B Cell Tolerance in Health and Disease

Abstract
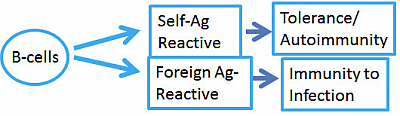
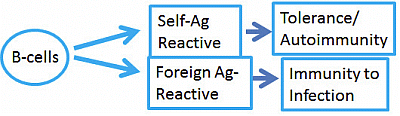
:
{kind=link}
{kind=link}
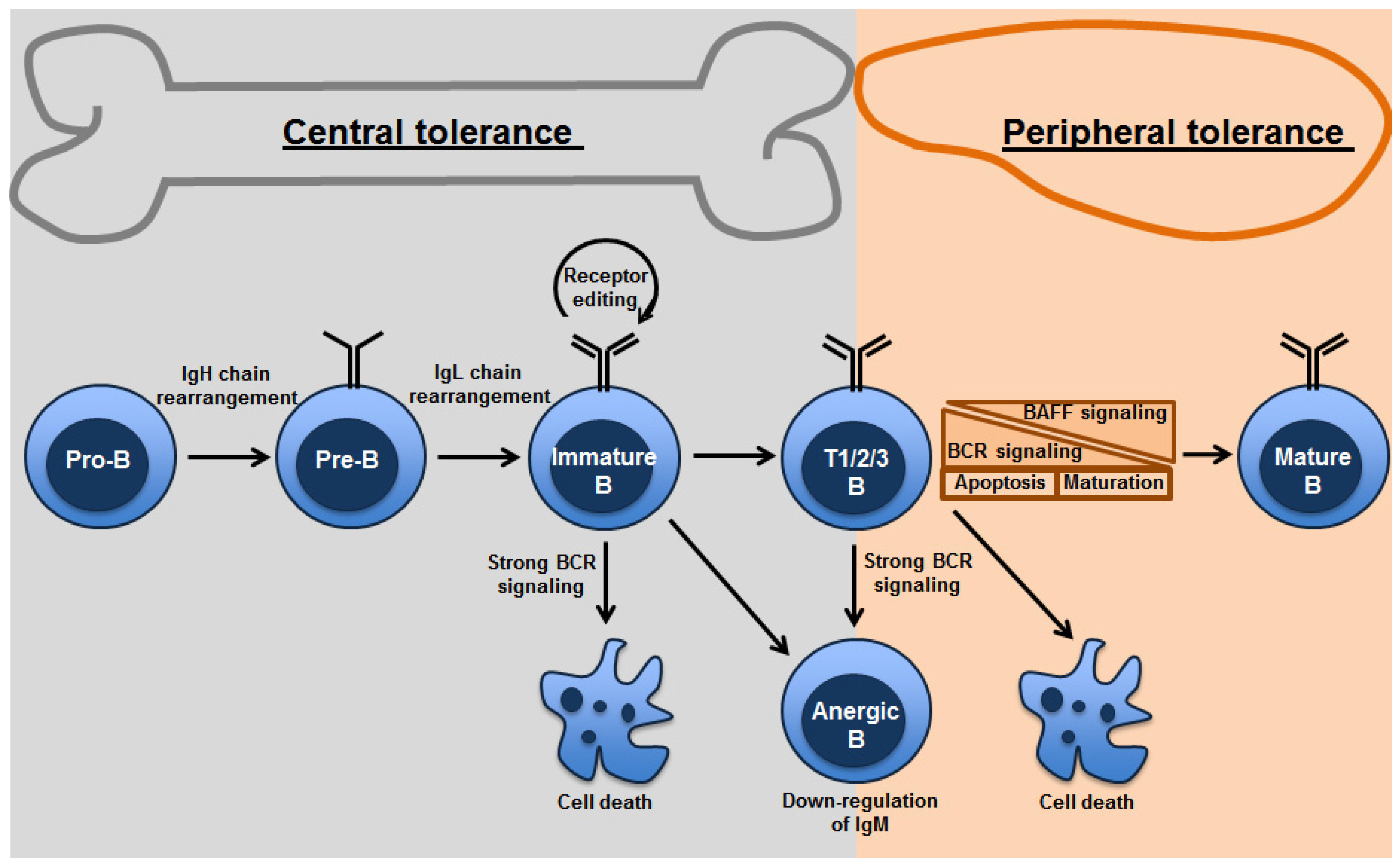
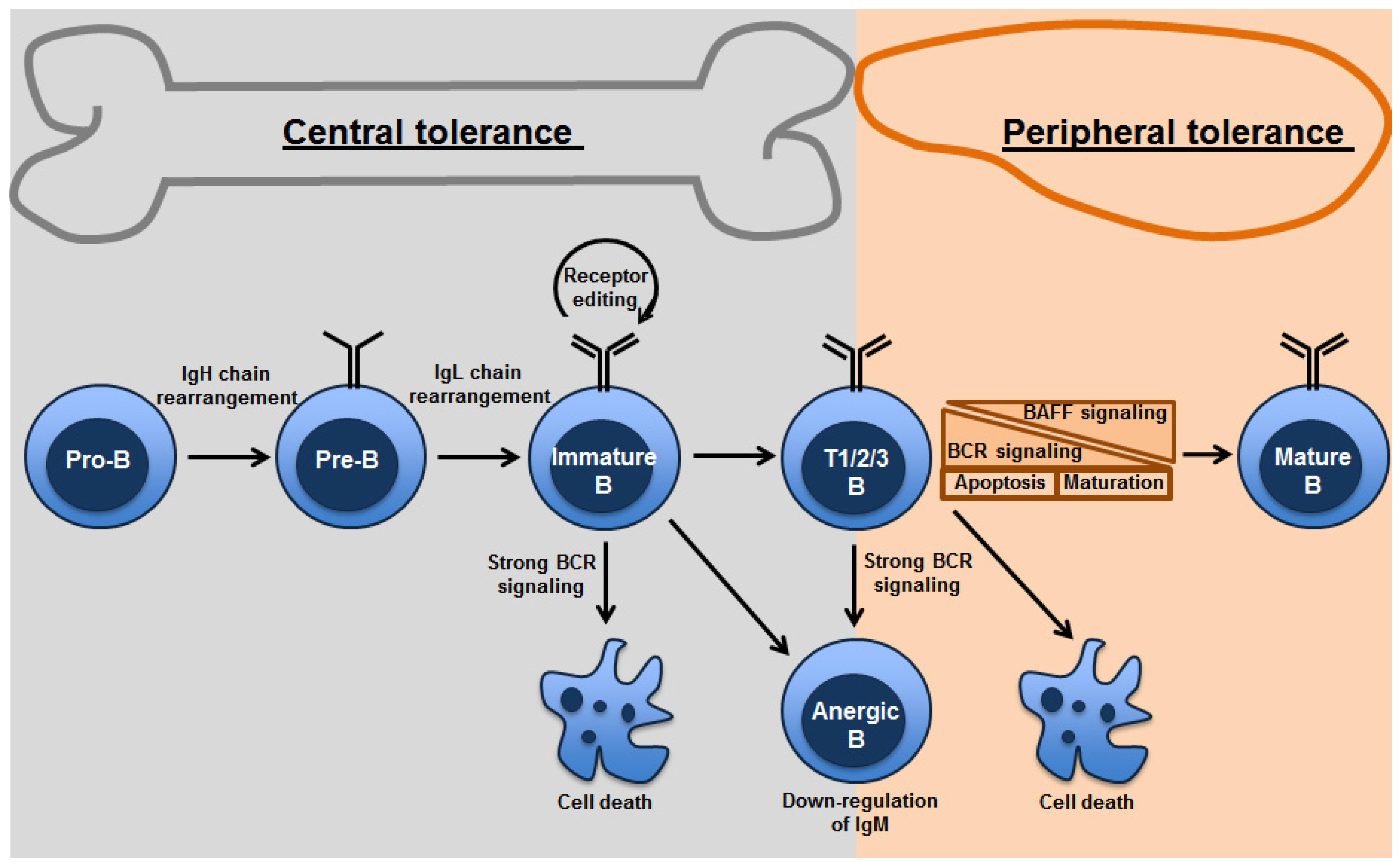{kind=link}
1. Introduction
2. Central Tolerance: Clonal Deletion, Anergy and Receptor Editing
3. Peripheral Tolerance – Role of BAFF (BLyS)
4. Peripheral Tolerance - Role of Toll-Like Receptors
5. Role of Dendritic Cells in Central and Peripheral Tolerance
6. Human B Cell Tolerance and Its Impact on Human Autoimmune Diseases
7. Targeting B Cell Anergy for Therapy
8. Conclusions
Acknowledgements
Author Contributions
Conflicts of interest
References
- Tze, L.E.; Schram, B.R.; Lam, K.P.; Hogquist, K.A.; Hippen, K.L.; Liu, J.; Shinton, S.A.; Otipoby, K.L.; Rodine, P.R.; Vegoe, A.L.; et al. Basal immunoglobulin signaling actively maintains developmental stage in immature B cells. PLoS Biol. 2005, 3, e82. [Google Scholar] [CrossRef]
- Monroe, J.G.; Bannish, G.; Fuentes-Panana, E.M.; King, L.B.; Sandel, P.C.; Chung, J.; Sater, R. Positive and negative selection during B lymphocyte development. Immunol. Res. 2003, 27, 427–442. [Google Scholar] [CrossRef]
- Healy, J.I.; Goodnow, C.C. Positive versus negative signaling by lymphocyte antigen receptors. Annu. Rev. Immunol. 1998, 16, 645–670. [Google Scholar] [CrossRef]
- Allman, D.; Lindsley, R.C.; DeMuth, W.; Rudd, K.; Shinton, S.A.; Hardy, R.R. Resolution of three nonproliferative immature splenic B cell subsets reveals multiple selection points during peripheral B cell maturation. J. Immunol. 2001, 167, 6834–6840. [Google Scholar]
- Allman, D.M.; Ferguson, S.E.; Lentz, V.M.; Cancro, M.P. Peripheral B cell maturation. Ii. Heat-stable antigen(hi) splenic B cells are an immature developmental intermediate in the production of long-lived marrow-derived B cells. J. Immunol. 1993, 151, 4431–4444. [Google Scholar]
- Hardy, R.R.; Hayakawa, K. Positive and negative selection of natural autoreactive B cells. Adv. Exp. Med. Biol. 2012, 750, 227–238. [Google Scholar] [CrossRef]
- Hayakawa, K.; Asano, M.; Shinton, S.A.; Gui, M.; Allman, D.; Stewart, C.L.; Silver, J.; Hardy, R.R. Positive selection of natural autoreactive B cells. Science 1999, 285, 113–116. [Google Scholar] [CrossRef]
- Wardemann, H.; Yurasov, S.; Schaefer, A.; Young, J.W.; Meffre, E.; Nussenzweig, M.C. Predominant autoantibody production by early human B cell precursors. Science 2003, 301, 1374–1377. [Google Scholar] [CrossRef]
