
Variations in the Distribution and Genetic Relationships among Luciola unmunsana Populations in South Korea


Abstract
:1. Introduction
2. Materials and Methods
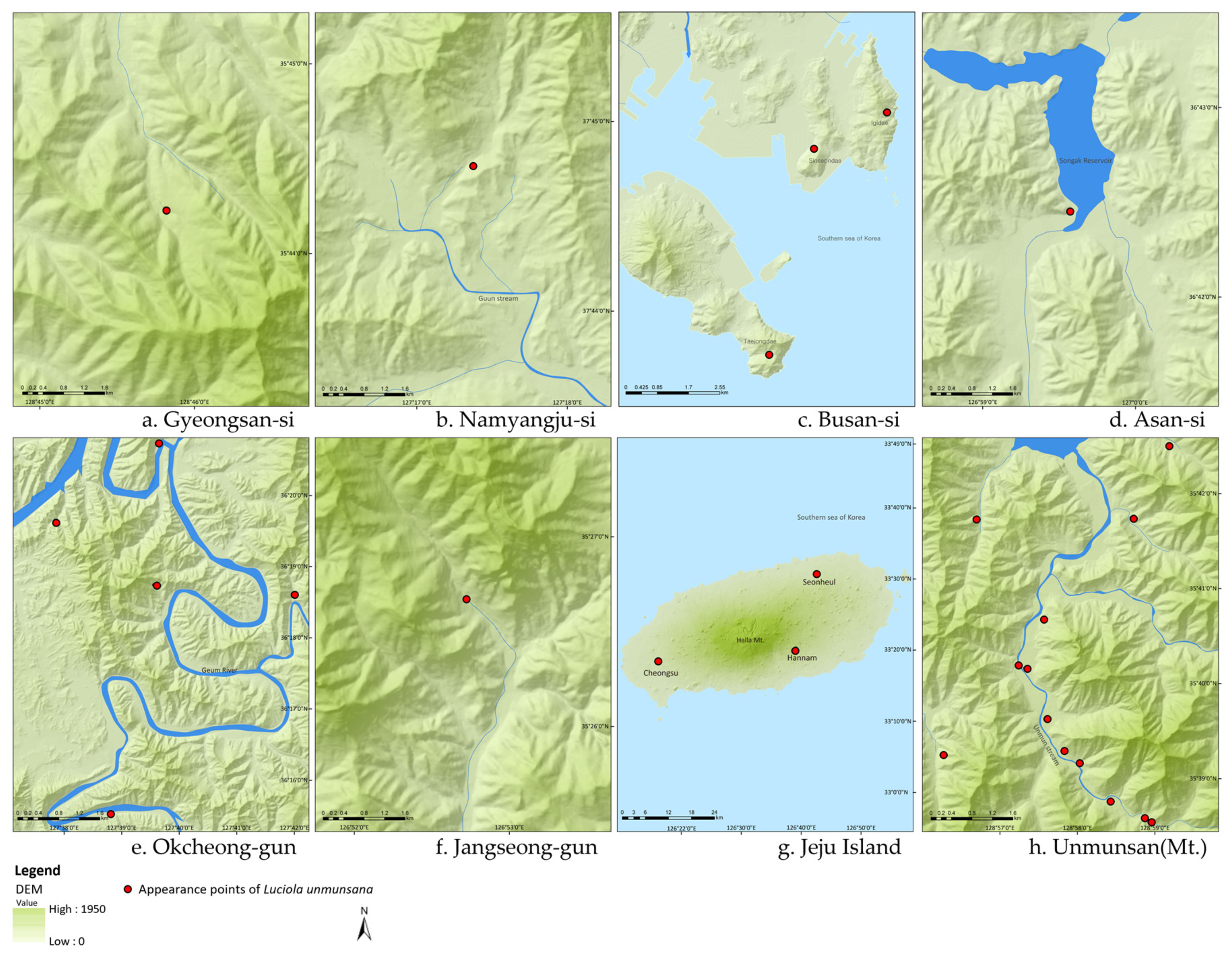
2.1. Sampling Sites for L. unmunsana
2.2. DNA Extraction
2.3. Sequencing
2.4. AMOVA Test
3. Results and Discussion
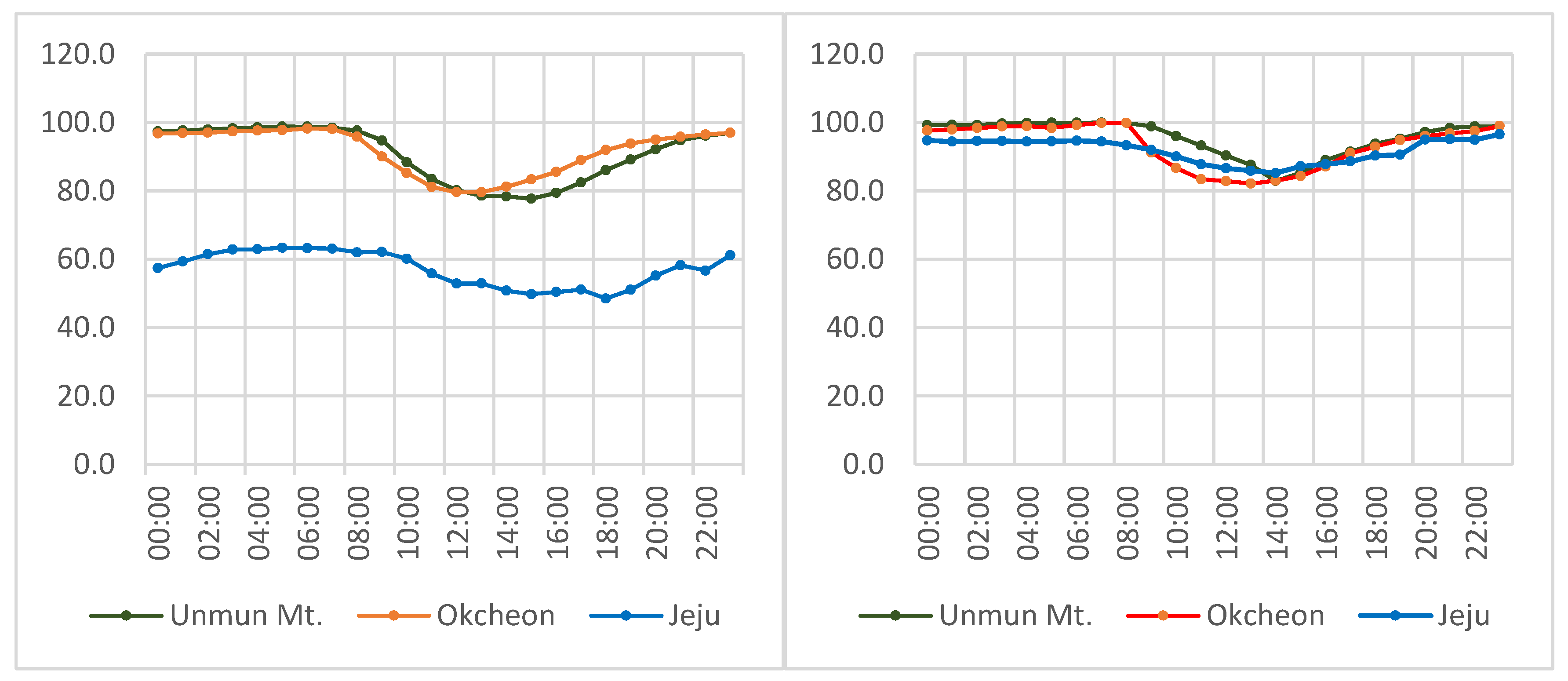
3.1. Population Density of L. unmunsana
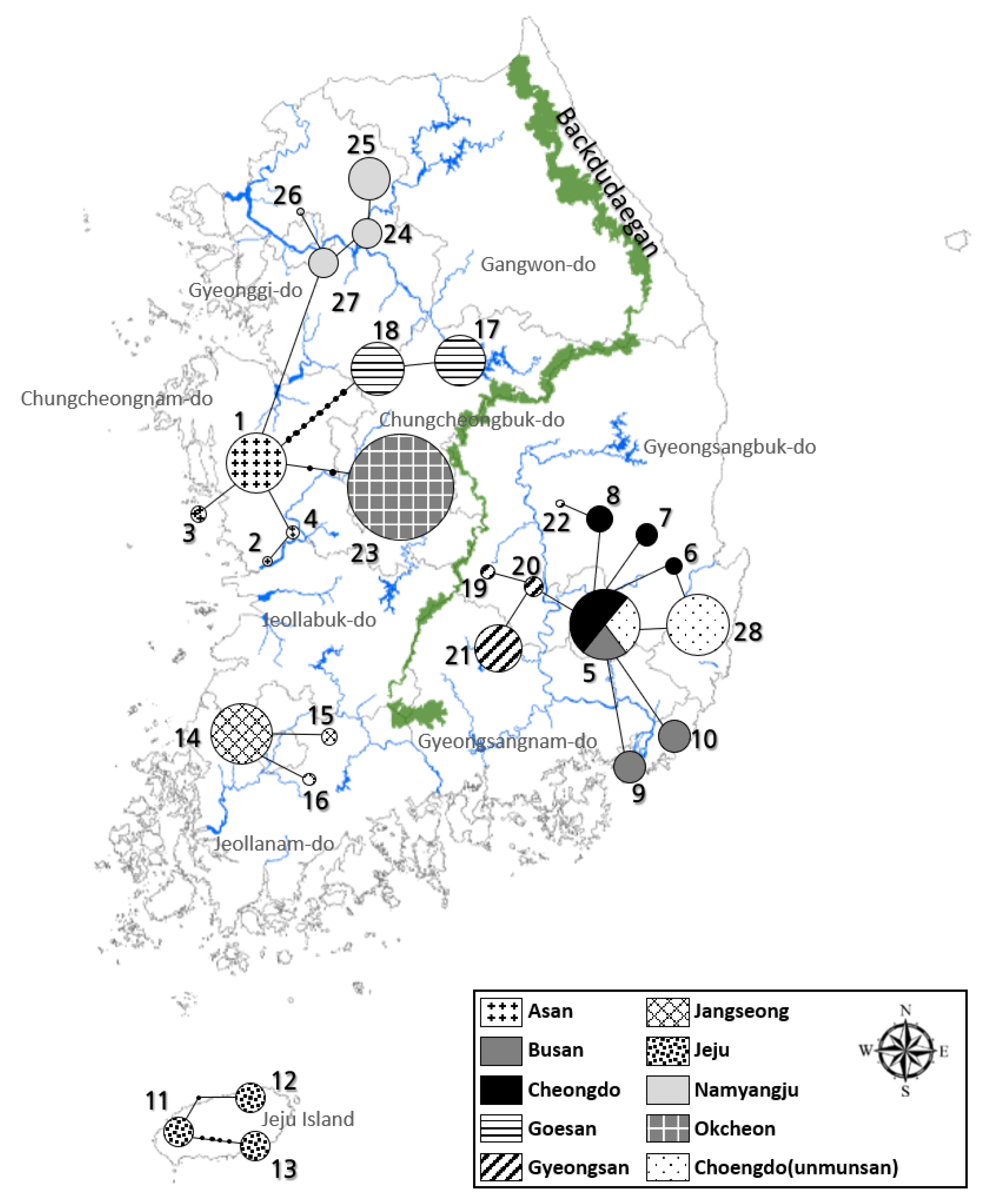
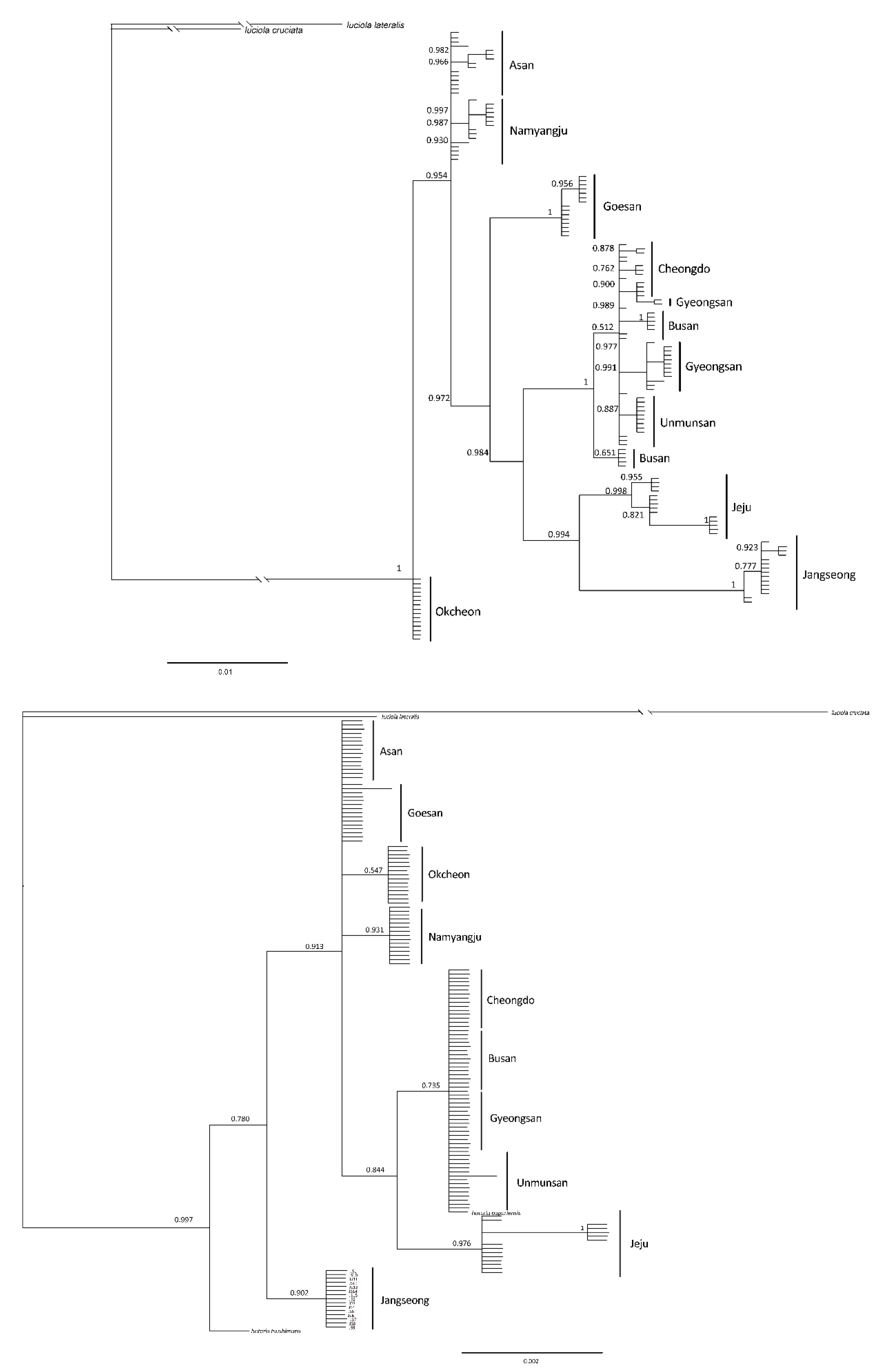
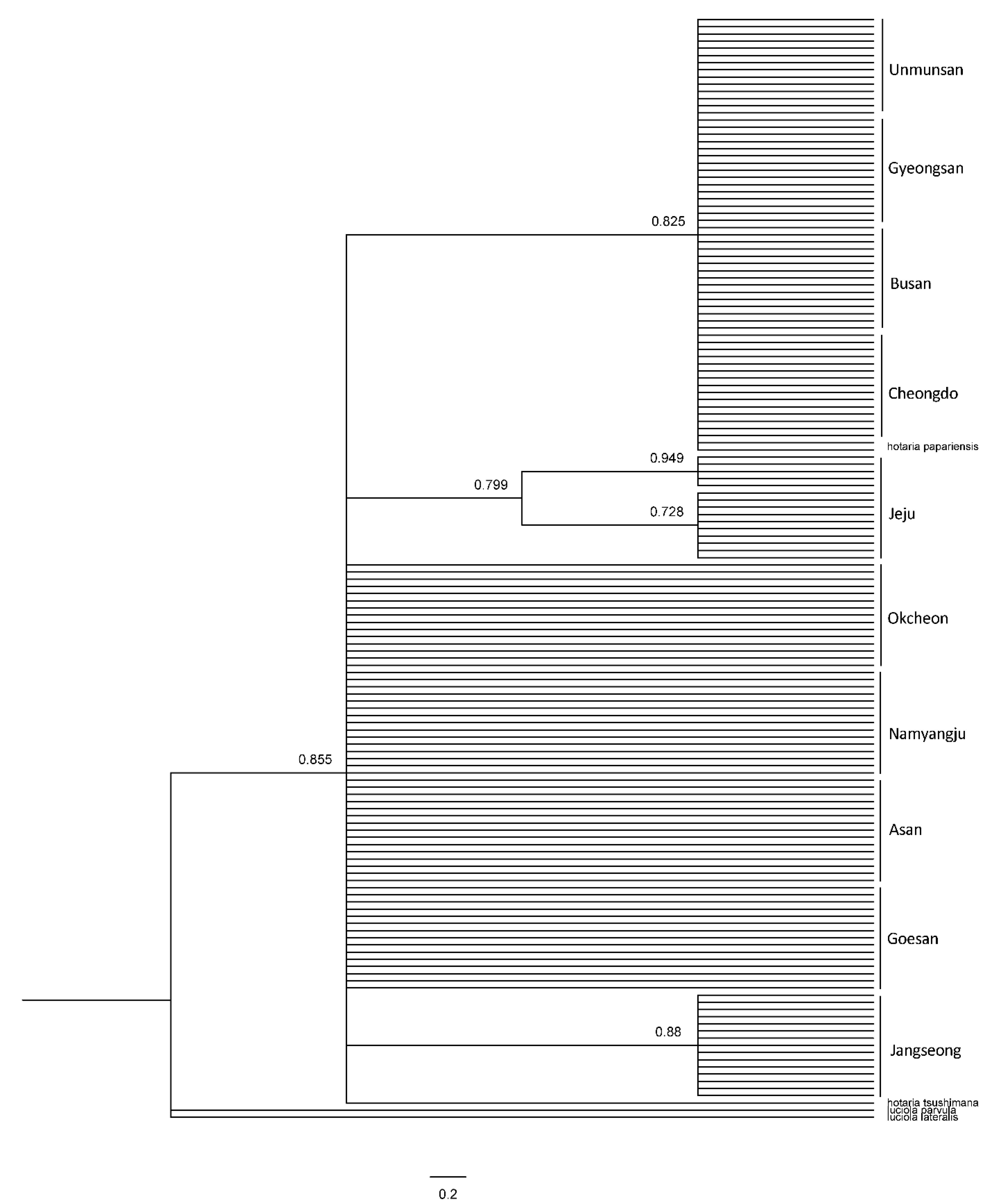
3.2. Genetic Analysis
3.3. AMOVA Test
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cho, K.H.; Lee, D.; Kim, T.; Jang, G. Measurement of 30-year urban expansion using spatial entropy in changwon and gimhae, Korea. Sustainability 2021, 13, 632. [Google Scholar] [CrossRef]
- Lee, S.; Miller-Rushing, A.J. Degradation, urbanization, and restoration: A review of the challenges and future of conservation on the Korean Peninsula. Biol. Conserv. 2014, 176, 262–276. [Google Scholar] [CrossRef]
- Sánchez-Bayo, F.; Wyckhuys, K.A.G. Worldwide decline of the entomofauna: A review of its drivers. Biol. Conserv. 2019, 232, 8–27. [Google Scholar] [CrossRef]
- Lewis, S.M.; Wong, C.H.; Owens, A.C.S.; Fallon, C.; Jepsen, S.; Thancharoen, A.; Wu, C.; De Cock, R.; Novák, M.; López-Palafox, T.; et al. A global perspective on firefly extinction threats. BioScience 2020, 70, 157–167. [Google Scholar] [CrossRef] [Green Version]
- Owens, A.C.S.; Cochard, P.; Durrant, J.; Farnworth, B.; Perkin, E.K.; Seymoure, B. Light pollution is a driver of insect declines. Biol. Conserv. 2020, 241, 108259. [Google Scholar] [CrossRef]
- Grubisic, M.; Grunsven, R.H.A.; van Kyba, C.C.M.; Manfrin, A.; Hölker, F. Insect declines and agroecosystems: Does light pollution matter? Ann. Appl. Biol. 2018, 173, 180–189. [Google Scholar] [CrossRef]
- Hagen, O.; Santos, R.M.; Schlindwein, M.N.; Viviani, V.R. Artificial Night Lighting Reduces Firefly (Coleoptera: Lampyridae) Occurrence in Sorocaba, Brazil. Adv. Entomol. 2015, 3, 24. [Google Scholar] [CrossRef] [Green Version]
