


Evaluating Spatial Coverage of the Greater Sage-Grouse Umbrella to Conserve Sagebrush-Dependent Species Biodiversity within the Wyoming Basins
 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
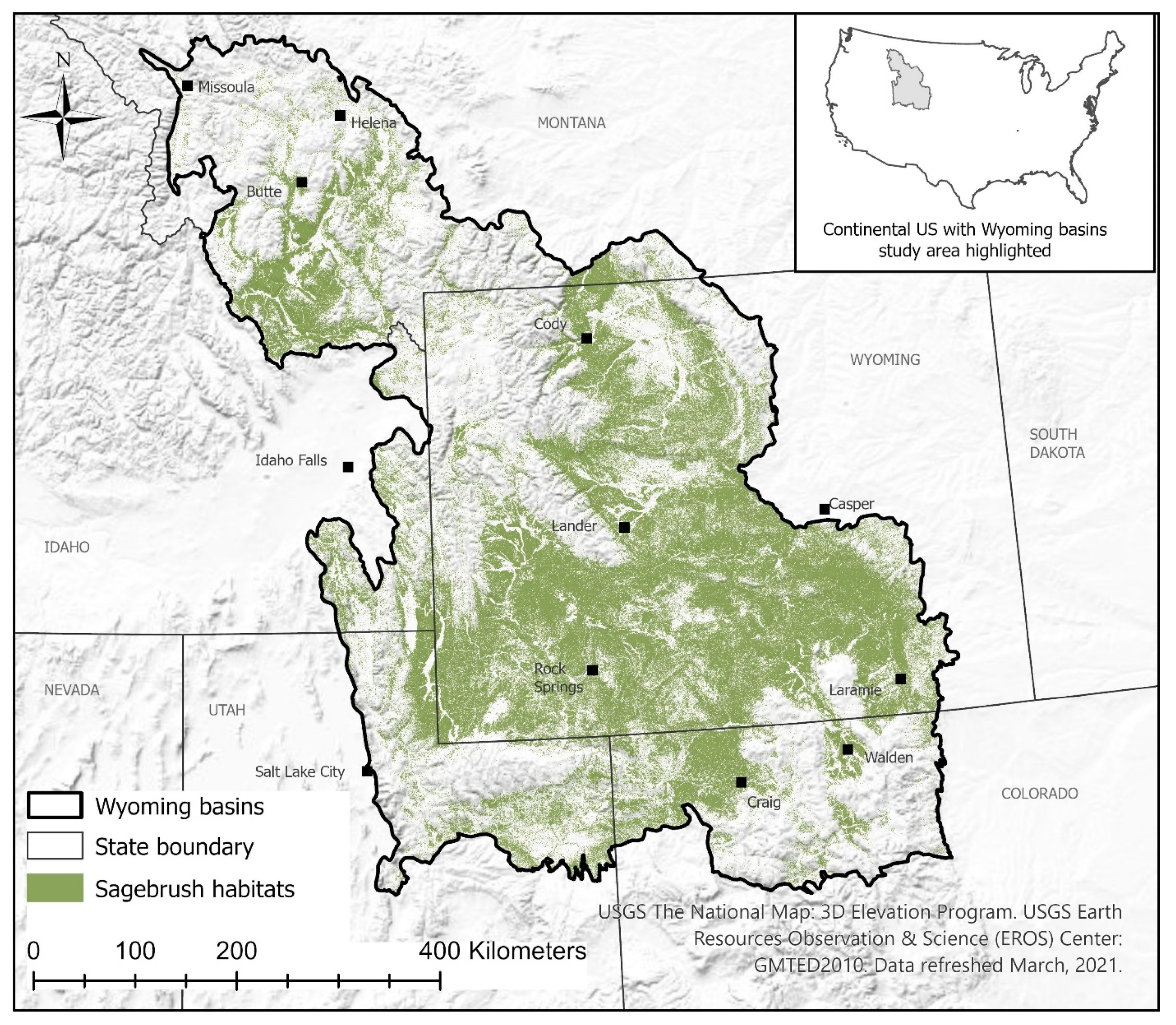
2.1. Study Area
2.2. Background Species
2.3. Data Analyses
3. Results
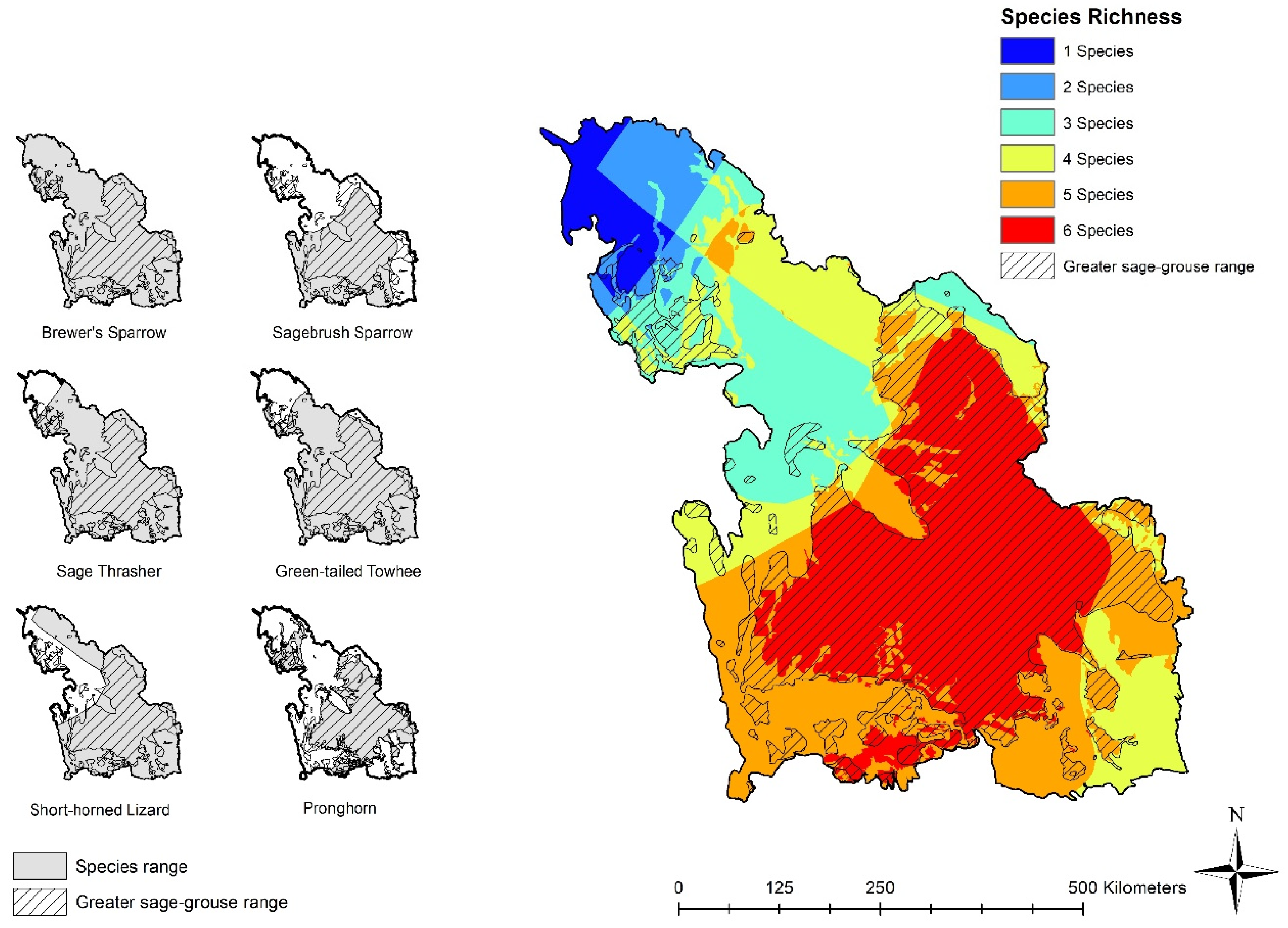
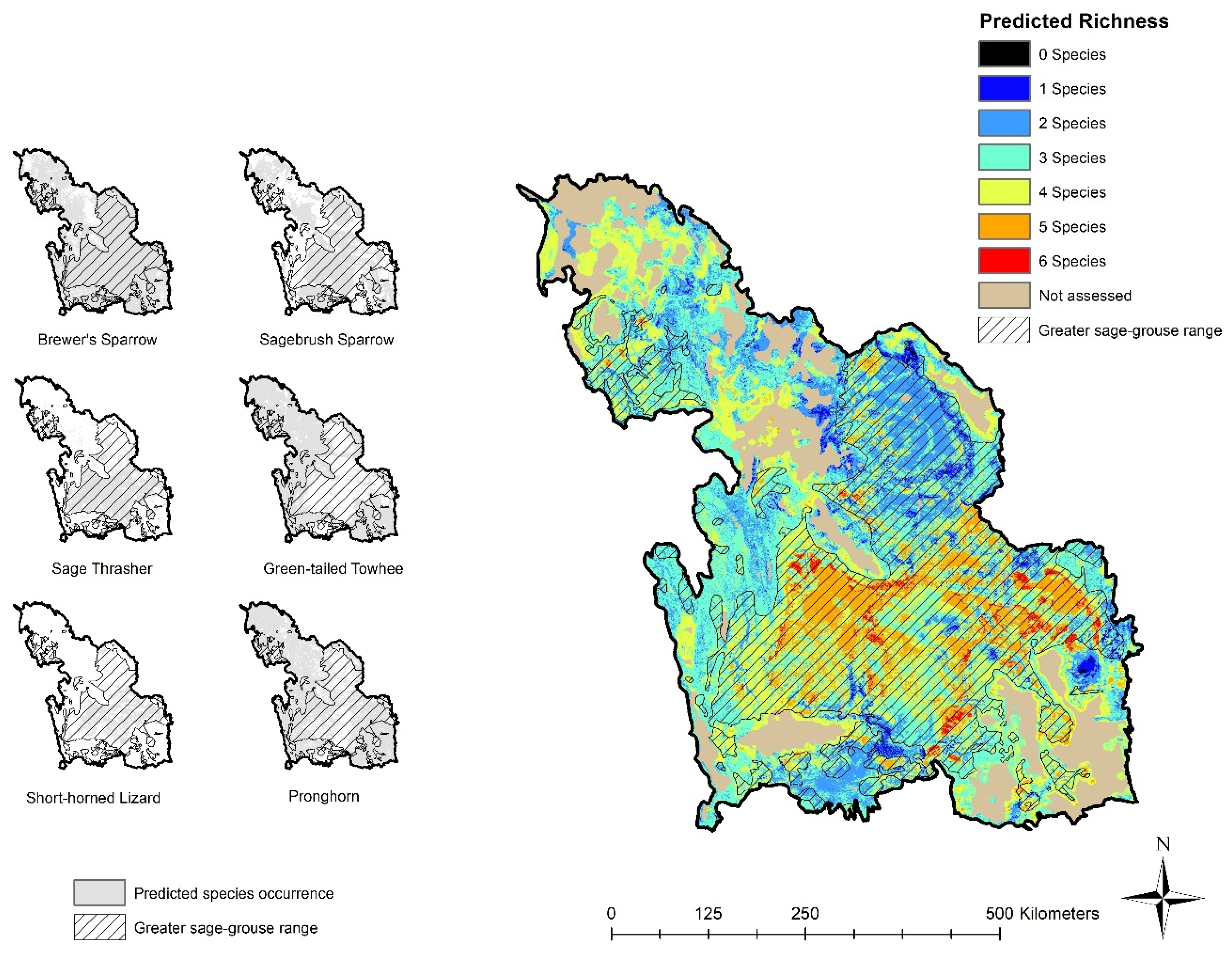
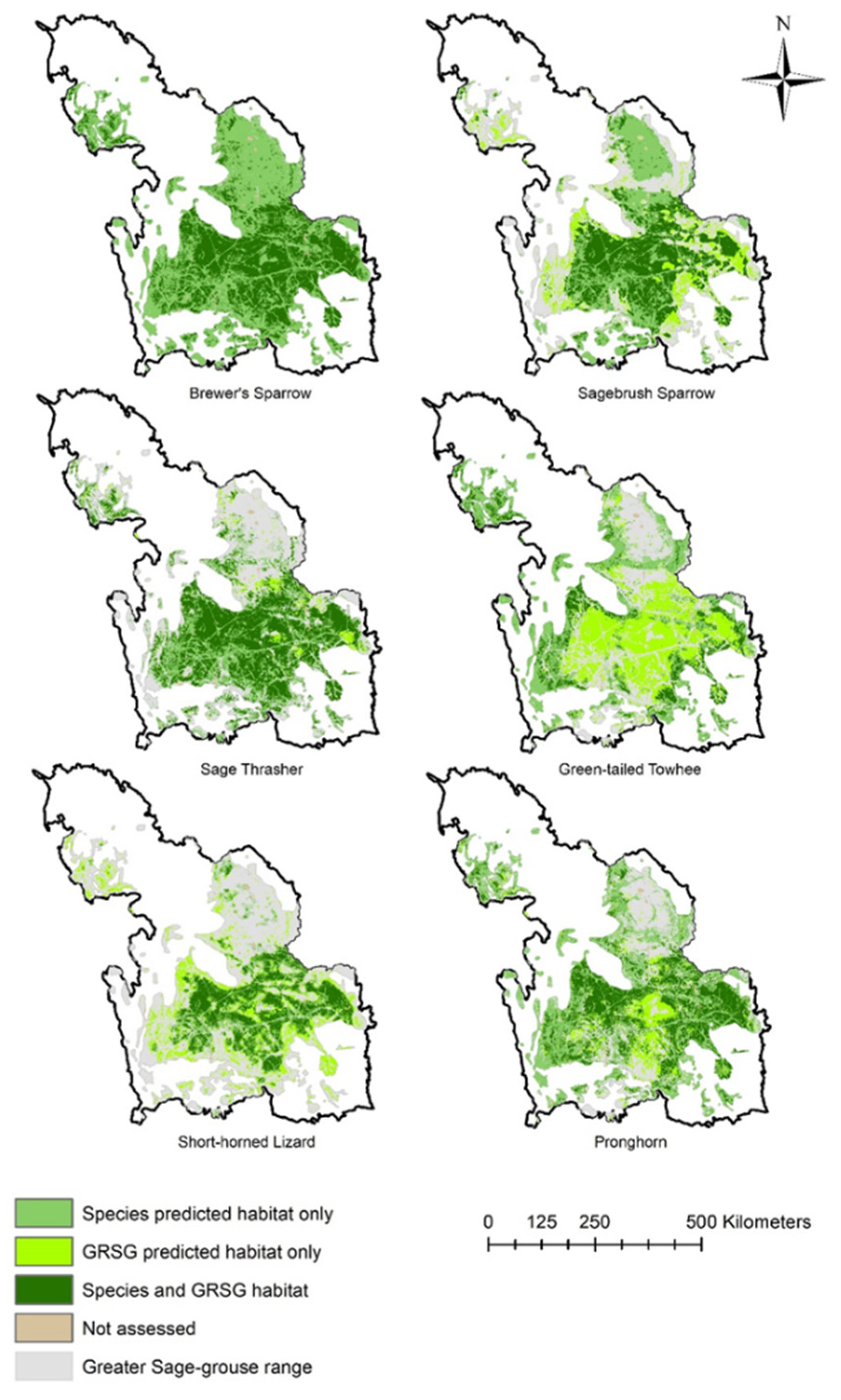
3.1. Sage-Grouse and Species Distribution
3.1.1. Range Maps and Sage-Grouse Overlap
3.1.2. Species Occurrence Models and Sage-Grouse Overlap
3.1.3. Management Areas and Sage-Grouse Overlap

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Area within Sage-Grouse Range (km2) | Percent of Sage-Grouse Range (%) | Percent Overlap with Sage-Grouse Predicted Occurrence (%) |
|---|---|---|---|
| Brewer’s sparrow | 161,253 | 99 | 39 |
| sagebrush sparrow | 90,827 | 56 | 48 |
| sage thrasher | 95,512 | 59 | 58 |
| green-tailed towhee | 68,532 | 42 | 27 |
| short-horned lizard | 46,031 | 28 | 68 |
| pronghorn | 98,501 | 60 | 44 |
| greater sage-grouse | 63,486 | 39 |
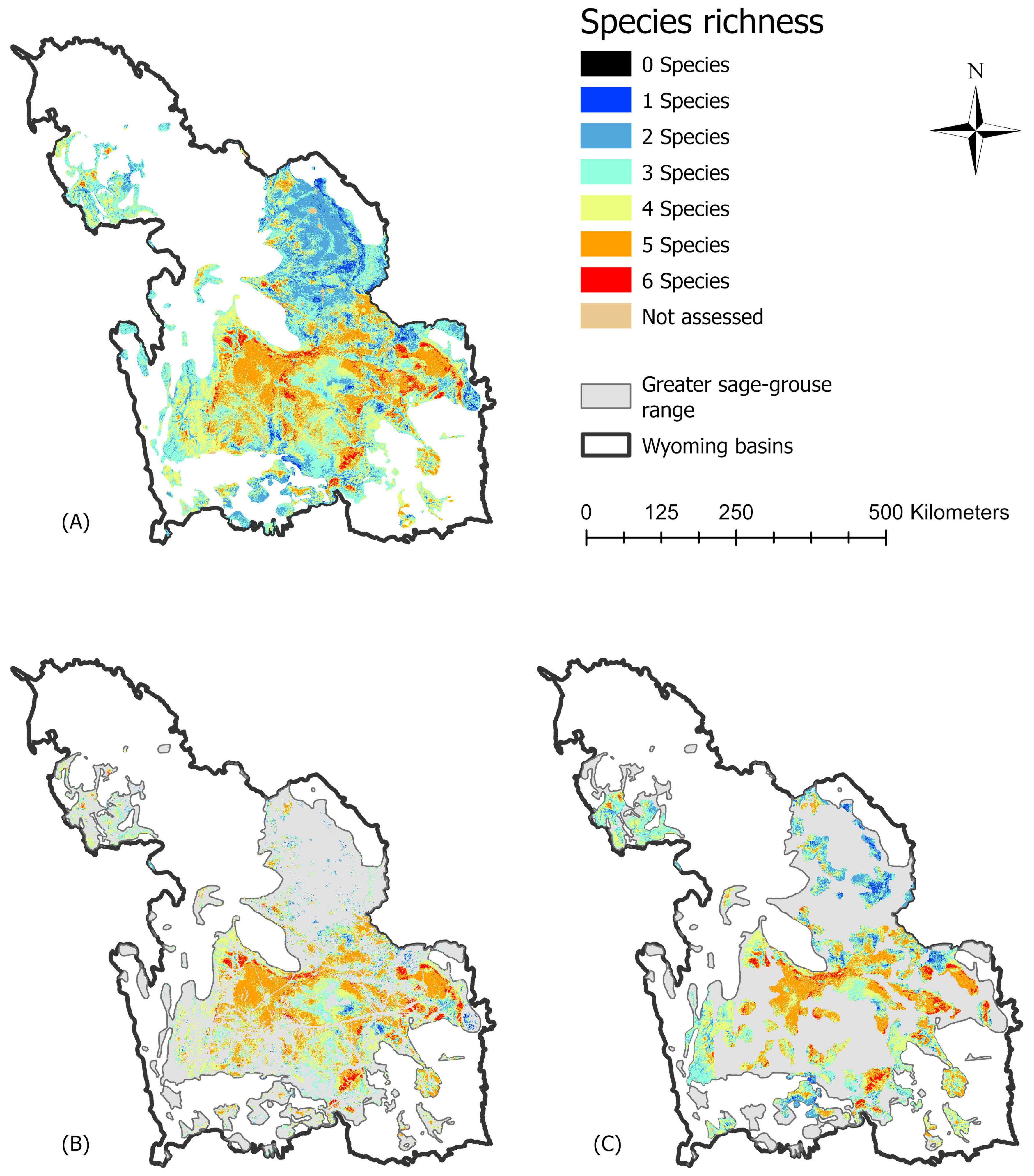
3.2. Sage-Grouse and Species Richness
3.2.1. Range Maps and Richness
3.2.2. Species Occurrence Models and Richness