- Burnet, F.M. A modification of jerne’s theory of antibody production using the concept of clonal selection. CA Cancer J. Clin. 1976, 26, 119–121. [Google Scholar] [CrossRef]
- Mahana, W.; Guilbert, B.; Avrameas, S. Suppression of anti-DNA antibody production in MRL mice by treatment with anti-idiotypic antibodies. Clin. Exp. Immunol. 1987, 70, 538–545. [Google Scholar]
- Rajewsky, K. From a dream to reality. Eur J. Immunol. 2007, 37 (Suppl. 1), S134–S137. [Google Scholar] [CrossRef]
- Allison, A.C.; Denman, A.M. Self-tolerance and autoimmunity. Br. Med. Bull. 1976, 32, 124–129. [Google Scholar]
- Coutinho, A.; Moller, G. Editorial: Immune activation of B cells: Evidence for ‘one nonspecific triggering signal’ not delivered by the ig receptors. Scand. J. Immunol. 1974, 3, 133–146. [Google Scholar] [CrossRef]
- Taussig, M.J. Induction of hapten-specific B cell tolerance by low doses of hapten-carrier conjugate. Nature 1973, 245, 34–36. [Google Scholar] [CrossRef]
- Chiller, J.M.; Romball, C.G.; Weigle, W.O. Induction of immunological tolerance in neonatal and adult rabbits. Differences in the cellular events. Cell. Immunol. 1973, 8, 28–39. [Google Scholar] [CrossRef]
- Siskind, G.W.; Benacerraf, B. Cell selection by antigen in the immune response. Adv. Immunol. 1969, 10, 1–50. [Google Scholar] [CrossRef]
- Denman, A.M.; Frenkel, E.P. Mode of action of anti-lymphocyte globulin. Ii. Changes in the lymphoid cell population in rats treated with anti-lymphocyte globulin. Immunology 1968, 14, 115–126. [Google Scholar]
- Nemazee, D.; Kouskoff, V.; Hertz, M.; Lang, J.; Melamed, D.; Pape, K.; Retter, M. B-cell-receptor-dependent positive and negative selection in immature B cells. Curr. Top. Microbiol. Immunol. 2000, 245, 57–71. [Google Scholar]
- Radic, M.Z.; Erikson, J.; Litwin, S.; Weigert, M. B lymphocytes may escape tolerance by revising their antigen receptors. J. Exp. Med. 1993, 177, 1165–1173. [Google Scholar] [CrossRef]
- Tiegs, S.L.; Russell, D.M.; Nemazee, D. Receptor editing in self-reactive bone marrow B cells. J. Exp. Med. 1993, 177, 1009–1020. [Google Scholar] [CrossRef]
- Luning Prak, E.T.; Monestier, M.; Eisenberg, R.A. B cell receptor editing in tolerance and autoimmunity. Ann. N. Y. Acad. Sci. 2011, 1217, 96–121. [Google Scholar]
- Forster, I.; Rajewsky, K. The bulk of the peripheral B-cell pool in mice is stable and not rapidly renewed from the bone marrow. Proc. Natl. Acad. Sci. USA 1990, 87, 4781–4784. [Google Scholar] [CrossRef]