- Marques, S.M.; Silva, J.E.D. Quantitative analysis of organophosphorus pesticides in freshwater using an optimized firefly luciferase-based coupled bioluminescent assay. Luminescence 2014, 29, 378–385. [Google Scholar] [CrossRef]
- Jusoh, W.; Hashim, N.R. The effect of habitat modification on firefly populations at the rembau-linggi estuary, peninsular malaysia. Lampyrid 2012, 2, 149–155. [Google Scholar]
- Vaz, S.; Manes, S.; Gama-Maia, D.; Silveira, L.; Mattos, G.; Paiva, P.C.; Figueiredo, M.; Lorini, M.L. light pollution is the fastest growing potential threat to firefly conservation in the atlantic forest hotspot. Insect Conserv. Divers. 2021, 14, 211–224. [Google Scholar] [CrossRef]
- Lee, K.; Kim, Y.; Lee, J.; Song, M.; Nam, S. Toxicity of Firefly, Luciola Lateralis (Coleoptera: Lampyridae) to Commercially Registered Insecticides and Fertilizers. Korean J. Appl. Entomol. 2008, 47, 265–272. [Google Scholar] [CrossRef]
- The Eontomological Society of Korea; Korean Society of Applied Entomology. Check List of Insects from Korea; Konkuk University Press: Seoul, Korea, 1994; p. 676. [Google Scholar]
- Lewis, S.M.; Cratsley, C.K. Flash Signal Evolution, Mate Choice, and Predation in Fireflies. Annu. Rev. Entomol. 2008, 53, 293–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, T.; Kim, S.; Yoon, H.J.; Gyun Park, I.; Park, H. Evolutionary History of Species of the Firefly Subgenus Hotaria (Coleoptera, Lampyridae, Luciolinae, Luciola) Inferred from DNA Barcoding Data. Contrib. Zool. 2020, 89, 127–145. [Google Scholar] [CrossRef]
- Doi, H. A New Species of Luciola from Korea. J. Chosen Nat. Hist. Soc. 1931, 12, 54–55. [Google Scholar]
- Choi, Y.S.; Lee, K.S.; Bae, J.S.; Lee, K.M.; Kim, S.R.; Kim, I.; Lee, S.M.; Sohn, H.D.; Jin, B.R. Molecular Cloning and Expression of a cDNA Encoding the Luciferase from the Firefly, Hotaria Unmunsana. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2002, 132, 661–670. [Google Scholar] [CrossRef]
- Choi, Y.S.; Bae, J.S.; Lee, K.S.; Kim, S.R.; Kim, I.; Kim, J.G.; Kim, K.Y.; Kim, S.E.; Suzuki, H.; Lee, S.M. Genomic Structure of the Luciferase Gene and Phylogenetic Analysis in the Hotaria-Group Fireflies. Comp. Biochem. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2003, 134, 199–214. [Google Scholar] [CrossRef]