3.2.3. Management Areas within Sage-Grouse Range and Richness from Predicted Occurrence
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pimm, S.L.; Jenkins, C.N.; Abell, R.; Brooks, T.M.; Gittleman, J.L.; Joppa, L.N.; Raven, P.H.; Roberts, C.M.; Sexton, J.O. The biodiversity of species and their rates of extinction, distribution, and protection. Science 2014, 344, 1246752. [Google Scholar] [CrossRef]
- Ceballos, G.; Ehrlich, P.R.; Barnosky, A.D.; García, A.; Pringle, R.M.; Palmer, T.M. Accelerated modern human-induced species losses: Entering the sixth mass extinction. Sci. Adv. 2015, 1, e1400253. [Google Scholar] [CrossRef] [PubMed]
- International Union for Conservation of Nature and Natural Resources [IUCN]. The IUCN Red List of Threatened Species. Version 2020-2. Available online: https://www.iucnredlist.org (accessed on 13 November 2020).
- Sala, O.E.; Chapin, F.S.; Armesto, J.J.; Berlo, E.; Bloomfield, J.; Dirzo, R.; Huber-Sanwald, E.; Huenneke, L.F.; Jackson, R.B.; Kinzig, A.; et al. Global biodiversity scenarios for the year 2100. Science 2000, 287, 1770–1774. [Google Scholar] [CrossRef] [PubMed]
- Dirzo, R.; Raven, P.H. Global state of biodiversity and loss. Annu. Rev. Environ. Resour. 2003, 28, 137–167. [Google Scholar] [CrossRef]
