


Response of Water-Use Efficiency (WUE) in Alpine Grasslands to Hydrothermal and Radiative Factors Across Elevation Gradients

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Data Source
2.3. Methods
2.3.1. The Calculations of VPD
2.3.2. The Calculations of WUE
2.3.3. Partial Correlation Analysis
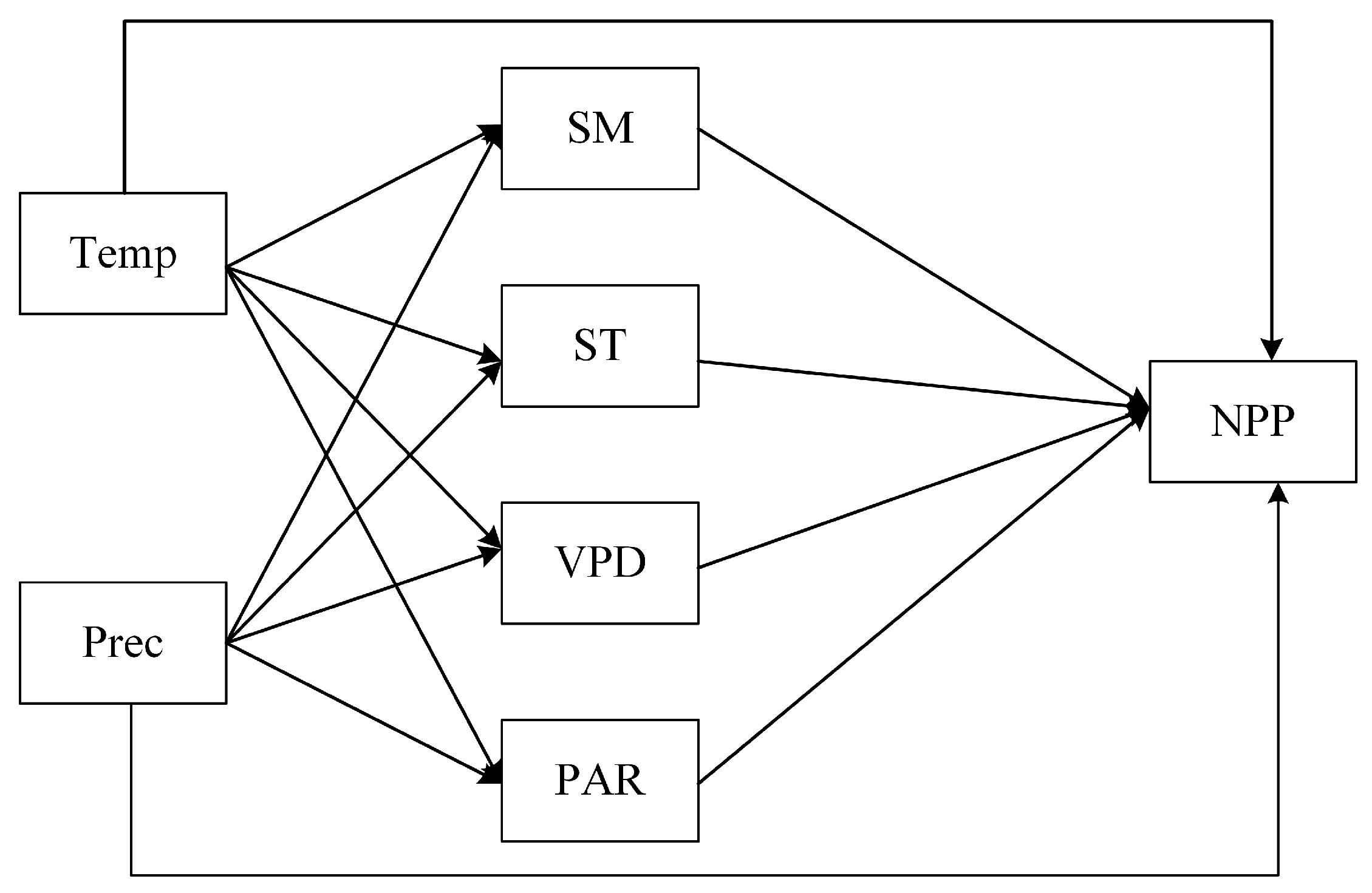
2.3.4. Structural Equation Modeling (SEM)
2.3.5. Contribution Rate Calculation
3. Results
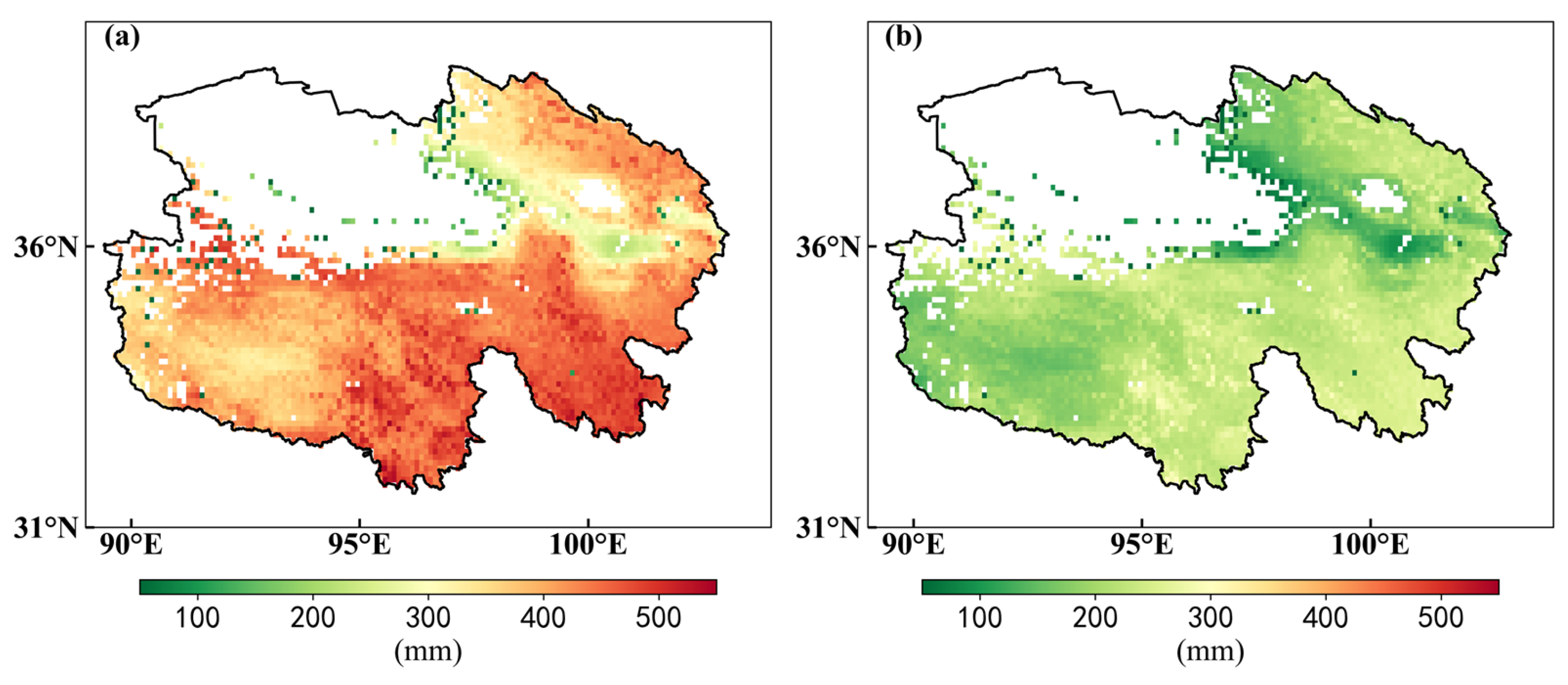
3.1. Changes in Hydrothermal and Radiation Factors
3.2. Spatial and Temporal Characteristics
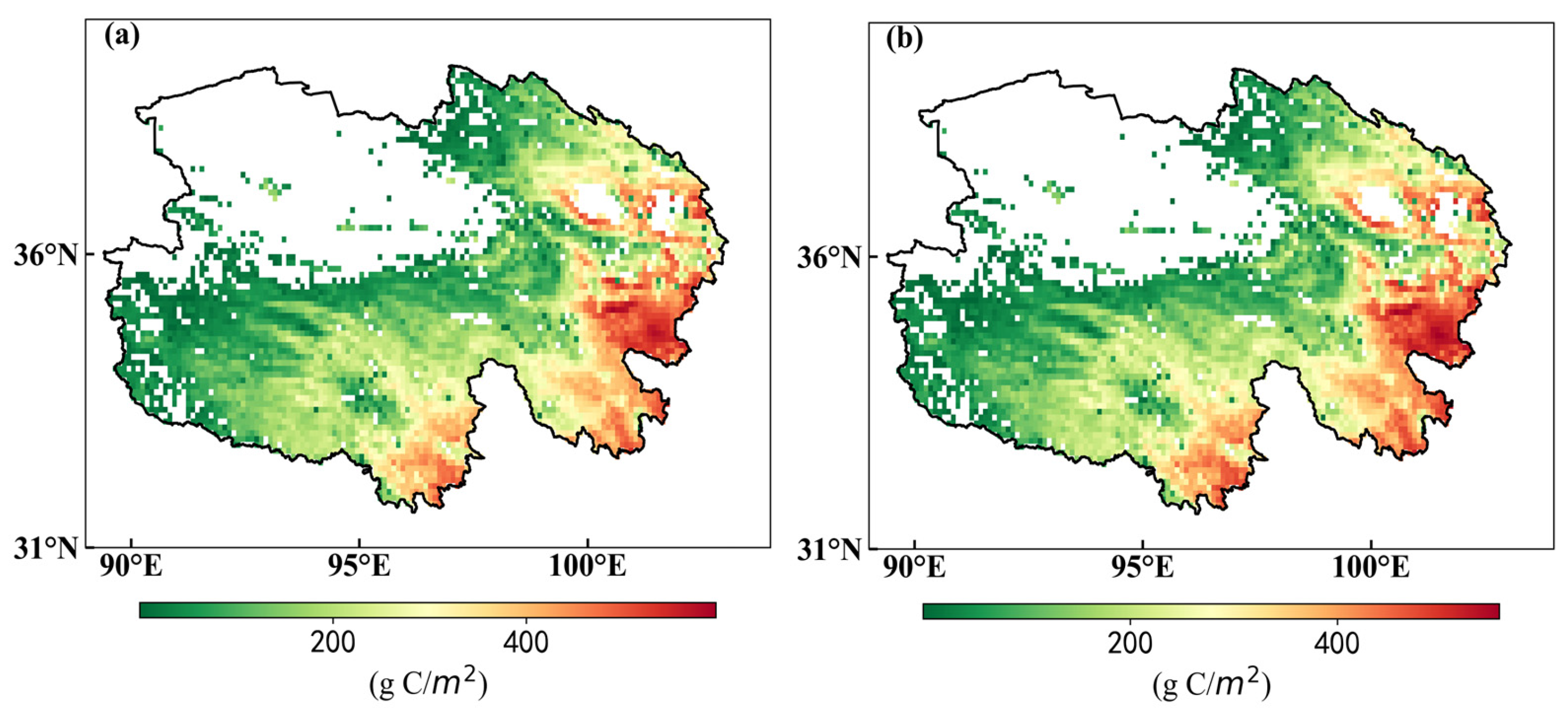
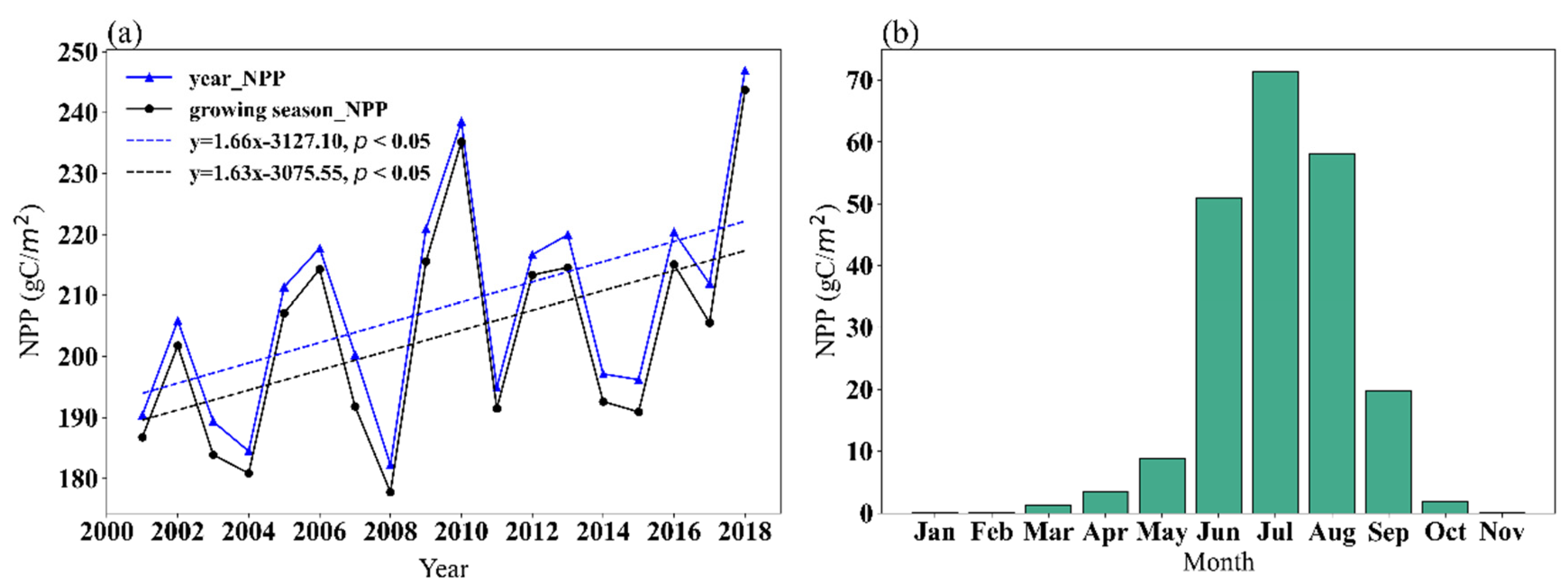
3.2.1. Spatial and Temporal Characteristics of NPP
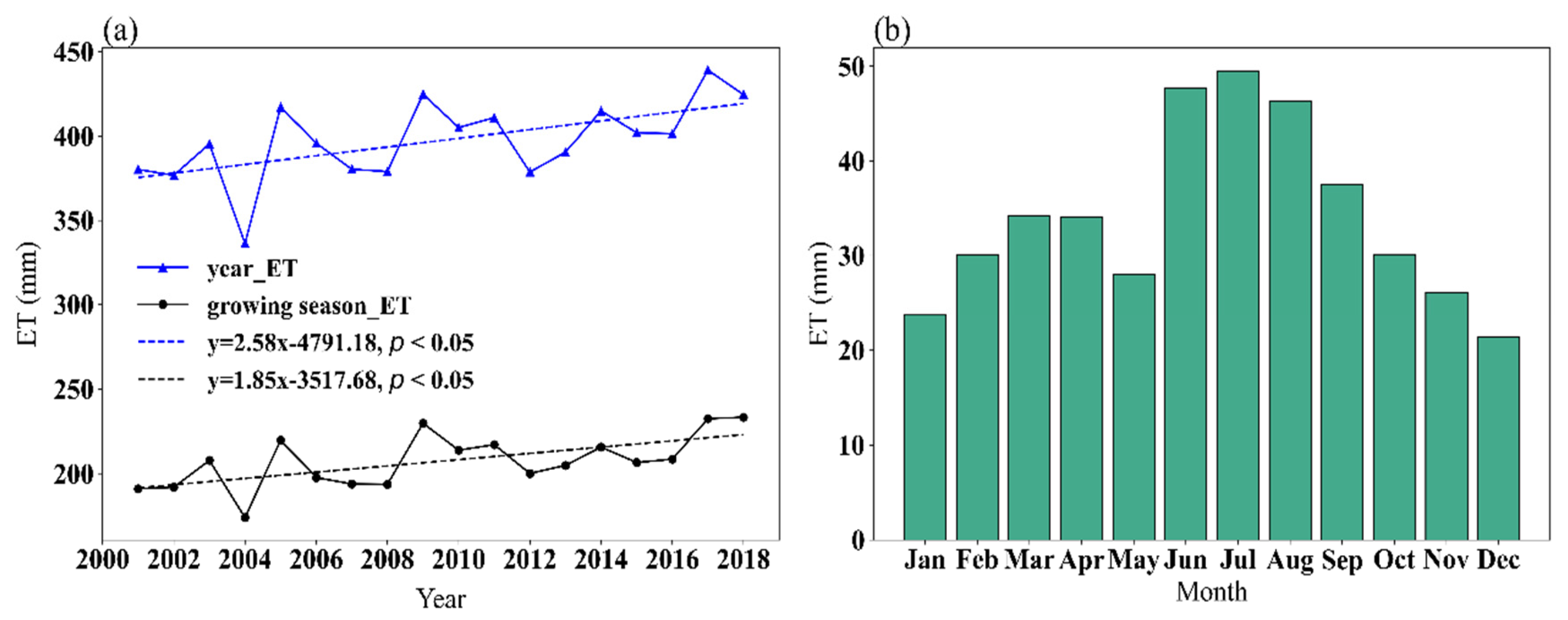
3.2.2. Spatial and Temporal Characteristics of ET
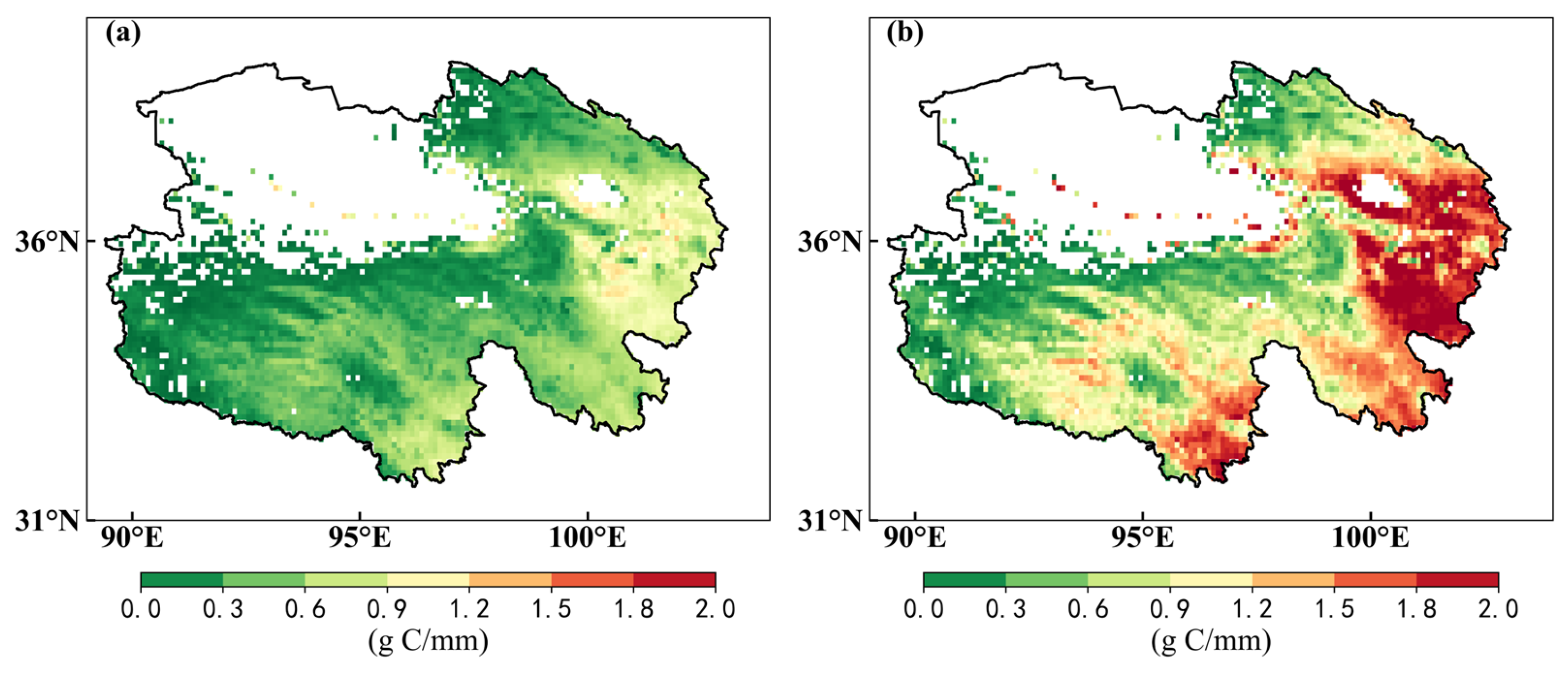
3.2.3. Spatial and Temporal Characteristics of WUE
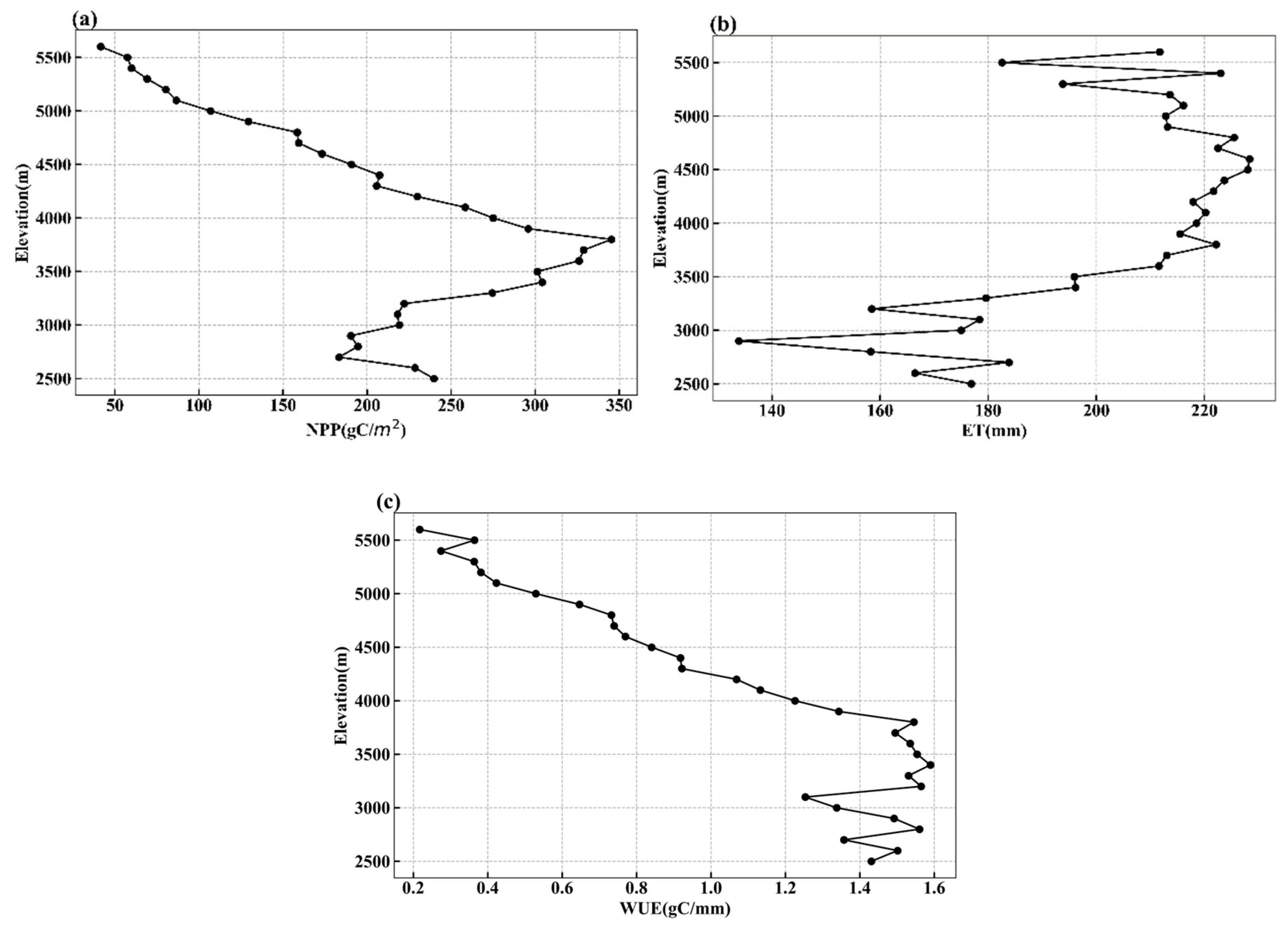
3.3. Variation Characteristics Along Elevation Gradient
3.4. Response Mechanism Analysis
3.4.1. Results of Partial Correlation Analysis
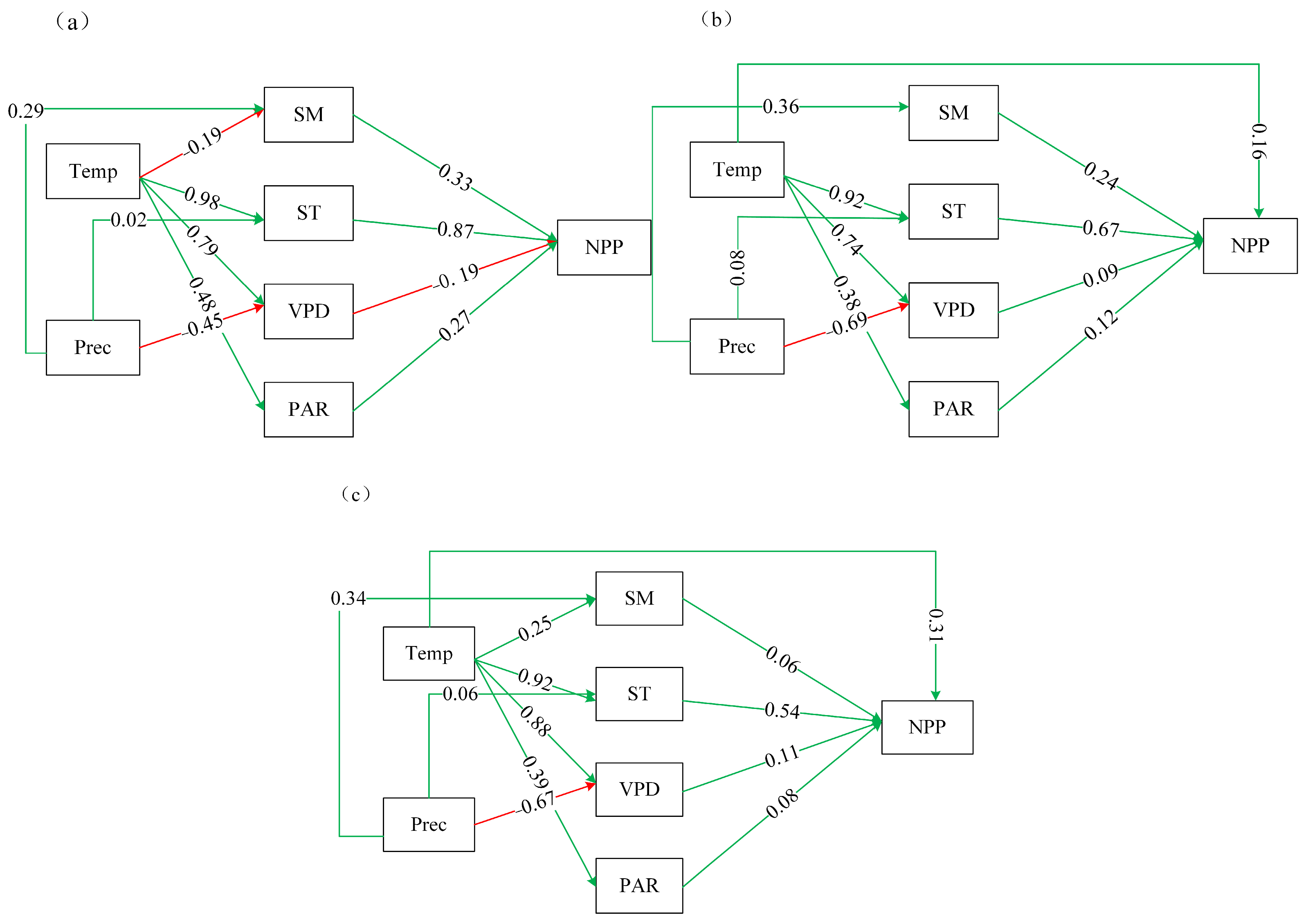
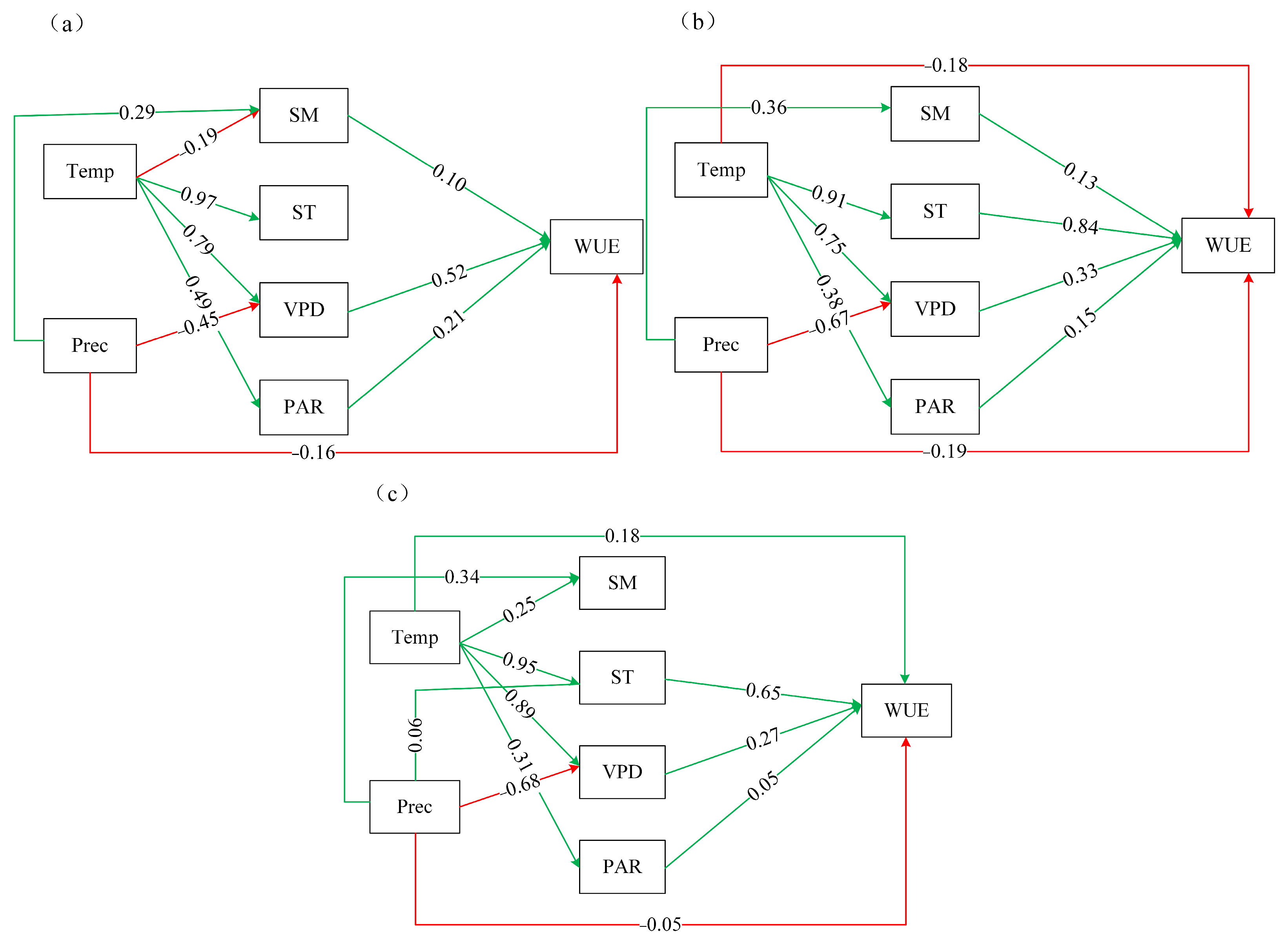
3.4.2. Results of Structural Equation Modeling
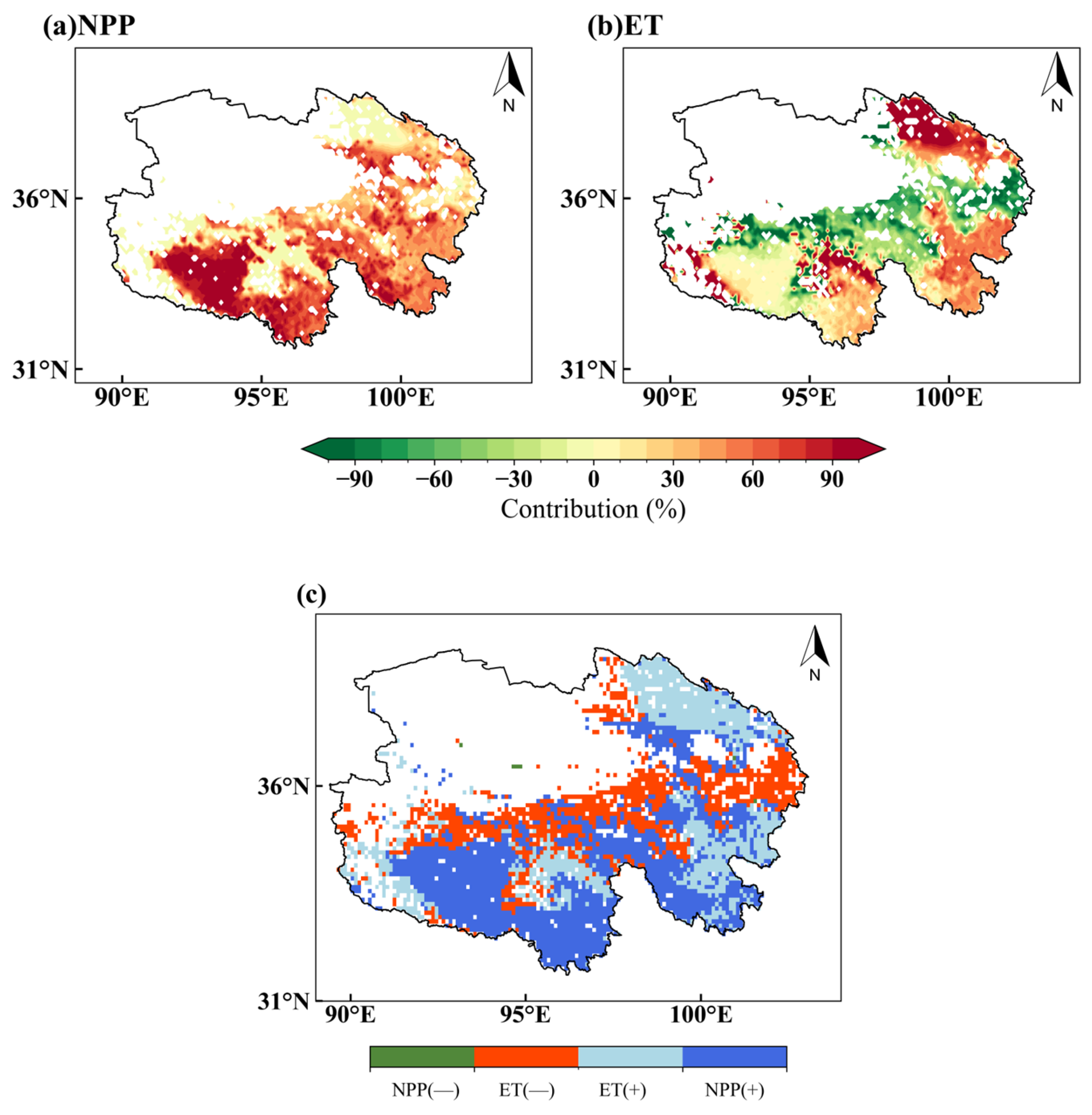
3.4.3. Results of Contribution Rate Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Lin, S.; Wang, G.; Feng, J.; Dan, L.; Sun, X.; Hu, Z.; Chen, X.; Xiao, X. A Carbon Flux Assessment Driven by Environmental Factors Over the Tibetan Plateau and Various Permafrost Regions. J. Geophys. Res. Biogeosci. 2019, 124, 1132–1147. [Google Scholar] [CrossRef]
- Du, Q.; Sun, Y.; Guan, Q.; Pan, N.; Wang, Q.; Ma, Y.; Li, H.; Liang, L. Vulnerability of Grassland Ecosystems to Climate Change in the Qilian Mountains, Northwest China. J. Hydrol. 2022, 612, 128305. [Google Scholar] [CrossRef]
- Jin, J.; Wang, Y.; Zhang, Z.; Magliulo, V.; Jiang, H.; Cheng, M. Phenology Plays an Important Role in the Regulation of Terrestrial Ecosystem Water-Use Efficiency in the Northern Hemisphere. Remote Sens. 2017, 9, 664. [Google Scholar] [CrossRef]
- Shen, M.; Tang, Y.; Chen, J.; Zhu, X.; Zheng, Y. Influences of Temperature and Precipitation before the Growing Season on Spring Phenology in Grasslands of the Central and Eastern Qinghai-Tibetan Plateau. Agric. For. Meteorol. 2011, 151, 1711–1722. [Google Scholar] [CrossRef]