- Kim, I.; Lee, S.; Bae, J.; Jin, B.; Kim, S.; Kim, J.; Yoon, H.; Yang, S.; Lim, S.; Sohn, H. Genetic Divergence and Phylogenetic Relationships among the Korean Fireflies, Hotaria Papariensis, Luciola Lateralis, and Pyrocoelia Rufa (Coleoptera: Lampyridae), using Mitochondrial DNA Sequences. Korean J. Appl. Entomol. 2000, 39, 211–226. [Google Scholar]
- Lee, S.; Kim, I.; Bae, J.; Jin, B.; Kim, S.; Kim, J.; Yoon, H.; Yang, S.; Lim, S.; Sohn, H. Mitochondrial DNA Sequence Variation of the Firefly, Pyrocoelia Rufa (Coleoptera: Lampyridae), in Korea. Korean J. Appl. Entomol. 2000, 39, 181–191. [Google Scholar]
- Kim, I.; Kim, J.G.; Jin, B.R. Phylogenetic Relationships of the Fireflies Co-Occurring in Korean and Japanese Territories Analyzed by Luciferase and Mitochondrial DNA Sequences. Int. J. Ind. Entomol. 2004, 9, 155–165. [Google Scholar]
- Kim, M.J.; Park, J.S.; Kim, I. Complete Mitochondrial Genome of the Korean Endemic Firefly, Luciola Unmunsana (Coleoptera: Lampyridae). Mitochondrial DNA Part B 2020, 5, 3165–3167. [Google Scholar] [CrossRef] [PubMed]
- Kang, T.W. Cantharioids I (Arthropoda, Insecta, Coleoptera). Insect Fauna Korea 2012, 12, 1–102. [Google Scholar]
- Jang, G.; Kim, T.; Kwon, K.; Lee, J.; Kim, A.; Hwang, S.; Kim, J. Research on Ecological Restoration of Luciola Unmunsana (1st Year); Daegu Regional Environment Agency: Daegu, Korea, 2013. [Google Scholar]
- Jang, G.; Kim, T.; Kwon, K.; Lee, J.; Kim, A.; Hwang, S.-H.; Kim, J.-H. Research on Ecological Restoration of Luciola Unmunsana (2nd Year); Daegu Regional Environment Agency: Daegu, Korea, 2014. [Google Scholar]
- Langellotto, G.A.; Denno, R.F.; Ott, J.R. A Trade-Off between Flight Capability and Reproduction in Males of a Wing-Dimorphic Insect. Ecology 2000, 81, 865–875. [Google Scholar] [CrossRef]
- Hajibabaei, M.; Singer, G.A.C.; Hebert, P.D.N.; Hickey, D.A. DNA Barcoding: How it Complements Taxonomy, Molecular Phylogenetics and Population Genetics. Trends Genet. 2007, 23, 167–172. [Google Scholar] [CrossRef]