- Grandien, A.; Fucs, R.; Nobrega, A.; Andersson, J.; Coutinho, A. Negative selection of multireactive B cell clones in normal adult mice. Eur. J. Immunol. 1994, 24, 1345–1352. [Google Scholar] [CrossRef]
- Wen, L.; Brill-Dashoff, J.; Shinton, S.A.; Asano, M.; Hardy, R.R.; Hayakawa, K. Evidence of marginal-zone B cell-positive selection in spleen. Immunity 2005, 23, 297–308. [Google Scholar] [CrossRef]
- Miller, J.P.; Stadanlick, J.E.; Cancro, M.P. Space, Selection, and surveillance: Setting boundaries with BLyS. J. Immunol. 2006, 176, 6405–6410. [Google Scholar]
- Smith, S.H.; Reth, M. Perspectives on the nature of BCR-mediated survival signals. Mol. Cell 2004, 14, 696–697. [Google Scholar] [CrossRef]
- Scholz, J.L.; Oropallo, M.A.; Sindhava, V.; Goenka, R.; Cancro, M.P. The role of B lymphocyte stimulator in B cell biology: Implications for the treatment of lupus. Lupus 2013, 22, 350–360. [Google Scholar] [CrossRef]
- Schneider, P.; MacKay, F.; Steiner, V.; Hofmann, K.; Bodmer, J.L.; Holler, N.; Ambrose, C.; Lawton, P.; Bixler, S.; Acha-Orbea, H.; et al. BAFF, a novel ligand of the tumor necrosis factor family, stimulates B cell growth. J. Exp. Med. 1999, 189, 1747–1756. [Google Scholar] [CrossRef]
- Moore, P.A.; Belvedere, O.; Orr, A.; Pieri, K.; LaFleur, D.W.; Feng, P.; Soppet, D.; Charters, M.; Gentz, R.; Parmelee, D.; et al. BLyS: Member of the tumor necrosis factor family and B lymphocyte stimulator. Science 1999, 285, 260–263. [Google Scholar] [CrossRef]
- Yamada, T.; Jiang, X.; Kubo, S.; Sakashita, M.; Narita, N.; Yamamoto, H.; Sunaga, H.; Fujieda, S. B type CpG-DNA suppresses poly(I:C)-induced BLyS expression and production in human tonsillar fibroblasts. Clin. Immunol. 2011, 141, 365–371. [Google Scholar] [CrossRef]
- Darce, J.R.; Arendt, B.K.; Wu, X.; Jelinek, D.F. Regulated expression of BAFF-binding receptors during human B cell differentiation. J. Immunol. 2007, 179, 7276–7286. [Google Scholar]
- Zhang, X.; Park, C.S.; Yoon, S.O.; Li, L.; Hsu, Y.M.; Ambrose, C.; Choi, Y.S. BAFF supports human B cell differentiation in the lymphoid follicles through distinct receptors. Int. Immunol. 2005, 17, 779–788. [Google Scholar] [CrossRef]
- Ji, F.; Chen, R.; Liu, B.; Zhang, X.; Han, J.; Wang, H.; Shen, G.; Tao, J. BAFF induces spleen CD4+ T cell proliferation by down-regulating phosphorylation of Foxo3a and activates cyclin D2 and D3 expression. Biochem. Biophys. Res. Commun. 2012, 425, 854–858. [Google Scholar] [CrossRef]
