

Comparative Study of Formation Conditions of Fe-Mn Ore Microbialites Based on Mineral Assemblages: A Critical Self-Overview

Abstract
1. Introduction
- (1)
- Why the enzymatic Mn oxidation under obligatory oxic conditions is the main primary process during formation of manganese ore deposits; why enzymatic activity requires obligatory oxic redox conditions. Details are given in Ref. [1].
- (2)
- (3)
- (4)
- (5)
- We highlighted the selective element enrichments resulting from microbial mediation and pointed out that geochemical ratio studies must be used with caution (e.g., Mg selective enrichment on cell material, microbial oxidation of Ce and Co by Mn oxidizing bacteria) [1].
- (6)
- (7)
- We pointed out the role of interpretation of mineral assemblages in the deposits, which also represent aspects of environmental conditions. The fact is that the primary ore formation processes are similar, but the output can be very different in terms of the local effects and the mass balance conditions (e.g., concentration of buried organic matter, its reactivity, concentration of metal oxides and other minerals, accumulation rate).
- (8)
- These Fe and Mn ore systems are dual microbial systems, as the Fe microbial system is present together with the Mn microbial system (Table 1). The strength of each can differ, but they occur together in an intimate connection (Raman profiles are offered in the papers). Diagenesis results in variable mineralogy of the original Fe and Mn biomats.
2. Studied Deposits
3. General Framework
3.1. Sketch of Syngenetic and Diagenetic Network
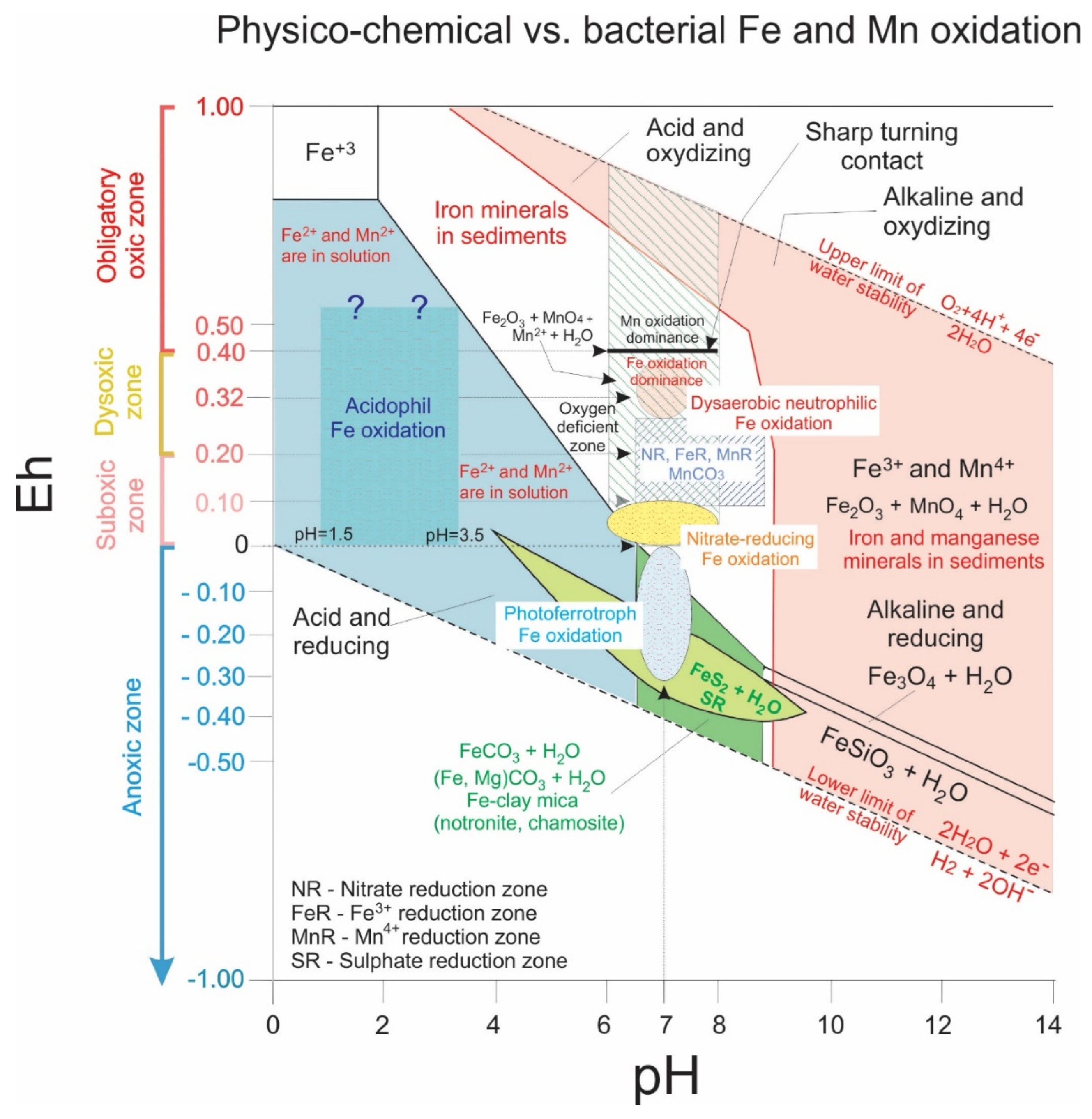
3.2. Fe Accumulations
- -
- acidophilic and oxic;
- -
- photoferrotroph, driven by light, occurring in anoxic/anaerobic and neutrophilic conditions;
- -
- suboxic/anaerobic, where neutrophilic NO3− reducers coupled with Fe(II) oxidizers contribute to the biochemical milieu;
- -
- suboxic and neutrophilic, e.g., Gallionella (Mariprofundus)-like Fe-oxidizing microbes, which are common in many environments.
3.3. Mn Accumulations
3.4. Consequences of Very Small Grain Size
3.5. Preservation of Metastable Minerals
- -
- The geodynamic situation refers to a rifting (or failed rifting) zone, and distal hydrothermal discharge is the metal source;
- -
- Oxygen supply is important: it is suboxic in the case of Fe and obligatory oxic in the case of Mn (to form a deposit, a very effective enzymatic enrichment engine is needed). The oxygen supply can be due to currents or cyanobacterial ventilation, and even small variations can determine what kind of deposit will be formed or not formed (suboxic: Fe, obligatory oxic: Mn);
- -
- Starving basin conditions are required. If a considerable contribution of debris takes place, the microbial biomat system will be destroyed;
- -
- Enrichment process: enzymatic selective element enrichment is very effective (enzymatic engine and its redox demand).
- -
- What cause differences? We can mention accumulation ratio differences and mass balance differences (accumulating organic matter (type, mass), metal oxide concentration and type, other forming minerals and clay minerals). These factors result in differences among deposits both in ore and other mineral content (Table 1).
