminal, oval endospores | Terminal or subterminal, oval, thermally resistant endospores (viable after 30 min heating at 110 °C) |
| Growth with: | ||
| Fe | + 2 (2 culture transfers) | + (2 culture transfers) |
| Fe + YE 1 | + | + (optimal) |
| S0 | – 3 | + (2 culture transfers) |
| S0 + YE | – | + |
| YE | + | + (6 culture transfers) |
| References | Present study; [4] | [26,27] |
| Microorganism | Iron oxidation Rate (mM h−1) | Reference |
|---|---|---|
| S. thermosulfidooxidans VKM B-1269T | 2.9 | [29] |
| S. thermosulfidooxidans VKM B-1269T | 1.91 | [30] |
| S. thermotolerans Kr1T | 2.1 | [31] |
| S. thermotolerans Kr1T | 2.2 | [29] |
| L. ferriphilum DSM 14647T | 1.7 | [30] |
| L. ferriphilum P1 | 2.2 | [32] |
| Ad. aeolicum V1T | 2.1 | [33] |
| Ad. cupricumulans BH2T | 2.1 | [33] |
| F. acidiphilum YT | 0.94 | [34] |
| Al. tolerans K1T | 0.36 | [31] |
| Al. tolerans AMF | 0.83 | This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Panyushkina, A.; Muravyov, M.; Fomchenko, N. A Case of Predominance of Alicyclobacillus tolerans in Microbial Community during Bioleaching of Pentlandite-Chalcopyrite Concentrate. Minerals 2022, 12, 396. https://doi.org/10.3390/min12040396
Panyushkina A, Muravyov M, Fomchenko N. A Case of Predominance of Alicyclobacillus tolerans in Microbial Community during Bioleaching of Pentlandite-Chalcopyrite Concentrate. Minerals. 2022; 12(4):396. https://doi.org/10.3390/min12040396
Chicago/Turabian StylePanyushkina, Anna, Maxim Muravyov, and Natalya Fomchenko. 2022. "A Case of Predominance of Alicyclobacillus tolerans in Microbial Community during Bioleaching of Pentlandite-Chalcopyrite Concentrate" Minerals 12, no. 4: 396. https://doi.org/10.3390/min12040396
APA StylePanyushkina, A., Muravyov, M., & Fomchenko, N. (2022). A Case of Predominance of Alicyclobacillus tolerans in Microbial Community during Bioleaching of Pentlandite-Chalcopyrite Concentrate. Minerals, 12(4), 396. https://doi.org/10.3390/min12040396