- Wen, L.; Chen, S.J.; Zhang, W.; Ma, H.W.; Zhang, S.Q.; Chen, L. hsBAFF regulates proliferation and response in cultured CD4(+) T lymphocytes by upregulation of intracellular free Ca(2+) homeostasis. Cytokine 2011, 53, 215–222. [Google Scholar] [CrossRef]
- Lavie, F.; Miceli-Richard, C.; Ittah, M.; Sellam, J.; Gottenberg, J.E.; Mariette, X. B-cell activating factor of the tumour necrosis factor family expression in blood monocytes and T cells from patients with primary sjogren’s syndrome. Scand. J. Immunol. 2008, 67, 185–192. [Google Scholar] [CrossRef]
- Yoshimoto, K.; Takahashi, Y.; Ogasawara, M.; Setoyama, Y.; Suzuki, K.; Tsuzaka, K.; Abe, T.; Takeuchi, T. Aberrant expression of BAFF in T cells of systemic lupus erythematosus, which is recapitulated by a human T cell line, Loucy. Int. Immunol. 2006, 18, 1189–1196. [Google Scholar] [CrossRef]
- Gross, J.A.; Dillon, S.R.; Mudri, S.; Johnston, J.; Littau, A.; Roque, R.; Rixon, M.; Schou, O.; Foley, K.P.; Haugen, H.; et al. TACI-Ig neutralizes molecules critical for B cell development and autoimmune disease. Impaired B cell maturation in mice lacking BLyS. Immunity 2001, 15, 289–302. [Google Scholar] [CrossRef]
- Schiemann, B.; Gommerman, J.L.; Vora, K.; Cachero, T.G.; Shulga-Morskaya, S.; Dobles, M.; Frew, E.; Scott, M.L. An essential role for BAFF in the normal development of B cells through a BCMA-independent pathway. Science 2001, 293, 2111–2114. [Google Scholar] [CrossRef]
- Hondowicz, B.D.; Alexander, S.T.; Quinn, W.J., 3rd; Pagan, A.J.; Metzgar, M.H.; Cancro, M.P.; Erikson, J. The role of BLyS/BLyS receptors in anti-chromatin B cell regulation. Int. Immunol. 2007, 19, 465–475. [Google Scholar] [CrossRef]
- Lesley, R.; Xu, Y.; Kalled, S.L.; Hess, D.M.; Schwab, S.R.; Shu, H.B.; Cyster, J.G. Reduced competitiveness of autoantigen-engaged B cells due to increased dependence on BAFF. Immunity 2004, 20, 441–453. [Google Scholar] [CrossRef]
- Cheema, G.S.; Roschke, V.; Hilbert, D.M.; Stohl, W. Elevated serum B lymphocyte stimulator levels in patients with systemic immune-based rheumatic diseases. Arthritis Rheum. 2001, 44, 1313–1319. [Google Scholar] [CrossRef]
- Mariette, X.; Roux, S.; Zhang, J.; Bengoufa, D.; Lavie, F.; Zhou, T.; Kimberly, R. The level of BLyS (BAFF) correlates with the titre of autoantibodies in human sjogren’s syndrome. Ann. Rheum. Dis. 2003, 62, 168–171. [Google Scholar] [CrossRef]
- Stohl, W.; Metyas, S.; Tan, S.M.; Cheema, G.S.; Oamar, B.; Xu, D.; Roschke, V.; Wu, Y.; Baker, K.P.; Hilbert, D.M. B lymphocyte stimulator overexpression in patients with systemic lupus erythematosus: Longitudinal observations. Arthritis Rheum. 2003, 48, 3475–3486. [Google Scholar] [CrossRef]
