Land-Cover and Climatic Controls on Water Temperature, Flow Permanence, and Fragmentation of Great Basin Stream Networks

 ,
,  ,
,
Abstract
1. Introduction
2. Materials and Methods
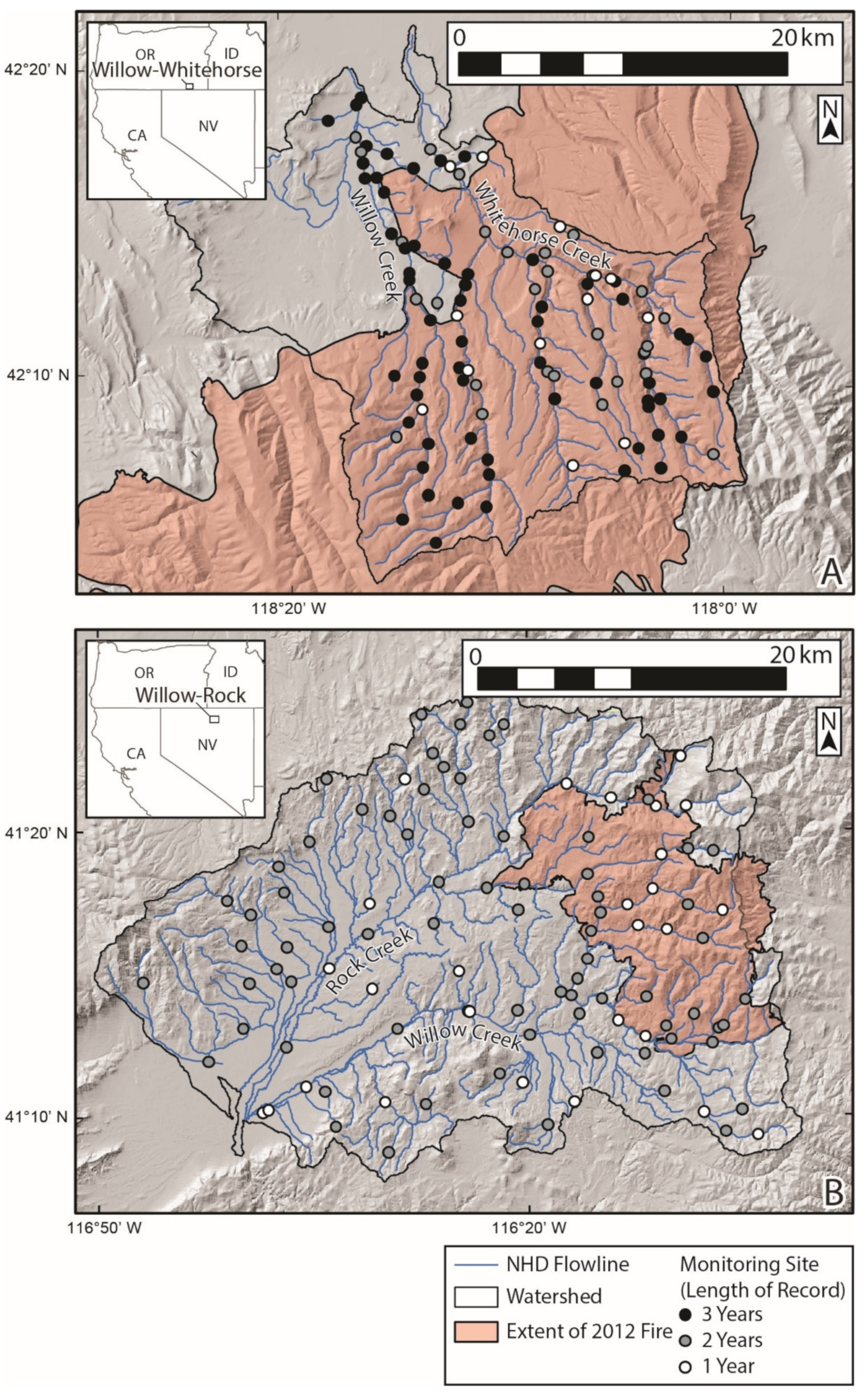
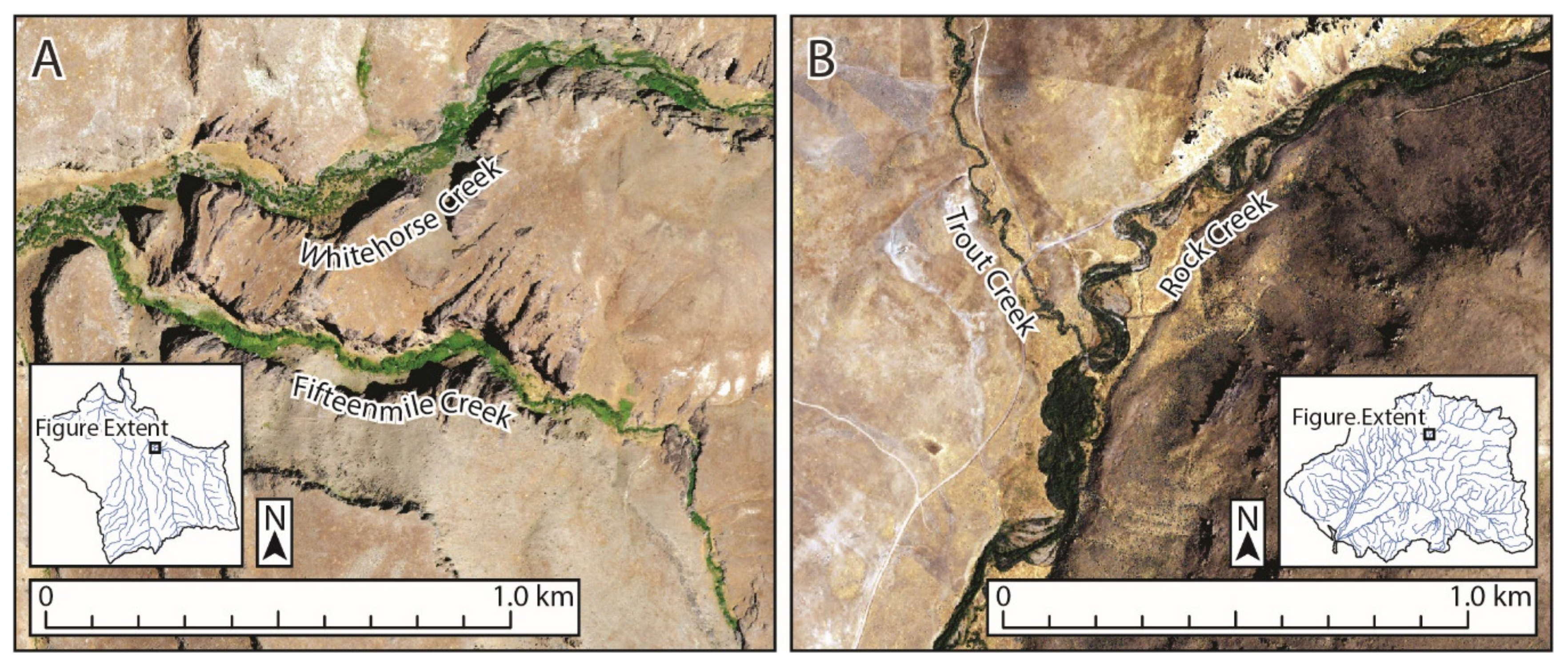
2.1. Study Sites
2.2. Data Collection
2.3. Statistical Models of Flow Presence and Stream Temperature
2.4. Flow Presence and Temperature Metric Development
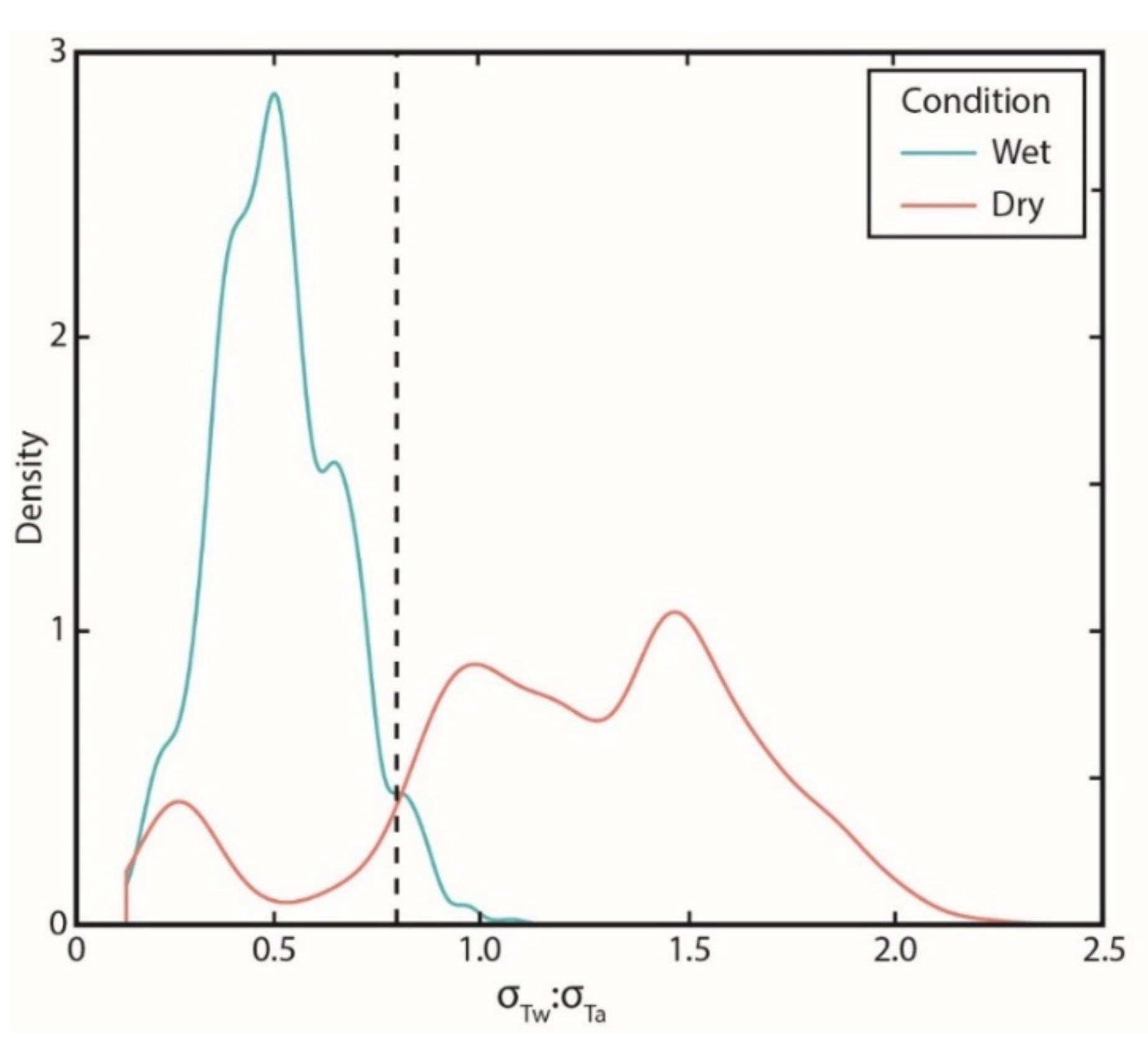
2.4.1. Diagnosis of Flow Presence/Absence
2.4.2. Summary of Temperatures and Flow Presence/Stream Drying
2.5. Model Covariates and Hydrography
3. Results
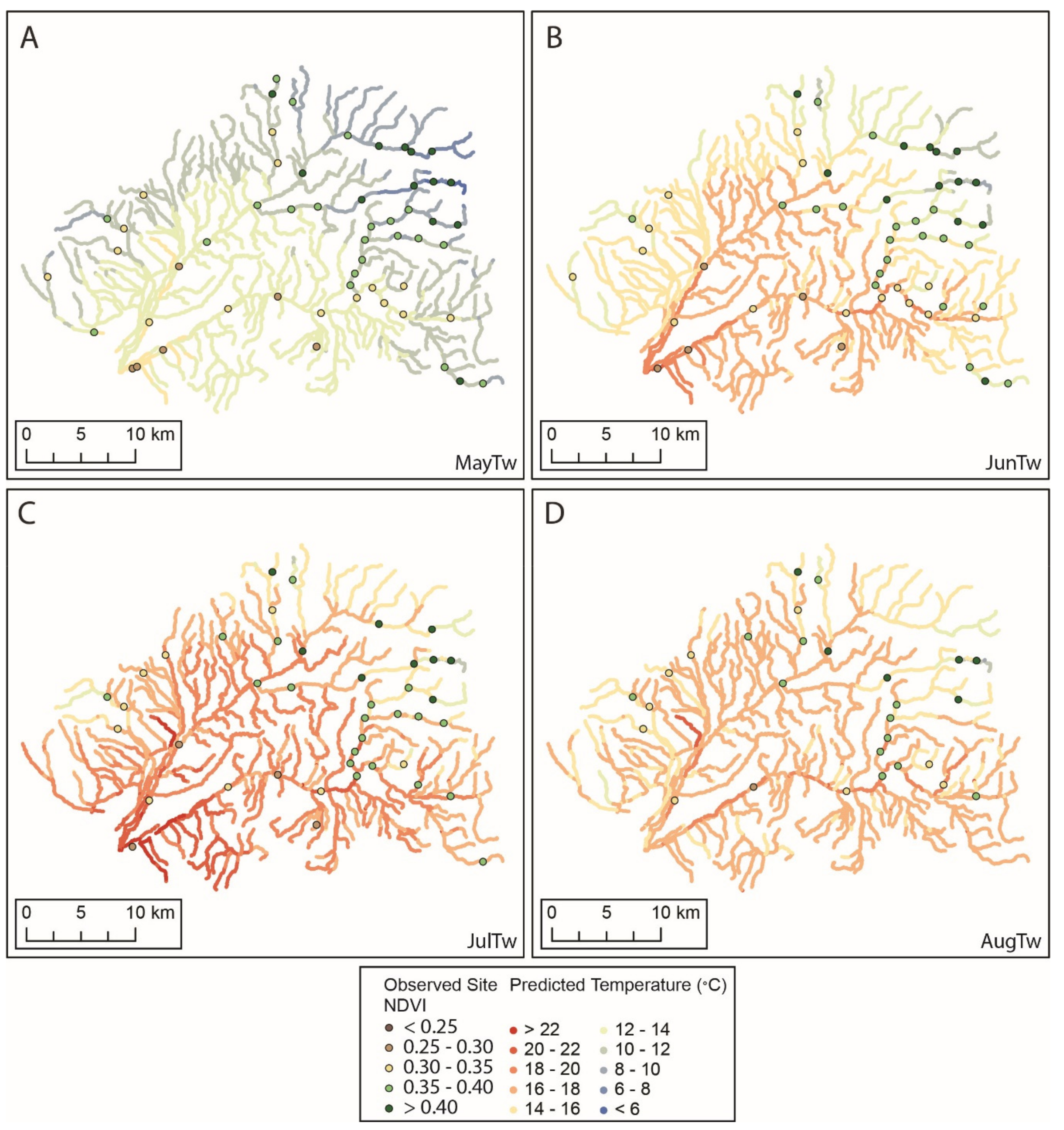
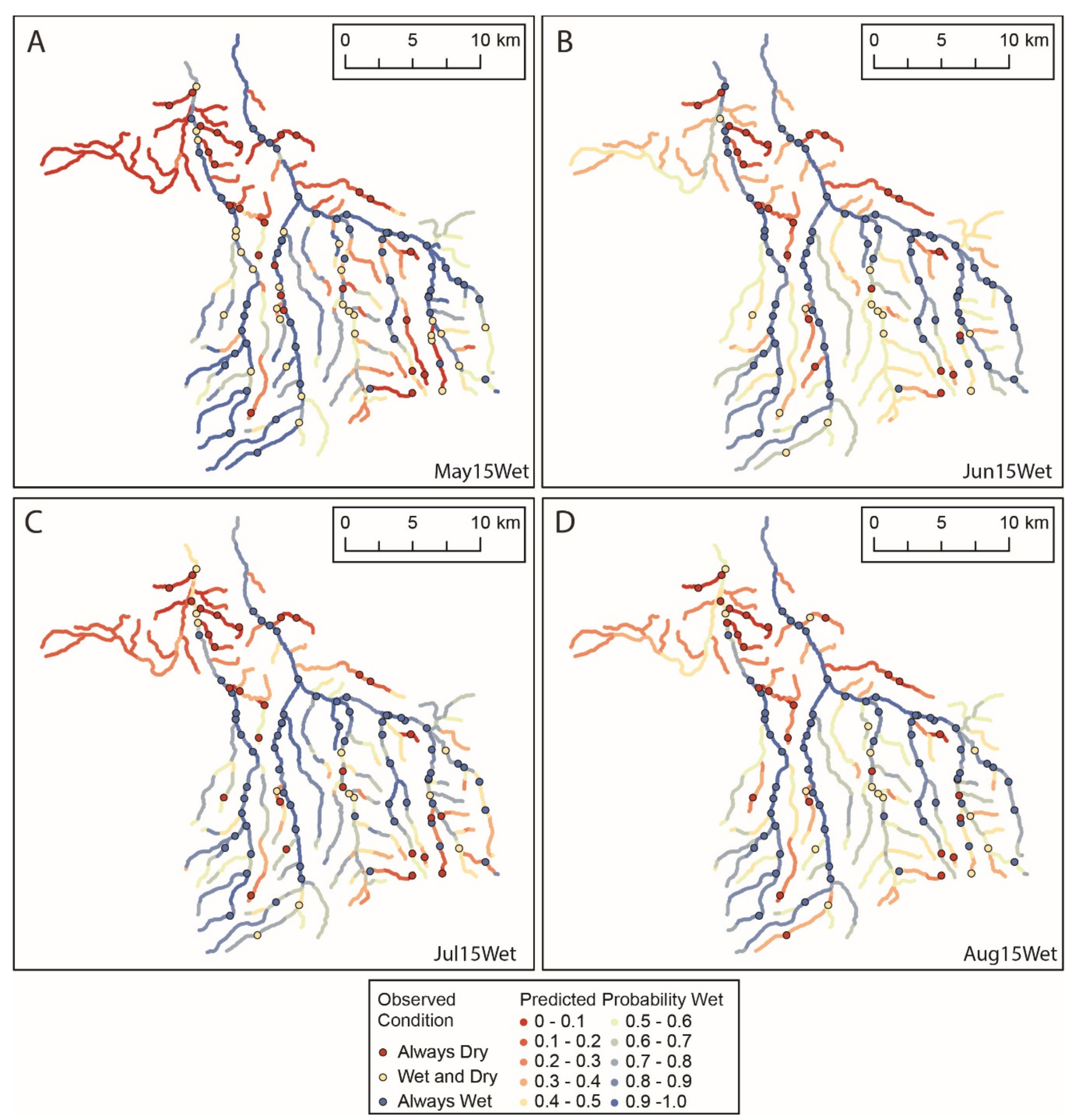
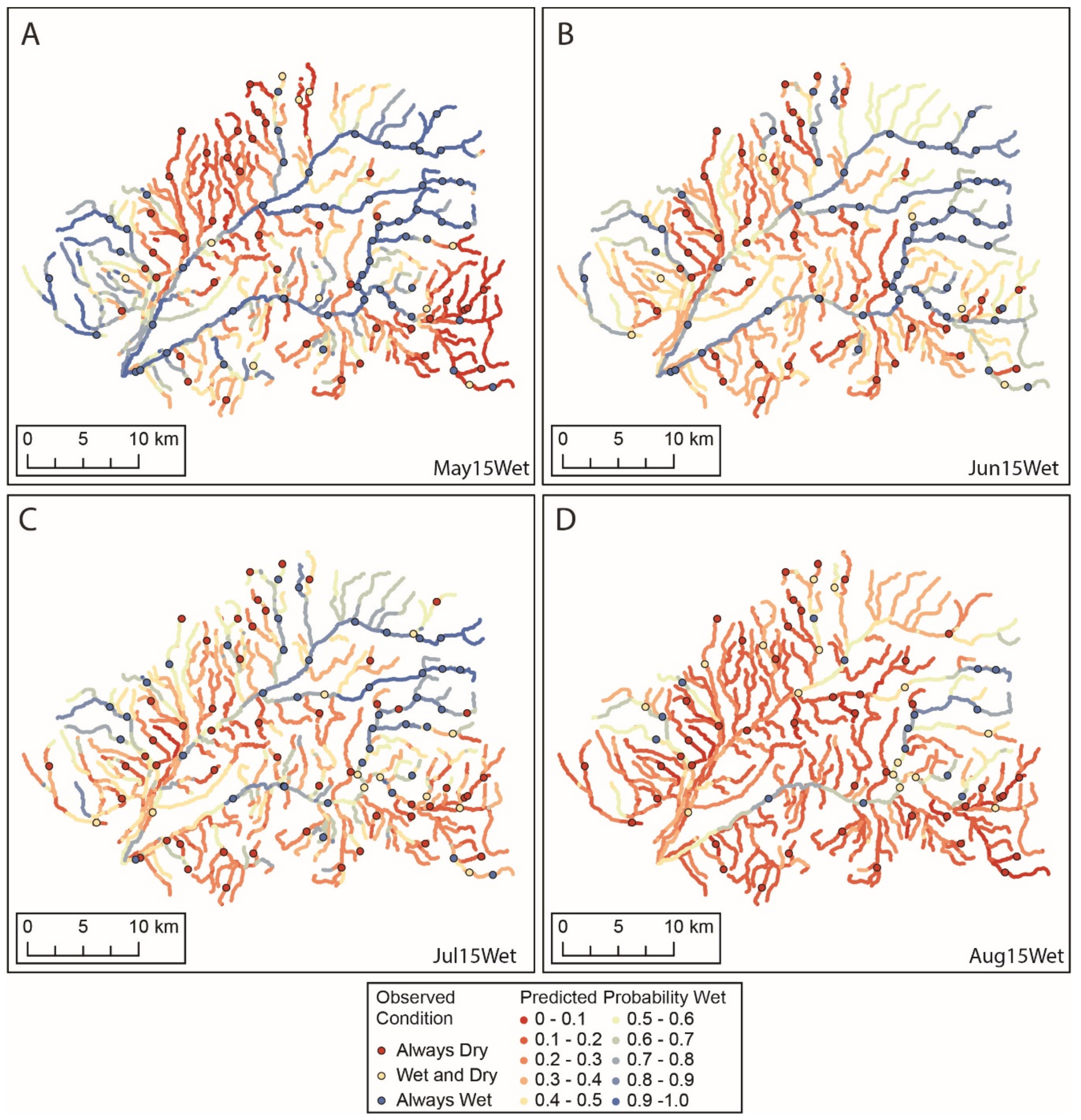
3.1. Flow Presence and Temperature
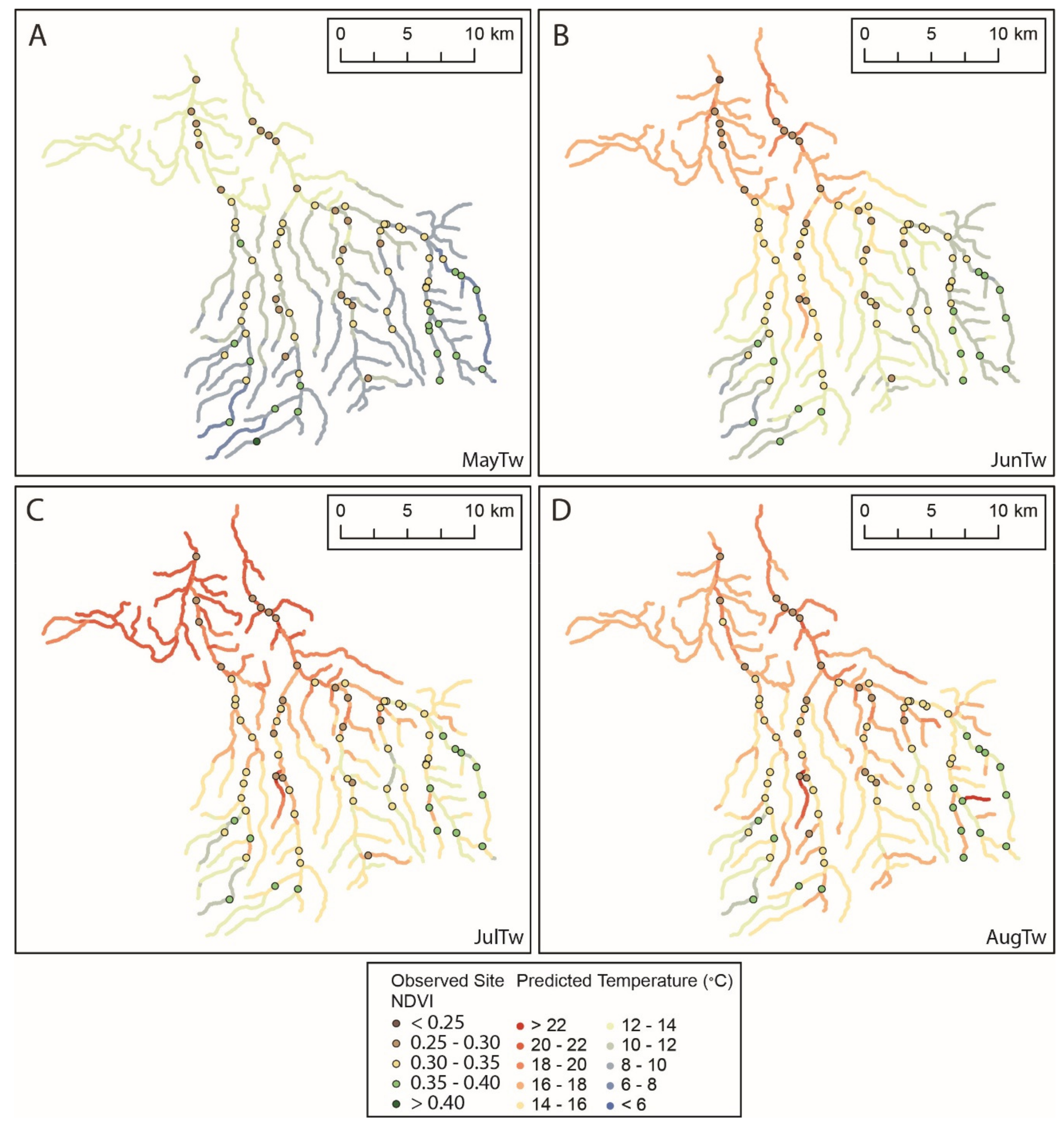
3.2. Seasonal Changes in Spatial Dependence of Stream Temperature
3.3. Models of Stream Temperature
3.4. Models of Flow Presence
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zwick, P. Stream habitat fragmentation—A threat to biodiversity. Biodivers. Conserv. 1992, 1, 80–97. [Google Scholar] [CrossRef]
- Fagan, W.F. Connectivity, fragmentation, and extinction risk in dendritic metapopulations. Ecology 2002, 83, 3243–3249. [Google Scholar] [CrossRef]
- Eros, T.; Campbell Grant, E.H. Unifying research on the fragmentation of terrestrial and aquatic habitats: Patches, connectivity and the matrix in riverscapes. Freshw. Biol. 2015, 60, 1487–1501. [Google Scholar] [CrossRef]
- Chelgren, N.D.; Dunham, J.B. Connectivity and conditional models of access and abundance of species in stream networks. Ecol. Appl. 2015, 25, 1357–1372. [Google Scholar] [CrossRef]
- Perkin, J.S.; Gido, K.B.; Costigan, K.H.; Daniels, M.D.; Johnson, E.R. Fragmentation and drying ratchet down Great Plains stream fish diversity. Aquat. Conserv.: Mar. Freshw. Ecosyst. 2015, 25, 639–655. [Google Scholar] [CrossRef]
- Matthews, W.J.; Harvey, B.C.; Power, M.E. Spatial and temporal patterns in the fish assemblages of individual pools in a midwestern stream (USA). Environ. Biol. Fishes 1994, 39, 381–397. [Google Scholar] [CrossRef]
- Gilliam, J.F.; Fraser, D.F. Movement in corridors: Enhancement by predation threat, disturbance, and habitat structure. Ecology 2001, 82, 258–273. [Google Scholar] [CrossRef]
- Meisner, J.D. Potential loss of thermal habitat for brook trout, due to climatic warming, in two southern Ontario streams. Trans. Am. Fish. Soc. 1990, 119, 282–291. [Google Scholar] [CrossRef]
- Eaton, J.G.; McCormick, J.H.; Goodno, B.E.; O’Brien, D.G.; Stefany, H.G.; Hondzo, M.; Scheller, R.M. A field information-based system for estimating fish temperature tolerances. Fisheries 1995, 20, 10–18. [Google Scholar] [CrossRef]
- Isaak, D.J.; Wenger, S.J.; Young, M.K. Big biology meets microclimatology: Defining thermal niches of ectotherms at landscape scales for conservation planning. Ecol. Appl. 2017, 27, 977–990. [Google Scholar] [CrossRef]
- Davey, A.J.H.; Kelly, D.J. Fish community responses to drying disturbances in an intermittent stream: A landscape perspective. Freshw. Biol. 2007, 52, 1719–1733. [Google Scholar] [CrossRef]