- Cuo, L.; Zhang, Y.; Ri, X.; Zhou, B. Decadal Change and Inter-Annual Variability of Net Primary Productivity on the Tibetan Plateau. Clim. Dyn. 2021, 56, 1837–1857. [Google Scholar] [CrossRef]
- Gao, Q.; Zhu, W.; Schwartz, M.W.; Ganjurjav, H.; Wan, Y.; Qin, X.; Ma, X.; Williamson, M.A.; Li, Y. Climatic Change Controls Productivity Variation in Global Grasslands. Sci. Rep. 2016, 6, 26958. [Google Scholar] [CrossRef]
- Yisilayili, G.; He, B.; Song, Y.; Luo, X.; Yang, W.; Chen, Y. Simulation of Vegetation NPP in Typical Arid Regions Based on the CASA Model and Quantification of Its Driving Factors. Land 2025, 14, 371. [Google Scholar] [CrossRef]
- Yan, D.; Yang, Y.; Hao, H.; Zhu, J.; Fu, Y.; Meng, N.; Li, Z.; Dai, X.; Li, R.; Zheng, H. Quantifying the Variation in Water Use Efficiency across Climates and Biomes on the Qinghai-Tibetan Plateau. Ecol. Indic. 2023, 157, 111274. [Google Scholar] [CrossRef]
- Khalifa, M.; Elagib, N.A.; Ribbe, L.; Schneider, K. Spatio-Temporal Variations in Climate, Primary Productivity and Efficiency of Water and Carbon Use of the Land Cover Types in Sudan and Ethiopia. Sci. Total Environ. 2018, 624, 790–806. [Google Scholar] [CrossRef]
- Zhang, X.; Chen, Y.; Zhang, Q.; Xia, Z.; Hao, H.; Xia, Q. Potential Evapotranspiration Determines Changes in the Carbon Sequestration Capacity of Forest and Grass Ecosystems in Xinjiang, Northwest China. Glob. Ecol. Conserv. 2023, 48, e02737. [Google Scholar] [CrossRef]
- USGCRP. 2018: Second State of the Carbon Cycle Report (SOCCR2): A Sustained Assessment Report; Cavallaro, N., Shrestha, G., Birdsey, R., Mayes, M.A., Najjar, R.G., Reed, S.C., Romero-Lankao, P., Zhu, Z., Eds.; U.S. Global Change Research Program: Washington, DC, USA, 2018; p. 878. Available online: https://carbon2018.globalchange.gov/ (accessed on 26 May 2025).
- Yang, S.; Li, Y.; Zhao, Y.; Lan, A.; Zhou, C.; Lu, H.; Zhou, L. Changes in Vegetation Ecosystem Carbon Sinks and Their Response to Drought in the Karst Concentration Distribution Area of Asia. Ecol. Inform. 2024, 84, 102907. [Google Scholar] [CrossRef]