3.6. Geochemical Conditions
3.7. Aspects of Biogenicity
3.7.1. Biogenic Origin
- (1)
- Microbial microtexture: biomat, filamentous (often sequented), coccoid like, vermiform; brain-like and stromatolite-like macrotexture—fine lamination multiple mineral cyclicity;
- (2)
- Bioindicator minerals that can be modified by diagenetic and other processes, mineralized cycles, and the importance of mineral assemblages; cyclicity, as population growth cycles of microbes are a very important feature, originating from the fundamental behavior of microbial life, and if there is a mineralizing microbe type it will result in ore lamination on the micrometer scale. In the case of non-mineralizing types, the cyclic character will also occur, as during diagenesis the cell and EPS material will also mineralize;
- (3)
- Presence of organic matter embedded in minerals; even in the oxic Urucum Fe formation, there is variable embedded organic matter;
- (4)
- Biosignatures like isotope signals (“vital effect”) (C, S, N, Fe), shape of minerals, and selective enrichment of bioessential elements (Fe, Mn, Si, Zn, Co, As, Be, U, P, Ce, Mg, Ca, K, Na, S), the biological character of Mn and Fe and their atomic structural causes and consequences [46], and mechanisms in the background of microbial element enrichments [60];
- (5)
- Recent analogies of biomineralization and biomarker organic matter;
- (6)
- Paleoenvironmental analogies (sedimentary);
- (7)
- Preservation.We can add an 8th and 9th aspect based on our experience:
- (8)
- The extremely large horizontal and vertical extension and the giant mass in the case of sedimentary ore deposits (e.g., areal extension of the Urucum Fe-Mn deposit is around 800 km2, and its thickness is 400 m; it contains hundreds of millions of tonnes of ore);
- (9)
- The complexity of natural systems can also be a new aspect; natural systems are complex systems with variable microbial systems (e.g., dual Fe and Mn and also cyanobacterial), several mineral types, as direct microbial ones occur with accompanying mineral assemblage, all in a coherent way (texture, minerals, embedded organic matter, isotopic composition of given elements). Investigation of ore deposits aims at as many of the above aspects as possible.
3.7.2. Overview of the Recent State of the Art in Biogenic Thinking
3.7.3. Mechanisms in the Background of Element Enrichments
- (1)
- Direct oxidation of main ore-forming metals to sequestrate giant masses of biominerals (Fe-, Mn-oxidizing microbial activity); direct oxidation can include trace elements as well (e.g., Co and Ce are oxidized by Mn-oxidizing bacteria);
- (2)
- Structure (mineral) stabilizing role, such as Mg bound from seawater (sinking process) by EPS [34];
- (3)
- Adsorption, as in Ni adsorption from seawater or hydrothermal fluids (biogenic signature–decay of organic matter–plankton) by sheet MnOOH (birnessite), resulting in Ni–Mn(II)–sheet MnOOH (birnessite–buserite) [47]. In another example, it is common to find U, Th, Ra, and Rh adsorption on active Fe-biomats [83].
- (4)
- Detoxication, for instance Ce(III) to Ce(IV) by MnOB on MnOOH, which is a microbial oxidation path with P.
- (5)
- Vitamin demand, such as the need of Co for vitamin B12 [71].
- (6)
- Enzymatic demand, metalloproteins [84].
- (7)
- Protection against UV radiation and high Fe concentration (such as Si); FeOB consumes silica against stress, and it is also used for protection by microbes [58].
- (8)
3.7.4. Mineralogical Interpretation—Source of Elements
- (1)
- Abiogenic mixed syngenetic carbonate formation. Based on the geological environment, crystallization was syn-diagenetic and would not have caused significant changes in the carbonate compositions. Therefore, the water composition of the primary sedimentary system affects the Fe-Mn enrichment of the carbonates, depending on the particular metal-endowed fluid-rock system in which carbonate was precipitated or recrystallized.
- (2)
- Cyanobacterially mediated syngenetic calcite formation and Mn- and Fe-replacement via early diagenesis. Fe- and Mn diagenetic replacement of Ca in carbonates resulted in mixed forms of highly variable composition.
- (3)
- Diagenetic mixed carbonate formation via organic matter decomposition. The most likely formation of carbonates is diagenetic mixed carbonate formation via organic matter decomposition. This scenario is supported by mineral assemblages, micro-textural features, and negative δ13CPDB-carb values.
4. Methodology
- (1)
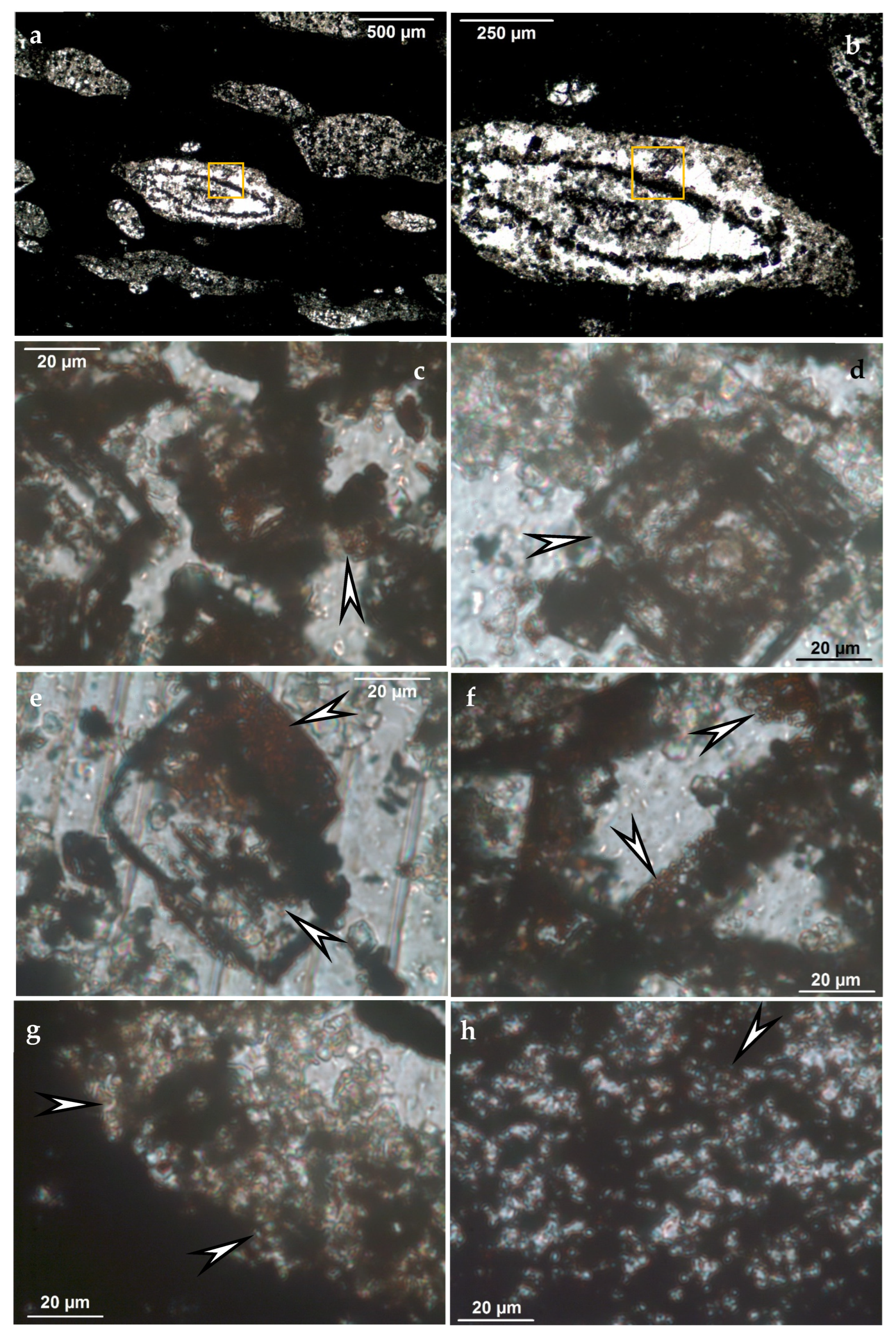
- The dimension of investigations must fall into the microbial size dimension (i.e., there is a need for high magnification microtextural observations as a first step to investigate the possible occurrence of microbial mediation). To solve this, high resolution in situ measurements and optical rock microscopy observations are needed.