- Perkin, J.S.; Gido, K.B.; Falke, J.A.; Fausch, K.D.; Crockett, H.; Johnson, E.R.; Sanderson, J. Groundwater declines are linked to changes in Great Plains stream fish assemblages. Proc. Natl. Acad. Sci. USA 2017, 114, 7373–7378. [Google Scholar] [CrossRef] [PubMed]
- Hwan, J.L.; Carlson, S.M. Fragmentation of an intermittent stream during seasonal drought: Intra-annual and interannual patterns and biological consequences. River Res. Appl. 2016, 32, 856–870. [Google Scholar] [CrossRef]
- Matthews, W.J.; Marsh-Matthews, E. Effects of drought on fish across axes of space, time and ecological complexity. Freshw. Biol. 2003, 48, 1232–1253. [Google Scholar] [CrossRef]
- Magoulick, D.D.; Kobza, R.M. The role of refugia for fishes during drought: A review and synthesis. Freshw. Biol. 2003, 48, 1186–1198. [Google Scholar] [CrossRef]
- Ruhi, A.; Holmes, E.E.; Rinne, J.N.; Sabo, J.L. Anomalous droughts, not invasion, decrease persistence of native fishes in a desert river. Glob. Change Biol. 2015, 21, 1482–1496. [Google Scholar] [CrossRef]
- Lake, P.S. Drought and Aquatic Ecosystems: Effects and Responses; John Wiley & Sons: West Sussex, UK, 2011. [Google Scholar]
- Van Loon, A.F.; Gleeson, T.; Clark, J.; Van Dijk, A.I.; Stahl, K.; Hannaford, J.; Di Baldassarre, G.; Teuling, A.J.; Tallaksen, L.M.; Uijlenhoet, R. Drought in the Anthropocene. Nat. Geosci. 2016, 9, 89–91. [Google Scholar] [CrossRef]
- Vose, J.M.; Miniat, C.F.; Luce, C.H.; Asbjornsen, H.; Caldwell, P.V.; Campbell, J.L.; Grant, G.E.; Isaak, D.J.; Loheide, S.P.; Sun, G. Ecohydrological implications of drought for forests in the United States. For. Ecol. Manag. 2016, 380, 335–345. [Google Scholar] [CrossRef]
- Fagan, W.E.; Aumann, C.; Kennedy, C.M.; Unmack, P.J. Rarity, fragmentation, and the scale dependence of extinction risk in desert fishes. Ecology 2005, 86, 34–41. [Google Scholar] [CrossRef]
- Kovach, R.P.; Dunham, J.B.; Al-Chokhachy, R.; Snyder, C.D.; Letcher, B.H.; Young, J.A.; Beever, E.A.; Pederson, G.T.; Lynch, A.J.; Hitt, N.P.; et al. An integrated framework for ecological drought across riverscapes of North America. Bioscience 2019, 69, 418–431. [Google Scholar] [CrossRef]
- Costigan, K.H.; Jaeger, K.L.; Goss, C.W.; Fritz, K.M.; Goebel, P.C. Understanding controls on flow permanence in intermittent rivers to aid ecological research: Integrating meteorology, geology and land cover. Ecohydrology 2016, 9, 1141–1153. [Google Scholar] [CrossRef]
- Costigan, K.; Kennard, M.; Leigh, C.; Sauquet, E.; Datry, T.; Boulton, A. Flow Regimes in Intermittent Rivers and Ephemeral Streams. In Intermittent Rivers and Ephemeral Streams; Datry, T., Bonada, N., Boulton, A., Eds.; Elsevier: London, UK, 2017; pp. 51–78. [Google Scholar] [CrossRef]
- Jaeger, K.L.; Olden, J.D.; Pelland, N.A. Climate change poised to threaten hydrologic connectivity and endemic fishes in dryland streams. Proc. Natl. Acad. Sci. USA 2014, 111, 13894–13899. [Google Scholar] [CrossRef] [PubMed]
- Torgersen, C.E.; Price, D.M.; Li, H.W.; McIntosh, B.A. Multiscale thermal refugia and stream habitat associations of chinook salmon in northeastern Oregon. Ecol. Appl. 1999, 9, 301–319. [Google Scholar] [CrossRef]
- Fausch, K.D.; Torgersen, C.E.; Baxter, C.V.; Li, H.W. Landscapes to riverscapes: Bridging the gap between research and conservation of stream fishes. Bioscience 2002, 52, 483–498. [Google Scholar] [CrossRef]