- Tang, X.; Li, H.; Desai, A.R.; Nagy, Z.; Luo, J.; Kolb, T.E.; Olioso, A.; Xu, X.; Yao, L.; Kutsch, W.; et al. How Is Water-Use Efficiency of Terrestrial Ecosystems Distributed and Changing on Earth? Sci. Rep. 2014, 4, 7483. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Liu, H.; Cao, G.; Ma, Z.; Li, Y.; Zhang, F.; Zhao, X.; Zhao, X.; Jiang, L.; Sanders, N.J.; et al. Alpine Grassland Plants Grow Earlier and Faster but Biomass Remains Unchanged over 35 Years of Climate Change. Ecol. Lett. 2020, 23, 701–710. [Google Scholar] [CrossRef]
- Franks, P.J.; Adams, M.A.; Amthor, J.S.; Barbour, M.M.; Berry, J.A.; Ellsworth, D.S.; Farquhar, G.D.; Ghannoum, O.; Lloyd, J.; McDowell, N.; et al. Sensitivity of Plants to Changing Atmospheric CO2 Concentration: From the Geological Past to the next Century. New Phytol. 2013, 197, 1077–1094. [Google Scholar] [CrossRef]
- Qu, Q.; Jian, S.; Chen, A.; Xiao, C. Investigating the Dynamic Change and Driving Force of Vegetation Carbon Sink in Taihang Mountain, China. Land 2024, 13, 1348. [Google Scholar] [CrossRef]
- Chen, N.; Zhang, Y.; Song, C.; Xu, M.; Zhang, T.; Li, M.; Cong, N.; Zu, J.; Zheng, Z.; Ma, G.; et al. The Chained Effects of Earlier Vegetation Activities and Summer Droughts on Ecosystem Productivity on the Tibetan Plateau. Agric. For. Meteorol. 2022, 321, 108975. [Google Scholar] [CrossRef]
- Xi, X.; Yuan, X. Remote Sensing of Atmospheric and Soil Water Stress on Ecosystem Carbon and Water Use during Flash Droughts over Eastern China. Sci. Total Environ. 2023, 868, 161715. [Google Scholar] [CrossRef]
- Wall, S.; Cockram, J.; Vialet-Chabrand, S.; Van Rie, J.; Gallé, A.; Lawson, T. The Impact of Growth at Elevated [CO2] on Stomatal Anatomy and Behavior Differs between Wheat Species and Cultivars. J. Exp. Bot. 2023, 74, 2860–2874. [Google Scholar] [CrossRef]
- Martínez-Goñi, X.S.; Miranda-Apodaca, J.; Pérez-López, U. Enhanced Photosynthesis, Transpiration Regulation, Water Use-Efficiency and Growth in Buckwheat Outperforms Wheat Response to High [CO2], High Temperature and Drought. Environ. Exp. Bot. 2024, 222, 105756. [Google Scholar] [CrossRef]
- Pernicová, N.; Urban, O.; Čáslavský, J.; Kolář, T.; Rybníček, M.; Sochová, I.; Peñuelas, J.; Bošeľa, M.; Trnka, M. Impacts of Elevated CO2 Levels and Temperature on Photosynthesis and Stomatal Closure along an Altitudinal Gradient Are Counteracted by the Rising Atmospheric Vapor Pressure Deficit. Sci. Total Environ. 2024, 921, 171173. [Google Scholar] [CrossRef]
