
Fluctuating Asymmetry as a Measure of Stress in Natural Populations of Woody Plants: Influence of Ecological and Geographical Factors on Developmental Stability



Abstract
1. Introduction
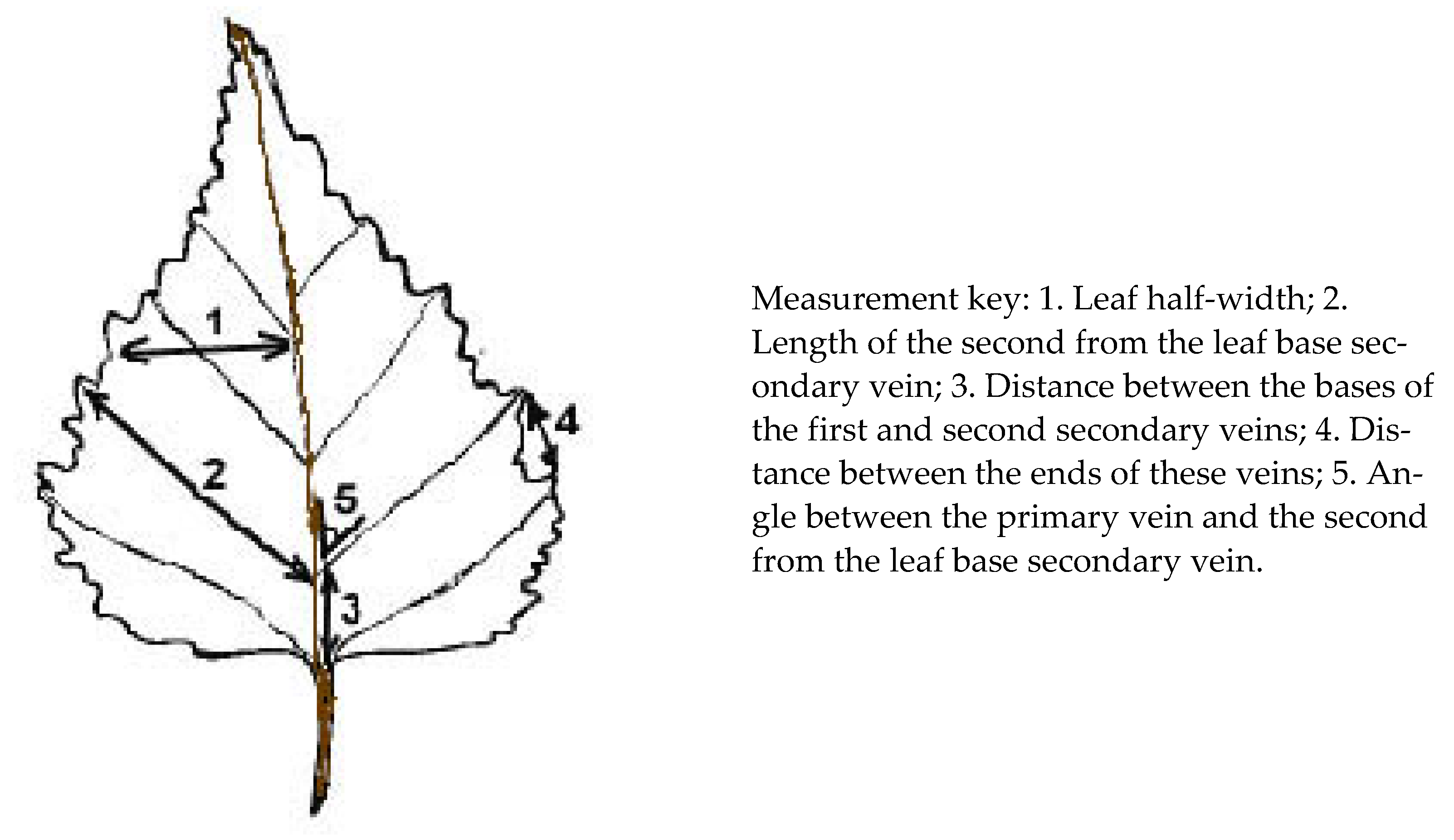
2. Materials and Methods
3. Results and Discussion
3.1. Abiotic Conditions of the Studied Regions
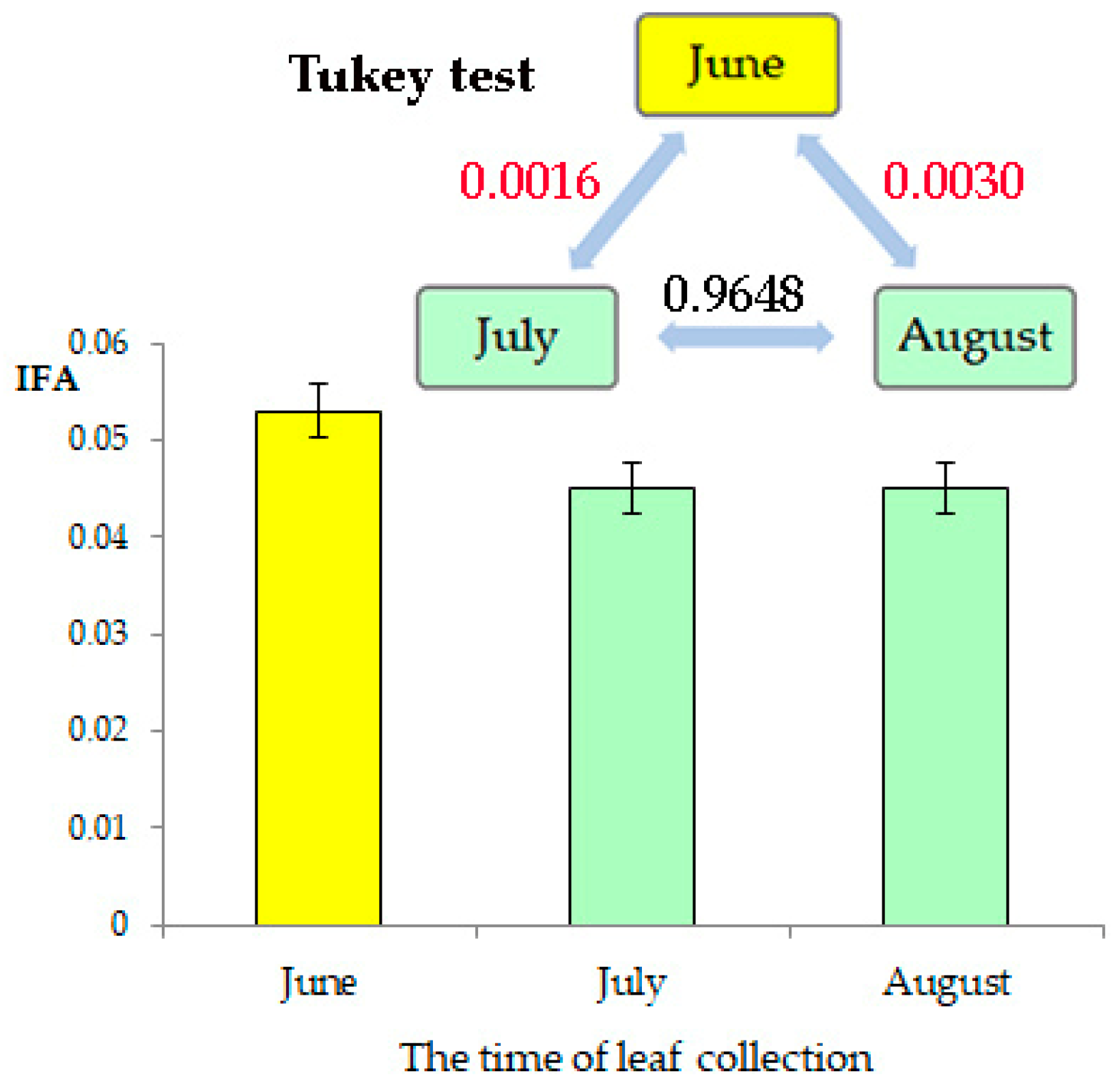
3.2. Intraindividual and Chronological Variability
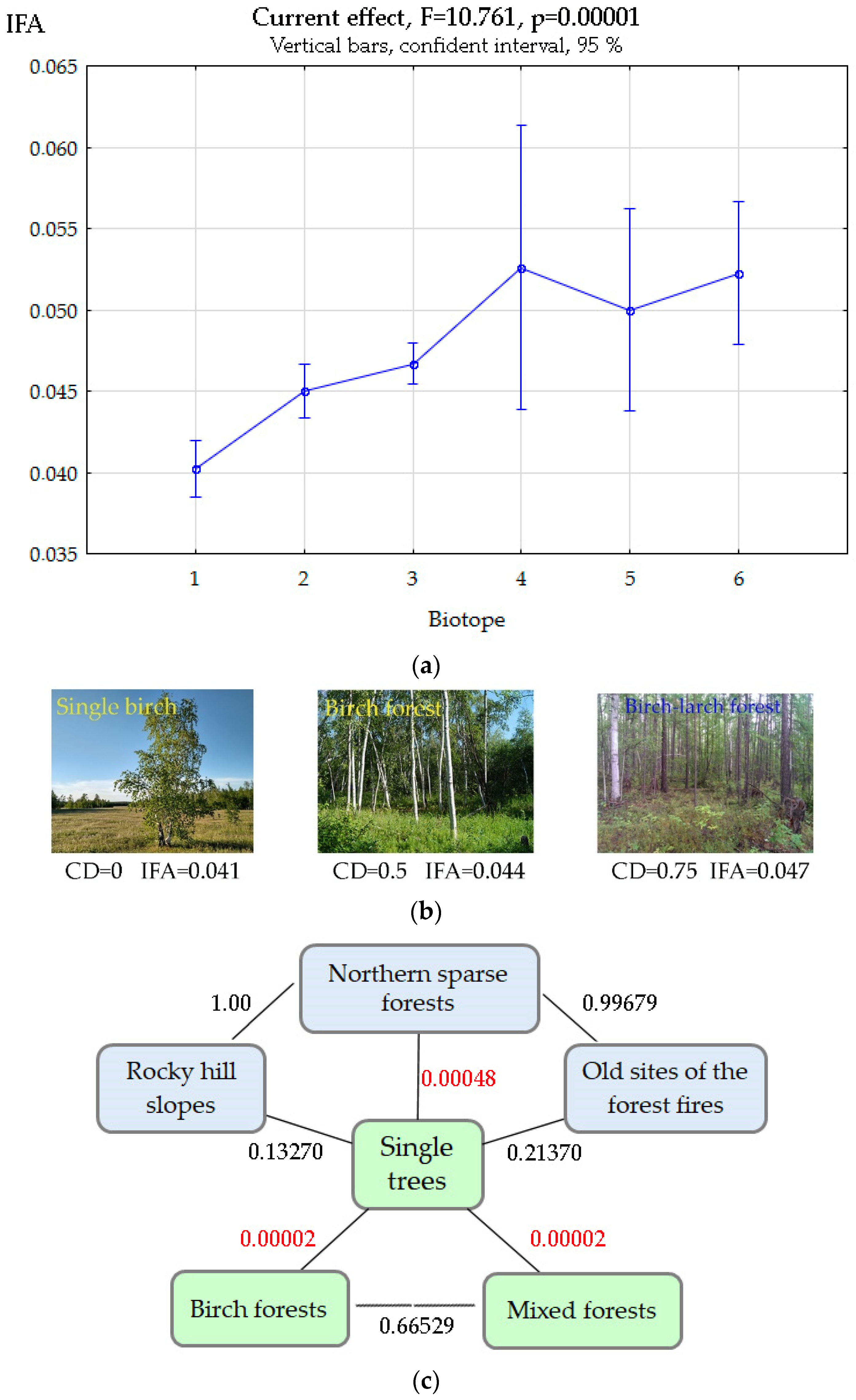
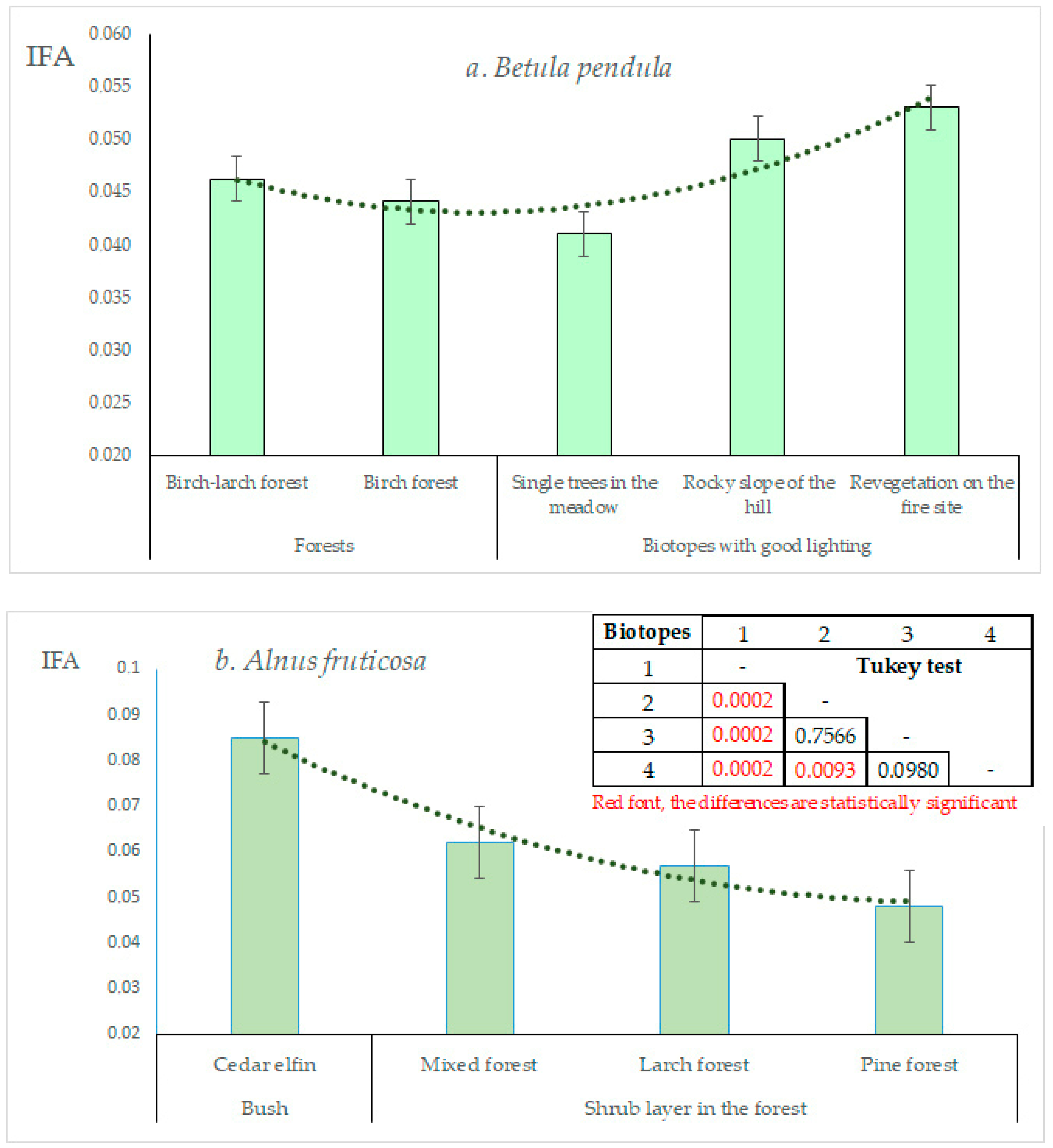
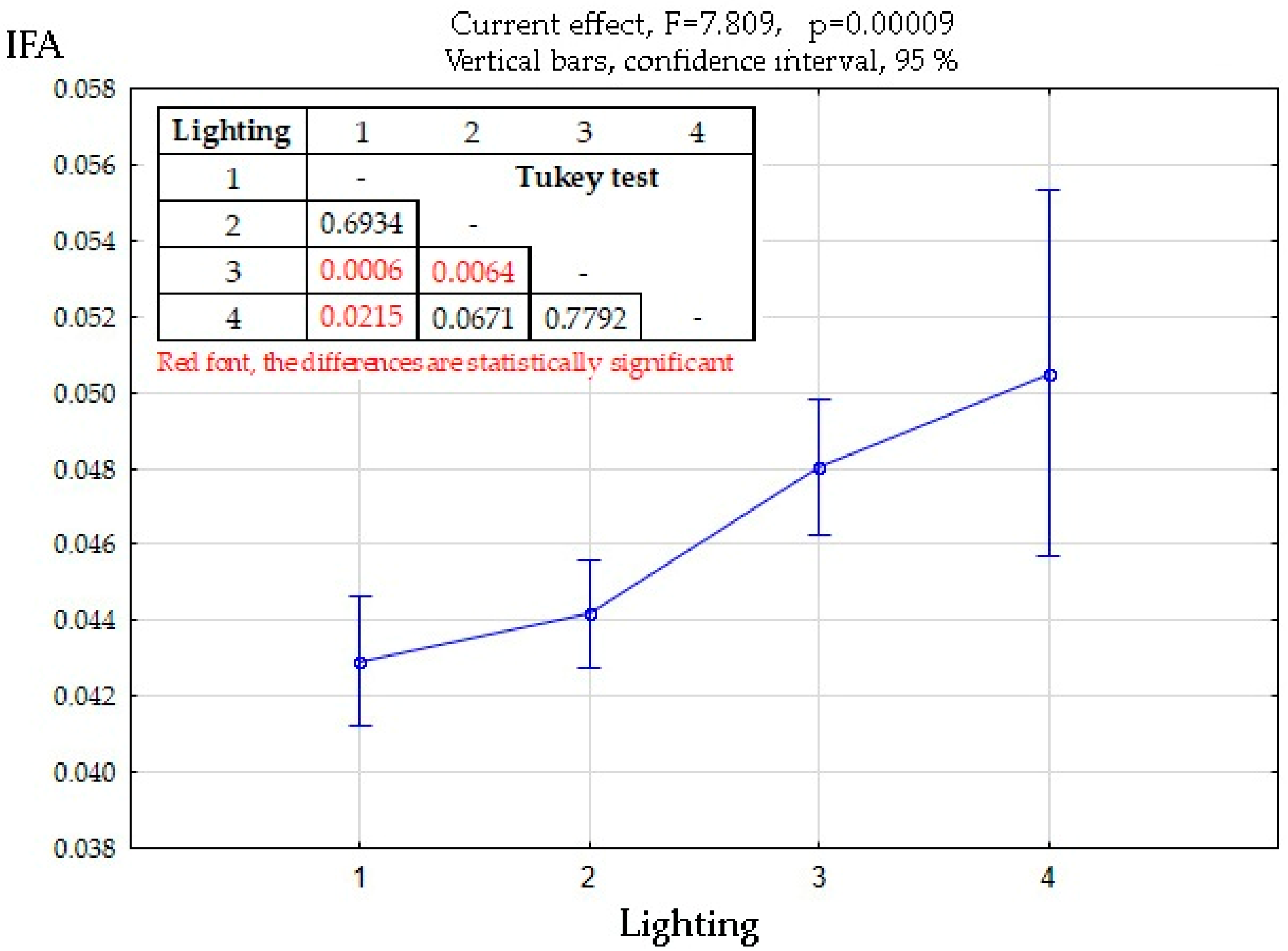
3.3. Biotopic Differences in FA Manifestations
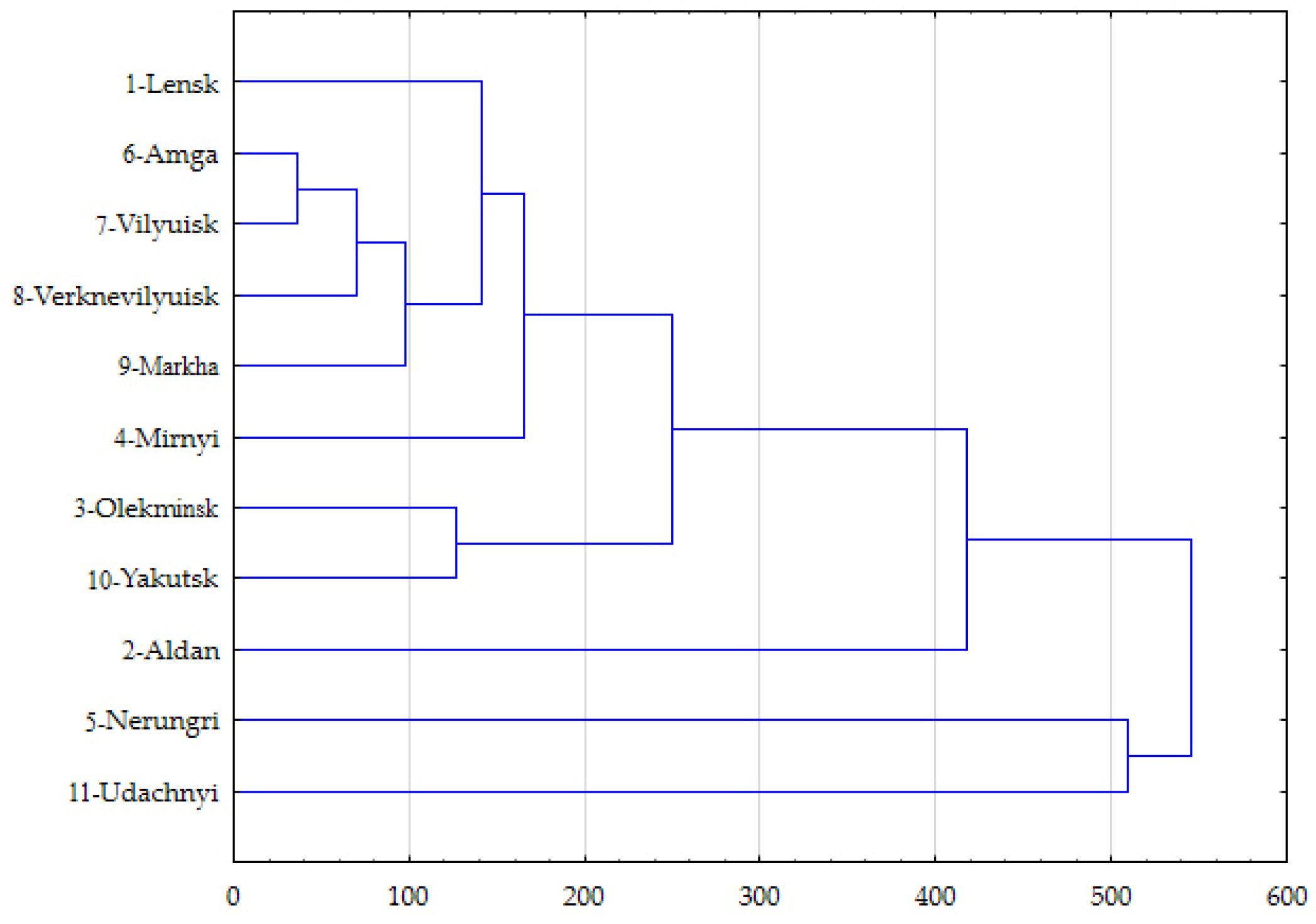
3.4. Geographical Differences in FA Manifestations
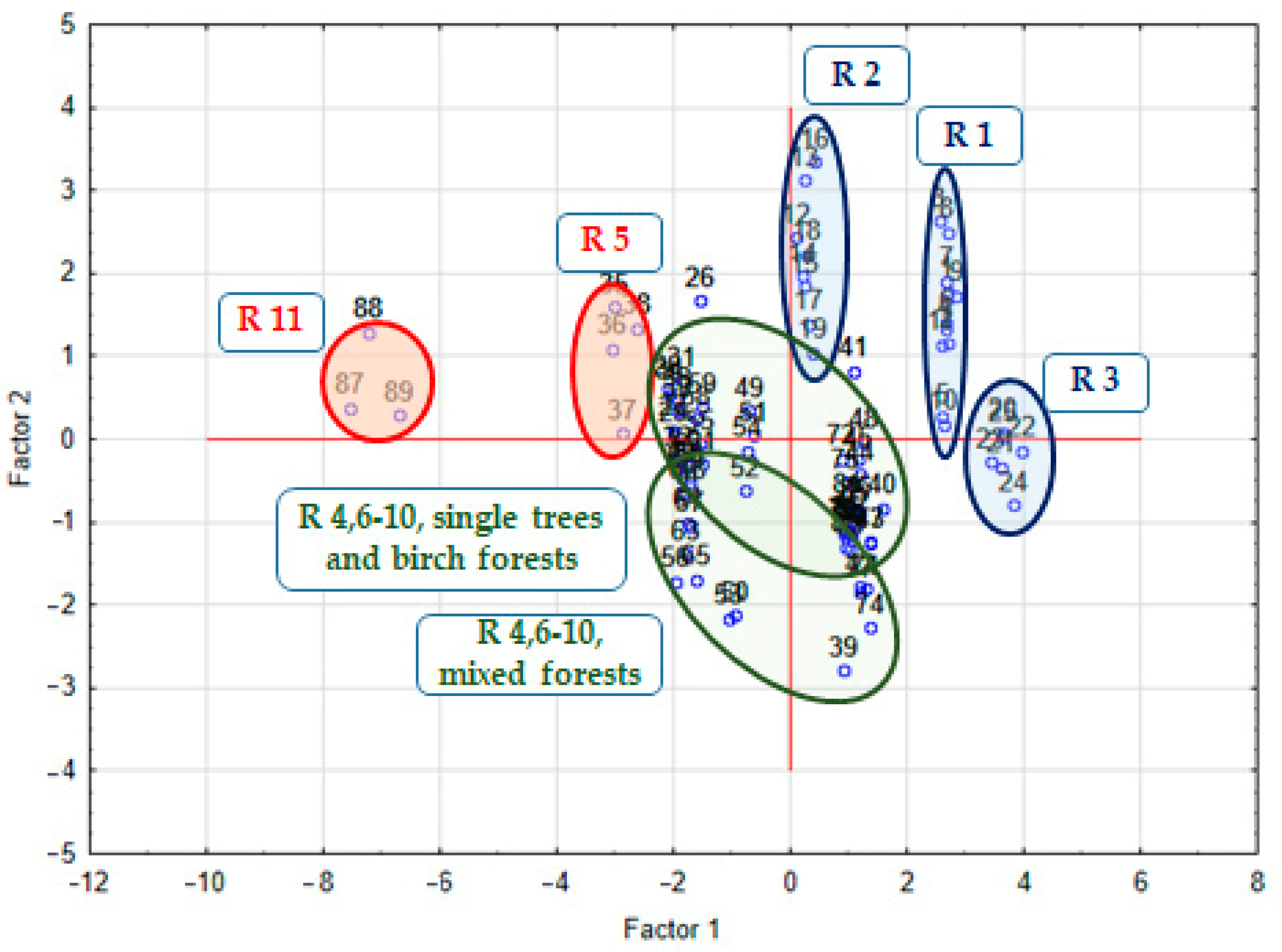
3.5. Combined Effect of the Climatic, Biotic, and Abiotic Conditions on FA
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zakharov, V.M. Asimmetriya Zhivotnykh: Populyatsionno-Fenogeneticheskii Podkhod (Animal Asymmetry: Population-Phenogenetic Approach); Nauka: Moscow, Russia, 1987; p. 216. (In Russian) [Google Scholar]
- Leary, R.F.; Allendorf, F.W. Fluctuating asymmetry as an indicator of stress: Implications for conservation biology. Trends Ecol. Evol. 1989, 4, 214–217. [Google Scholar] [CrossRef] [PubMed]
- Palmer, R.A.; Strobeck, C. Fluctuating assymmetry: A measurement, analysis, patterns. Ann. Rev. Ecol. Syst. 1986, 17, 391–421. [Google Scholar] [CrossRef]
- Parsons, P.A. Fluctuating asymmetry: An epigenetic measure of stress. Biol. Rev. 1990, 65, 131–145. [Google Scholar] [CrossRef] [PubMed]
- Parsons, P.A. Fluctuating asymmetry: A biological monitor of environmental and genomic stress. Heredity 1992, 68, 361–364. [Google Scholar] [CrossRef] [PubMed]
- Zakharov, V.M.; Krysanov, E.Y. (Eds.) Posledstviya Chernobyl’skoi Katastrofy: Zdorov’e Sredy (Consequences of the Chernobyl Disaster: Environmental Health); Tsentr Ekologicheskoi Politiki Rossii: Moscow, Russia, 1996; p. 170. (In Russian) [Google Scholar]
- Oleksyk, T.K.; Novak, J.M.; Purdue, J.R.; Gashchak, S.P.; Smith, M.H. High levels of fluctuating asymmetry in populations of Apodemus flavicollis from the most contaminated areas in Chornobyl. J. Environ. Radioactiv. 2004, 73, 1–20. [Google Scholar] [CrossRef]
- Shadrina, E.G.; Soldatova, V.Y.; Shadrin, D.Y.; Kapitonova, N.N. Evaluation of Environmental Quality by the Indices of Disturbance of Stable Development in Plants and Animals in the Conditions of Permanently Increased Radiation Background over the Yakutia Territory. Reg. Environ. Issues 2008, 2, 91–95. [Google Scholar]
- Fuller, N.; Smith, J.T.; Nagorskaya, L.L.; Gudkov, D.I.; Ford, A.T. Does Chernobyl-derived radiation impact the developmental stability of Asellus aquaticus 30 years on? Sci. Total Environ. 2017, 576, 242–250. [Google Scholar] [CrossRef]