- (2)
- Besides bulk analyses, in situ determination and distribution of mineral assemblage and embedded organic matter is needed to detect mineralized microbial cycles (mineralized biomats) and is also important to distinguish authigenic and allothigenic minerals (FTIR, Raman spectroscopy, cathodoluminescence microscopy).
- (3)
- If the possibility of microbial mediation in a formation is suspected (through microtextural evidence), the choice of appropriate methods is essential. For instance, XRD does not detect X-ray amorphous poorly crystallized microbially mediated minerals (e.g., ferrihydrite); furthermore, attention must be paid to levels of excitation energy, which is capable of converting minerals like ferrihydrite to more stable hematite. Thus, along with Raman spectroscopy, FTIR spectroscopy at lower excitation energy is suggested.
- (4)
- Debris character can be a "virtual" outlook, showing large minerals of authigenic processes and influencing textural features; this can be clarified easily using cathodoluminescence microscopy.
- (5)
- The authigenic formation of minerals that are well known as being of a particular origin may actually appear as authigenic materials. One example is magmatic or metamorphic conditions (quartz, feldspar—earlier known), but also pyroxene as aegirine, amphibol as riebeckite, and others, like dickite as high T and p minerals also common as low T authigenic minerals. Thus, a more detailed investigation of mineral type may be needed, and a complex approach is proposed to avoid a false interpretation.
- (6)
- Due to selective element enrichment (isotopes) resulting from microbial processes, we must use geochemical ratio methods with caution. This enrichment can indicate a microbially influenced element source, which often is not recognized as a possible interpretation. It also demonstrates the need for a multi-methodology approach and complex interpretation.
- (7)
- It is very important to distinguish whether clay-size dimensions or clay minerals occur. Certain data must be gathered: size dimension, the minerals in the fraction, and the origin of clay minerals (detrital, hydrothermal, or microbial). Often, microbial origin is indeed the case. This must be investigated in the case of marls or claystones (other clay-bearing rocks or ores), since the general assumption with clay-rich rocks is increased wet weathering and run off from terrestrial parts. In many cases, that may be a false interpretation.
- (8)
- It is extremely important to distinguish between syngenetic and diagenetic anoxia, since this determines original formation conditions. Environmental mineralogy based on mineral assemblage can provide clarification.
- (9)
- Mass balance considerations must be taken into consideration; the ratio of accumulating metal oxides, organic matter, and other components influences the diagenetic processes and the final character of the deposits (oxide, carbonate).
- (10)
- Identifying suboxic zones is important due to the consummation of organic matter by metal oxides and hydroxides, and the forming of diagenetic minerals.
Comparison of Observation by Optical Rock Microscopy and Electron Optical Methodology
5. Deposits and indications: Sample Investigations
6. Summary
- (1)
- The atomic scale features of elements, which determine microbial behavior and processes (e.g., Mn and Fe are “microbial” elements);
- (2)
- The connections between the Periodic Table and microbial life;
- (3)
- The background of selective microbial element enrichments;
- (4)
- Time duration differences on biological and geological scales;
- (5)
- Biomineralization, such as microbial metabolism, the metabolic types of given microbes (e.g., Fe-oxidizing microbes, Fe-reducing microbes), the role of prokaryots, autotrophy and heterotrophy;
- (6)
- Microbial systems in the frame of population growth cycles;
- (7)
- The main natural environments of microbial activity.
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Polgári, M.; Hein, J.R.; Vigh, T.; Szabó-Drubina, M.; Fórizs, I.; Bíró, L.; Müller, A.; Tóth, A.L. Microbial processes and the origin of the Úrkút manganese deposit, Hungary. Ore Geol. Rev. 2012, 47, 87–109. [Google Scholar] [CrossRef]
- Polgári, M.; Hein, J.R.; Tóth, A.L.; Pál-Molnár, E.; Vigh, T.; Bíró, L.; Fintor, K. Microbial action formed Jurassic Mn-carbonate ore deposit in only a few hundred years (Úrkút, Hungary). Geology 2012, 40, 903–906. [Google Scholar] [CrossRef]
- Gyollai, I.; Polgári, M.; Fintor, K.; Pál-Molnár, E.; Popp, F.; Koeberl, C. Microbial activity records in Marinoan Snowball Earth postglacial transition layers connecting diamictite with cap carbonate (Otavi Group, NW-Namibia). Austrian J. Earth Sci. 2017, 110, 2–18. [Google Scholar] [CrossRef]
- Rajabzadeh, M.A.; Haddad, F.; Polgári, M.; Fintor, K.; Walter, H.; Molnár, Z.; Gyollai, I. Investigation on the role of microorganisms in manganese mineralization from Abadeh-Tashk area, Fars Province, southwestern Iran by using petrographic and geochemical data. Ore Geol. Rev. 2017, 80, 229–249. [Google Scholar] [CrossRef]
- Yu, W.; Polgári, M.; Gyollai, I.; Fintor, K.; Szabó, M.; Kovács, I.; Fekete, J.; Du, Y.; Zhou, Q. Microbial metallogenesis of Cryogenian manganese ore deposits in South China. Precambrian Res. 2019, 322, 122–135. [Google Scholar] [CrossRef]
- Biondi, J.C.; Polgári, M.; Gyollai, I.; Fintor, K.; Kovács, I.; Fekete, J.; Mojzsis, S.J. Biogenesis of the Neoproterozoic kremydilite manganese ores from Urucum (Brazil)–A new manganese ore type. Precambrian Res. 2020, 340, 105624. [Google Scholar] [CrossRef]
- Gracheva, M.; Homonnay, Z.; Kovács, K.; Béres, K.A.; Biondi, J.C.; Wenchao, Y.; Kovács Kis, V.; Gyollai, I.; Polgári, M. Mössbauer characterization of microbially mediated iron and manganese ores of variable geological ages. Ore Geol. Rev. 2021, 134, 104124. [Google Scholar] [CrossRef]