- Foley, J.A.; DeFries, R.; Asner, G.P.; Barford, C.; Bonan, G.; Carpenter, S.R.; Chapin, F.S.; Coe, M.T.; Daily, G.C.; Gibbs, H.K.; et al. Global consequences of land use. Science 2005, 309, 570–574. [Google Scholar] [CrossRef] [PubMed]
- Young, H.S.; McCauley, D.M.; Galetti, M.; Dirzo, R. Patterns, causes, and consequences of Anthropocene defaunation. Annu. Rev. Ecol. Evol. Syst. 2016, 47, 333–358. [Google Scholar] [CrossRef]
- Hooper, D.U.; Adair, E.C.; Cardinale, B.J.; Byrnes, J.E.K.; Hungate, B.A.; Matulich, K.L.; Gonzalez, A.; Duffy, J.E.; Gamfeldt, L.; O’Connor, M.I. A global synthesis reveals biodiversity loss as a major driver of ecosystem change. Nature 2012, 486, 105–108. [Google Scholar] [CrossRef]
- Dirzo, R.; Young, H.; Galetti, M.; Ceballow, G.; Isaac, N.J.B.; Collen, B. Defaunation in the Anthropocene. Science 2014, 345, 401–406. [Google Scholar] [CrossRef]
- Butchart, S.H.M.; Walpole, M.; Collen, B.; van Strien, A.; Scharlemann, J.P.W.; Almond, R.E.A.; Baillie, J.E.M.; Bomhard, B.; Brown, C.; Bruno, J.; et al. Global biodiversity: Indicators of recent declines. Science 2010, 328, 1164–1168. [Google Scholar] [CrossRef]
- America the Beautiful. Conserving and Restoring America the Beautiful. 2021. Available online: https://www.doi.gov/sites/doi.gov/files/report-conserving-and-restoring-america-the-beautiful-2021.pdf (accessed on 15 January 2022).