- Tibayrenc, M. Bridging the Gap between Molecular Epidemiologists and Evolutionists. Trends Microbiol. 2005, 13, 575–580. [Google Scholar] [CrossRef]
- Prijovic, M.; Skaljac, M.; Drobnjakovic, T.; Zanic, K.; Peric, P.; MarÄic, D.; Puizina, J. Genetic Variation of the Greenhouse Whitefly, Trialeurodes Vaporariorum (Hemiptera: Aleyrodidae), among Populations from Serbia and Neighbouring Countries, as Inferred from COI Sequence Variability. Bull. Entomol. Res. 2014, 104, 357. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Velázquez, E.J.; Santillán-Galicia, M.T.; Novelli, V.M.; Nunes, M.A.; Mora-Aguilera, G.; Valdez-Carrasco, J.M.; Otero-Colina, G.; Freitas-Astúa, J. Diversity and Genetic Variation among Brevipalpus Populations from Brazil and Mexico. PLoS ONE 2015, 10, e0133861. [Google Scholar] [CrossRef] [Green Version]
- Sun, P.; Shi, Z.; Yin, F.; Peng, S. Genetic variation analysis of mugil cephalus in china sea based on mitochondrial coi gene sequences. Biochem. Genet. 2012, 50, 180–191. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Chen, J.; Cheng, D.; Yong, L.; Frédéric, F. Genetic Variation among the Geographic Population of the Grain Aphid, Sitobion Avenae (Hemiptera: Aphididae) in China Inferred from Mitochondrial COI Gene Sequence. Agric. Sci. China 2011, 10, 1041–1048. [Google Scholar] [CrossRef]
- Islam, S.U.; Qasim, M.; Lin, W.; Islam, W.; Arif, M.; Ali, H.; Du, Z.; Wu, Z. Genetic Interaction and Diversity of the Families Libellulidae and Gomphidae through COI Gene from China and Pakistan. Acta Trop. 2018, 182, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Pfeiler, E.; Bitler, B.G.; Ramsey, J.M.; Palacios-Cardiel, C.; Markow, T.A. Genetic Variation, Population Structure, and Phylogenetic Relationships of Triatoma Rubida and T. Recurva (Hemiptera: Reduviidae: Triatominae) from the Sonoran Desert, Insect Vectors of the Chagas’ Disease Parasite Trypanosoma Cruzi. Mol. Phylogenet. Evol. 2006, 41, 209–221. [Google Scholar] [CrossRef]
- Avise, J.C. Molecular Markers, Natural History and Evolution; Chapman & Hall: New York, NY, USA, 2012. [Google Scholar]