- Pankakoski, E.; Koivisto, I.; Hyvärinen, H. Reduced developmental stability as an indicator of heavy metal pollution in the common shrew Sorex araneus. Acta Zool. Fenn. 1992, 191, 137–144. [Google Scholar]
- Chubinishvili, A.T. Homeostasis in the populations of marsh frog (Rana ridibunda Pall) living in conditions of chemical pollution in the area of the Middle Volga. Russ. J. Ecol. 1998, 1, 71–74. [Google Scholar]
- Zakharov, V.M.; Chubinishvili, A.T.; Dmitriev, S.G.; Baranov, A.S.; Borisov, V.I.; Valetskii, A.V.; Krysanov, E.U.; Kryazheva, N.G.; Pronin, A.V.; Chistyakova, E.K. Zdorov’e Sredy: Praktika Otsenki (Health of the Environment: Assessment Practice); Tsentr Ekologicheskoi Politiki Rossii: Moscow, Russia, 2000; p. 320. (In Russian) [Google Scholar]
- Shadrina, E.G.; Vol’pert, Y.L.; Danilov, V.A.; Shadrin, D.Y. Bioindikatsiya Vozdeistviya Gornodobyvayushchei Promyshlennosti Na Nazemnye Ekosistemy Severa (Morfogeneticheskii Podkhod) (Bioindication Impact of Mining on Terrestrial Ecosystems of the North (Morphogenetic Approach)); Savvinov, D.D., Ed.; Nauka, Sib. Otdelenie: Novosibirsk, Russia, 2003; p. 110. (In Russian) [Google Scholar]
- Lens, L.; Eggermont, H. Fluctuating asymmetry as a putative marker of human-induced stress in avian conservation. Bird Conserv. Int. 2008, 18, S125–S143. [Google Scholar] [CrossRef]
- Shadrina, E.G.; Vol’pert, Y.L. Developmental Instability of the Organism as a Result of Pessimization of Environment under Anthropogenic Transformation of Natural Landscapes. Russ. J. Dev. Biol. 2014, 45, 117–126. [Google Scholar] [CrossRef]
- Zhelev, Z.M.; Popgeorgiev, G.S.; Arnaudov, A.D.; Georgieva, K.N.; Mehterov, N.H. Fluctuating Asymmetry in Pelophylax Ridibundus (Amphibia: Ranidae) as a Response to Anthropogenic Pollution in South Bulgaria. Arch. Biol. Sci. 2015, 67, 1009–1023. [Google Scholar] [CrossRef]
- Lutterschmidt, W.I.; Martin, S.L.; Schaefer, J.F. Fluctuating Asymmetry in Two Common Freshwater Fishes as a Biological Indicator of Urbanization and Environmental Stress within the Middle Chattahoochee Watershed. Symmetry 2016, 8, 124. [Google Scholar] [CrossRef]
- Guo, R.; Zhang, W.; Ai, S.; Ren, L.; Zhang, Y. Fluctuating asymmetry rather than oxidative stress in Bufo raddei can be an accurate indicator of environmental pollution induced by heavy metals. Environ. Monit. Assess 2017, 189, 293. [Google Scholar] [CrossRef] [PubMed]
- Shadrina, E.G.; Vol’pert, Y.L. Experience of Applying Plant and Animal Fluctuating Asymmetry in Assessment of Environmental Quality in Terrestrial Ecosystems: Results of 20-Year Studies of Wildlife and Anthropogenically Transformed Territories. Russ. J. Dev. Biol. 2018, 49, 23–35. [Google Scholar] [CrossRef]