- Lowe, W.H.; Likens, G.E.; Power, M.E. Linking scales in stream ecology. Bioscience 2006, 56, 591–597. [Google Scholar] [CrossRef]
- Schultz, L.D.; Heck, M.P.; Hockman-Wert, D.; Allai, T.; Wenger, S.; Cook, N.A.; Dunham, J.B. Spatial and temporal variability in the effects of wildfire and drought on thermal habitat for a desert trout. J. Arid Environ. 2017, 145, 60–68. [Google Scholar] [CrossRef]
- Dettinger, M.D.; Diaz, H.F. Global characteristics of stream flow seasonality and variability. J. Hydrometeorol. 2000, 1, 289–310. [Google Scholar] [CrossRef]
- Luce, C.H.; Abatzoglou, J.T.; Holden, Z.A. The missing mountain water: Slower westerlies decrease orographic enhancement in the Pacific Northwest USA. Science 2013, 342, 1360–1364. [Google Scholar] [CrossRef]
- Black, B.A.; van der Sleen, P.; Di Lorenzo, E.; Griffin, D.; Sydeman, W.J.; Dunham, J.B.; Rykaczewski, R.R.; Garcia-Reyes, M.; Safeeq, M.; Arismendi, I.; et al. Rising synchrony controls western North American ecosystems. Glob. Change Biol. 2018, 24, 2305–2314. [Google Scholar] [CrossRef]
- Moore, R.D.; Spittlehouse, D.L.; Story, A. Riparian microclimate and stream temperature response to forest harvesting: A review. J. Am. Water Resour. Assoc. 2005, 41, 813–834. [Google Scholar] [CrossRef]
- Daly, C.; Conklin, D.R.; Unsworth, M.H. Local atmospheric decoupling in complex topography alters climate change impacts. Int. J. Climatol. 2010, 30, 1857–1864. [Google Scholar] [CrossRef]
- Roper, B.B.; Saunders, W.C.; Ojala, J.V. Did changes in western federal land management policies improve salmonid habitat in streams on public lands within the Interior Columbia River Basin? Environ. Monit. Assess. 2019, 191, 574. [Google Scholar] [CrossRef]
- Spies, T.A.; Long, J.W.; Charnley, S.; Hessburg, P.F.; Marcot, B.G.; Reeves, G.H.; Lesmeister, D.B.; Reilly, M.J.; Cerveny, L.K.; Stine, P.A. Twenty-five years of the Northwest Forest Plan: What have we learned? Front. Ecol. Environ. 2019, 17, 511–520. [Google Scholar] [CrossRef]
- Lyon, J.G. GIS for Water Resources and Watershed Management; CRC Press: London, UK, 2002; pp. 17–22. [Google Scholar]
- Crausbay, S.D.; Ramirez, A.R.; Carter, S.L.; Cross, M.S.; Hall, K.R.; Bathke, D.J.; Betancourt, J.L.; Colt, S.; Cravens, A.E.; Dalton, M.S.; et al. Defining ecological drought for the twenty-first century. B. Am. Meteorol. Soc. 2017, 98, 2543–2550. [Google Scholar] [CrossRef]
- Dunham, J.B.; Vinyard, G.L.; Rieman, B.E. Habitat fragmentation and extinction risk of Lahontan cutthroat trout. N. Am. J. Fish. Manag. 1997, 17, 1126–1133. [Google Scholar] [CrossRef]
- Dunham, J.B.; Rieman, B.E.; Peterson, J.T. Patch-based models to predict species occurrence: Lessons from salmonid fishes in streams. In Predicting Species Occurrences: Issues of Accuracy and Scale; Scott, J., Morrison, M., Heglund, P., Eds.; Island Press: Washington, DC, USA, 2002; pp. 327–334. [Google Scholar]
- Dunham, J.B.; Peacock, M.M.; Rieman, B.E.; Schroeter, R.E.; Vinyard, G.L. Local and geographic variability in the distribution of stream-living Lahontan cutthroat trout. Trans. Am. Fish. Soc. 1999, 128, 875–889. [Google Scholar] [CrossRef]
- Warren, D.R.; Dunham, J.B.; Hockman-Wert, D. Geographic variability in elevation and topographic constraints on the distribution of native and nonnative trout in the Great Basin. Trans. Am. Fish. Soc. 2014, 143, 205–218. [Google Scholar] [CrossRef]