- Kang, T.H.; Han, T.M.; Okushima, Y.; Park, H.C. Integrative Taxonomy of Asiopodabrus Fragiliformis (Kang and Kim, 2000) (Coleoptera: Cantharidae) and its Related Species. Zootaxa 2012, 3259, 1–33. [Google Scholar]
- Lewter, J.A.; Szalanski, A.L.; Nagoshi, R.N.; Meagher Jr, R.L.; Owens, C.B.; Luttrell, R.G. Genetic Variation within and between Strains of the Fall Armyworm, Spodoptera Frugiperda (Lepidoptera: Noctuidae). Fla. Entomol. 2006, 89, 63–68. [Google Scholar] [CrossRef]
- Clements, K.M.; Wiegmann, B.M.; Sorenson, C.E.; Smith, C.F.; Neese, P.A.; Roe, R.M. Genetic Variation in the Myzus Persicae Complex (Homoptera: Aphididae): Evidence for a Single Species. Ann. Entomol. Soc. Am. 2000, 93, 31–46. [Google Scholar] [CrossRef]
- Frati, F.; Spinsanti, G.; Dallai, R. Genetic Variation of mtCOII Gene Sequences in the Collembolan Isotoma Klovstadi from Victoria Land, Antarctica: Evidence for Population Differentiation. Polar Biol. 2001, 24, 934–940. [Google Scholar] [CrossRef]
- Chen, B.; Harbach, R.E.; Butlin, R.K. Genetic Variation and Population Structure of the Mosquito Anopheles Jeyporiensis in Southern China. Mol. Ecol. 2004, 13, 3051–3056. [Google Scholar] [CrossRef]
- Clements, J.; Garcia, M.; Bradford, B.; Crubaugh, L.; Piper, S.; Duerr, E.; Zwolinska, A.; Hogenhout, S.; Groves, R.L. Genetic Variation among Geographically Disparate Isolates of Aster Yellows Phytoplasma in the Contiguous United States. J. Econ. Entomol. 2020, 113, 604–611. [Google Scholar] [CrossRef]
- Mrázek, J.; Štrosová, L.; Fliegerova, K.; Kott, T.; Kopečný, J. Diversity of insect intestinal microflora. Folia Microbiol. 2008, 53, 229–233. [Google Scholar] [CrossRef] [PubMed]
- Vobis, M.; D’Haese, J.; Mehlhorn, H.; Mencke, N.; Blagburn, B.L.; Bond, R.; Denholm, I.; Dryden, M.W.; Payne, P.; Rust, M.K. Molecular Phylogeny of Isolates of Ctenocephalides Felis and Related Species Based on Analysis of ITS1, ITS2 and Mitochondrial 16S rDNA Sequences and Random Binding Primers. Parasitol. Res. 2004, 94, 219–226. [Google Scholar] [CrossRef]
- Heiri, O.; Lotter, A.F.; Lemcke, G. Loss on Ignition as a Method for Estimating Organic and Carbonate Content in Sediments: Reproducibility and Comparability of Results. J. Paleolimnol. 2001, 25, 101–110. [Google Scholar] [CrossRef]