- Hoffmann, M.; Hilton-Taylor, C.; Angulo, A.; Böhm, M.; Brooks, T.M.; Butchart, S.H.M.; Carpenter, K.E.; Chanson, J.; Collen, B.; Cox, N.A.; et al. The impact of conservation on the status of the world’s vertebrates. Science 2010, 330, 1503–1509. [Google Scholar] [CrossRef]
- Maxwell, S.L.; Cazalis, V.; Dudley, N.; Hoffmann, M.; Rodrigues, A.S.L.; Stolton, S.; Visconti, P.; Woodley, S.; Kingston, N.; Lewis, E.; et al. Area-based conservation in the twenty-first century. Nature 2020, 586, 217–227. [Google Scholar] [CrossRef] [PubMed]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fenseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Reid, W.V. Biodiversity hotspots. Trends Ecol. Evol. 1998, 13, 275–280. [Google Scholar] [CrossRef] [PubMed]
- Lawton, J.H. Population dynamic principles. Philos. Trans. R. Soc. Lond. Biol. Sci. 1994, 334, 61–68. [Google Scholar] [CrossRef]
- Grinnell, J. The niche-relationships of the California Thrasher. Auk 1917, 34, 427–433. [Google Scholar] [CrossRef]
- Hutchinson, G.E. Concluding remarks. In Cold Spring Symposium on Quantitative Biology; Yale University: New Haven, CT, USA, 1957; pp. 415–427. [Google Scholar]
- Soberon, J. Grinnellian and Eltonian niches and geographic distributions of species. Ecol. Lett. 2007, 10, 1115–1123. [Google Scholar] [CrossRef]
- Pulliam, H.R. Sources, Sinks, and Population Regulation. Am. Nat. 1988, 132, 652–661. [Google Scholar] [CrossRef]
- Knick, S.T.; Hanser, S.E.; Preston, K.L. Modeling ecological minimum requirements for distribution of greater sage-grouse leks: Implications for population connectivity across their western range, USA. Ecol. Evol. 2013, 3, 1539–1551. [Google Scholar] [CrossRef]
- Scott, J.M.; Davis, F.; Csuti, B.; Noss, R.; Butterfield, B.; Groves, C.; Anderson, H.; Caicco, S.; D’Erchia, F.; Edwards, T.C., Jr.; et al. Gap analysis: A geographic approach to protection of biological diversity. Wildl. Monogr. 1993, 123, 1–41. [Google Scholar]
- Flather, C.H.; Knowles, M.S.; Kendall, I.A. Threatened and endangered species geography: Characteristics of hot spots in the conterminous United States. BioScience 1998, 48, 365–376. [Google Scholar] [CrossRef]
- Keinath, D.A.; Andersen, M.D.; Beauvais, G.P. Range and Modeled Distribution of Wyoming’s Species of Greatest Conservation Need; The Wyoming Natural Diversity Database, Laramie, Wyoming for the Wyoming Game and Fish Department, Cheyenne, Wyoming and the U.S. Geological Survey: Fort Collins, CO, USA, 2010; Available online: https://www.uwyo.edu/wyndd/_files/docs/reports/wynddreports/u10kei01wyus.pdf (accessed on 19 November 2020).
- Berger, J. Population constraints associated with the use of black rhinos as an umbrella species for desert herbivores. Conserv. Biol. 1997, 11, 69–78. [Google Scholar] [CrossRef]
- Branton, M.; Richardson, J.S. Assessing the value of the umbrella-species concept for conservation planning with meta-analysis. Conserv. Biol. 2011, 25, 9–20. [Google Scholar] [CrossRef]
- Noss, R.F. Indicators for monitoring biodiversity: A hierarchical approach. Conserv. Biol. 1990, 4, 355–364. [Google Scholar] [CrossRef]
- Smith, I.T.; Knetter, S.J.; Svancara, L.K.; Karl, J.W.; Johnson, T.R.; Rachlow, J.L. Overlap Between Sagebrush Habitat Specialists Differs Among Seasons: Implications for Umbrella Species Conservation. Rangel. Ecol. Manag. 2021, 78, 142–154. [Google Scholar] [CrossRef]
- Ozaki, K.; Isono, M.; Kawahara, T.; Iida, S.; Kudo, T.; Fukuyama, K. A mechanistic approach to evaluation of umbrella species as conservation surrogates. Conserv. Biol. 2006, 20, 1507–1515. [Google Scholar] [CrossRef] [PubMed]
- Roberge, J.M.; Mikusinski, G.; Svensson, S. The white-backed woodpecker: Umbrella species for forest conservation planning? Biodivers. Conserv. 2008, 17, 2479–2494. [Google Scholar] [CrossRef]
- Lambeck, R.J. Focal species: A multi-species umbrella for nature conservation. Conserv. Biol. 1997, 11, 849–856. [Google Scholar] [CrossRef]
- Carlisle, J.D.; Keinath, D.A.; Albeke, S.A.; Chalfoun, A.D. Identifying holes in the greater sage-grouse conservation umbrella. J. Wildl. Manag. 2018, 82, 948–957. [Google Scholar] [CrossRef]
- Pendergast, J.R.; Quinn, R.M.; Lawton, J.H.; Eversham, B.C.; Gibbons, D.W. Rare species, the coincidence of diversity hotspots and conservation strategies. Nature 1993, 365, 335–337. [Google Scholar] [CrossRef]
- Duchardt, C.J.; Monroe, A.P.; Heinrichs, J.A.; O’Donnell, M.S.; Edmunds, D.R.; Aldridge, C.L. Prioritizing restoration areas to conserve multiple sagebrush-associated wildlife species. Biol. Conserv. 2021, 260, 109212. [Google Scholar] [CrossRef]
- Remington, T.E.; Welty, J.L.; Aldridge, C.L.; Jakes, A.F.; Pilliod, D.S.; Rachlow, J.L.; Smith, I.T. Chapter Q. Sage-grouse Management as an Umbrella for Conservation of Sagebrush. In Sagebrush Conservation Strategy—Challenges to Sagebrush Conservation; Remington, T.E., Deibert, P.A., Hanser, S.E., Davis, D.M., Robb, L.A., Welty, J.L., Eds.; U.S. Geological Survey Open-File Report; 2020-1125; U.S. Geological Survey, 2021; pp. 163–178. [Google Scholar] [CrossRef]
- Aldridge, C.L.; Nielsen, S.E.; Beyer, H.L.; Boyce, M.S.; Connelly, J.W.; Knick, S.T.; Schroeder, M.A. Range-wide patterns of greater sage-grouse persistence. Divers. Distrib. 2008, 14, 983–994. [Google Scholar] [CrossRef]
- Connelly, J.W.; Rinkes, E.T.; Braun, C.E. Characteristics of greater sage-grouse habitat—A landscape species at micro- and macroscales. In Greater Sage-Grouse: Ecology and Conservation of a Landscape Species and Its Habitats; Studies in Avian Biology; Knick, S.T., Connelly, J.W., Eds.; University of California Press: Berkeley, CA, USA, 2011; Volume 38, pp. 69–83. [Google Scholar]
- Knick, S.T.; Hanser, S.E. Connecting pattern and process in greater sage-grouse populations and sagebrush landscapes. In Greater Sage-Grouse: Ecology and Conservation of a Landscape Species and Its Habitats; Studies in Avian Biology; Knick, S.T., Connelly, J.W., Eds.; University of California Press: Berkeley, CA, USA, 2011; Volume 38, pp. 383–405. [Google Scholar]