- Zhang, J.; Roschke, V.; Baker, K.P.; Wang, Z.; Alarcon, G.S.; Fessler, B.J.; Bastian, H.; Kimberly, R.P.; Zhou, T. Cutting edge: A role for B lymphocyte stimulator in systemic lupus erythematosus. J. Immunol. 2001, 166, 6–10. [Google Scholar]
- Lamore, R., 3rd; Parmar, S.; Patel, K.; Hilas, O. Belimumab (Benlysta): A breakthrough therapy for systemic lupus erythematosus. Pharm. Ther. 2012, 37, 212–226. [Google Scholar]
- Stohl, W.; Scholz, J.L.; Cancro, M.P. Targeting BLyS in rheumatic disease: The sometimes-bumpy road from bench to bedside. Curr. Opin. Rheumatol. 2011, 23, 305–310. [Google Scholar] [CrossRef]
- Stohl, W.; Hiepe, F.; Latinis, K.M.; Thomas, M.; Scheinberg, M.A.; Clarke, A.; Aranow, C.; Wellborne, F.R.; Abud-Mendoza, C.; Hough, D.R.; et al. Belimumab reduces autoantibodies, normalizes low complement levels, and reduces select B cell populations in patients with systemic lupus erythematosus. Arthritis Rheum. 2012, 64, 2328–2337. [Google Scholar] [CrossRef]
- Coca, A.; Sanz, I. Updates on B-cell immunotherapies for systemic lupus erythematosus and Sjogren’s syndrome. Curr. Opin Rheumatol. 2012, 24, 451–456. [Google Scholar]
- Stohl, W.; Merrill, J.T.; McKay, J.D.; Lisse, J.R.; Zhong, Z.J.; Freimuth, W.W.; Genovese, M.C. Efficacy and safety of Belimumab in patients with rheumatoid arthritis: A phase ii, randomized, double-blind, placebo-controlled, dose-ranging study. J. Rheumatol. 2013, 40, 579–589. [Google Scholar] [CrossRef]
- Wallace, D.J.; Stohl, W.; Furie, R.A.; Lisse, J.R.; McKay, J.D.; Merrill, J.T.; Petri, M.A.; Ginzler, E.M.; Chatham, W.W.; McCune, W.J.; et al. A phase II, randomized, double-blind, placebo-controlled, dose-ranging study of belimumab in patients with active systemic lupus erythematosus. Arthritis Rheum. 2009, 61, 1168–1178. [Google Scholar] [CrossRef]
- Medzhitov, R.; Janeway, C. The Toll receptor family and microbial recognition. Trends Microbiol. 2000, 8, 452–456. [Google Scholar]
- Medzhitov, R.; PrestonHurlburt, P.; Janeway, C.A. A human homologue of the drosophila toll protein signals activation of adaptive immunity. Nature 1997, 388, 394–397. [Google Scholar] [CrossRef]
- Steinman, R.M.; Hemmi, H. Dendritic cells: Translating innate to adaptive immunity. Curr. Top. Microbiol. Immunol. 2006, 311, 17–58. [Google Scholar]