- Zhao, P.; Zeng, X.; Peng, S.; Sun, G. Daily variation of gas exchange, stomatal conductance and water use efficiency in summer leaves of Ormosia pinnata. J. Trop. Subtrop. Bot. 2000, 8, 35–42. [Google Scholar]
- Grossiord, C.; Buckley, T.N.; Cernusak, L.A.; Novick, K.A.; Poulter, B.; Siegwolf, R.T.W.; Sperry, J.S.; McDowell, N.G. Plant Responses to Rising Vapor Pressure Deficit. New Phytol. 2020, 226, 1550–1566. [Google Scholar] [CrossRef] [PubMed]
- Seleiman, M.F.; Al-Suhaibani, N.; Ali, N.; Akmal, M.; Alotaibi, M.; Refay, Y.; Dindaroglu, T.; Abdul-Wajid, H.H.; Battaglia, M.L. Drought Stress Impacts on Plants and Different Approaches to Alleviate Its Adverse Effects. Plants 2021, 10, 259. [Google Scholar] [CrossRef]
- Bai, Y.; Wu, J.; Xing, Q.; Pan, Q.; Huang, J.; Yang, D.; Han, X. Primary Production and Rain Use Efficiency Across a Precipitation Gradient on the Mongolia Plateau. Ecology 2008, 89, 2140–2153. [Google Scholar] [CrossRef]
- Lu, X.; Zhuang, Q. Evaluating Evapotranspiration and Water-Use Efficiency of Terrestrial Ecosystems in the Conterminous United States Using MODIS and AmeriFlux Data. Remote Sens. Environ. 2010, 114, 1924–1939. [Google Scholar] [CrossRef]
- Marenco, R.A.; Nascimento, H.C.S.; Magalhães, N.S. Stomatal Conductance in Amazonian Tree Saplings in Response to Variations in the Physical Environment. Photosynthetica 2014, 52, 493–500. [Google Scholar] [CrossRef]
- Urban, J.; Ingwers, M.; McGuire, M.A.; Teskey, R.O. Stomatal Conductance Increases with Rising Temperature. Plant Signal. Behav. 2017, 12, e1356534. [Google Scholar] [CrossRef]
- Ma, L.; Liu, Y.; Zhao, J. The Relationship between Environmental Change and Stable Carbon Isotopes Records from Tree-Ring in Mt. Helan. Huan Jing Ke Xue = Huanjing Kexue 2003, 24, 49–53. [Google Scholar]
- Sun, Y.; Piao, S.; Huang, M.; Ciais, P.; Zeng, Z.; Cheng, L.; Li, X.; Zhang, X.; Mao, J.; Peng, S.; et al. Global Patterns and Climate Drivers of Water-Use Efficiency in Terrestrial Ecosystems Deduced from Satellite-Based Datasets and Carbon Cycle Models. Glob. Ecol. Biogeogr. 2016, 25, 311–323. [Google Scholar] [CrossRef]