- Zhelev, Z.M.; Tsonev, S.V.; Angelov, M.V. Fluctuating asymmetry in Pelophylax ridibundus meristic morphological traits and their importance in assessing environmental health. Ecol. Indic. 2019, 107, 105589. [Google Scholar] [CrossRef]
- Benítez, H.A.; Lemic, D.; Villalobos-Leiva, A.; Bažok, R.; Órdenes-Claveria, R.; Pajač Živković, I.; Mikac, K.M. Breaking Symmetry: Fluctuating Asymmetry and Geometric Morphometrics as Tools for Evaluating Developmental Instability under Diverse Agroecosystems. Symmetry 2020, 12, 1789. [Google Scholar] [CrossRef]
- Turmukhametova, N.V. Evaluation of the State of the Environment in Yoshkar-Ola Using Morphometric Indicators of Betula pendula Roth. Biol. Bull. 2020, 47, 191–197. [Google Scholar] [CrossRef]
- Kryazheva, N.G.; Chistyakova, E.K.; Zakharov, V.M. Analysis of stability of the development of Betula pendula under conditions of chemical pollution. Russ. J. Ecol. 1996, 27, 422–424. [Google Scholar]
- Turmukhametova, N.V.; Shivtsova, I.V. Morfologicheskii podhod k ocenke sostoyaniya sredy po asimmetrii lista Betula pendula Roth i Fragaria vesca L. (Morphological approach to assessing the state of the environment based on leaf asymmetry of Betula pendula Roth and Fragaria vesca L.). Lesn. Vestn. (For. Bull.) 2007, 5, 140–143. (In Russian) [Google Scholar]
- Vasil’ev, A.G.; Vasil’eva, I.A.; Marin, Y.F. Phenogenetic monitoring of the weeping birch (Betula pendula Roth.) in the Middle Urals: Testing a new method for assessing developmental instability in higher plants. Russ. J. Ecol. 2008, 39, 25–31. [Google Scholar] [CrossRef]
- Shadrina, E.G.; Volpert, Y.L.; Alekseeva, N.N.; Danilov, V.A.; Stepanova, T.M. Bioindikacionnaya ocenka izmeneniya kachestva okruzhayushchei sredy v rezul’tate vozdejstviya almazodobyvayushchih predpriyatii (Bioindicational Estimation of Environment Quality Changing As A Result of The Influence of Diamond Mining Companies). Gorn. Zhurnal 2012, 2, 84–87. (In Russian) [Google Scholar]
- Youssef, N.A.; Gurbanov, E.M.; Haciyeva, S.R.; Mammedova, A.O.; Khalilov, R.I. Antioxidant enzymes, fluctuating asymmetry and morphological changes of urban trees as an ecological indicators of heavy metal stress. Int. J. Pharm. Sci. Health Care 2013, 3. Available online: http://www.rspublication.com/ijphc/index.html (accessed on 15 April 2022).
- Erofeeva, E.A. Fluktuiruyushchaya asimmetriya lista Betula pendula (Betulacea) v usloviyah avtotransportnogo zagryazneniya (g. Nizhni Novgorod) (Dependence of leaf fluctuating asymmetry of Betula pendula (Betulacea) on motor traffic pollution intensity). Rastit. Resur. 2015, 51, 366–383. (In Russian) [Google Scholar]