- Wisdom, M.J.; Meinke, C.W.; Knick, S.T.; Schroeder, M.A. Factors associated with extirpation of sage-grouse. In Greater Sage-Grouse: Ecology and Conservation of a Landscape Species and Its Habitats; Studies in Avian Biology; Knick, S.T., Connelly, J.W., Eds.; University of California Press: Berkeley, CA, USA, 2011; Volume 38, pp. 451–472. [Google Scholar]
- Schroeder, M.A.; Aldridge, C.L.; Apa, A.D.; Bohne, J.R.; Braun, C.E.; Bunnell, S.D.; Connelly, J.W.; Deibert, P.A.; Gardner, S.C.; Hilliard, M.A.; et al. Distribution of sage-grouse in North America. Condor 2004, 106, 363–376. [Google Scholar] [CrossRef]
- Coates, P.S.; Prochazka, B.G.; O’Donnell, M.S.; Aldridge, C.L.; Edmunds, D.R.; Monroe, A.P.; Ricca, M.A.; Wann, G.T.; Hanser, S.E.; Wiechman, L.A.; et al. Range-Wide Greater Sage-Grouse Hierarchical Monitoring Framework—Implications for Defining Population Boundaries, Trend Estimation, and a Targeted Annual Warning System; U.S. Geological Survey Open-File Report 2020–1154; U.S. Geological Survey: Reston, VA, USA, 2021; p. 239. [Google Scholar] [CrossRef]
- Remington, T.E.; Deibert, P.A.; Hanser, S.E.; Davis, D.M.; Robb, L.A.; Welty, J.L. Sagebrush Conservation Strategy—Challenges to Sagebrush Conservation; U.S. Geological Survey Open-File Report 2020-1125; U.S. Geological Survey: Reston, VA, USA, 2021; p. 327. [Google Scholar] [CrossRef]
- U.S. Fish and Wildlife Service. Greater Sage-Grouse (Centrocercus urophasianus) Conservation Objectives; Final Report; U.S. Department of the Interior, Fish and Wildlife Service: Denver, CO, USA, 2013; p. 91. [Google Scholar]
- Wisdom, M.J.; Rowland, M.M.; Suring, L.H. Habitat Threats in the Sagebrush Ecosystem—Methods of Regional Assessment and Applications in the Great Basin; Alliance Communications Group: Lawrence, KA, USA, 2005; p. 301. [Google Scholar]
- Rich, T.; Altman, B. Under the sage-grouse umbrella: Bird Conservation. Mag. Am. Bird Conserv. 2001, 14, 10. [Google Scholar]
- Braun, C.E. Multi-species benefits of the proposed North American Sage-Grouse Management plan. In Bird Conservation Implementation and Integration in the Americas, Proceedings of the Third International Partners in Flight Symposium; Asilomar, CA, USA, 20–24 March 2005, Ralph, C.J., Rich, T.D., Eds.; General Technical Report PSW-GTR-191; USDA Forest Service, Pacific Southwest Research Station: Albany, CA, USA, 2005; pp. 1162–1164. [Google Scholar]
- Rich, T.D.; Wisdom, M.J.; Saab, V.A. Conservation of priority birds in sagebrush ecosystems. In Proceedings of the Third International Partners in Flight Symposium; General Technical Report PSW-GTR-191; USDA Forest Service, Pacific Southwest Research Station: Albany, CA, USA, 2005; pp. 598–606. [Google Scholar]
- Rowland, M.M.; Wisdom, M.J.; Suring, L.H.; Meinke, C.W. Greater sage-grouse as an umbrella species for sagebrush-associated vertebrates. Biol. Conserv. 2006, 129, 323–335. [Google Scholar] [CrossRef]
- Hanser, S.E.; Knick, S.T. Greater sage-grouse as an umbrella species for shrubland passerine birds: A multiscale assessment. In Greater Sage-Grouse: Ecology and Conservation of a Landscape Species and Its Habitats; Studies in Avian Biology; Knick, S.T., Connelly, J.W., Eds.; University of California Press: Berkeley, CA, USA, 2011; Volume 38, pp. 451–472. [Google Scholar]
- Simberloff, D. Flagships, umbrellas, and keystones: Is single-species management passé in the landscape era? Biol. Conserv. 1998, 83, 247–257. [Google Scholar] [CrossRef]
- Caro, T. Conservation by Proxy: Indicator, Umbrella, Keystone, Flagship, and Other Surrogate Species; Island Press: Washington, DC, USA, 2010. [Google Scholar]
- Caro, T.M.; O’Doherty, G. On the use of surrogate species in conservation biology. Conserv. Biol. 1999, 13, 805–814. [Google Scholar] [CrossRef]
- Fleishman, E.; Murphy, D.D.; Brussard, P.F. A new method for selection of umbrella species for conservation planning. Ecol. Appl. 2000, 10, 569–579. [Google Scholar] [CrossRef]
- Rubinoff, D. Evaluating the California Gnatcatcher as an umbrella species for conservation of southern California coastal sage scrub. Conserv. Biol. 2001, 15, 1374–1383. [Google Scholar] [CrossRef]
- Hanser, S.E.; Leu, M.; Knick, S.T.; Aldridge, C.L. Sagebrush Ecosystem Conservation and Management: Ecoregional Assessment Tools and Models for the Wyoming Basins; Allen Press: Lawrence, KS, USA, 2011. [Google Scholar]
- Leu, M.; Hanser, S.E.; Aldridge, C.L.; Nielsen, S.E.; Cade, B.S.; Knick, S.T. Chapter 4: A sampling and analytical approach to develop spatial distribution models for sagebrush-associated species. In Sagebrush Ecosystem Conservation and Management: Ecoregional Assessment Tools and Models for the Wyoming Basins; Hanser, S.E., Leu, M., Knick, S.T., Aldridge, C.L., Eds.; Allen Press: Lawrence, KS, USA, 2011; pp. 88–111. [Google Scholar]
- WBEA Model Outputs. 2011. Available online: https://www.sciencebase.gov/catalog/item/64a5e82ed34ef77fcb0624f9?community=Forest+and+Rangeland+Ecosystem+Science+Center+%28FRESC%29 (accessed on 15 September 2023).
- Smith, I.T.; Rachlow, J.L.; Svancara, L.K.; McMahon, L.A.; Knetter, S.J. Habitat specialists as conservation umbrellas: Do areas managed for greater sage-grouse also protect pygmy rabbits? Ecosphere 2019, 10, e02827. [Google Scholar] [CrossRef]
- Pilliod, D.S.; Jeffries, M.I.; Arkle, R.S.; Olson, D.H. Reptiles under the conservation umbrella of the greater sage-grouse. J. Wildl. Manag. 2020, 84, 478–491. [Google Scholar] [CrossRef]
- Fleishman, E.; Noss, R.F.; Noon, B.R. Utility and limitations of species richness metrics for conservation planning. Ecol. Indic. 2006, 6, 543–553. [Google Scholar] [CrossRef]
- LANDFIRE, LANDFIRE 1.0.0 Existing Vegetation Type Layer. U.S. Department of Interior, Geological Survey. 2007. Available online: http://landfire.cr.usgs.gov/viewer/ (accessed on 20 September 2011).