- Xu, X.; Jiao, F.; Gong, H.; Xue, P.; Lin, N.; Liu, J.; Zhang, K.; Qiu, J.; Lin, D.; Yang, Y.; et al. Observed Divergence in the Trends of Temperature Controls on Chinese Ecosystem Water Use Efficiency. Ecol. Indic. 2023, 157, 111241. [Google Scholar] [CrossRef]
- Shao, W.; Guan, Q.; Liu, H.; Sun, W.; Sun, Y. Temporal and Spatial Dynamics of Carbon and Water Use Efficiency on the Qinghai-Tibet Plateau and Their Ecosystem Responses. J. Clean. Prod. 2024, 475, 143581. [Google Scholar] [CrossRef]
- Chen, J.; Pu, J.; Li, J.; Zhang, T. Response of Carbon- and Water-Use Efficiency to Climate Change and Human Activities in China. Ecol. Indic. 2024, 160, 111829. [Google Scholar] [CrossRef]
- Ma, L.; Zhang, Z.; Yao, B.; Ma, Z.; Huang, X.; Zhou, B.; Xu, M.; Guo, J.; Zhou, H. Effects of Drought and Heat on the Productivity and Photosynthetic Characteristics of Alpine Meadow Plants on the Qinghai-Tibetan Plateau. J. Mt. Sci. 2021, 18, 2079–2093. [Google Scholar] [CrossRef]
- Cheng, M.; Jin, J.; Jiang, H. Strong Impacts of Autumn Phenology on Grassland Ecosystem Water Use Efficiency on the Tibetan Plateau. Ecol. Indic. 2021, 126, 107682. [Google Scholar] [CrossRef]
- Wang, Q.; Fan, X.; Wang, M. Recent Warming Amplification over High Elevation Regions across the Globe. Clim. Dyn. 2014, 43, 87–101. [Google Scholar] [CrossRef]
- Lu, S.; Chen, F.; Zhou, J.; Hughes, A.C.; Ma, X.; Gao, W. Cascading Implications of a Single Climate Change Event for Fragile Ecosystems on the Qinghai-Tibetan Plateau. Ecosphere 2020, 11, e03243. [Google Scholar] [CrossRef]
- Zhang, S.; Wu, T.; Guo, L.; Zhao, Y. Assessing Ecological Risk on the Qinghai-Tibet Plateau Based on Future Land Use Scenarios and Ecosystem Service Values. Ecol. Indic. 2023, 154, 110769. [Google Scholar] [CrossRef]
- Zhang, Y.; Bao, G.; Bao, Y.; Yuan, Z.; Rina, W.; Tong, S. Altitudinal Differences in Decreasing Heat Deficit at the End of the Growing Season of Alpine Grassland on the Qinghai–Tibetan Plateau from 1982 to 2022. Land 2025, 14, 758. [Google Scholar] [CrossRef]
- Sun, J.; Zhou, T.-C.; Liu, M.; Chen, Y.-C.; Liu, G.-H.; Xu, M.; Shi, P.-L.; Peng, F.; Tsunekawa, A.; Liu, Y.; et al. Water and Heat Availability Are Drivers of the Aboveground Plant Carbon Accumulation Rate in Alpine Grasslands on the Tibetan Plateau. Glob. Ecol. Biogeogr. 2020, 29, 50–64. [Google Scholar] [CrossRef]