- Ivanov, V.P.; Ivanov, Y.V.; Marchenko, S.I.; Kuznetsov, V.V. Application of fluctuating asymmetry indexes of silver birch leaves for diagnostics of plant communities under technogenic pollution. Russ. J. Plant Physiol. 2015, 62, 340–348. [Google Scholar] [CrossRef]
- Rodrigues, R.A.R.; Vaz, V.C.; Sato, A.; Arruda, R.C.O.; de Castro, W.A.C.; da Silva-Matos, D.M. Structural leaf changes in trees around a subway air duct. Rev. Árvore Viçosa-MG 2015, 39, 417–421. [Google Scholar] [CrossRef]
- Shadrina, E.; Vol’pert, Y.; Soldatova, V.; Alekseeva, N.; Pudova, T. Evaluation of Environmental Conditions in Two Cities of East Siberia Using Bioindication Methods (Fluctuating Asymmetry Value and Mutagenic Activity of Soils). Int. J. Biol. 2015, 7, 20–32. [Google Scholar] [CrossRef]
- Shadrina, E.G.; Pudova, T.M.; Soldatova, V.Y.; Legostaeva, Y.B. Bioindicational Assessment Of Environmental Quality In Habitats Exposed To Geological Exploration For Hydrocarbon (Oil And Gas) Deposits In The West Yakutia. In Proceedings of the 15th International Multidisciplinary Scientific Geoconference SGEM 2015—Ecology, Economics, Education and Legislation, Albena, Bulgaria, 18–24 June 2015; Volume 1, pp. 95–102. [Google Scholar]
- Mendes, G.; Boaventura, M.G.; Cornelissen, T. Fluctuating Asymmetry as a Bioindicator of Environmental Stress Caused by Pollution in a Pioneer Plant Species. Environ. Entomol. 2018, 47, 1479–1484. [Google Scholar] [CrossRef]
- Erofeeva, E.A.; Yakimov, B.N. Change of Leaf Trait Asymmetry Type in Tilia cordata Mill. and Betula pendula Roth under Air Pollution. Symmetry 2020, 12, 727. [Google Scholar] [CrossRef]
- Prihod’ko, S.A.; Shtirc, Y.A. Primenenie morfogeometricheskogo metoda dlya opredeleniya pokazatelej fluktuiruyushchej asimmetrii listovoj plastinki Aser pseudoplatanus L. (Application of the morphometric method for determining the indicators of fluctuating asymmetry leaf plate Acer pseudoplatanus L.). Ukr. J. Ecol. 2018, 8, 194–210. (In Russian) [Google Scholar]
- Shadrina, E.; Turmukhametova, N.; Soldatova, V.; Vol’pert, Y.; Korotchenko, I.; Pervyshina, G. Fluctuating Asymmetry in Morphological Characteristics of Betula Pendula Roth Leaf under Conditions of Urban Ecosystems: Evaluation of the Multi-Factor Negative Impact. Symmetry 2020, 12, 1317. [Google Scholar] [CrossRef]
- Turmukhametova, N.; Shadrina, E. Changes in the Fluctuating Asymmetry of the Leaf and Reproductive Capacity of Betula pendula Roth Reflect Pessimization of Anthropogenically Transformed Environment. Symmetry 2020, 12, 1970. [Google Scholar] [CrossRef]
- Zakharov, V.M.; Shadrina, E.G.; Turmukhametova, N.V.; Ivantsova, E.N.; Shikalova, E.A.; Soldatova, V.Y.; Sharova, N.A.; Trofimov, I.E. Assessment of Plant Status by Stability of Development in Natural and Anthropogenic Conditions (Fluctuating Asymmetry of Leaf Characters of Silver Birch, Betula pendula Roth). Biol. Bull. 2020, 47, 186–190. [Google Scholar] [CrossRef]
