Distribution and Drivers of Marine Isoprene Concentration across the Southern Ocean

 , , , , ,
, , , , ,  , ,
, ,  and
and
Abstract

1. Introduction
2. Materials and Methods
2.1. The PEGASO, TransPEGASO and ACE Cruises
2.2. Isoprene Concentration Measurements
2.3. Biological, Physical, and Environmental Variables
2.4. Other Data Sources in the Southern Ocean
2.5. Statistical Analysis and Model Development
3. Results and Discussion
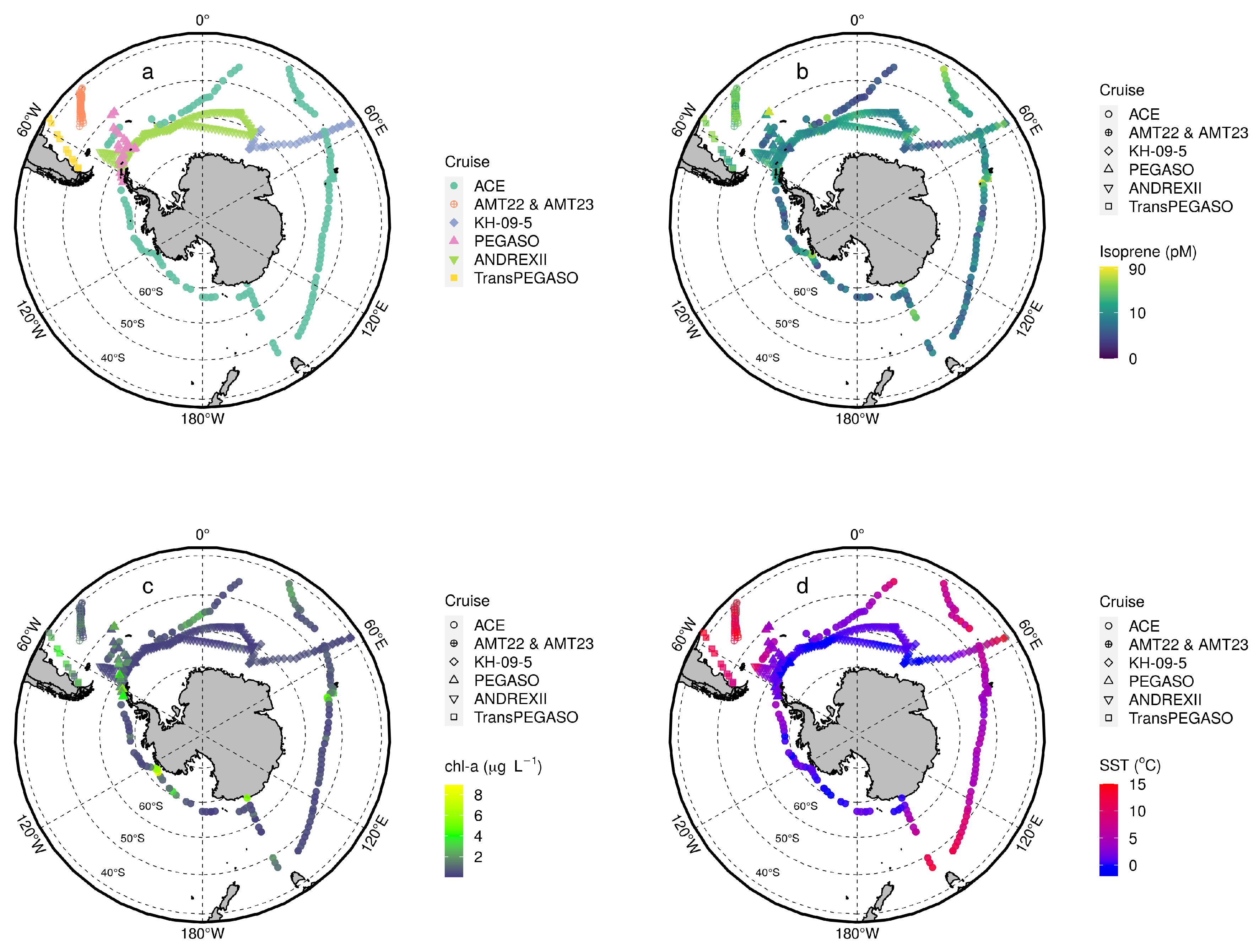
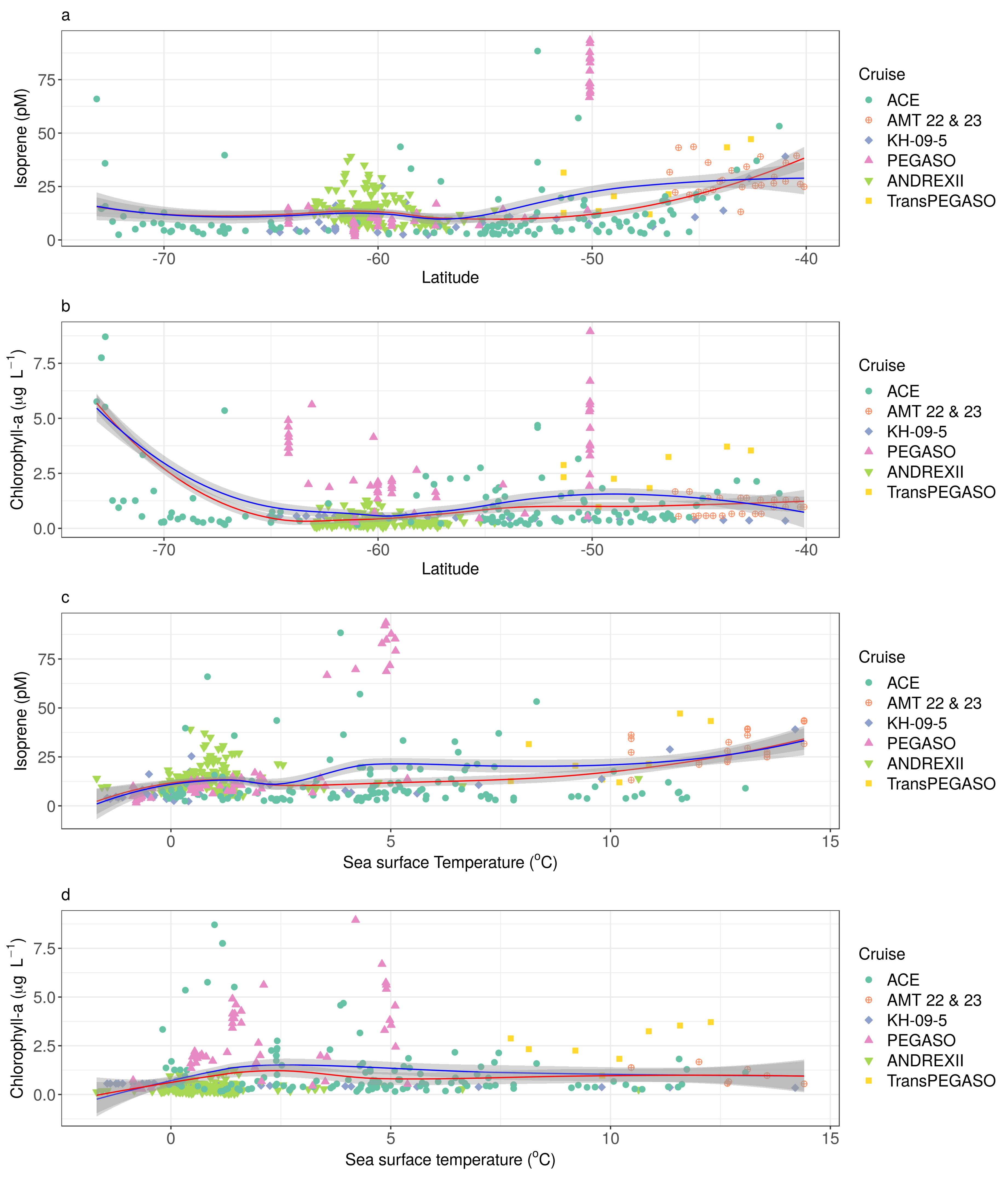
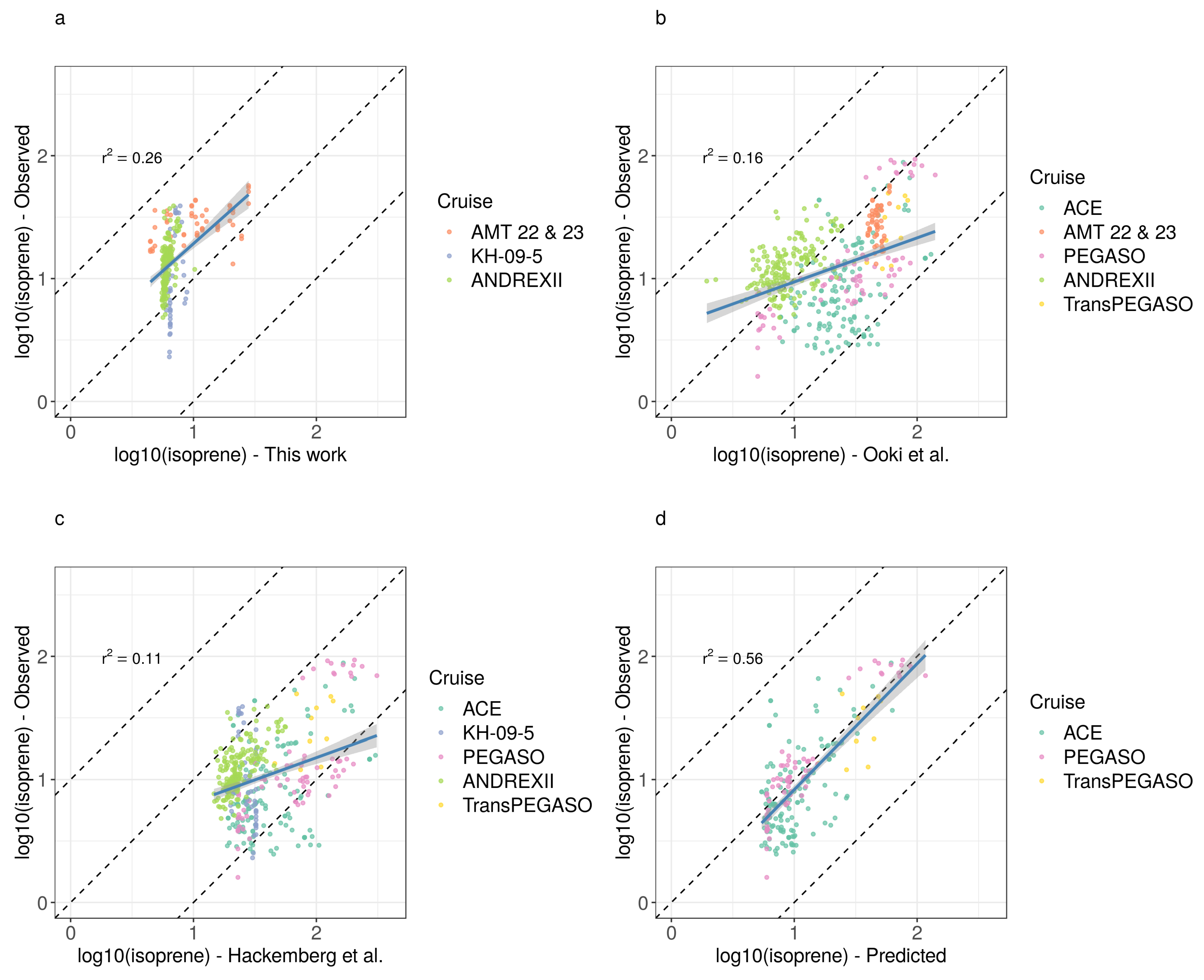