- Suzuki, H. Studies on Biological Diversity of Firefly in Japan. Int. J. Ind. Entomol. 2001, 2, 91–105. [Google Scholar]
- Suzuki, H.; Sato, Y.; Ohba, N.; Bae, J.; Jin, B.; Sohn, H.; Kim, S. Phylogeographic Analysis of the Firefly, Luciola Lateralis, in Japan and Korea Based on Mitochondrial Cytochrome Oxidase II Gene Sequences (Coleoptera: Lampyridae). Biochem. Genet. 2004, 42, 287–300. [Google Scholar] [CrossRef] [PubMed]
- Kimura, M. A Simple Method for Estimating Evolutionary Rates of Base Substitutions through Comparative Studies of Nucleotide Sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Holsinger, K.E.; Mason-Gamer, R.J. Hierarchical Analysis of Nucleotide Diversity in Geographically Structured Populations. Genetics 1996, 142, 629–639. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Ronquist, F. Mrbayes: Bayesian Inference of Phylogenetic Trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Excoffier, L.; Lischer, H.E.L. Arlequin Suite Ver 3.5: A New Series of Programs to Perform Population Genetics Analyses Under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Sampling Site (Population) | Sampling Date | Haplotype | PCR Success | ||||
|---|---|---|---|---|---|---|---|---|
| 16s rRNA (9) Hd: 0.9566 | COII (28) Hd: 0.9566 | 16s rRNA | COII | |||||
| 1 | Asan-si | 15 | 14.05.31 | H1 | H1, H2 H3. H4 | 15 | 15 | |
| 2 | Cheongdo-gun | 15 | 14.06.08 | H2 | H5, H6 H7, H8 | 15 | 14 | |
| 3 | Busan city | Igidae | 5 | 13.06.04 14.06.05 | H2 | H9 H10 | 15 | 12 |
| Sinseondae | 5 | 13.06.19 14.06.05 | ||||||
| Taejongdae | 5 | 13.06.19 14.06.05 | ||||||
| 4 | Gyeongsan-si | 15 | 14.05.30 | H2 | H19, H20 H21, H22 | 15 | 14 | |
| 5 | Mt. Unmun | 15 | 13.06.10 14.05.29 14.07.14 14.07.16 14.07.18 | H2 H9 | H28 | 15 | 13 | |
| 6 | Namyangju-si | 15 | 13.06.19 | H8 | H24, H25 H26, H27 | 15 | 15 | |
| 7 | Okcheon-gun | 15 | 13.06.08 14.05.31 | H7 | H23 | 15 | 15 | |
| 8 | Goesan-gun | 15 | 14.05.31 | H1 H6 | H17 H18 | 15 | 15 | |
| 9 | Jeju Island | Cheongsu | 5 | 13.07.08 14.06.26 | H3 H4 | H11 H12 H13 | 15 | 15 |
| Hannam | 5 | 13.06.24 14.06.27 | ||||||