- Turmukhametova, N.V.; Shadrina, E.G.; Soldatova, V.Y.; Ivantsova, E.N. Fluctuating asymmetry of the lamina of Betula pendula Roth in the context of different cities and industrial load. IOP Conf. Ser. Earth Environ. Sci. 2021, 839, 052011. [Google Scholar] [CrossRef]
- Murphy, H.T.; Lovett-Doust, J. Landscape-Level Effects on Developmental Instability: Fluctuating Asymmetry across the Range of Honey Locust, Gleditsia triacanthos (Fabaceae). Int. J. Plant Sci. 2004, 165, 795–803. [Google Scholar] [CrossRef]
- Ambo-Rappe, R.; Lajus, D.L.; Schreider, M.J. Translational Fluctuating Asymmetry and Leaf Dimension in Seagrass, Zostera capricorni Aschers in a Gradient of Heavy Metals. Environ. Bioindic. 2007, 2, 99–116. [Google Scholar] [CrossRef]
- Koroteeva, E.V.; Veselkin, D.V.; Kuyantseva, N.B.; Chashchina, O.E. The size, but not the fluctuating asymmetry of the leaf, of silver birch changes under the gradient influence of emissions of the Karabash Copper Smelter Plant. Dokl. Biol. Sci. 2015, 460, 36–39. [Google Scholar] [CrossRef]
- Sandner, T.M.; Matthies, D. Fluctuating asymmetry of leaves is a poor indicator of environmental stress and genetic stress by inbreeding in Silene vulgaris. Ecol. Indic. 2017, 79, 247–253. [Google Scholar] [CrossRef]
- Zverev, V.; Lama, A.D.; Kozlov, M.V. Fluctuating asymmetry of birch leaves did not increase with pollution and drought stress in a controlled experiment. Ecol. Indic. 2018, 84, 283–289. [Google Scholar] [CrossRef]
- Zakharov, V.M.; Zhdanova, N.P.; Kirik, E.F.; Shkil, F.N. Ontogenesis and Population: Evaluation of Developmental Stability in Natural Populations. Russ. J. Dev. Biol. 2001, 32, 336–351. [Google Scholar] [CrossRef]
- Freeman, D.C.; Graham, J.H.; Emlen, J.M. Developmental stability in plants: Symmetries, stress and epigenesis. Genetica 1993, 89, 97–119. [Google Scholar] [CrossRef]
- Rettig, J.E.; Fuller, R.C.; Corbett, A.L.; Getty, T. Fluctuating Asymmetry Indicates Levels of Competition in an Even-Aged Poplar Clone. Oikos 1997, 80, 123–127. [Google Scholar] [CrossRef]
- Wilsey, B.J.; Haukioja, E.; Koricheva, J.; Sulkinoja, M. Leaf fluctuating asymmetry increases with Hybridization and elevation in tree-line birches. Ecology 1998, 79, 2092–2099. [Google Scholar] [CrossRef]
- Hagen, S.B.; Folstad, I.; Jakobsen, S.W. Autumn colouration and herbivore resistance in mountain birch (Betula pubescens). Ecol. Lett. 2003, 6, 807–811. [Google Scholar] [CrossRef]
- Freeman, D.C.; Brown, M.L.; Duda, J.J.; Graraham, J.H.; Emlen, J.M.; Krzysik, A.J.; Balbach, H.; Kovacic, D.A.; Zak, J.C. Leaf fluctuating asymmetry, soil disturbance and plant stress: A multiple year comparison using two herbs, Ipomoea pandurata and Cnidoscolus stimulosus. Ecol. Indic. 2005, 5, 85–95. [Google Scholar] [CrossRef]