- Sindhava, V.J.; Bondada, S. Multiple regulatory mechanisms control B-1 B cell activation. Front. Immunol. 2012, 3. [Google Scholar] [CrossRef]
- Gururajan, M.; Jacob, J.; Pulendran, B. Toll-like receptor expression and responsiveness of distinct murine splenic and mucosal B-cell subsets. PLoS One 2007, 2, e863. [Google Scholar] [CrossRef]
- Lau, C.M.; Broughton, C.; Tabor, A.S.; Akira, S.; Flavell, R.A.; Mamula, M.J.; Christensen, S.R.; Shlomchik, M.J.; Viglianti, G.A.; Rifkin, I.R.; et al. Rna-associated autoantigens activate B cells by combined B cell antigen receptor/Toll-like receptor 7 engagement. J. Exp. Med. 2005, 202, 1171–1177. [Google Scholar] [CrossRef]
- Leadbetter, E.A.; Rifkin, I.R.; Marshak-Rothstein, A. Toll-like receptors and activation of autoreactive B cells. Curr. Dir. Autoimmun. 2003, 6, 105–122. [Google Scholar]
- Leadbetter, E.A.; Rifkin, I.R.; Hohlbaum, A.M.; Beaudette, B.C.; Shlomchik, M.J.; Marshak-Rothstein, A. Chromatin-IgG complexes activate B cells by dual engagement of IgM and Toll-like receptors. Nature 2002, 416, 603–607. [Google Scholar] [CrossRef]
- Celhar, T.; Magalhaes, R.; Fairhurst, A.M. TLR7 and TLR9 in SLE: When sensing self goes wrong. Immunol. Res. 2012, 53, 58–77. [Google Scholar] [CrossRef]
- Herlands, R.A.; Christensen, S.R.; Sweet, R.A.; Hershberg, U.; Shlomchik, M.J. T cell-independent and toll-like receptor-dependent antigen-driven activation of autoreactive B cells. Immunity 2008, 29, 249–260. [Google Scholar] [CrossRef]
- Finkelman, F.D.; Lees, A.; Birnbaum, R.; Gause, W.C.; Morris, S.C. Dendritic cells can present antigen in vivo in a tolerogenic or immunogenic fashion. J. Immunol. 1996, 157, 1406–1414. [Google Scholar]
- Rotta, G.; Edwards, E.W.; Sangaletti, S.; Bennett, C.; Ronzoni, S.; Colombo, M.P.; Steinman, R.M.; Randolph, G.J.; Rescigno, M. Lipopolysaccharide or whole bacteria block the conversion of inflammatory monocytes into dendritic cells in vivo. J. Exp. Med. 2003, 198, 1253–1263. [Google Scholar] [CrossRef]
- Tisch, R. Immunogenic versus tolerogenic dendritic cells: A matter of maturation. Int. Rev. Immunol. 2010, 29, 111–118. [Google Scholar] [CrossRef]
- Sindhava, V.J.; Tuna, H.; Gachuki, B.W.; DiLillo, D.J.; Avdiushko, M.G.; Onami, T.M.; Tedder, T.F.; Cohen, D.A.; Bondada, S. Bone marrow dendritic cell-mediated regulation of TLR and B cell receptor signaling in B cells. J. Immunol. 2012, 189, 3355–3367. [Google Scholar] [CrossRef]
- Santos, L.; Draves, K.E.; Boton, M.; Grewal, P.K.; Marth, J.D.; Clark, E.A. Dendritic cell-dependent inhibition of B cell proliferation requires CD22. J. Immunol. 2008, 180, 4561–4569. [Google Scholar]