- Liu, M.; Jiao, J.; Pan, J.; Song, J.; Che, Y.; Li, L. Spatial and Temporal Patterns of Planting NPP and Its Driving Factors in Qinghai Province. Acta Ecol. Sin. 2020, 40, 5306–5317. [Google Scholar]
- Li, M.C.; Zhu, J.J.; Li, L.X. Occurrence and Altitudinal Pattern of C4 Plants on Qinghai Plateau, Qinghai Province, China. Photosynthetica 2009, 47, 298–303. [Google Scholar] [CrossRef]
- Zhou, Y.; Fan, J.; Zhong, H.; Zhang, W. Relationships between Altitudinal Gradient and Plant Carbon Isotope Composition of Grassland Communities on the Qinghai-Tibet Plateau, China. Sci. China Earth Sci. 2013, 56, 311–320. [Google Scholar] [CrossRef]
- He, J.; Yang, K.; Tang, W.; Lu, H.; Qin, J.; Chen, Y.; Li, X. The First High-Resolution Meteorological Forcing Dataset for Land Process Studies over China. Sci. Data 2020, 7, 25. [Google Scholar] [CrossRef]
- Li, Q.; Shi, G.; Shangguan, W.; Nourani, V.; Li, J.; Li, L.; Huang, F.; Zhang, Y.; Wang, C.; Wang, D.; et al. A 1 Km Daily Soil Moisture Dataset over China Using in Situ Measurement and Machine Learning. Earth Syst. Sci. Data 2022, 14, 5267–5286. [Google Scholar] [CrossRef]
- Tang, W.; Qin, J.; Yang, K.; Jiang, Y.; Pan, W. Mapping Long-Term and High-Resolution Global Gridded Photosynthetically Active Radiation Using the ISCCP H-Series Cloud Product and Reanalysis Data. Earth Syst. Sci. Data 2022, 14, 2007–2019. [Google Scholar] [CrossRef]
- Ding, J.; Yang, T.; Zhao, Y.; Liu, D.; Wang, X.; Yao, Y.; Peng, S.; Wang, T.; Piao, S. Increasingly Important Role of Atmospheric Aridity on Tibetan Alpine Grasslands. Geophys. Res. Lett. 2018, 45, 2852–2859. [Google Scholar] [CrossRef]
- Zhang, Q.; Kong, D.; Shi, P.; Singh, V.P.; Sun, P. Vegetation Phenology on the Qinghai-Tibetan Plateau and Its Response to Climate Change (1982–2013). Agric. For. Meteorol. 2018, 248, 408–417. [Google Scholar] [CrossRef]
- Yang, L.; Shen, F.; Zhang, L.; Cai, Y.; Yi, F.; Zhou, C. Quantifying Influences of Natural and Anthropogenic Factors on Vegetation Changes Using Structural Equation Modeling: A Case Study in Jiangsu Province, China. J. Clean. Prod. 2021, 280, 124330. [Google Scholar] [CrossRef]
- Zhang, M.; Yuan, N.; Lin, H.; Liu, Y.; Zhang, H. Quantitative Estimation of the Factors Impacting Spatiotemporal Variation in NPP in the Dongting Lake Wetlands Using Landsat Time Series Data for the Last Two Decades. Ecol. Indic. 2022, 135, 108544. [Google Scholar] [CrossRef]