- Gelashvili, D.B.; Lobanova, I.V.; Yerofeeva, E.A.; Naumova, M.M. Vliyanie lesopatologicheskogo sostoyaniya breezy povisloj na velichinu fluktuiruyushchej asimmetrii listovoj plastinki (Influence of the phytopathologic state of Betula pendula Roth. fluctuating asymmetry of its leaf laminae). Povolzhskiy J. Ecol. 2007, 2, 106–115. (In Russian) [Google Scholar]
- Hagen, S.B.; Ims, R.A.; Yoccoz, N.G.; Sørlibraten, O. Fluctuating asymmetry as an indicator of elevation stress and distribution limits in mountain birch (Betula pubescens). Plant Ecol. 2008, 195, 157–163. [Google Scholar] [CrossRef]
- Xu, Z.; Hu, T.; Zhang, Y. Effects of experimental warming on phenology, growth and gas exchange of treeline birch (Betula utilis) saplings, Eastern Tibetan Plateau, China. Eur. J. Forest Res. 2012, 131, 811–819. [Google Scholar] [CrossRef]
- Santos, J.C.; Alves-Silva, E.; Cornelissen, T.G.; Fernandes, G.W. The effect of fluctuating asymmetry and leaf nutrients on gall abundance and survivorship. Basic Appl. Ecol. 2013, 14, 489–495. [Google Scholar] [CrossRef]
- Shadrina, E.; Volpert, Y. Fluctuating Asymmetry of Craniological Features of Small Mammals as a Reflection of Heterogeneity of Natural Populations. Symmetry 2016, 8, 142. [Google Scholar] [CrossRef]
- Shadrina, E.G.; Soldatova, V.Y. Vliyanie bioticheskih i abioticheskih faktorov na proyavleniya fluktuiruyushchei asimmetrii v populyaciyah drevesnyh rastenii (na primere Yakutii) (Influence of biotic and abiotic factors on the manifestations of fluctuating asymmetry in populations of woody plants (on the example of Yakutia)). In Proceedings of the Problemy Populyacionnoi Biologii (Problems of Population Biology), Yoshkar-Ola, Russia, 11–14 April 2017; p. 260262. (In Russian). [Google Scholar]
- Zakharov, V.M.; Sikorski, M.D. Inbreeding and developmental stability in a laboratory strain of the bank vole Clethrionomys glareolus. Acta Theriol. 1997, 4, 73–78. [Google Scholar] [CrossRef]
- Siikamaki, P. Developmental instability in hybrids between Lychnis viscaria and Lychnis alpina (Caryophyllaceae). Am. J. Bot. 1999, 86, 1683–1686. [Google Scholar] [CrossRef] [PubMed]
- Botanic Gardens, Kew and Missouri Botanical (Garden). The Plant List. A working List of All Plant Species. Betula pendula Roth is an Accepted Name. Version 1.1. Published on the Internet; 2013. Available online: http://www.theplantlist.org/ (accessed on 31 July 2020).
- Afonin, A.N.; Greene, S.; Frolov, A. Interactive Agricultural Ecological Atlas of Russia and Neighboring Countries. Economic Plants and their Diseases, Pests and Weeds. 2009. Available online: http://www.agroatlas.ru (accessed on 22 April 2022).