- Rowland, M.M.; Leu, M. Study area description. In Sagebrush Ecosystem Conservation and Management: Ecoregional Assessment Tools and Models for the Wyoming Basins; Hanser, S.E., Leu, M., Knick, S.T., Aldridge, C.L., Eds.; Allen Press: Lawrence, KS, USA, 2011; pp. 10–45. [Google Scholar]
- Knick, S.T.; Hanser, S.E.; Leu, M.; Aldridge, C.L.; Wisdom, M.J. Introduction: An Ecoregional Assessment of the Wyoming Basins. In Sagebrush Ecosystem Conservation and Management: Ecoregional Assessment Tools and Models for the Wyoming Basins; Hanser, S.E., Leu, M., Knick, S.T., Aldridge, C.L., Eds.; Allen Press: Lawrence, KS, USA, 2011; pp. 1–9. [Google Scholar]
- Hanser, S.E.; Aldridge, C.L.; Leu, M.; Rowland, M.M.; Nielsen, S.E.; Knick, S.T. Greater Sage-Grouse: General Use and Roost Site Occurrence with Pellet Counts as a Measure of Relative Abundance. In Sagebrush Ecosystem Conservation and Management: Ecoregional Assessment Tools and Models for the Wyoming Basins; Hanser, S.E., Leu, M., Knick, S.T., Aldridge, C.L., Eds.; Allen Press: Lawrence, KS, USA, 2011; pp. 112–140. [Google Scholar]
- Aldridge, C.L.; Hanser, S.E.; Nielsen, S.E.; Leu, M.; Cade, B.S.; Saher, D.J.; Knick, S.T. Detectability Adjusted Count Models of Songbird Abundance. In Sagebrush Ecosystem Conservation and Management: Ecoregional Assessment Tools and Models for the Wyoming Basins; Hanser, S.E., Leu, M., Knick, S.T., Aldridge, C.L., Eds.; Allen Press: Lawrence, KS, USA, 2011; pp. 141–220. [Google Scholar]
- Wyoming Game and Fish Department [WGFD]. State Wildlife Action Plan; Wyoming Game and Fish Department: Cheyenne, WY, USA, 2017. [Google Scholar]
- Leu, M.; Hanser, S.E.; Aldridge, C.L.; Nielsen, S.E.; Suring, L.H.; Knick, S.T. Chapter 8: Occurrence of large and medium-sized mammals: Occurrence but not count models predict pronghorn distribution. In Sagebrush Ecosystem Conservation and Management: Ecoregional Assessment Tools and Models for the Wyoming Basins; Hanser, S.E., Leu, M., Knick, S.T., Aldridge, C.L., Eds.; Allen Press: Lawrence, KS, USA, 2011; pp. 315–336. [Google Scholar]
- Ngugi, K.R.; Powell, J.; Hinds, F.C.; Olson, R.A. Range animal diet composition in southcentral Wyoming. J. Range Manag. 1992, 45, 542–545. [Google Scholar] [CrossRef]
- Jakes, A.F. Chapter, F. Pronghorn. In Sagebrush Conservation Strategy—Challenges to Sagebrush Conservation; Remington, T.E., Deibert, P.A., Hanser, S.E., Davis, D.M., Robb, L.A., Welty, J.L., Eds.; U.S. Geological Survey Open-File Report 2020-1125; U.S. Geological Survey: Reston, VA, USA, 2021; pp. 37–42. [Google Scholar] [CrossRef]
- Outdoor Industry Association. The Outdoor Recreation Economy: Wyoming; Outdoor Industry Association: Boulder, CO, USA, 2017; Available online: https://outdoorindustry.org/wp-content/uploads/2017/07/OIA_RecEcoState_WY.pdf (accessed on 19 November 2020).
- Wyoming Game and Fish Department [WGFD]. Annual Reports of Big and Trophy Game Harvest: Antelope; Wyoming Game and Fish Department: Cheyenne, WY, USA, 2019. Available online: https://wgfd.wyo.gov/WGFD/media/content/PDF/Hunting/Harvest%20Reports/HR2019_Antelope.pdf (accessed on 19 November 2020).
- Knick, S.T.; Dobkin, D.S.; Rotenberry, J.T.; Schroeder, M.A.; Vander Haegen, W.M.; van Riper, C., III. Teetering on the edge or too late? Conservation and research issues for avifauna of sagebrush habitats. Condor 2003, 105, 611–634. [Google Scholar] [CrossRef]
- Green, A.W.; Aldridge, C.L.; O’Donnell, M.S. Investigating impacts of oil and gas development on Greater sage-grouse. J. Wildl. Manag. 2017, 81, 46–57. [Google Scholar] [CrossRef]
- Naugle, D.E.; Doherty, K.E.; Walker, B.L.; Copeland, H.E.; Holloran, M.J.; Tack, J.D. Sage-grouse and cumulative impacts of energy development. In Energy Development and Wildlife Conservation in North America; Naugle, D.E., Ed.; Island Press: Washington, DC, USA, 2011. [Google Scholar]
- Smith, J.T.; Evans, J.S.; Martin, B.H.; Baruch-Mordo, S.; Kiesecker, J.M.; Naugle, D.E. Reducing cultivation risk for at-risk species: Predicting outcomes of conservation easements for sage-grouse. Biol. Conserv. 2016, 201, 10–19. [Google Scholar] [CrossRef]
- Ingelfinger, F.; Anderson, S. Passerine response to roads associated with natural gas extraction in a sagebrush steppe habitat. West. N. Am. Nat. 2004, 64, 385–395. [Google Scholar]
- Gilbert, M.M.; Chalfoun, A.D. Energy development affects populations of sagebrush songbirds in Wyoming. J. Wildl. Manag. 2011, 75, 816–824. [Google Scholar] [CrossRef]
- Mutter, M.; Pavlacky, D.C.; Van Lanen, N.J.; Grenyer, R. Evaluating the impact of gas extraction infrastructure on the occupancy of sagebrush-obligate songbirds. Ecol. Appl. 2015, 25, 1175–1186. [Google Scholar] [CrossRef]
- Carlisle, J.D.; Chalfoun, A.D.; Smith, K.T.; Beck, J.L. Nontarget effects on songbirds from habitat manipulation for greater sage-grouse: Implications for the umbrella species concept. Condor Ornithol. Appl. 2018, 120, 439–455. [Google Scholar] [CrossRef]
- Hanser, S.E.; Leu, M.; Aldridge, C.L.; Nielsen, S.E.; Rowland, M.M.; Knick, S.T. Occurrence and Abundance of Ants, Reptiles, and Mammals. In Sagebrush Ecosystem Conservation and Management: Ecoregional Assessment Tools and Models for the Wyoming Basins; Hanser, S.E., Leu, M., Knick, S.T., Aldridge, C.L., Eds.; Allen Press: Lawrence, KS, USA, 2011; pp. 221–314. [Google Scholar]
- Dobbs, R.C.; Martin, P.R.; Martin, T.E. Green-tailed Towhee (Pipilo chlorurus), Version 1.0. In Birds of the World; Poole, A.F., Ed.; Cornell Lab of Ornithology: Ithaca, NY, USA, 2020. [Google Scholar]
- Holmes, A.L.; Maestas, J.D.; Naugle, D.E. Bird responses to removal of western juniper in sagebrush-steppe. Rangel. Ecol. Manag. 2017, 70, 87–94. [Google Scholar] [CrossRef]
- Ridgely, R.S.; Allnutt, T.F.; Brooks, T.; McNicol, D.K.; Mehlman, D.W.; Young, B.E.; Zook, J.R. Digital Distribution of the Birds of the Western Hemisphere, Version 1.0; NatureServe: Arlington, VA, USA, 2003; Available online: http://www.natureserve.org/getData/birdMaps.jsp (accessed on 20 September 2001).