- Karlik, M.; Gyollai, I.; Vancsik, A.; Fintor, K.; Szalai, Z.; Mindrescu, M.; Grădinaru, I.; Vágási, I.; Bozsó, G.; Pál-Molnár, E.; et al. High resolution mineralogical characterization of sediments-lake Bolatau-Feredeu (Romania). Carpathian J. Earth Environ. Sci. 2021, 16, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; Polgári, M.; Fintor, K.; Gyollai, I.; Szabó, M.; Velledits, F.; Liu, Z.; Du, Y. Contribution of microbial processes to the enrichment of Middle Permian manganese deposits in northern Guizhou, South China. Ore Geol. Rev. 2021, 136, 104259. [Google Scholar] [CrossRef]
- Yu, W.; Polgári, M.; Gyollai, I.; Fintor, K.; Huang, H.; Szabó, M.; Du, Y. Microbial metallogenesis of early carboniferous manganese deposit in central Guangxi, South China. Ore Geol. Rev. 2021, 136, 104251. [Google Scholar] [CrossRef]
- Polgári, M.; Biondi, J.C.; Gyollai, I.; Fintor, K.; Szabó, M. Origin of the Urucum iron formations (Neoproterozoic, Brazil): Textural and mineralogical evidence (Mato Grosso do Sul–Brazil). Ore Geol. Rev. 2021, 139, 104456. [Google Scholar] [CrossRef]
- Polgári, M.; Gyollai, I.; Fintor, K.; Horváth, H.; Pál-Molnár, E.; Biondi, J.C. Microbially mediated ore-forming processes and cell mineralization. Front. Microbiol. 2019, 10, 2731. [Google Scholar] [CrossRef]
- Varga, G. Kálitrachit és káliumdús kőzetek a Mátrában (Potassium trachyte and potassium rich rocks in the Mátra Mts.). Annu. Rep. Hung. Geol. Inst. 1990, 1992, 241–276. [Google Scholar]
- Nagy, B. K-rich rocks and their relation to mineralization in the Mátra Mountains (North Hungary). Acta Geol. Hung. 2006, 49, 33–41. [Google Scholar] [CrossRef]
- Polgári, M.; Nagy, B.; Fintor, K.; Gyollai, I.; Kovács, I.; Szabó, M.; Mojzsis, S. Contribution to the origin of K-rich rocks in the Mátra Mountains (North Hungary) 2022. (Research Report, National Scientific Research No. 125060, manuscript in Hungarian).
- Roy, S. Manganese Deposits; Academic Press: London, UK, 1981; p. 458. [Google Scholar]
- Polgári, M.; Hein, J.R.; Németh, T.; Pál-Molnár, E.; Vigh, T. Celadonite and smectite formation in the Úrkút Mn-carbonate ore deposit (Hungary). Sediment. Geol. 2013, 294, 157–163. [Google Scholar] [CrossRef]
- Polgári, M.; Hein, J.R.; Bíró, L.; Gyollai, I.; Németh, T.; Sajgó, C.; Fekete, J.; Schwark, L.; Pál-Molnár, E.; Vigh, T.; et al. Mineral and chemostratigraphy of a Toarcian black shale hosting Mn-carbonate microbialites (Úrkút, Hungary). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2016, 459, 99–120. [Google Scholar] [CrossRef]
- Polgári, M.; Németh, T.; Pál-Molnár, E.; Futó, I.; Vigh, T.; Mojzsis, S.J. Correlated chemostratigraphy of Mn-carbonate microbialites (Úrkút, Hungary). Gondwana Res. 2016, 29, 278–289. [Google Scholar] [CrossRef]
- Berner, R.A. Early Diagenesis: A Theoretical Approach; Princeton University Press: Princeton, NJ, USA, 1980; p. 250. [Google Scholar]
- Coleman, M.L. Geochemistry of diagenetic non-silicate minerals: Kinetic considerations. Philos. Trans. R. Soc. Lond. Ser. A Math. Phys. Sci. 1985, 315, 39–56. [Google Scholar]
- Listova, L.P. Experimental Studies of Physic-Chemical Conditions of Sedimentation of Manganese Oxides and Carbonate. Ocherki Metallogen. Osadochn. Porod.; Publication of the AN USSR: Moscow, Russia, 1961; pp. 319–351. [Google Scholar]
- Harder, H. Nontronite synthesis at low temperatures. Chem. Geol. 1976, 18, 169–180. [Google Scholar] [CrossRef]
- Harder, H. Synthesis of iron layer silicate minerals under natural conditions. Clays Clay Miner. 1978, 26, 65–72. [Google Scholar] [CrossRef]
- Trudinger, P.A.; Swaine, D.J. Biogeochemical Cycling of Mineral-Forming Elements; Elsevier: Amsterdam, The Netherlands, 1979; p. 608. [Google Scholar]
- Giovanoli, R. On natural and synthetic manganese nodules. In Geology and Geochemistry of Manganese, 1st ed.; Varentsov, I.M., Grasselly, G., Eds.; Akadémiai Publishing House: Budapest, Hungary, 1980; pp. 159–203. [Google Scholar]
- Sung, W.; Morgan, J.J. Oxidative removal of Mn (II) from solution catalysed by the γ-FeOOH (lepidocrocite) surface. Geochim. Cosmochim. Acta 1981, 45, 2377–2383. [Google Scholar] [CrossRef]
- Cole, T.G.; Shaw, H.F. The nature and origin of authigenic smectites in some recent marine sediments. Clay Miner. 1983, 18, 239–252. [Google Scholar] [CrossRef]
- Ewers, W.E. Chemical Factors in the Deposition and Diagenesis of Banded Iron-Formation. In Developments in Precambrian Geology 6, Iron Formation: Facts and Problems; Trendall, A.F., Morris, R.C., Eds.; Elsevier: Amsterdam, The Netherlands, 1983; pp. 491–512. [Google Scholar]
- Maynard, J.B. Geochemistry of Sedimentary Ore Deposits; Springer: New York, NY, USA, 1983. [Google Scholar]
- Skinner, H.C.W. A review of apatites, iron and manganese minerals and their roles as indicators of biological activity in black shales. Precambrian Res. 1993, 61, 209–229. [Google Scholar] [CrossRef]
- Ehrenreich, A.; Widdel, F. Anaerobic oxidation of ferrous iron by purple bacteria, a new type of phototrophic metabolism. Appl. Environ. Microbiol. 1994, 60, 4517–4526. [Google Scholar] [CrossRef]
- Wignall, P.B. Black Shales; Oxford Clarendon Press: Oxford, UK, 1994; p. 124. [Google Scholar]
- Mandernack, K.W.; Post, J.; Tebo, B.M. Manganese mineral formation by bacterial spores of the marine Bacillus, strain SG-1: Evidence for the direct oxidation of Mn (II) to Mn (IV). Geochim. Cosmochim. Acta 1995, 59, 4393–4408. [Google Scholar] [CrossRef]
- Straub, K.L.; Benz, M.; Schink, B.; Widdel, F. Anaerobic, nitrate-dependent microbial oxidation of ferrous iron. Appl. Environ. Microbiol. 1996, 62, 1458–1460. [Google Scholar] [CrossRef]
- Banfield, J.F.; Nealson, K.H. Geomicrobiology: Interactions between Microbes and Minerals. In Reviews in Mineralogy 35; Mineralogical Society of America: Washington, DC, USA, 1997; p. 448. [Google Scholar]