- Kuznetcova, L.V.; Zakharova, V.I. Konspekt Flory Yakutii: Sosudistye Rasteniya (A Compendium of the Flora of Yakutia); Nauka: Novosibirsk, Russia, 2012; p. 272. (In Russian) [Google Scholar]
- Zhukova, L.A.; Dorogova, Y.A.; Turmuhametova, N.V.; Zhukova, L.A. (Eds.) Ekologicheskie Shkaly i Metody Analiza Ekologicheskogo Raznoobraziya Rastenii (Ecological Indicator Values and Methods of Analysis of Ecological Diversity of Plants); Mari State University: Yoshkar-Ola, Russia, 2010; p. 368. [Google Scholar]
- Dorogova, Y.A.; Zhukova, L.A.; Turmuhametova, N.V.; Polyanskaya, T.A.; Notov, A.A.; Dementyeva, S.M. Methods of Analysis of Environmental Diversity of Plants. Biol. Med. 2016, 8, 354. [Google Scholar] [CrossRef]
- Volpert, Y.K.; Danilov, P.P.; Danilov, Y.G.; Isaev, A.P.; Nikolin, E.G.; Pakhomova, L.S.; Pestryakova, L.A.; Petrova, A.N.; Savvinov, D.D.; Savvinov, G.N.; et al. Republic of Sakha (Yakutia). 7.3. Physical geography. In Geography of Siberia in the Early 21st Century; Korytny, L.M., Tulokhonov, A., Eds.; Geo: Novosibirsk, Russia, 2016; Volume 6, pp. 317–336. (In Russian) [Google Scholar]
- Ivanov, I.N. (Ed.) Yakutia, Historical and Cultural Atlas; Publishing and Production Center “Design, Information. Cartography”: Moscow, Russia, 2007; p. 869. (In Russian) [Google Scholar]
- Shcherbakov, I.P. Lesnoi Pokrov Severo-Vostoka SSSR (Forest Cover of the North-East of the USSR). Nauka, Siberian Branch: Novosibirsk, Russia, 1975. (In Russian) [Google Scholar]
- Timofeev, P.A. Lesa Yakutii. Sostav, Resursy, Ispol’zovanie i Ohrana (Forests of Yakutia. Composition, Resources, Use and Protection); Isaev, A.P., Ed.; Publishing House of the Siberian Branch of the Russian Academy of Sciences: Novosibirsk, Russia, 2003; p. 194. (In Russian) [Google Scholar]
- Sukachev, V.N.; Zonn, S.V. Metodicheskie Ukazaniya k Izucheniyu Tipov Lesa (Methodological Guidelines for the Study of Forest Types); Publishing House of the USSR Academy of Sciences: Moscow, Russia, 1961. [Google Scholar]
- Weather Archives. Yakutia, Russia. Pogodaiklimat (Weather and Climate). Available online: http://www.pogodaiklimat.ru/history/24959_3.htm (accessed on 15 April 2020). (In Russian).
- Agroklimaticheskij Spravochnik po Yakutskoi ASSR (Agro-Climatic Guide of the Yakut ASSR); Hydrometeoizdat: Leningrad, Russia, 1963. (In Russian)
- Smirnova, N.S. (Ed.) Nauchno-Prikladnoj Spravochnik po Klimatu SSSR (Scientific and Applied Reference Book on the Climate of the USSR); Ser. 3, Book 1; Hydrometeoizdat: Leningrad, Russia, 1989; Volume 24, p. 607. (In Russian) [Google Scholar]
- Zakharov, V.M.; Baranov, A.S.; Borisov, V.I.; Valetskii, A.V.; Kryazheva, N.G.; Chistyakova, E.K.; Chubinishvili, A.T. Zdorov’eSredy: Metodika Otsenki (Health of the Environment: Metodology Evaluation); Tsentr Ekologicheskoi Politiki Rossii: Moscow, Russia, 2000; p. 68. (In Russian) [Google Scholar]
- Statistica. StatSoft Russia. Available online: http://statsoft.ru/company/ (accessed on 19 April 2021).
- Sokal, R.R.; Rohlf, F.J. Biometry: The Principles and Practice of Statistics in Biological Research, 3rd ed.; W.H. Freeman and Co.: New York, NY, USA, 1995; p. 887. [Google Scholar]
- Voropaev, G.V.; Avakian, A. (Eds.) Vodohranilishcha i ih Vozdejstvie na Okruzhayushchuyu Sredu (Reservoirs and Their Impact on the Environment); Nauka: Moscow, Russia, 1986; p. 366. (In Russian) [Google Scholar]
- Shadrina, E.G. Vilyujskij Gidrouzel: Social’no-Ekologicheskie Posledstviya (Vilyuysky Hydroelectric Power Plant: Socio-ECOLOGICAL Consequences). Available online: https://www.plotina.net/experts/vilyujskij-gidrouzel-socialno-ekologicheskie-posledstviya/ (accessed on 22 April 2022). (In Russian).
- Soldatova, V.Y.; Samsonova, A.P. Vliyanie ekologicheskih uslovij gorodskoj sredy na kachestvo semyan i morfologicheskie pokazateli listovoj plastinki berezy povisloj Betula pendula Roth (The influence of environmental conditions of the urban environment on the quality of seeds and morphological indicators of the Birch leaf blade (Betula pendula Roth). Probl. Reg. Ekol. 2018, 6, 12–15. (In Russian) [Google Scholar]
- Dolgoshov, V.I. Urozhainost’ plodov glavneishikh drevesnykh porod i kustarnikov (Fruit yield of the main species of trees and shrubs). Geogr. Sb. 1963, 16, 131–149. (In Russian) [Google Scholar]
- Stukov, V.I. Osobennosti tsveteniya i plodonosheniya berezy borodavchatoi v Saratovskom Povolzh’e (Specifics of blossoming and fruit producing in the silver birch in Povolzhye in the vicinity of Saratov). Lesn. Khozyaistvo (For. Manag.) 1967, 8, 37–38. (In Russian) [Google Scholar]
- Danchenko, A.M. Biologiya Plodonosheniya i Osnovy Semenodostva Berezy (Biology of Fruit Bearing and the Basics of Seed Farming of the Birch); ILID Publisher: Krasnoyarsk, Russia, 1992; p. 126. (In Russian) [Google Scholar]