- Wenger, S.J.; Isaak, D.J.; Dunham, J.B.; Fausch, K.D.; Luce, C.H.; Neville, H.M.; Rieman, B.E.; Young, M.K.; Nagel, D.E.; Horan, D.L. Role of climate and invasive species in structuring trout distributions in the interior Columbia River Basin, USA. Can. J. Fish. Aquat. Sci. 2011, 68, 988–1008. [Google Scholar] [CrossRef]
- Leasure, D.R.; Wenger, S.J.; Chelgren, N.D.; Neville, H.M.; Dauwalter, D.C.; Bjork, R.; Fesenmyer, K.A.; Dunham, J.B.; Peacock, M.M.; Luce, C.H.; et al. Hierarchical multi-population viability analysis. Ecology 2019, 100, e02538. [Google Scholar] [CrossRef]
- Snyder, K.A.; Evers, L.; Chambers, J.C.; Dunham, J.; Bradford, J.B.; Loik, M.E. Effects of changing climate on the hydrological cycle in cold desert ecosystems of the Great Basin and Columbia Plateau. Rangel. Ecol. Manag. 2019, 72, 1–12. [Google Scholar] [CrossRef]
- Grayson, D. The Great Basin: A Natural Prehistory; University of California Press: Berkeley, CA, USA, 2011. [Google Scholar]
- Omernik, J.M.; Griffith, G.E. Ecoregions of the conterminous United States: Evolution of a hierarchical spatial framework. Environ. Manag. 2014, 54, 1249–1266. [Google Scholar] [CrossRef] [PubMed]
- Coffin, P.D.; Cowan, W.F. Lahontan Cutthroat Trout (Oncorhynchus Clarki Henshawi) Recovery Plan; US Fish and Wildlife Service, Region 1: Portland, OR, USA, 1995. [Google Scholar]
- Jones, K.K.; Dambacher, J.M.; Lovatt, B.G.; Talabere, A.G.; Bowers, W. Status of Lahontan cutthroat trout in the Coyote Lake basin, southeast Oregon. N. Am. J. Fish. Manag. 1998, 18, 308–317. [Google Scholar] [CrossRef]
- Peacock, M.; Neville, H.; Finger, A. The Lahontan Basin Evolutionary Lineage of Cutthroat Trout. In Cutthroat Trout: Evolutionary Biology and Taxonomy; Trotter, P., Bisson, P., Schultz, L., Roper, B., Eds.; American Fisheries Society: Bethesda, MD, USA, 2018; Volume 36, pp. 231–259. [Google Scholar]
- Carter, D.T.; Ely, L.L.; O'Connor, J.E.; Fenton, C.R. Late Pleistocene outburst flooding from pluvial Lake Alvord into the Owyhee River, Oregon. Geomorphology 2006, 75, 346–367. [Google Scholar] [CrossRef]
- PRISM Climate Group. PRISM Gridded Climate Data; Oregon State University: Corvallis, OR, USA, 2004. [Google Scholar]
- National Operational Hydrologic Remote Sensing Center. Snow Data Assimilation System (SNODAS) Data Products at NSIDC, Version 1; NSIDC: National Snow and Ice Data Center: Boulder, CO, USA, 2004. [Google Scholar] [CrossRef]
- Dunham, J.; Schroeter, R.; Rieman, B. Influence of maximum water temperature on occurrence of Lahontan cutthroat trout within streams. N. Am. J. Fish. Manag. 2003, 23, 1042–1049. [Google Scholar] [CrossRef]
- Falke, J.A.; Dunham, J.B.; Hockman-Wert, D.; Pahl, R. A simple prioritization tool to diagnose impairment of stream temperature for coldwater fishes in the Great Basin. N. Am. J. Fish. Manag. 2016, 36, 147–160. [Google Scholar] [CrossRef]
- Swanson, S.R.; Wyman, S.; Evans, C. Practical grazing management to meet riparian objectives. J. Rangel. Appl. 2015, 2, 1–28. [Google Scholar]
- Booth, D.T.; Cox, S.E.; Simonds, G.; Sant, E.D. Willow cover as a stream-recovery indicator under a conservation grazing plan. Ecol. Indic. 2012, 18, 512–519. [Google Scholar] [CrossRef]