- Konhauser, K.O. Diversity of bacterial iron mineralization. Earth Sci. Rev. 1998, 43, 91–121. [Google Scholar] [CrossRef]
- Herdianita, N.R.; Browne, P.R.L.; Rodgers, K.A.; Campbell, K.A. Mineralogical and textural changes accompanying ageing of silica sinter. Miner. Depos. 2000, 35, 48–62. [Google Scholar] [CrossRef]
- Chaudhuri, S.K.; Lack, J.G.; Coates, J.D. Biogenic magnetite formation through anaerobic biooxidation of Fe (II). Appl. Environ. Microbiol. 2001, 67, 2844–2848. [Google Scholar] [CrossRef]
- Ehrlich, H.L. Geomicrobiology, 4th ed.; Marcell Dekker Inc.: New York, NY, USA, 2002; pp. 183–274. [Google Scholar]
- Bazylinski, D.A.; Frankel, R.B. Biologically controlled mineralization in prokaryotes. Rev. Mineral. Geochem. 2003, 54, 217–247. [Google Scholar] [CrossRef]
- Lee, S.H.; Lee, I.; Roh, Y. Biomineralization of a poorly crystalline Fe (III) oxide, akaganeite, by an anaerobic Fe (III)-reducing bacterium (Shewanella alga) isolated from marine environment. Geosci. J. 2003, 7, 217–226. [Google Scholar] [CrossRef]
- Villalobos, M.; Toner, B.; Bargar, J.; Sposito, G. Characterization of the manganese oxide produced by Pseudomonas putida strain MnB1. Geochim. Cosmochim. Acta 2003, 67, 2649–2662. [Google Scholar] [CrossRef]
- Bargar, J.R.; Tebo, B.M.; Bergmann, U.; Webb, S.M.; Glatzel, P.; Chiu, V.Q.; Villalobos, M. Biotic and abiotic products of Mn (II) oxidation by spores of the marine Bacillus sp. strain SG-1. Am. Mineral. 2005, 90, 143–154. [Google Scholar] [CrossRef]
- Dupraz, C.; Visscher, P.T. Microbial lithification in marine stromatolites and hypersaline mats. Trends Microbiol. 2005, 13, 429–438. [Google Scholar] [CrossRef] [PubMed]
- Morgan, J.J. Kinetics of reaction between O2 and Mn (II) species in aqueous solutions. Geochim. Cosmochim. Acta 2005, 69, 35–48. [Google Scholar] [CrossRef]
- Bodeï, S.; Manceau, A.; Geoffroy, N.; Baronnet, A.; Buatier, M. Formation of todorokite from vernadite in Ni-rich hemipelagic sediments. Geochim. Cosmochim. Acta 2007, 71, 5698–5716. [Google Scholar] [CrossRef]
- Lemos, V.P.; Lima da Costa, M.; Lemos, R.L. Vivianite and siderite in lateritic iron crust: An example of bioreduction. Quim. Nova 2007, 30, 36–40. [Google Scholar] [CrossRef]
- Schwertmann, U.; Cornell, R.M. Iron oxides in the laboratory: Preparation and characterization; John Wiley & Sons: Hoboken, NJ, USA, 2007; p. 188. [Google Scholar]
- Dupraz, C.; Reid, R.P.; Braissant, O.; Decho, A.W.; Norman, R.S.; Visscher, P.T. Processes of carbonate precipitation in modern microbial mats. Earth Sci. Rev. 2009, 96, 141–162. [Google Scholar] [CrossRef]
- Chan, C.S.; Fakra, S.C.; Emerson, D.; Fleming, E.J.; Edwards, K.J. Lithotrophic iron oxidizing bacteria produce organic stalks to control mineral growth: Implications for biosignature formation. Multidiscip. J. Int. Soc. Microb. Ecol. 2011, 5, 717–727. [Google Scholar] [CrossRef]
- Piepenbrock, A.; Dippon, U.; Porsch, K.; Appel, E.; Kappler, A. Dependence of microbial magnetite formation on humic substance and ferrihydrite concentrations. Geochim. Cosmochim. Acta 2011, 75, 6844–6858. [Google Scholar] [CrossRef]
- Biagioni, C.; Capalbo, C.; Lezzerini, M.; Pasero, M. Ferrihollandite, BaMn4+ 6Fe3+ 2O16, from Apuan Alps, Tuscany, Italy: Description and crystal structure. Eur. J. Mineral. 2014, 26, 171–178. [Google Scholar] [CrossRef]
- Zeyen, N.; Benzerara, K.; Li, J.; Groleau, A.; Balan, E.; Robert, J.-L.; Estève, I.; Tavera, R.; Moreira, D.; López-García, P. Formation of low-T hydrated silicates in modern microbialites from Mexico and implications for microbial fossilization. Front. Earth Sci. 2015, 3, 64. [Google Scholar] [CrossRef]
- Liu, D.; Zhang, Q.; Wu, L.; Zeng, Q.; Dong, H.; Bishop, M.E.; Wang, H. Humic acid-enhanced illite and talc formation associated with microbial reduction of Fe(III) in nontronite. Chem. Geol. 2016, 447, 199–207. [Google Scholar] [CrossRef]
- Johnson, J.E.; Webb, S.M.; Ma, C.; Fischer, W.W. Manganese mineralogy and diagenesis in the sedimentary rock record. Geochim. Cosmochim. Acta 2016, 173, 210–231. [Google Scholar] [CrossRef]
- Gyollai, I.; Polgári, M.P.; Fintor, K.; Popp, F.; Mader, D.; Pál-Molnár, E.; Nagy, S.; Koeberl, C. Microbially mediated deposition of postglacial transition layers from the Neoproterozoic Otavi Group, Namibia: Evidence of rapid deglaciation after the Sturtian cryogenic period. Carpathian J. Earth Environ. Sci. 2015, 10, 63–76. [Google Scholar]
- Młoszewska, A.M.; Cole, D.B.; Planavsky, N.J.; Kappler, A.; Whitford, D.S.; Owttrim, G.W.; Konhauser, K. UV radiation limited the expansion of cyanobacteria in early marine photic environments. Nat. Commun. 2018, 9, 1–8. [Google Scholar] [CrossRef]
- Sanz Montero, M.E.; Rodríguez Aranda, J.P. Magnesite formation by microbial activity: Evidence from a Miocene hypersaline lake. Sediment. Geol. 2020, 12, 51. [Google Scholar] [CrossRef]
- Polgári, M.; Gyollai, I. Geochemical constraints on the element enrichments of microbially mediated manganese and iron ores–An overview. Ore Geol. Rev. 2021, 136, 104203. [Google Scholar] [CrossRef]
- Fortin, D.; Ferris, F.G.; Beveridge, T.J. Surface-mediated mineral development by bacteria. In Geomicrobiology: Interactions between Microbes and Minerals; Banfield, J., Nealson, K.H., Eds.; Mineralogical Society of America: Chantilly, VA, USA, 1997; Volume 35, pp. 162–180. [Google Scholar]
- Polgári, M.; Gyollai, I.; Bérczi, S. Terraforming on Early Mars. In Terraforming Mars; Beech, M., Seckbach, J., Gordon, R., Eds.; Wiley: Hoboken, NJ, USA, 2022; pp. 161–279. [Google Scholar]
- Madondo, J.; Polgári, M.; Gyollai, I.; Fintor, K.; Kovács, I.; Szabó, M. Contribution to the origin of Molango manganese deposit, Mexico. 2022. (Research Report, National Scientific Research No. 125060, manuscript in Hungarian).