| Seonheul | 5 | 13.06.26 14.06.29 | ||||||
| 10 | Jangseong-gun | 15 | 14.06.05 | H5 | H14, H15 H16 | 15 | 15 | |
| Gene | Sequence | Reference |
|---|---|---|
| COII | ||
| TK2-J-3037 | 5′-ATGGCAGATTAGTGCAATGG-3′ | [44,45] |
| TK-N-3785 | 5′-GTTTAAGAGACCAGTACTTG-3′ | |
| 16s rRNA | ||
| LR-J-12887-1 | 5′-CCGGTTTAAACTCAGATCATGT-3′ | [18] |
| LR-N-13398-1 | 5′-TGCCTGTTTATTAAAAACAT-3′ | |
| No. | Observation Sites | Date (2013) | No. of Males | Date (2014) | No. of Males |
|---|---|---|---|---|---|
| 1 | Mt. Unmun | 6 June 2013 | 8 | 8 June 2014 | 4 |
| 29 June 2014 | 28 | ||||
| 14 June 2013 | 12 | 14 July 2014 | 14 | ||
| 16 July 2014 | 7 | ||||
| 18 June 2013 | 7 | 18 July 2014 | 5 | ||
| 2 | Cheongdo-gun | No field investigation | 8 June 2014 | 43 | |
| 3 | Gyeongsan-si | No field investigation | 30 May 2014 | 78 | |
| 7 June 2014 | 17 | ||||
| 3 | Seonheul-ri (Jeju) | 26 June 2013 | 141 | 29 June 2014 | 17 |
| 29 June 2013 | 81 | ||||
| 4 | Hannam-ri (Jeju) | 27 June 2013 | 250 | 27 June 2014 | 97 |
| 1 July 2014 | 78 | ||||
| 5 | Cheongsu-ri (Jeju) | 34 June 2013 | 452 | 26 June 2014 | 327 |
| 28 June 2014 | 289 | ||||
| 30 June 2014 | 143 | ||||
| 6 | Okcheon-gun | 7 June 2013 | 144 | 31 May 2014 | 24 |
| 7 June 2014 | 20 | ||||
| 8 | Igidae (Busan) | 5 June 2013 | 27 | 5 June 2014 | 23 |
| 10 June 2014 | 24 | ||||
| 11 June 2014 | 7 | ||||
| 9 | Sinseondae (Busan) | 6 June 2013 | 15 | 5 June 2014 | 47 |
| 10 June 2014 | 62 | ||||
| 11 June 2014 | 47 | ||||
| 10 | Taejongdae (Busan) | 4 June 2013 | 32 | 5 June 2014 | 12 |
| 11 June 2014 | 34 | ||||
| 11 | Namyangju-si | 19 June 2013 | 55 | No field investigation | |
| 12 | Asan-si | No field investigation | 31 May 2014 | 17 | |
| 13 | Jangseong-gun | No field investigation | 5 June 2014 | 78 | |
| 14 | Goesan-gun | No field investigation | |||
| Site Name | DW | WLI | LOI (% wt. loss) | TOC | OM (%) | Soil Color | |
|---|---|---|---|---|---|---|---|
| Cheongsu | 10 | 3.8 | 38 | 17.00 | 29.31 | Brownish black |  |
| Hannam | 10 | 5.1 | 51 | 22.96 | 39.58 | ||
| Seonheul | 10 | 4.4 | 44 | 19.75 | 34.05 | ||