- Fesenmyer, K.A.; Dauwalter, D.C.; Evans, C.; Allai, T. Livestock management, beaver, and climate influences on riparian vegetation in a semi-arid landscape. PLoS ONE 2018, 13, e0208928. [Google Scholar] [CrossRef]
- Blasch, K.W.; Ferre, T.P.A.; Christensen, A.H.; Hoffmann, J.P. New field method to determine streamflow timing using electrical resistance sensors. Vadose Zone J. 2002, 1, 289–299. [Google Scholar] [CrossRef]
- Arismendi, I.; Dunham, J.B.; Heck, M.P.; Schultz, L.D.; Hockman-Wert, D. A statistical method to predict flow permanence in dryland streams from time series of stream temperature. Water 2017, 9, 946. [Google Scholar] [CrossRef]
- Som, N.A.; Monestiez, P.; Hoef, J.M.V.; Zimmerman, D.L.; Peterson, E.E. Spatial sampling on streams: Principles for inference on aquatic networks. Environmetrics 2014, 25, 306–323. [Google Scholar] [CrossRef]
- Heck, M.P.; Schultz, L.D.; Hockman-Wert, D.; Dinger, E.C.; Dunham, J.B. Monitoring Stream Temperatures—A Guide for Non-Specialists; U.S. Geological Survey: Reston, VA, USA, 2018; Book 3, Chapter A25; p. 76. [Google Scholar]
- Ruesch, A.S.; Torgersen, C.E.; Lawler, J.J.; Olden, J.D.; Peterson, E.E.; Volk, C.J.; Lawrence, D.J. Projected climate-induced habitat loss for salmonids in the John Day River network, Oregon, USA. Conserv. Biol. 2012, 26, 873–882. [Google Scholar] [CrossRef] [PubMed]
- Detenbeck, N.E.; Morrison, A.C.; Abele, R.W.; Kopp, D.A. Spatial statistical network models for stream and river temperature in New England, USA. Water Resour. Res. 2016, 52, 6018–6040. [Google Scholar] [CrossRef]
- Isaak, D.J.; Wenger, S.J.; Peterson, E.E.; Ver Hoef, J.M.; Nagel, D.E.; Luce, C.H.; Hostetler, S.W.; Dunham, J.B.; Roper, B.B.; Wollrab, S.P.; et al. The NorWeST summer stream temperature model and scenarios for the western US: A crowd-sourced database and new geospatial tools foster a user community and predict broad climate warming of rivers and streams. Water Resour. Res. 2017, 53, 9181–9205. [Google Scholar] [CrossRef]
- Gardner, K.K.; McGlynn, B.L. Seasonality in spatial variability and influence of land use/land cover and watershed characteristics on stream water nitrate concentrations in a developing watershed in the Rocky Mountain West. Water Resour. Res. 2009, 45, W08411. [Google Scholar] [CrossRef]
- Brennan, S.R.; Torgersen, C.E.; Hollenbeck, J.P.; Fernandez, D.P.; Jensen, C.K.; Schindler, D.E. Dendritic network models: Improving isoscapes and quantifying influence of landscape and in-stream processes on strontium isotopes in rivers. Geophys. Res. Lett. 2016, 43, 5043–5051. [Google Scholar] [CrossRef]
- Frieden, J.C.; Peterson, E.E.; Webb, J.A.; Negus, P.M. Improving the predictive power of spatial statistical models of stream macroinvertebrates using weighted autocovariance functions. Environ. Model. Softw. 2014, 60, 320–330. [Google Scholar] [CrossRef]
- Isaak, D.J.; Peterson, E.E.; Hoef, J.M.V.; Wenger, S.J.; Falke, J.A.; Torgersen, C.E.; Sowder, C.; Steel, E.A.; Fortin, M.J.; Jordan, C.E.; et al. Applications of spatial statistical network models to stream data. Wiley Interdiscip. Rev. Water 2014, 1, 277–294. [Google Scholar] [CrossRef]
- Ver Hoef, J.M.; Peterson, E.E. A moving average approach for spatial statistical models of stream networks. J. Am. Stat. Assoc. 2010, 105, 6–18. [Google Scholar] [CrossRef]