- Stebbins, R.C. A Field Guide to Western Reptiles and Amphibians, 3rd ed.; Houghton Mifflin Company: Boston, MA, USA, 2003. [Google Scholar]
- Rowland, M.M.; Suring, L.H.; Leu, M.; Knick, S.T. Sagebrush-associated species of conservation concern. In Sagebrush Ecosystem Conservation and Management: Ecoregional Assessment Tools and Models for the Wyoming Basins; Hanser, S.E., Leu, M., Knick, S.T., Aldridge, C.L., Eds.; Allen Press: Lawrence, KS, USA, 2011; pp. 46–68. [Google Scholar]
- Gaston, K.J. How large is a species’ geographic range? Oikos 1991, 61, 434–438. [Google Scholar] [CrossRef]
- Fertig, W.; Reiners, W.A. Predicting presence/absence of plant species for range mapping: A case study from Wyoming. In Predicting Species Occurrences: Issues of Accuracy and Scale; Scott, J.M., Heglund, P.J., Morrison, M.L., Haufler, J.B., Raphael, M.G., Wall, W.A., Sanson, F.B., Eds.; Island Press: Washington, DC, USA, 2002; pp. 483–489. [Google Scholar]
- Dobkin, D.S.; Sauder, J.D. Shrub-Steppe Landscapes in Jeopardy. In Distributions, Abundances, and the Uncertain Future of Birds and Small Mammals in the Intermountain West; High Desert Ecological Research Institute: Bend, OR, USA, 2004. [Google Scholar]
- Doherty, K.E.; Naugle, D.E.; Copeland, H.E.; Pocewicz, A.; Kiesecker, J.M. Energy development and conservation tradeoffs: Systematic planning for Greater Sage-Grouse in their eastern range. In Greater Sage-Grouse: Ecology and Conservation of a Landscape Species and Its Habitats; Studies in Avian Biology; Knick, S.T., Connelly, J.W., Eds.; University of California Press: Berkeley, CA, USA, 2011; Volume 38, pp. 505–516. [Google Scholar]
- Fedy, B.C.; Doherty, K.E.; Aldridge, C.L.; O’Donnell, M.S.; Beck, J.L.; Bedrosian, B.; Gummer, D.L.; Holloran, M.J.; Johnson, G.D.; Kaczor, N.W.; et al. Habitat prioritization across large landscapes, multiple seasons, and novel areas: An example using greater sage-grouse in Wyoming. Wildl. Monogr. 2014, 190, 2795–2813. [Google Scholar] [CrossRef]
- Connelly, J.W.; Knick, S.T.; Schroeder, M.A.; Stiver, S.J. Conservation Assessment of Greater Sage-Grouse and Sagebrush Habitats; Western Association of Fish and Wildlife Agencies: Cheyenne, WY, USA, 2004. [Google Scholar]
- McGarigal, K.; Wan, H.Y.; Zeller, K.A.; Timm, B.C.; Cushman, S.A. Multi-scale habitat selection modeling: A review and outlook. Landsc. Ecol. 2016, 31, 1161–1175. [Google Scholar] [CrossRef]
- Van Lanen, N.J.; Shyvers, J.E.; Duchardt, C.J.; Aldridge, C.L. A multi-ecosystem prioritization framework to balance competing habitat conservation needs of multiple species in decline. Landsc. Ecol. 2023, 38, 1–19. [Google Scholar] [CrossRef]
- Sauer, J.R.; Hines, J.E.; Fallon, J.E.; Pardieck, K.L.; Ziolkowski, D.J., Jr.; Link, W.A. The North American Breeding Bird Survey, Results and Analysis 1966–2009; Version 3.23.2011; USGS Patuxent Wildlife Research Center: Laurel, MD, USA, 2011. [Google Scholar]
- Van Horne, B. Density as a misleading indicator of habitat quality. J. Wildl. Manag. 1983, 47, 893–901. [Google Scholar] [CrossRef]
- Dinkins, J.B.; Beck, J.L. Comparison of conservation policy benefits for and umbrella and related sagebrush-obligate species. Hum. Wildl. Interact. 2019, 13, 447–458. [Google Scholar] [CrossRef]
- Carlisle, J.D.; Stewart, D.R.; Chalfoun, A.D. An invertebrate ecosystem engineer under the umbrella of sage-grouse conservation. West. N. Am. Nat. 2017, 77, 450–463. [Google Scholar] [CrossRef]
- Duchardt, C.J.; Monroe, A.P.; Edmunds, D.R.; Holloran, M.J.; Holloran, A.G.; Aldridge, C.L. Using neutral landscape models to evaluate the umbrella species concept in an ecotone. Landsc. Ecol. 2023, 38, 1447–1462. [Google Scholar] [CrossRef]
- Van Lanen, N.J.; Monroe, A.P.; Aldridge, C.L. A hidden cost of single species management: Habitat-relationships reveal potential negative effects of conifer removal on a non-target species. Biol. Conserv. 2023, 280, 109959. [Google Scholar] [CrossRef]
- Aldridge, C.L.; Saher, D.J.; Childers, T.; Stahlnecker, K.E.; Bowen, Z.H. Crucial nesting habitat for Gunnison sage-grouse: A spatially explicit hierarchical approach. J. Wildl. Manag. 2012, 76, 391–406. [Google Scholar] [CrossRef]
- Kirol, C.P.; Beck, J.L.; Huzurbazar, S.V.; Holloran, M.J.; Miller, S.N. Identifying greater sage-grouse source and sink habitats for conservation planning in an energy development landscape. Ecol. Appl. 2015, 25, 968–990. [Google Scholar] [CrossRef] [PubMed]
- Aldridge, C.L.; Boyce, M.S. Linking occurrence and fitness to persistence: Habitat-based approach for endangered Greater Sage-Grouse. Ecol. Appl. 2007, 117, 508–526. [Google Scholar] [CrossRef] [PubMed]
- Lennon, J.J.; Koleff, P.; Greenwood, J.J.D.; Gaston, K.J. Contribution of rarity and commonness to patterns of species richness. Ecol. Lett. 2004, 7, 81–87. [Google Scholar] [CrossRef]
- Pilliod, D.S.; Jeffries, M.I. 2021. Chapter I. Sagebrush Amphibians and Reptiles. In Sagebrush Conservation Strategy—Challenges to Sagebrush Conservation; Remington, T.E., Deibert, P.A., Hanser, S.E., Davis, D.M., Robb, L.A., Welty, J.L., Eds.; U.S. Geological Survey Open-File Report 2020-1125; U.S. Geological Survey: Reston, VA, USA, 2021; pp. 67–75. [Google Scholar] [CrossRef]
- Andelman, S.J.; Fagan, W.F. Umbrellas and flagships: Efficient conservation surrogates or expensive mistakes? Proc. Natl. Acad. Sci. USA 2000, 97, 5954–5959. [Google Scholar] [CrossRef] [PubMed]
- Henry, E.; Brammer-Robbins, E.; Aschehoug, E.; Haddad, N. Do substitute species help or hinder endangered species management? Biol. Conserv. 2019, 232, 127–130. [Google Scholar] [CrossRef]
- Zipkin, E.F.; DeWan, A.; Royle, J.A. Impacts of forest fragmentation on species richness: A hierarchical approach to community modelling. J. Appl. Ecol. 2009, 46, 815–822. [Google Scholar] [CrossRef]
- Monroe, A.P.; Edmunds, D.R.; Aldridge, C.L.; Holloran, M.J.; Assal, T.J.; Holloran, A.G. Prioritizing landscapes for grassland bird conservation with hierarchical community models. Landsc. Ecol. 2021, 36, 1023–1038. [Google Scholar] [CrossRef]
- Pacifici, K.; Zipkin, E.F.; Collazo, J.A.; Irizarry, J.I.; DeWan, A. Guidelines for a priori grouping of species in hierarchical community models. Ecol. Evol. 2014, 4, 877–888. [Google Scholar] [CrossRef] [PubMed]
- Carlisle, J.D.; Chalfoun, A.D. The abundance of Greater sage-grouse as a proxy for the abundance of sagebrush-associated songbirds in Wyoming, USA. Avian Conserv. Ecol. 2020, 15, 16. [Google Scholar] [CrossRef]
- Lindenmayer, D.B.; Manning, A.D.; Smith, P.L.; Possingham, H.P.; Fischer, J.; Oliver, I.; McCarthy, M.A. The focal-species approach and landscape restoration: A critique. Conserv. Biol. 2002, 16, 338–345. [Google Scholar] [CrossRef]
- Gregg, M.A.; Crawford, J.A. Survival of Greater sage-grouse chicks and broods in the Northern Great Basin. J. Wildl. Manag. 2009, 73, 904–913. [Google Scholar] [CrossRef]
- Dumroese, R.K.; Luna, T.; Pinto, J.R.; Landis, T.D. Forbs: Foundation for restoration of monarch butterflies, other pollinators, and greater sage-grouse in the Western United States. Nat. Areas J. 2016, 36, 499–511. [Google Scholar] [CrossRef]
- Copeland, H.E.; Sawyer, H.; Monteith, K.L.; Naugle, D.E.; Pocewicz, A.; Graf, N.; Kauffman, M.J. Conserving migratory mule deer through the umbrella of sage-grouse. Ecosphere 2014, 5, 117. [Google Scholar] [CrossRef]
- Johnson, M.D. Measuring habitat quality: A review. Condor 2007, 109, 489–504. [Google Scholar] [CrossRef]
- Timmer, J.M.; Aldridge, C.L.; Fernández-Giménez, M.E. Managing for multiple species: Greater sage-grouse and sagebrush songbirds. J. Wildl. Manag. 2019, 83, 1043–1056. [Google Scholar] [CrossRef]
- Lukacs, P.M.; Seglund, A.; Boyle, S. Effects of Gunnison Sage-Grouse habitat treatment efforts on associated avifauna and vegetation structure. Avian Conserv. Ecol. 2015, 10, 7. [Google Scholar] [CrossRef]
- Barlow, N.L.; Kirol, C.P.; Doherty, K.E.; Fedy, B.C. Evaluation of the umbrella species concept at fine spatial scales. J. Wildl. Manag. 2020, 84, 237–248. [Google Scholar] [CrossRef]
- Johnson, D.H. The comparison of usage and availability measurements for evaluating resource preference. Ecology 1980, 61, 65–71. [Google Scholar] [CrossRef]
- Manier, D.J.; Wood, D.J.A.; Bowen, Z.H.; Donovan, F.M.; Holloran, M.J.; Juliusson, L.M.; Oyler-McCance, S.J.; Quamen, F.R.; Saher, D.J.; Titolo, A.J. Summary of Science Activities, Programs, and Policies That Influence the Rangewide Conservation of Greater Sage-Grouse (Centrocercus urophasianus); U.S. Geological Survey Open-File Report 2013-1098; U.S. Geological Survey: Reston, VA, USA, 2013; p. 170. Available online: http://pubs.usgs.gov/of/2013/1098/ (accessed on 15 September 2023).