- Dong, Z.; Polgári, M.; Gyollai, I.; Fintor, K.; Kovács, I.; Szabó, M. Contribution to the origin of manganese deposit at Kunlun Mts., China 2022. (Research Report, National Scientific Research No. 125060, manuscript in Hungarian).
- Dong, Z.; Polgári, M.; Gyollai, I.; Fintor, K.; Kovács, I.; Szabó, M. Contribution to the origin of manganese deposit at Tianshan Mts., China 2022. (Research Report, National Scientific Research No. 125060, manuscript in Hungarian).
- Tosca, N.J.; Macdonald, F.A.; Strauss, J.V.; Johnston, D.T.; Knoll, A.H. Sedimentary talc in Neoproterozoic carbonate successions. Earth Planet. Sci. Lett. 2011, 306, 11–22. [Google Scholar] [CrossRef]
- Bodor, S.; Polgári, M.; Szentpétery, I.; Földessy, J. Microbially mediated iron ore formation, Silicic Superunit, Rudabánya, Hungary. Ore Geol. Rev. 2016, 72, 391–401. [Google Scholar] [CrossRef]
- Rosing, M.T.; Rose, N.M.; Bridgwater, D.; Thomsen, H.S. Earliest part of Earth’s stratigraphic record: A reappraisal of the >3.7 Ga Isua (Greenland) supracrustal sequence. Geology 1996, 24, 43–46. [Google Scholar] [CrossRef]
- Katsuta, N.; Shimizu, I.; Helmstaedt, H.; Takano, M.; Kawakami, S.; Kumazawa, M. Major element distribution in Archean banded iron formation (BIF): Influence of metamorphic differentiation. J. Metamorph. Geol. 2012, 30, 457–472. [Google Scholar] [CrossRef]
- Gutzmer, J.; Beukes, N.J. Origin and paleoenvironmental significance of major iron formations at the Archean-Paleoproterozoic boundary. In Banded Iron Formation-Related High-Grade Iron Ore.—Reviews in Economic Geology; Hagemann, S., Rosière, C.A., Gutzmer, J., Beukes, N.J., Eds.; Society of Economic Geologists (SEG): Littleton, CO, USA, 2008; Volume 15. [Google Scholar] [CrossRef]
- Knoll, A.H.; Canfield, D.E.; Konhauser, K.O. Fundamentals of Geobiology; Wiley-Blackwell: Oxford, UK, 2012; p. 456. [Google Scholar]
- Webb, S.; Dick, G.J.; Bargar, J.R.; Tebo, B.M. Evidence for the presence of Mn (III) intermediates in the bacterial oxidation of Mn(II). Microbiology 2005, 102, 5558–5563. [Google Scholar] [CrossRef] [PubMed]
- Mandernack, K.W.; Tebo, B.M. Manganese scavenging and oxidation at hydrothermal vents and in vent plumes. Geochim. Cosmochim. Acta 1993, 57, 3907–3923. [Google Scholar] [CrossRef]
- Maynard, J.B. Manganiferous sediments, rocks, and ores. In Treatise on Geochemistry; Elsevier: Amsterdam, The Netherlands, 2014; Volume 7, pp. 327–349, 407. [Google Scholar]
- Konhauser, K. Biomineralization. In Introduction to Geomicrobiology; Kohnhauser, K., Ed.; Blackwell Publishing: Oxford, UK, 2007; pp. 139–191. [Google Scholar]
- Garrels, R.M.; MacKenzie, F.T. Evolution of Sedimentary Rocks; Nortom: New York, NY, USA, 1971. [Google Scholar]
- Hallbeck, L.; Pedersen, K. Culture parameters regulating stalk formation and growth rate of Gallionella ferruginea. Microbiology 1990, 136, 1675–1680. [Google Scholar] [CrossRef]
- Emerson, D.; Fleming, E.J.; McBeth, J.M. Iron-oxidizing bacteria: An environmental and genomic perspective. Annu. Rev. Microbiol. 2010, 64, 561–583. [Google Scholar] [CrossRef]
- Tyson, R.V.; Pearson, T.H. Modern and ancient continental shelf anoxia: An overview. Geol. Soc. Lond. Spec. Publ. 1991, 58, 1–24. [Google Scholar] [CrossRef]
- Noffke, N.; Hazen, R.; Nhleko, N. Earth’s earliest microbial mats in a siliciclastic marine environment (2.9 Ga Mozaan Group, South Africa). Geology 2003, 31, 673–676. [Google Scholar] [CrossRef]
- Noffke, N.; Eriksson, K.A.; Hazen, R.M.; Simpson, E.L. A new window into Early Archean life: Microbial mats in Earth’s oldest siliciclastic tidal deposits (3.2 Ga Moodies Group, South Africa). Geology 2006, 34, 253–256. [Google Scholar] [CrossRef]
- Cady, S.L.; Farmer, J.D.; Grotzinger, J.P.; Schopf, J.W.; Steele, A. Morphological Biosignatures and the Search for Life on Mars. Astrobiology 2003, 3, 351–369. [Google Scholar] [CrossRef] [PubMed]
- Tsezos, M.; Baird, M.H.I.; Shemilt, L.W. The elution of radium adsorbed by microbial mats. Chem. Eng. J. 1987, 34, B57–B64. [Google Scholar] [CrossRef]
- Dupont, C.L.; Butcher, A.; Ruben, E.; Valas, R.E.; Bourne, P.E.; Caetano-Anollés, G. History of biological metal utilization inferred through phylogenomic analysis of protein structures. Proc. Natl. Acad. Sci. USA 2010, 107, 10567–10572. [Google Scholar] [CrossRef] [PubMed]
- Glamoclija, M.; Steele, A.; Fries, M.; Schieber, J.; Voytek, M.; Cockell, C. Association of anatase (TiO2) and microbes: Unusual fossilization effect or a potential biosignature? Geol. Soc. Am. Spec. Pap. 2009, 458, 965–975. [Google Scholar]
- Burke, I.T.; Kemp, A.E.S. Microfabric analysis of Mn-carbonate laminae deposition and Mn-sulfide formation in the Gotland Deep, Baltic Sea. Geochim. Cosmochim. Acta 2002, 66, 1589–1600. [Google Scholar] [CrossRef]
- Murowchick, J.B.; Barnes, H.L. Marcasite precipitation from hydrothermal solutions. Geochim. Cosmochim. Acta 1986, 50, 2615–2629. [Google Scholar] [CrossRef]
- Schoonen, M.A.A.; Barnes, H.L. Reactions forming pyrite and marcasite from solution: I. Nucleation of FeS2 below 100 °C. Geochim. Cosmochim. Acta 1991, 55, 1495–1504. [Google Scholar] [CrossRef]
- Egger, M.; Hagens, M.; Sapart, C.J.; Dijkstra, N.; van Helmond, N.A.G.M.; Mogoll´on, J.M.; Risgaard-Petersen, N.; van der Veen, C.; Kasten, S.; Riedinger, N.; et al. Iron oxide reduction in methane-rich deep Baltic Sea sediments. Geochim. Cosmochim. Acta 2017, 207, 256–276. [Google Scholar] [CrossRef]
- Baele, J.M.; Bouvain, F.; De Jong, J.; Matielli, N.; Papier, S.; Préat, A. Iron microbial mats in Modern and Phanerozoic environments. In Instruments, Methods, and Missions for Astrobiology XI; International Society for Optics and Photonics: Washington, DC, USA, 2008; Volume 7097, p. 70970N12. [Google Scholar]
- Müller, A. Morphology and genesis of chalcedony and opal in S-Mátra Mts. Dél-mátrai kalcedon és opál mintázatok morfológiája és genetikája. 2009. Research Report, National Scientific Research No. 68992, manuscript in Hungarian.