- Kaisho, T.; Akira, S. Dendritic-cell function in Toll-like receptor- and MyD88-knockout mice. Trends Immunol. 2001, 22, 78–83. [Google Scholar] [CrossRef]
- Jenner, J.; Kerst, G.; Handgretinger, R.; Muller, I. Increased alpha2,6-sialylation of surface proteins on tolerogenic, immature dendritic cells and regulatory T cells. Exp. Hematol. 2006, 34, 1212–1218. [Google Scholar]
- Duong, B.H.; Tian, H.; Ota, T.; Completo, G.; Han, S.; Vela, J.L.; Ota, M.; Kubitz, M.; Bovin, N.; Paulson, J.C.; et al. Decoration of T-independent antigen with ligands for CD22 and SIGLEC-G can suppress immunity and induce B cell tolerance in vivo. J. Exp. Med. 2010, 207, 173–187. [Google Scholar] [CrossRef]
- Kilmon, M.A.; Rutan, J.A.; Clarke, S.H.; Vilen, B.J. Low-affinity, smith antigen-specific B cells are tolerized by dendritic cells and macrophages. J. Immunol. 2005, 175, 37–41. [Google Scholar]
- Jellusova, J.; Nitschke, L. Regulation of B cell functions by the sialic acid-binding receptors SIGLEC-G and CD22. Front. Immunol. 2011, 2. [Google Scholar] [CrossRef]
- Jellusova, J.; Wellmann, U.; Amann, K.; Winkler, T.H.; Nitschke, L. CD22 x SIGLEC-G double-deficient mice have massively increased B1 cell numbers and develop systemic autoimmunity. J. Immunol. 2010, 184, 3618–3627. [Google Scholar] [CrossRef]
- McDonnell, M.; Liang, Y.; Noronha, A.; Coukos, J.; Kasper, D.L.; Farraye, F.A.; Ganley-Leal, L.M. Systemic Toll-like receptor ligands modify B-cell responses in human inflammatory bowel disease. Inflamm. Bowel Dis. 2011, 17, 298–307. [Google Scholar] [CrossRef]
- Jagannathan, M.; McDonnell, M.; Liang, Y.; Hasturk, H.; Hetzel, J.; Rubin, D.; Kantarci, A.; van Dyke, T.E.; Ganley-Leal, L.M.; Nikolajczyk, B.S. Toll-like receptors regulate B cell cytokine production in patients with diabetes. Diabetologia 2010, 53, 1461–1471. [Google Scholar] [CrossRef]
- Jube, S.; Rivera, Z.S.; Bianchi, M.E.; Powers, A.; Wang, E.; Pagano, I.; Pass, H.I.; Gaudino, G.; Carbone, M.; Yang, H. Cancer cell secretion of the damp protein hmgb1 supports progression in malignant mesothelioma. Cancer Res. 2012, 72, 3290–3301. [Google Scholar] [CrossRef]
- Munoz, L.E.; Lauber, K.; Schiller, M.; Manfredi, A.A.; Herrmann, M. The role of defective clearance of apoptotic cells in systemic autoimmunity. Nat. Rev. Rheumatol. 2010, 6, 280–289. [Google Scholar]