- Guo, B.; Zang, W.; Yang, F.; Han, B.; Chen, S.; Liu, Y.; Yang, X.; He, T.; Chen, X.; Liu, C.; et al. Spatial and Temporal Change Patterns of Net Primary Productivity and Its Response to Climate Change in the Qinghai-Tibet Plateau of China from 2000 to 2015. J. Arid Land 2020, 12, 1–17. [Google Scholar] [CrossRef]
- Zheng, Z.; Zhu, W.; Zhang, Y. Seasonally and Spatially Varied Controls of Climatic Factors on Net Primary Productivity in Alpine Grasslands on the Tibetan Plateau. Glob. Ecol. Conserv. 2020, 21, e00814. [Google Scholar] [CrossRef]
- Tang, Z.; Zhang, Y.; Lei, M.; Li, Z.; Zhao, G.; Chen, Y.; Zhu, W. Climate-Driven Effects on NPP in the Tibetan Plateau Alpine Grasslands Diminish with Increasing Elevation. Remote Sens. 2024, 16, 4754. [Google Scholar] [CrossRef]
- Lv, G.; Jin, J.; He, M.; Wang, C. Soil Moisture Content Dominates the Photosynthesis of C3 and C4 Plants in a Desert Steppe after Long-Term Warming and Increasing Precipitation. Plants 2023, 12, 2903. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, C.; Li, Z.; Yang, L.; Jin, X.; Gao, X. Analysis on the Temporal and Spatial Characteristics of the Shallow Soil Temperature of the Qinghai-Tibet Plateau. Sci. Rep. 2022, 12, 19746. [Google Scholar] [CrossRef]
- Gandin, A.; Koteyeva, N.K.; Voznesenskaya, E.V.; Edwards, G.E.; Cousins, A.B. The Acclimation of Photosynthesis and Respiration to Temperature in the C3–C4 Intermediate Salsola Divaricata: Induction of High Respiratory CO2 Release under Low Temperature. Plant Cell Environ. 2014, 37, 2601–2612. [Google Scholar] [CrossRef]
- Piao, S.; Nan, H.; Huntingford, C.; Ciais, P.; Friedlingstein, P.; Sitch, S.; Peng, S.; Ahlström, A.; Canadell, J.G.; Cong, N.; et al. Evidence for a Weakening Relationship between Interannual Temperature Variability and Northern Vegetation Activity. Nat. Commun. 2014, 5, 5018. [Google Scholar] [CrossRef]
- Xu, W.; Gu, S.; Zhao, X.; Xiao, J.; Tang, Y.; Fang, J.; Zhang, J.; Jiang, S. High Positive Correlation between Soil Temperature and NDVI from 1982 to 2006 in Alpine Meadow of the Three-River Source Region on the Qinghai-Tibetan Plateau. Int. J. Appl. Earth Obs. Geoinf. 2011, 13, 528–535. [Google Scholar] [CrossRef]