- Angerer, T.; Hagemann, S.G.; Walde, D.H.; Halverson, G.P.; Boyce, A.J. Multiple metal sources in the glaciomarine facies of the Neoproterozoic Jacadigo iron formation in the “Santa Cruz deposit”, Corumbá, Brazil. Precambrian Res. 2016, 275, 369–393. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Minerals/ Processes | Chemical Formula | Locality of Deposits | Occurrence of Minerals in the Deposits x/9 | Eh | pH | Microbially | S | Mn | Fe | D | Mn ** | Fe | C | EPS | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mn mineral assemblage | oxic | suboxic | anoxic | acidic | neutral-slightly alkaline | alkaline | mediated | |||||||||||
| Oxides and hydroxides | ||||||||||||||||||
| Pyrolusite | Mn4+O2 | 1,2,4,6,7,9 | 6 | * | * | * | ||||||||||||
| Ramsdellite | Mn4+O2 | 1,4,7 | 3 | * | * | * | ||||||||||||
| Nsutite | (Mn4+Mn2+)(O,OH)2 | 1 | 1 | * | * | * | * | |||||||||||
| Hausmannite | Mn3+3O4 | 1 | 1 | * | * | * | ||||||||||||
| Cryptomelane | KMn4+6Mn2+2O16 | 1,4,6,9 | 4 | * | * | (*) | ||||||||||||
| Jacobsite | Mn2+0.6Fe2+0.3Mg0.1Fe3+1.5Mn3+0.5O4 | 1,2,4,5,7 | 5 | * | * | (*) | * | * | ||||||||||
| Manganite | Mn3+OOH | 1,2,3,4,6,7,9 | 7 | * | * | * | * | |||||||||||
| Vernadite δMnO2 (wad) | (Mn4+Fe3+CaNa)(OOH)2·nH2O | 1,9 | 2 | * | * | * | * | |||||||||||
| Todorokite | Na0.2Ca0.05K0.02Mn4+4Mn3+2O12•3(H2O) | 1,2,4,7,8,9 | 6 | * | * | * | * | * | ||||||||||
| Birnessite | Na0.7Ca0.3(Mn3+ Mn4+)7O14·2.8H2O | 1,8 | 2 | * | * | * | * | * | ||||||||||
| Romanèchite (psilomelane) | [(Ba,H2O,Mn5O10, Ba(Mn4+, Mn3+)O10·1.4H2O)] | 1,4,5,6,7 | 5 | * | * | (*) | ||||||||||||
| Hollandite* | Ba(Mn4+,Mn2+)8O16 | 1,4,9 | 3 | * | * | (*) | ||||||||||||
| Manjiroite | Na(Mn4+7Mn3+)O16 | 1,2,4,5,6,7,8 | 7 | * | * | (*) | ||||||||||||
| Pyrophanite | MnTiO3 | 4,5,6,7,8 | 5 | * | ||||||||||||||
| Arsenate Geigerite | Mn5(AsO3OH)2(AsO4)2·10H2O | 5 | 1 | |||||||||||||||
| Carbonates | ||||||||||||||||||
| Rhodochrosite | MnCO3 | 1,2,3,4,5,6,7,8,9 | 9 | * | * | * | * | (*) | ||||||||||
| Mn-Calcite | Mn-CaCO3 | 2,4,7 | 3 | * | ||||||||||||||
| Kutnohorite | (Ca,Mn)(CO3)2 | 2,3,4,5,6,7,8,9 | 8 | * | * | (*) | ||||||||||||
| Oxides-silicates | ||||||||||||||||||
| Serandite (pyroxmangite/rhodonite) | NaMn2+1.5Ca0.5Si3O8(OH) | 1,4,5,7 | 4 | * | * | |||||||||||||
| Braunite | Mn2+Mn3+6SiO12 | 1,2,4,5,6,7,8 | 7 | * | * | * | * | * | ||||||||||
| Sulfides | ||||||||||||||||||
| Alabandite | MnS | 2,7 | 2 | * | * | * | * | |||||||||||
| Rambergite | MnS | 4,5,6,7,8 | 5 | * | * | |||||||||||||
| Fe mineral assemblage | ||||||||||||||||||
| Oxides and hydroxides | ||||||||||||||||||
| Ferrihydrite | FeOOH | 1,2,3,4,5,6,7,8 | 8 | * | * | * | * | * | ||||||||||
| Lepidocrocite | Fe3+O(OH) | 1,3,4,7,8 | 5 | * | * | * | * | * | ||||||||||
| Hematite | Fe2O3 | 1,2,3,4,5,6,7,8,9 | 9 | * | * | * | * | * | * | |||||||||
| Goethite | FeOOH | 1,2,4,5,6,7,8,9 | 8 | * | * | * | * | * | * | |||||||||
| Magnetite | Fe2O3 | 1,5,6,7,8,9 | 6 | * | * | * | * | * | ||||||||||
| Anatase | TiO2 − FexTi(1 − x)O(2 − x)OHx | 2,4,5,6,7,8 | 6 | * | * | * | * | * | ||||||||||
| Ilmenite | FeTiO3 | 8 | 1 | |||||||||||||||
| Brookite | TiO2 | 5,6 | 2 | |||||||||||||||
| Carbonates | ||||||||||||||||||
| Siderite | FeCO3 | 2,3,4,5,6,7,8,9 | 8 | * | * | * | * | * | ||||||||||
| Ankerite | Ca(Fe2+,Mg)(CO3)2 | 1,2,3,4,6,7,8 | 7 | * | * | * | * | |||||||||||
| Sulfides | ||||||||||||||||||
| Pyrite | FeS2 | 1,3,4,5,6,7,8,9 | 8 | * | * | * | * | * | ||||||||||
| Marcasite | FeS2 | 2,6,7,8,9 | 5 | * | * | * | * | |||||||||||
| Sulphates | ||||||||||||||||||
| Na-jarosite | NaFe3+3(SO4)2(OH)6 | 4,6,7,8 | 4 | * | * | * | * | (*) | ||||||||||
| Silicates | ||||||||||||||||||
| Aegirine | Ca0.75Na0.25Mg0.5Fe2+0.25Fe3+0.25(Si2O6) | 1,2,7 | 3 | * | * | * | (*) | |||||||||||
| Riebeckite | Na2(Fe2+3Fe3+2)Si8O22(OH)2 | 1 | 1 | * | (*) | |||||||||||||
| Celadonite | KMg0.8Fe2+0.2Fe3+0.9Al0.1Si4O10(OH)2 | 1,5,6,7,8,9 | 6 | * | * | * | * | (*) | ||||||||||