- Gendaszek, A.; Hockman-Wert, D.; Dunham, J.B.; Torgersen, C.E. Stream Temperature and Water Presence Models of Willow/Whitehorse and Willow/Rock Watersheds, Oregon and Nevada; U.S. Geological Survey: Reston, VA, USA, 2020. [Google Scholar] [CrossRef]
- Fielding, A.H.; Bell, J.F. A review of methods for the assessment of prediction errors in conservation presence/absence models. Environ. Conserv. 1997, 24, 38–49. [Google Scholar] [CrossRef]
- Akaike, H. A new look at the statistical model identification. IEEE Trans. Automat. Contr. 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Constantz, J.; Stonestrom, D.; Stewart, A.E.; Niswonger, R.; Smith, T.R. Analysis of streambed temperatures in ephemeral channels to determine streamflow frequency and duration. Water Resour. Res. 2001, 37, 317–328. [Google Scholar] [CrossRef]
- Blasch, K.W.; Ferre, T.P.A.; Hoffmann, J.P. A statistical technique for interpreting streamflow timing using streambed sediment thermographs. Vadose Zone J. 2005, 3, 936–946. [Google Scholar] [CrossRef]
- Webb, B.W.; Hannah, D.M.; Moore, R.D.; Brown, L.E.; Nobilis, F. Recent advances in stream and river temperature research. Hydrol. Process. 2008, 22, 902–918. [Google Scholar] [CrossRef]
- Helsel, D.R.; Hirsch, R.M. Statistical Methods in Water Resources; U.S. Geological Survey: Reston, VA, USA, 2002; Book 4, Chapter A3. [Google Scholar]
- Steel, E.A.; Beechie, T.J.; Torgersen, C.E.; Fullerton, A.H. Envisioning, quantifying, and managing thermal regimes on river networks. Bioscience 2017, 67, 506–522. [Google Scholar] [CrossRef]
- Jackson, F.L.; Fryer, R.J.; Hannah, D.M.; Malcolm, I.A. Can spatial statistical river temperature models be transferred between catchments? Hydrol. Earth Syst. Sci. 2017, 21, 4727–4745. [Google Scholar] [CrossRef]
- Sinokrot, B.A.; Stefan, H.G. Stream temperature dynamics—measurements and modeling. Water Resour. Res. 1993, 29, 2299–2312. [Google Scholar] [CrossRef]
- Hausner, M.B.; Huntington, J.L.; Nash, C.; Morton, C.; McEvoy, D.J.; Pilliod, D.S.; Hegewisch, K.C.; Daudert, B.; Abatzoglou, J.T.; Grant, G. Assessing the effectiveness of riparian restoration projects using Landsat and precipitation data from the cloud-computing application ClimateEngine.org. Ecol. Eng. 2018, 120, 432–440. [Google Scholar] [CrossRef]
- Constantz, J. Interaction between stream temperature, streamflow, and groundwater exchanges in Alpine streams. Water Resour. Res. 1998, 34, 1609–1615. [Google Scholar] [CrossRef]
- Senay, G.B. Satellite psychrometric formulation of the operational Simplified Surface Energy Balance (SSEBop) model for quantifying and mapping evapotranspiration. Appl. Eng. Agric. 2018, 34, 555–566. [Google Scholar] [CrossRef]
- Senay, G.; Gowda, P.H.; Bohms, S.; Howell, T.; Friedrichs, M.; Marek, T.; Verdin, J. Evaluating the SSEBop approach for evapotranspiration mapping with landsat data using lysimetric observations in the semi-arid Texas High Plains. Hydrol. Earth Syst. Sci. Discuss. 2014, 11, 723–756. [Google Scholar] [CrossRef]
- U.S. Geological Survey. National Water Information System: U.S. Geological Survey web interface. Available online: https://waterdata.usgs.gov/nwis (accessed on 1 October 2019).