- Garman, S.L. A simulation framework for assessing physical and wildlife impacts of oil and gas development scenarios in southwestern Wyoming. Environ. Model. Assess. 2018, 23, 39–56. [Google Scholar] [CrossRef]
- Brooks, M.L.; D’Antonio, C.M.; Richardson, D.M.; Grace, J.B.; Keeley, J.E.; DiTomaso, J.M.; Hobbs, R.J.; Pellant, M.; Pyke, D. Effects of invasive alien plants on fire regimes. Bioscience 2004, 54, 677–688. [Google Scholar] [CrossRef]
- Brooks, M.L.; Matchett, J.R.; Shinneman, D.J.; Coates, P.S. Fire Patterns in the Range of the Greater Sage-Grouse, 1984–2013—Implications for Conservation and Management; U.S. Geological Survey Open-File Report 2015-1167; U.S. Geological Survey: Reston, VA, USA, 2015; p. 66. [Google Scholar] [CrossRef]
- Balch, J.K.; Bradley, B.A.; D’Antonio, C.M.; Gómez-Dans, J. Introduced annual grass increases regional fire activity across the arid western USA (1980–2009). Glob. Chang. Biol. 2013, 19, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Homer, C.G.; Xian, G.; Aldridge, C.L.; Meyer, D.K.; Loveland, T.R.; O’Donnell, M.S. Forecasting sagebrush ecosystem components and greater sage-grouse habitat for 2050: Learning from past climate patterns and Landsat imagery to predict the future. Ecol. Indic. 2015, 55, 131–145. [Google Scholar] [CrossRef]
- Donnelly, J.P.; Naugle, D.E.; Hagen, C.A.; Maestas, J.D. Public lands and private waters: Scarce mesic resources structure land tenure and sage-grouse distributions. Ecosphere 2016, 7, e01208. [Google Scholar] [CrossRef]





| Range Maps | Species Occurrence Models | ||||||
|---|---|---|---|---|---|---|---|
| Species | Area within WBEA (km2) | Percent of WBEA (%) | Percent of Area within WBEA Captured by Sage-Grouse Range (%) | Area within WBEA (km2) | Percent of WBEA (%) | Percent of Area within WBEA Captured by Sage-Grouse Range (%) | Percent of Area within WBEA Captured by Sage-Grouse Occurrence (%) |
| Brewer’s sparrow | 345,300 | 100 | 47 | 267,335 | 92 | 60 | 64 |
| sagebrush sparrow | 189,915 | 55 | 69 | 136,686 | 47 | 66 | 44 |
| sage thrasher | 317,676 | 92 | 51 | 106,325 | 37 | 96 | 55 |
| green-tailed towhee | 303,864 | 88 | 53 | 179,686 | 62 | 38 | 10 |
| short-horned lizard | 265,881 | 77 | 56 | 46,833 | 16 | 98 | 67 |
| pronghorn | 158,838 | 46 | 86 | 204,399 | 71 | 48 | 21 |
| greater sage-grouse | 162,291 | 47 | 100 | 69,387 | 24 | 100 | |
| Range Maps | Species Occurrence Models | ||||||
|---|---|---|---|---|---|---|---|
| Richness | Area within WBEA (km2) | Percent of WBEA (%) | Percent of Area within WBEA Captured by Sage-Grouse Range (%) | Area within WBEA (km2) | Percent of WBEA (%) | Percent of Area within WBEA Captured by Sage-Grouse Range (%) | Percent of Area within WBEA Captured by Sage-Grouse Occurrence (%) |
| 1 Species | 13,814 | 4 | 2 | 7774 | 3 | 49 | 11 |
| 2 Species | 17,277 | 5 | 8 | 56,964 | 20 | 52 | 8 |
| 3 Species | 48,350 | 14 | 12 | 112,718 | 39 | 48 | 15 |
| 4 Species | 62,164 | 18 | 5 | 79,347 | 27 | 56 | 31 |
| 5 Species | 89,792 | 26 | 40 | 28,333 | 10 | 95 | 73 |
| 6 Species | 113,967 | 33 | 96 | 3701 | 1 | 99 | 65 |
| Richness | Area within WBEA Sage-Grouse Range (km2) | Percent of WBEA Sage-Grouse Range (%) | Percent within WBEA Sage-Grouse Range Captured by Sage-Grouse Occurrence (%) | Percent within WBEA Sage-Grouse Range Captured by PACs (%) |
|---|---|---|---|---|
| 1 Species | 3831 | 2 | 15 | 43 |
| 2 Species | 29,770 | 18 | 11 | 31 |
| 3 Species | 53,732 | 33 | 27 | 41 |
| 4 Species | 44,863 | 28 | 50 | 52 |
| 5 Species | 26,944 | 17 | 75 | 62 |
| 6 Species | 3650 | 2 | 64 | 71 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aldridge, C.L.; Saher, D.J.; Heinrichs, J.A.; Monroe, A.P.; Leu, M.; Hanser, S.E. Evaluating Spatial Coverage of the Greater Sage-Grouse Umbrella to Conserve Sagebrush-Dependent Species Biodiversity within the Wyoming Basins. Land 2024, 13, 123. https://doi.org/10.3390/land13010123
Aldridge CL, Saher DJ, Heinrichs JA, Monroe AP, Leu M, Hanser SE. Evaluating Spatial Coverage of the Greater Sage-Grouse Umbrella to Conserve Sagebrush-Dependent Species Biodiversity within the Wyoming Basins. Land. 2024; 13(1):123. https://doi.org/10.3390/land13010123
Chicago/Turabian StyleAldridge, Cameron L., D. Joanne Saher, Julie A. Heinrichs, Adrian P. Monroe, Matthias Leu, and Steve E. Hanser. 2024. "Evaluating Spatial Coverage of the Greater Sage-Grouse Umbrella to Conserve Sagebrush-Dependent Species Biodiversity within the Wyoming Basins" Land 13, no. 1: 123. https://doi.org/10.3390/land13010123
APA StyleAldridge, C. L., Saher, D. J., Heinrichs, J. A., Monroe, A. P., Leu, M., & Hanser, S. E. (2024). Evaluating Spatial Coverage of the Greater Sage-Grouse Umbrella to Conserve Sagebrush-Dependent Species Biodiversity within the Wyoming Basins. Land, 13(1), 123. https://doi.org/10.3390/land13010123