- Guo, L.; Chen, J.; Luedeling, E.; He, J.-S.; Cheng, J.; Wen, Z.; Peng, C. Early-Spring Soil Warming Partially Offsets the Enhancement of Alpine Grassland Aboveground Productivity Induced by Warmer Growing Seasons on the Qinghai-Tibetan Plateau. Plant Soil 2018, 425, 177–188. [Google Scholar] [CrossRef]
- Su, S.-L.; Ji, H.-J.; Zhang, D.; Zhang, S.-Q.; Li, S.-D.; Su, W.-J. Characteristics of Water Use Efficiency and Its Influencing Factors of Grassland Ecosystem in Qinghai Province. Acta Agrestia Sin. 2023, 31, 2814. [Google Scholar] [CrossRef]
- Liu, P.; Tong, X.; Meng, P.; Zhang, J.; Li, J.; Zhang, J.; Zhou, Y. Biophysical Controls on Water Use Efficiency of Six Plantations under Different Sky Conditions. Agric. For. Meteorol. 2022, 320, 108938. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, Y. Impacts of Climate, Phenology, Elevation and Their Interactions on the Net Primary Productivity of Vegetation in Yunnan, China under Global Warming. Ecol. Indic. 2023, 154, 110533. [Google Scholar] [CrossRef]
- Lin, S.; Wang, G.; Hu, Z.; Huang, K.; Sun, J.; Sun, X. Spatiotemporal Variability and Driving Factors of Tibetan Plateau Water Use Efficiency. J. Geophys. Res. Atmos. 2020, 125, e2020JD032642. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tian, Y.; Zhang, W.; Xu, X.; Zhou, B.; Cao, X.; Qiao, B. Response of Water-Use Efficiency (WUE) in Alpine Grasslands to Hydrothermal and Radiative Factors Across Elevation Gradients. Land 2025, 14, 1173. https://doi.org/10.3390/land14061173
Tian Y, Zhang W, Xu X, Zhou B, Cao X, Qiao B. Response of Water-Use Efficiency (WUE) in Alpine Grasslands to Hydrothermal and Radiative Factors Across Elevation Gradients. Land. 2025; 14(6):1173. https://doi.org/10.3390/land14061173
Chicago/Turabian StyleTian, Ye, Wan Zhang, Xiao Xu, Bingrong Zhou, Xiaoyun Cao, and Bin Qiao. 2025. "Response of Water-Use Efficiency (WUE) in Alpine Grasslands to Hydrothermal and Radiative Factors Across Elevation Gradients" Land 14, no. 6: 1173. https://doi.org/10.3390/land14061173
APA StyleTian, Y., Zhang, W., Xu, X., Zhou, B., Cao, X., & Qiao, B. (2025). Response of Water-Use Efficiency (WUE) in Alpine Grasslands to Hydrothermal and Radiative Factors Across Elevation Gradients. Land, 14(6), 1173. https://doi.org/10.3390/land14061173