- Alves-Silva, E.; Del-Claro, K. Effect of post-fire resprouting on leaf fluctuating asymmetry, extrafloral nectar quality, and ant–plant–herbivore interactions. Naturwissenschaften 2013, 100, 525–532. [Google Scholar] [CrossRef]
- Turskii, M.K. Lesovodstvo (Silviculture); Gosudarstvennoe Izdatel’stvo Selskokhozyaistvennoi Literatury: Moscow, Russia, 1954; p. 352. (In Russian) [Google Scholar]
- Tikhonov, A.S. Lesovedenie (Forestry); Oblizdat: Kaluga, Russia, 2011; p. 329. (In Russian) [Google Scholar]
- Alves-Silva, E. The influence of Ditylenchus (Nematoda) galls and shade on the fluctuating asymmetry of Miconia fallax (Melastomataceae). Ecol. Austral. 2012, 22, 53–61. [Google Scholar]
- Moller, A.P. Leaf-mining Insects and fluctuating asymmetry in elm Ulmus glabra leaves. J. Anim. Ecol. 1995, 64, 697–707. [Google Scholar] [CrossRef]
- Martel, J.; Lempa, K.; Haukioja, E. Effect of stress and rapid growth on fluctuating asymmetry and insect damage in birch leaves. OIKOS 1999, 86, 208–216. [Google Scholar] [CrossRef]
- Cuevas-Reyes, P.; Canché-Delgado, A.; Maldonado-López, Y.; Fernandes, G.W.; Oyama, K.; González-Rodríguez, A. Patterns of herbivory and leaf morphology in two Mexican hybrid oak complexes: Importance of fluctuating asymmetry as indicator of environmental stress in hybrid plants. Ecol. Ind. 2018, 90, 164–170. [Google Scholar] [CrossRef]
- Graham, J.H.; Raz, S.; Hel-Or, H.; Nevo, E. Fluctuating asymmetry: Methods, theory, and applications. Symmetry 2010, 2, 466–540. [Google Scholar] [CrossRef]
- Zakharov, V.M.; Shadrina, E.G.; Trofimov, I.E. Fluctuating Asymmetry, Developmental Noise Biology Approach. Symmetry 2020, 12, 1376. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
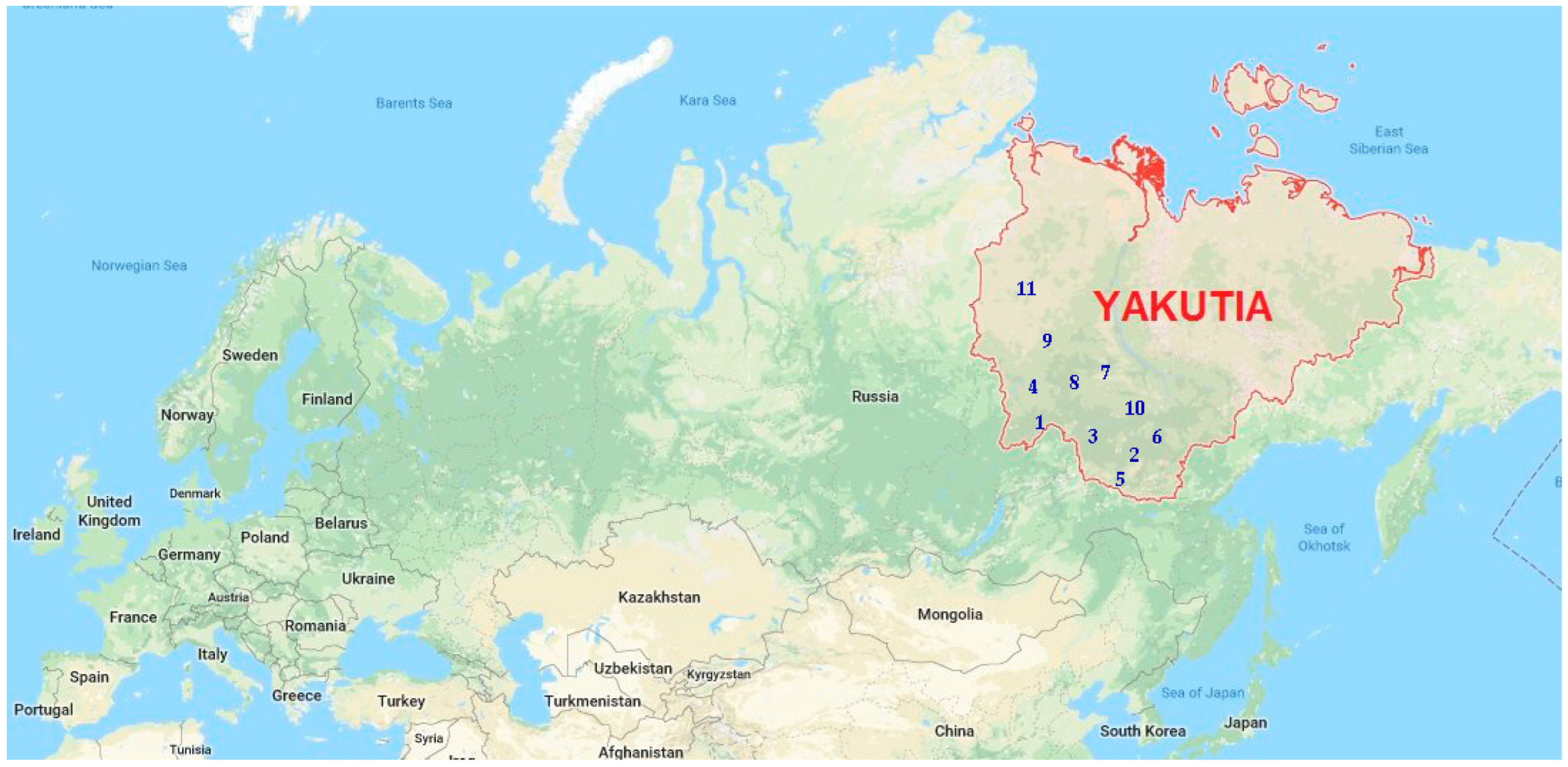
| Number | Region ** | Average Temperature, °C | Geographical Coordinates * | Material | ||
|---|---|---|---|---|---|---|
| Latitude, North | Longitude, East | Elevation, AMSL, m | Sites/Leaves Amount | |||
| 1 | Lensk | −5.29 | 60.72 | 114.8 | 241 | 14/1400 |
| 2 | Aldan | −5.53 | 58.6 | 125.3 | 679 | 10/1000 |
| 3 | Olekminsk | −5.85 | 60.4 | 120.4 | 230 | 7/700 |
| 4 | Mirny | −6.82 | 62.53 | 114.04 | 352 | 11/1100 |
| 5 | Nerungri | −6.94 | 56.83 | 124.86 | 858 | 4/400 |
| 6 | Amga | −9.97 | 60.9 | 131.9 | 146 | 12/1200 |
| 7 | Vilyuisk | −8.02 | 63.77 | 121.6 | 123 | 10/1000 |
| 8 | Verkhnevilyuisk | −8.06 | 63.45 | 120.2 | 115 | 24/2400 |
| 9 | Markha | −8.20 | 63.28 | 118.3 | 119 | 14/1400 |
| 10 | Yakutsk | −8.84 | 62.0 | 129.6 | 230 | 21/2100 |
| 11 | Udachny | −9.57 | 66.73 | 112.43 | 501 | 3/300 |
| Total | 130/13,000 | |||||
| Region 2 | Amount of Sites | IFA 3 | |
|---|---|---|---|
| n | M ± m | ||
| 1 | 12 | 1200 | 0.043 ± 0.0019 |
| 2 | 9 | 900 | 0.041 ± 0.0017 * |
| 4 | 11 | 1100 | 0.046 ± 0.0007 |
| 5 | 3 | 300 | 0.050 ± 0.0019 |
| 6 | 5 | 500 | 0.047 ± 0.0016 |
| 7 | 24 | 2400 | 0.044 ± 0.0010 |
| 8 | 24 | 2400 | 0.042 ± 0.0010 * |
| 9 | 14 | 1400 | 0.046 ± 0.0010 |
| 10 | 11 | 1100 | 0.046 ± 0.0005 |
| 11 | 3 | 300 | 0.051 ± 0.0018 * |
| Parameters | Components | |
|---|---|---|
| I | II | |
| IFA | 0.200 | −0.715 ** |
| Biotope | −0.281 | −0.772 ** |
| Soil moisture | −0.239 | −0.148 |
| Lighting | 0.105 | −0.704 ** |
| Tree age | 0.212 | −0.250 |
| Leafe size | 0.0442 | 0.031 |
| SAT-5 | 0.767 ** | 0.194 |
| Duration of vegetative period (from +5 °C) | 0.833 ** | 0.141 |
| Spring climate inclemency | −0.930 ** | 0.139 |
| Winter climate inclemency | −0.631 | −0.029 |
| Summer climate inclemency | −0.951 ** | 0.168 |
| General climate inclemency | −0.847 ** | 0.044 |
| LEG | −0.849 ** | −0.066 |
| Total variance | 0.394 | 0.138 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shadrina, E.; Soldatova, V.; Turmukhametova, N. Fluctuating Asymmetry as a Measure of Stress in Natural Populations of Woody Plants: Influence of Ecological and Geographical Factors on Developmental Stability. Symmetry 2023, 15, 700. https://doi.org/10.3390/sym15030700
Shadrina E, Soldatova V, Turmukhametova N. Fluctuating Asymmetry as a Measure of Stress in Natural Populations of Woody Plants: Influence of Ecological and Geographical Factors on Developmental Stability. Symmetry. 2023; 15(3):700. https://doi.org/10.3390/sym15030700
Chicago/Turabian StyleShadrina, Elena, Victoria Soldatova, and Nina Turmukhametova. 2023. "Fluctuating Asymmetry as a Measure of Stress in Natural Populations of Woody Plants: Influence of Ecological and Geographical Factors on Developmental Stability" Symmetry 15, no. 3: 700. https://doi.org/10.3390/sym15030700
APA StyleShadrina, E., Soldatova, V., & Turmukhametova, N. (2023). Fluctuating Asymmetry as a Measure of Stress in Natural Populations of Woody Plants: Influence of Ecological and Geographical Factors on Developmental Stability. Symmetry, 15(3), 700. https://doi.org/10.3390/sym15030700