- Menard, L.; Samuels, J.; Ng, Y.S.; Meffre, E. Inflammation-independent defective early B cell tolerance checkpoints in rheumatoid arthritis. Arthritis Rheum. 2011, 63, 1237–1245. [Google Scholar] [CrossRef]
- Cotsapas, C.; Voight, B.F.; Rossin, E.; Lage, K.; Neale, B.M.; Wallace, C.; Abecasis, G.R.; Barrett, J.C.; Behrens, T.; Cho, J.; et al. Pervasive sharing of genetic effects in autoimmune disease. PLoS Genet. 2011, 7, e1002254. [Google Scholar] [CrossRef]
- Edwards, J.C.; Szczepanski, L.; Szechinski, J.; Filipowicz-Sosnowska, A.; Emery, P.; Close, D.R.; Stevens, R.M.; Shaw, T. Efficacy of B-cell-targeted therapy with rituximab in patients with rheumatoid arthritis. N. Engl. J. Med. 2004, 350, 2572–2581. [Google Scholar] [CrossRef]
- Kinnunen, T.; Chamberlain, N.; Morbach, H.; Cantaert, T.; Lynch, M.; Preston-Hurlburt, P.; Herold, K.C.; Hafler, D.A.; O’Connor, K.C.; Meffre, E. Specific peripheral B cell tolerance defects in patients with multiple sclerosis. J. Clin. Invest. 2013, 123, 2737–2741. [Google Scholar]
- Arimura, Y.; Yagi, J. Comprehensive expression profiles of genes for protein tyrosine phosphatases in immune cells. Sci. Signal. 2010, 3, rs1. [Google Scholar]
- Menard, L.; Saadoun, D.; Isnardi, I.; Ng, Y.S.; Meyers, G.; Massad, C.; Price, C.; Abraham, C.; Motaghedi, R.; Buckner, J.H.; et al. The PTPN22 allele encoding an R620W variant interferes with the removal of developing autoreactive B cells in humans. J. Clin. Invest. 2011, 121, 3635–3644. [Google Scholar] [CrossRef]
- Apollonio, B.; Scielzo, C.; Bertilaccio, M.T.; Ten Hacken, E.; Scarfo, L.; Ranghetti, P.; Stevenson, F.; Packham, G.; Ghia, P.; Muzio, M.; et al. Targeting B-cell anergy in chronic lymphocytic leukemia. Blood 2013, 121, 3879–3888. [Google Scholar] [CrossRef]
- Woyach, J.A. Survival of the weak (signalers): Anergy in CLL. Blood 2013, 121, 3781–3783. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Gururajan, M.; Sindhava, V.J.; Bondada, S. B Cell Tolerance in Health and Disease. Antibodies 2014, 3, 116-129. https://doi.org/10.3390/antib3010116
Gururajan M, Sindhava VJ, Bondada S. B Cell Tolerance in Health and Disease. Antibodies. 2014; 3(1):116-129. https://doi.org/10.3390/antib3010116
Chicago/Turabian StyleGururajan, Murali, Vishal J. Sindhava, and Subbarao Bondada. 2014. "B Cell Tolerance in Health and Disease" Antibodies 3, no. 1: 116-129. https://doi.org/10.3390/antib3010116
APA StyleGururajan, M., Sindhava, V. J., & Bondada, S. (2014). B Cell Tolerance in Health and Disease. Antibodies, 3(1), 116-129. https://doi.org/10.3390/antib3010116