- Nagel, D.; Peterson, E.; Isaak, D.; Ver Hoef, J.; Horan, D. National Stream Internet Protocol and User Guide; U.S. Forest Service, Rocky Mountain Research Station Air, Water, and Aquatic Environments Program: Boise, ID, USA, 2015. [Google Scholar]
- McKay, L.; Bondelid, T.; Dewald, T.; Johnston, J.; Moore, R.; Rea, A. NHDPlus Version 2: User Guide; United States Environmental Protection Agency: Washington, DC, USA, 2012. [Google Scholar]
- Peterson, E.E.; Hoef, J.M.V. STARS: An ArcGIS toolset used to calculate the spatial information needed to fit spatial statistical models to stream network data. J. Stat. Softw. 2014, 56, 1–17. [Google Scholar] [CrossRef]
- Hoef, J.M.V.; Peterson, E.E.; Clifford, D.; Shah, R. SSN: An R package for spatial statistical modeling on stream networks. J. Stat. Softw. 2014, 56, 1–45. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014. [Google Scholar]
- Snyder, C.D.; Hitt, N.P.; Young, J.A. Accounting for groundwater in stream fish thermal habitat responses to climate change. Ecol. Appl. 2015, 25, 1397–1419. [Google Scholar] [CrossRef] [PubMed]
- Vatland, S.J.; Gresswell, R.E.; Poole, G.C. Quantifying stream thermal regimes at multiple scales: Combining thermal infrared imagery and stationary stream temperature data in a novel modeling framework. Water Resour. Res. 2015, 51, 31–46. [Google Scholar] [CrossRef]
- Paillex, A.; Siebers, A.R.; Ebi, C.; Mesman, J.; Robinson, C.T. High stream intermittency in an alpine fluvial network: Val Roseg, Switzerland. Limnol. Oceanogr. 2020, 65, 557–568. [Google Scholar] [CrossRef]
- Frissell, C.A.; Liss, W.J.; Warren, C.E.; Hurley, M.D. A hierarchical framework for stream habitat classification: Viewing streams in a watershed context. Environ. Manag. 1986, 10, 199–214. [Google Scholar] [CrossRef]
- Torgersen, C.E.; Ebersole, J.L.; Keenan, D.M. Primer for Identifying Cold-Water Refuges to Protect and Restore Thermal Diversity in Riverine Landscapes; U.S. Environmental Protection Agency: Seattle, WA, USA, 2012. [Google Scholar]
- Falke, J.A.; Dunham, J.B.; Jordan, C.E.; McNyset, K.M.; Reeves, G.H. Spatial ecological processes and local factors predict the distribution and abundance of spawning by steelhead (Oncorhynchus mykiss) across a complex riverscape. PLoS ONE 2013, 8, e79232. [Google Scholar] [CrossRef]
- McNyset, K.M.; Volk, C.J.; Jordan, C.E. Developing an effective model for predicting spatially and temporally continuous stream temperatures from remotely sensed land surface temperatures. Water 2015, 7, 6827–6846. [Google Scholar] [CrossRef]
- Jaeger, K.; Sando, R.; McShane, R.R.; Dunham, J.B.; Hockman-Wert, D.; Kaiser, K.E.; Hafen, K.; Risley, J.; Blasch, K. Probability of Streamflow Permanence Model (PROSPER): A spatially continuous model of annual streamflow permanence throughout the Pacific Northwest. J. Hydrol. X 2019, 2, 100005. [Google Scholar] [CrossRef]
- Dahm, C.N.; Cleverly, J.R.; Coonrod, J.E.A.; Thibault, J.R.; McDonnell, D.E.; Gilroy, D.F. Evapotranspiration at the land/water interface in a semi-arid drainage basin. Freshw. Biol. 2002, 47, 831–843. [Google Scholar] [CrossRef]
- Nash, C.S.; Selker, J.S.; Grant, G.E.; Lewis, S.L.; Noel, P. A physical framework for evaluating net effects of wet meadow restoration on late-summer streamflow. Ecohydrology 2018, 11, e1953. [Google Scholar] [CrossRef]
- Pettorelli, N.; Vik, J.O.; Mysterud, A.; Gaillard, J.M.; Tucker, C.J.; Stenseth, N.C. Using the satellite-derived NDVI to assess ecological responses to environmental change. Trends Ecol. Evol. 2005, 20, 503–510. [Google Scholar] [CrossRef]
- Minshall, G.W.; Jensen, S.E.; Platts, W.S. The Ecology of Stream and Riparian Habitats of the Great Basin Region: A Community Profile; Natonal Wetlands Research Center: Slidell, LA, USA, 1989. [Google Scholar]
- Caissie, D. The thermal regime of rivers: A review. Freshw. Biol. 2006, 51, 1389–1406. [Google Scholar] [CrossRef]
- Wondzell, S.M.; Diabat, M.; Haggerty, R. What matters most: Are future stream temperatures more sensitive to changing air temperatures, discharge, or riparian vegetation? J. Am. Water Resour. Assoc. 2019, 55, 116–132. [Google Scholar] [CrossRef]
- Cartwright, J.; Johnson, H.M. Springs as hydrologic refugia in a changing climate? A remote-sensing approach. Ecosphere 2018, 9, e02155. [Google Scholar] [CrossRef]
- Donnelly, J.P.; Naugle, D.E.; Hagen, C.A.; Maestas, J.D. Public lands and private waters: Scarce mesic resources structure land tenure and sage-grouse distributions. Ecosphere 2016, 7, e01208. [Google Scholar] [CrossRef]
- Donnelly, J.P.; Allred, B.W.; Perret, D.; Silverman, N.L.; Tack, J.D.; Dreitz, V.J.; Maestas, J.D.; Naugle, D.E. Seasonal drought in North America's sagebrush biome structures dynamic mesic resources for sage-grouse. Ecol. Evol. 2018, 8, 12492–12505. [Google Scholar] [CrossRef]
- Eidenshink, J.; Schwind, B.; Brewer, K.; Zhu, Z.-L.; Quayle, B.; Howard, S. A project for monitoring trends in burn severity. Fire Ecol. 2007, 3, 3–21. [Google Scholar] [CrossRef]
- Silverman, N.L.; Allred, B.W.; Donnelly, J.P.; Chapman, T.B.; Maestas, J.D.; Wheaton, J.M.; White, J.; Naugle, D.E. Low-tech riparian and wet meadow restoration increases vegetation productivity and resilience across semiarid rangelands. Restor. Ecol. 2019, 27, 269–278. [Google Scholar] [CrossRef]
- Shi, H.; Rigge, M.; Homer, C.G.; Xian, G.; Meyer, D.K.; Bunde, B. Historical cover trends in a sagebrush steppe ecosystem from 1985 to 2013: Links with climate, disturbance, and management. Ecosystems 2018, 21, 913–929. [Google Scholar] [CrossRef]
- Dauwalter, D.C.; Fesenmyer, K.A.; Miller, S.W.; Porter, T. Response of Riparian Vegetation, Instream Habitat, and Aquatic Biota to Riparian Grazing Exclosures. N. Am. J. Fish. Manag. 2018, 38, 1187–1200. [Google Scholar] [CrossRef]
- Neville, H.M.; Leasure, D.R.; Dauwalter, D.C.; Dunham, J.B.; Bjork, R.; Fesenmyer, K.A.; Chelgren, N.D.; Peacock, M.M.; Luce, C.H.; Isaak, D.J.; et al. Application of multiple-population viability analysis to evaluate species recovery alternatives. Conserv. Biol. 2020, 34, 482–493. [Google Scholar] [CrossRef] [PubMed]
- Milly, P.C.; Betancourt, J.; Falkenmark, M.; Hirsch, R.M.; Kundzewicz, Z.W.; Lettenmaier, D.P.; Stouffer, R.J. Stationarity is dead: Whither water management? Science 2008, 319, 573–574. [Google Scholar] [CrossRef] [PubMed]
- Jencso, K.G.; McGlynn, B.L.; Gooseff, M.N.; Wondzell, S.M.; Bencala, K.E.; Marshall, L.A. Hydrologic connectivity between landscapes and streams: Transferring reach-and plot-scale understanding to the catchment scale. Water Resour. Res. 2009, 45, W04428. [Google Scholar] [CrossRef]
- Ward, A.S.; Schmadel, N.M.; Wondzell, S.M. Simulation of dynamic expansion, contraction, and connectivity in a mountain stream network. Adv. Water Resour. 2018, 114, 64–82. [Google Scholar] [CrossRef]
- Raheem, N.; Cravens, A.E.; Cross, M.S.; Crausbay, S.; Ramirez, A.; McEvoy, J.; Zoanni, D.; Bathke, D.J.; Hayes, M.; Carter, S.; et al. Planning for ecological drought: Integrating ecosystem services and vulnerability assessment. Wiley Interdiscip. Rev. Water 2019, 6, e1352. [Google Scholar] [CrossRef]
- Diffenbaugh, N.S.; Swain, D.L.; Touma, D. Anthropogenic warming has increased drought risk in California. Proc. Natl. Acad. Sci. USA 2015, 112, 3931–3936. [Google Scholar] [CrossRef]
- Bogan, M.T.; Boersma, K.S.; Lytle, D.A. Resistance and resilience of invertebrate communities to seasonal and supraseasonal drought in arid-land headwater streams. Freshw. Biol. 2015, 60, 2547–2558. [Google Scholar] [CrossRef]