| Mt. Unmun | 10 | 0.4 | 4 | 1.43 | 2.46 | Grayish yellow |  |
| Igidae | 10 | 1.5 | 15 | 6.47 | 11.15 | Olive brown |  |
| Sinseondae | 10 | 0.8 | 8 | 3.26 | 5.627 | Yellowish brown |  |
| Namyangju | 10 | 0.3 | 3 | 0.97 | 1.679 | Dark grayish yellow |  |
| Source of Variation | df | Sum of Square | VC | PV | FI | ST |
|---|---|---|---|---|---|---|
| Among group | 3 | 698.70 | 6.33 | 69.54 | 0.75444 | 0.00000 |
| Among populations within group | 6 | 188.31 | 2.18 | 23.99 | 0.92869 | 0.00000 |
| Within populations | 133 | 78.31 | 0.59 | 6.47 | 0.70960 | 0.00098 |
| Total | 142 | 965.32 | 91 | 100 |
| Site | CD | UM | NC | BS | SD | AS | OC | GS | JS | JJ |
|---|---|---|---|---|---|---|---|---|---|---|
| CD | - | |||||||||
| UM | 0.3777 | - | ||||||||
| NC | 0.5644 | 0.6667 | - | |||||||
| BS | 0.3180 | 0.4609 | 0.5561 | - | ||||||
| SD | 0.9159 | 0.9392 | 0.8954 | 0.8807 | - | |||||
| AS | 0.9212 | 0.9455 | 0.9008 | 0.8868 | 0.6202 | - | ||||
| OC | 0.9662 | 0.9864 | 0.9473 | 0.9390 | 0.8905 | 0.8677 | - | |||
| GS | 0.9496 | 0.9691 | 0.9324 | 0.9220 | 0.9286 | 0.9262 | 0.9785 | - | ||
| JS | 0.9681 | 0.9794 | 0.9562 | 0.9503 | 0.9660 | 0.9699 | 0.9887 | 0.9800 | - | |
| JJ | 0.8717 | 0.8871 | 0.8575 | 0.8398 | 0.8601 | 0.8655 | 0.8980 | 0.8849 | 0.9080 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, T.-S.; Kwon, K.; Jang, G.-S. Variations in the Distribution and Genetic Relationships among Luciola unmunsana Populations in South Korea. Land 2021, 10, 730. https://doi.org/10.3390/land10070730
Kim T-S, Kwon K, Jang G-S. Variations in the Distribution and Genetic Relationships among Luciola unmunsana Populations in South Korea. Land. 2021; 10(7):730. https://doi.org/10.3390/land10070730
Chicago/Turabian StyleKim, Tae-Su, Kwanik Kwon, and Gab-Sue Jang. 2021. "Variations in the Distribution and Genetic Relationships among Luciola unmunsana Populations in South Korea" Land 10, no. 7: 730. https://doi.org/10.3390/land10070730
APA StyleKim, T.-S., Kwon, K., & Jang, G.-S. (2021). Variations in the Distribution and Genetic Relationships among Luciola unmunsana Populations in South Korea. Land, 10(7), 730. https://doi.org/10.3390/land10070730