| Nontronite | (Ca0.5Na)0.7Fe3+4[(Si7.3Al0.7)O20(OH)4]·nH2O | 6,7,8,9 | 4 | * | * | * | * | (*) | ||||||||||
| Chlorite | Mg3.75Fe2+1.25Si3Al2O10(OH)8 | 1,9 | 2 | * | (*) | |||||||||||||
| Chamosite | (Fe2+Mg)5Al(AlSi3O10)(OH)8 | 1 | 1 | * | * | (*) | ||||||||||||
| Phlogopite | KMg3AlSi3O10(F,OH)2 (Fe substitution, Fe-mica) | 6,8 | 2 | |||||||||||||||
| Chlinochlore | Mg5Al(AlSi3O10)(OH)8 (Fe2+) | 8 | 1 | |||||||||||||||
| Others | ||||||||||||||||||
| Oxides-hydroxides | ||||||||||||||||||
| Quartz | SiO2 | 1,2,3,4,5,6,7,8,9 | 9 | * | * | * | * | |||||||||||
| Rutile | TiO2 | 8 | 1 | |||||||||||||||
| Corundum | Al2O3 | 8 | 1 | |||||||||||||||
| Carbonates | ||||||||||||||||||
| Calcite | CaCO3 | 5,6,8 | 3 | |||||||||||||||
| Magnesite | MgCO3 | 8 | 1 | * | * | * | * | |||||||||||
| Dolomite | CaMg(CO3)2 | 1,2,3,4,6,7,8 | 7 | * | * | * | * | * | ||||||||||
| Strontianite | Sr(CO3) | 1,8 | 2 | * | * | * | * | |||||||||||
| Witherite | BaCO3 | 2 | 1 | * | * | |||||||||||||
| Silicates | ||||||||||||||||||
| Orthoclase feldspar | KAlSi3O8 | 1,3 | 2 | * | * | * | ||||||||||||
| Plagioclase | NaAlSi3O8 | 2 | 1 | * | * | |||||||||||||
| Albite feldspar | NaAlSi3O8 | 1,2,4,5,6,7,8,9 | 8 | * | * | * | ||||||||||||
| Kaolinite/dickite | Al2Si2O5(OH)4 | 1,2,5,6,7,8 | 6 | ? | ? | * | * | * | ||||||||||
| Muscovite/mica | KAl3Si3O10(OH)1.8F0.2 | 1,2,5,6,7,8 | 6 | * | * | |||||||||||||
| Montmorillonite | (Na,Ca)(Al,Mg)2Si4O10(OH)2·nH2O | 2,4,6,7,8,9 | 6 | * | * | * | * | |||||||||||
| Illite | (K,H3O)(Al,Mg,Fe)2(Si,Al)4O10[(OH)2,(H2O) | 3 | 1 | * | * | |||||||||||||
| Talk | Mg3Si4O10(OH)2 | 5,6 | 2 | * | * | * | ||||||||||||
| Clay mineral | 8 | 1 | ||||||||||||||||
| Pyroxene | 8 | 1 | ||||||||||||||||
| Zircon | ZrSiO4 | 2 | 1 | |||||||||||||||
| Zeolite | 9 | 1 | * | * | ||||||||||||||
| Silicate-carbonate-sulphate | ||||||||||||||||||
| Cancrinite | (Na,Ca)(Al6Si6O24)(CO3,SO4)2·2H2O | 1 | 1 | * | * | |||||||||||||
| Phosphates | ||||||||||||||||||
| Apatite | [(Ca10(PO4)6(OH,F,Cl)2] | 1,2,3,4,5,6,7,9 | 8 | * | * | * | * | * | * | |||||||||
| Xenotime | YPO4 | 2 | 1 | |||||||||||||||
| Monazite | (Ce,La,Th)PO4 | 5 | 1 | |||||||||||||||
| Sulfides | ||||||||||||||||||
| Sphalerite | ZnS | 5,8 | 2 | |||||||||||||||
| Galenite | PbS | 5 | 1 | |||||||||||||||
| Chalchopyrite | CuFeS2 | 5,8 | 2 | |||||||||||||||
| Cattierite | CoS2 | 8 | 1 | |||||||||||||||
| Orphiment | As2S | 8 | 1 | |||||||||||||||
| Sulphates | ||||||||||||||||||
| Barite | Ba(SO4) | 1,4,6,7,9 | 5 | * | * | * | * | * | ||||||||||
| Gypsum (anhidrite) | CaSO4·2H2O | 1,9 | 2 | * | * | * | * | * | ||||||||||
| Coelestite | SrSO4 | 2 | 1 | |||||||||||||||
| Johannite | Cu(UO2)2(SO4)2(OH)2 8H2O | 1,4,5,6,7 | 5 | * | * | |||||||||||||
| Organic material | * | |||||||||||||||||
| Environmental Oxygen Levels | Eh (V) | Dissolved Oxygen (DO) | FeOB Demand * | MnOB Demand |
|---|---|---|---|---|
| obligatory oxic | 0.4 < Eh < 1.0 | DO > 2 mL/L | x | |
| dysoxic | 0.2–0.4 | DO 0.2–2.0 mL/L | ≈ 0.3 | |
| suboxic | 0–0.2 | DO 0–0.2 mL/L | ||
| anoxic | < 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Polgári, M.; Gyollai, I. Comparative Study of Formation Conditions of Fe-Mn Ore Microbialites Based on Mineral Assemblages: A Critical Self-Overview. Minerals 2022, 12, 1273. https://doi.org/10.3390/min12101273
Polgári M, Gyollai I. Comparative Study of Formation Conditions of Fe-Mn Ore Microbialites Based on Mineral Assemblages: A Critical Self-Overview. Minerals. 2022; 12(10):1273. https://doi.org/10.3390/min12101273
Chicago/Turabian StylePolgári, Márta, and Ildikó Gyollai. 2022. "Comparative Study of Formation Conditions of Fe-Mn Ore Microbialites Based on Mineral Assemblages: A Critical Self-Overview" Minerals 12, no. 10: 1273. https://doi.org/10.3390/min12101273
APA StylePolgári, M., & Gyollai, I. (2022). Comparative Study of Formation Conditions of Fe-Mn Ore Microbialites Based on Mineral Assemblages: A Critical Self-Overview. Minerals, 12(10), 1273. https://doi.org/10.3390/min12101273