- Lynch, D.T.; Leasure, D.R.; Magoulick, D.D. The influence of drought on flow-ecology relationships in Ozark Highland streams. Freshw. Biol. 2018, 63, 946–968. [Google Scholar] [CrossRef]
- Morelli, T.L.; Daly, C.; Dobrowski, S.Z.; Dulen, D.M.; Ebersole, J.L.; Jackson, S.T.; Lundquist, J.D.; Millar, C.I.; Maher, S.P.; Monahan, W.B.; et al. Managing climate change refugia for climate adaptation. PLoS ONE 2016, 11, e0159909. [Google Scholar] [CrossRef] [PubMed]
- Beever, E.A.; Hall, L.E.; Varner, J.; Loosen, A.E.; Dunham, J.B.; Gahl, M.K.; Smith, F.A.; Lawler, J.J. Behavioral flexibility as a mechanism for coping with climate change. Front. Ecol. Environ. 2017, 15, 299–308. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Class | Covariate | Abbreviation | Description | Hypothesis (Water Temperature) | Hypothesis (Flow Presence) | Data Source |
|---|---|---|---|---|---|---|
| Land cover | Riparian NDVI (dimensionless) | NDVIrip | Seasonally averaged (1 May–30 August) NDVI for individual years. Values averaged within 200-m diameter riparian buffer upstream of site. | (−) Higher NDVI indicates increased vegetation, which may limit gains from solar insolation and indicates shallow groundwater input thus cooling stream temperature. | (+) Higher NDVI indicates increased shallow groundwater input and should be positively related to flow presence. | Earth Engine SSEBop ET Model. LANDSAT 8 (30-m resolution) |
| Climatic, Land cover | Riparian ET (mm/month) | ETrip | Seasonally averaged (1 May–1 August) ET for individual years. Values averaged within 200-m diameter riparian buffer upstream of site. | (−) Higher ET is associated with warmer air and water temperatures but ET cools stream temperature through evaporative heat loss. | (−) Higher ET removes water from the stream and riparian corridor and should be negatively related to flow presence. | Earth Engine SSEBop ET Model. LANDSAT 8 (30-m resolution) |
| Climatic | Watershed averaged May 1 SWE (categorical) | SWEws | Categorical variable of snow water equivalent (SWE) in contributing watershed of each site for individual years. Presence of snowmelt is associated with increased runoff and flow presence. | (−) Higher SWE should increase cool runoff and result in cooler stream temperature. | (+) Higher SWE should increase cool runoff and be positively related to flow presence. | National Operational Hydrologic Remote Sensing Center (2004) (1-km resolution) |
| Climatic | Monthly mean discharge (m3/km2) | MonthQ | Monthly mean discharge for individual years normalized by contributing basin area for nearby gages. | (−) Higher monthly discharge should correspond with lower stream temperature. | (+) Higher mean monthly discharge should correspond with increased flow presence. | USGS–NWIS |
| Climatic | Monthly mean air temperature (°C) | MonthTa | Basin mean monthly mean air temperature for individual years. Values were averaged over WW and WR watersheds. | (+) Higher monthly air temperature should correspond with higher stream temperature. | (−) Higher mean monthly air temperature should correspond with decreased discharge and be negatively related to flow presence. | PRISM |
| Watershed | Day | 2015 Count | 2016 Count | 2017 Count | 2015–2016 Change in State | 2016–2017 Change in State | |||
|---|---|---|---|---|---|---|---|---|---|
| Wet | Dry | Wet | Dry | Wet | Dry | Proportion of Sites | Proportion of Sites | ||
| WW | 15 May | 52 | 50 | 63 | 21 | 54 | 19 | 0.27 | 0.08 |
| WR | 15 May | n.m. | n.m. | 50 | 45 | 39 | 35 | n.m. | 0.15 |
| WW | 15 June | 71 | 30 | 61 | 23 | 50 | 23 | 0.07 | 0.1 |
| WR | 15 June | n.m. | n.m. | 54 | 41 | 35 | 39 | n.m. | 0.09 |
| WW | 15 July | 66 | 35 | 56 | 28 | 45 | 28 | 0.1 | 0.05 |
| WR | 15 July | n.m. | n.m. | 37 | 58 | 34 | 40 | n.m. | 0.16 |
| WW | 15 August | 64 | 37 | 56 | 28 | 47 | 26 | 0.09 | 0.11 |
| WR | 15 August | n.m. | n.m. | 17 | 57 | 28 | 46 | n.m. | 0.2 |
| Response Variable | ||||
| MayTw | JunTw | JulTw | AugTw | |
| Sample size | 244 | 250 | 215 | 193 |
| Spatial Models | ||||
| y-intercept | 12.90 *** | - | 45.50 *** | 47.80 *** |
| Covariate coefficient (significance) | ||||
| Riparian NDVI (NDVIrip) | −7.44 *** | −7.54 * | −15.82 *** | −14.72 *** |
| Riparian ET (ETrip) | - | - | - | - |
| Watershed May 1 SWE (SWEws) | −0.38 ** | −0.45 * | - | - |
| Monthly Mean Discharge (MonthQ) | −103 * | - | 13,308 *** | 23,664 ** |
| Monthly Mean Air Temperature (MonthTa) | - | 0.59 *** | −1.28 *** | −1.38 * |
| Covariance components (fraction variance explained) | ||||
| Covariates | 0.24 | 0.57 | 0.24 | 0.16 |
| Tail-up | 0.05 | 0.06 | 0.14 | 0.48 |
| Tail-down | 0 | 0.07 | 0.04 | 0.12 |
| Euclidean | 0.67 | 0.27 | 0.54 | 0 |
| Site | 0 | 0 | 0 | 0.13 |
| Year | 0 | 0 | 0 | 0 |
| Nugget | 0.03 | 0.04 | 0.04 | 0.1 |
| Model performance | ||||
| r2pred | 0.89 | 0.83 | 0.76 | 0.66 |
| RMSPE | 0.7 | 1.21 | 1.5 | 1.72 |
| MAPE | 0.49 | 0.82 | 0.95 | 1.03 |
| AIC | 588 | 885 | 832 | 794 |
| Nonspatial Models | ||||
| r2pred | 0.75 | 0.74 | 0.71 | 0.64 |
| RMSPE | 1.04 | 1.49 | 1.65 | 1.77 |
| MAPE | 0.76 | 1.1 | 1.12 | 1.13 |
| AIC | 775 | 991 | 896 | 808 |
| Variance Inflation Factor | ||||
| Riparian NDVI (NDVIrip) | 2.8 | 3.3 | 2.8 | 2.6 |
| Riparian ET (ETrip) | 2.4 | 2.7 | 2.7 | 2.5 |
| Watershed May 1 SWE (SWEws) | 1.4 | 1.4 | 1.3 | 1.2 |
| Monthly Mean Discharge (MonthQ) | 1.2 | 2.4 | 3.1 | 7.6 |
| Monthly Mean Air Temperature (MonthTa) | 1.4 | 3.4 | 3.5 | 7.7 |
| Response Variable | ||||
| May15Wet | Jun15Wet | Jul15Wet | Aug15Wet | |
| Sample size | 429 | 428 | 428 | 406 |
| Spatial Models | ||||
| y-intercept | −5.8 *** | −1.8 *** | −4.9 *** | −3.5 *** |
| Covariate coefficient (significance) | ||||
| Riparian NDVI (NDVIrip) | 21.0 *** | 4.4 ** | 15.4 *** | 7.8 *** |
| Riparian ET (ETrip) | - | - | - | - |
| Watershed May 1 SWE (SWEws) | 1.9 ** | - | - | - |
| Monthly Mean Discharge (MonthQ) | - | - | - | - |
| Monthly Mean Air Temperature (MonthTa) | - | - | - | - |
| Model performance | ||||
| AUC | 88.6 | 94.2 | 93.3 | 92.6 |
| Nonspatial Models | ||||
| AUC | 85.1 | 90.8 | 87.2 | 85.2 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gendaszek, A.S.; Dunham, J.B.; Torgersen, C.E.; Hockman-Wert, D.P.; Heck, M.P.; Thorson, J.; Mintz, J.; Allai, T. Land-Cover and Climatic Controls on Water Temperature, Flow Permanence, and Fragmentation of Great Basin Stream Networks. Water 2020, 12, 1962. https://doi.org/10.3390/w12071962
Gendaszek AS, Dunham JB, Torgersen CE, Hockman-Wert DP, Heck MP, Thorson J, Mintz J, Allai T. Land-Cover and Climatic Controls on Water Temperature, Flow Permanence, and Fragmentation of Great Basin Stream Networks. Water. 2020; 12(7):1962. https://doi.org/10.3390/w12071962
Chicago/Turabian StyleGendaszek, Andrew S., Jason B. Dunham, Christian E. Torgersen, David P. Hockman-Wert, Michael P. Heck, Justin Thorson, Jeffrey Mintz, and Todd Allai. 2020. "Land-Cover and Climatic Controls on Water Temperature, Flow Permanence, and Fragmentation of Great Basin Stream Networks" Water 12, no. 7: 1962. https://doi.org/10.3390/w12071962
APA StyleGendaszek, A. S., Dunham, J. B., Torgersen, C. E., Hockman-Wert, D. P., Heck, M. P., Thorson, J., Mintz, J., & Allai, T. (2020). Land-Cover and Climatic Controls on Water Temperature, Flow Permanence, and Fragmentation of Great Basin Stream Networks. Water, 12(7), 1962. https://doi.org/10.3390/w12071962