3.1. General Patterns of Isoprene Surface Concentration in the Southern Ocean
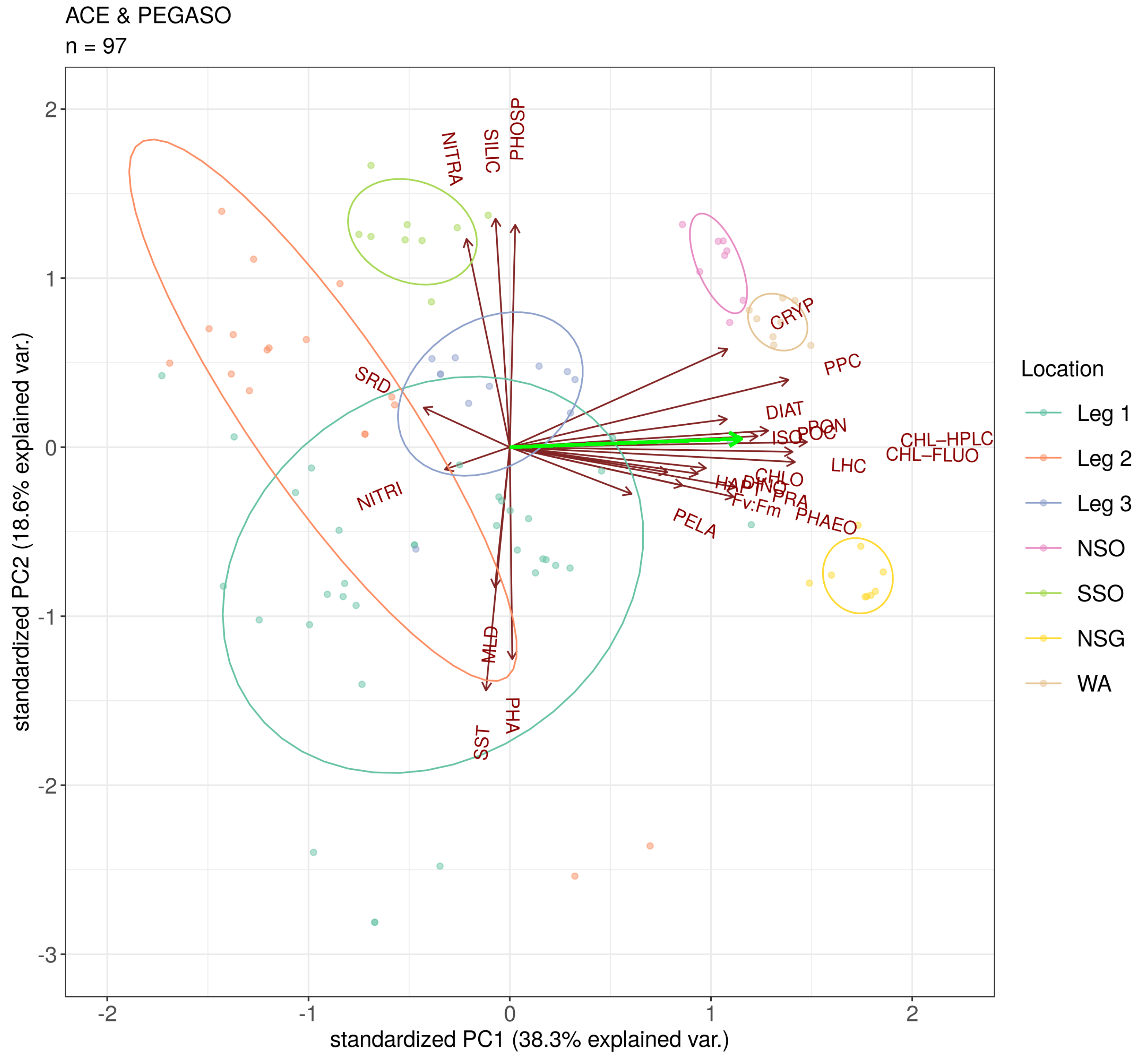
3.2. Drivers of Isoprene Concentration in the Southern Ocean
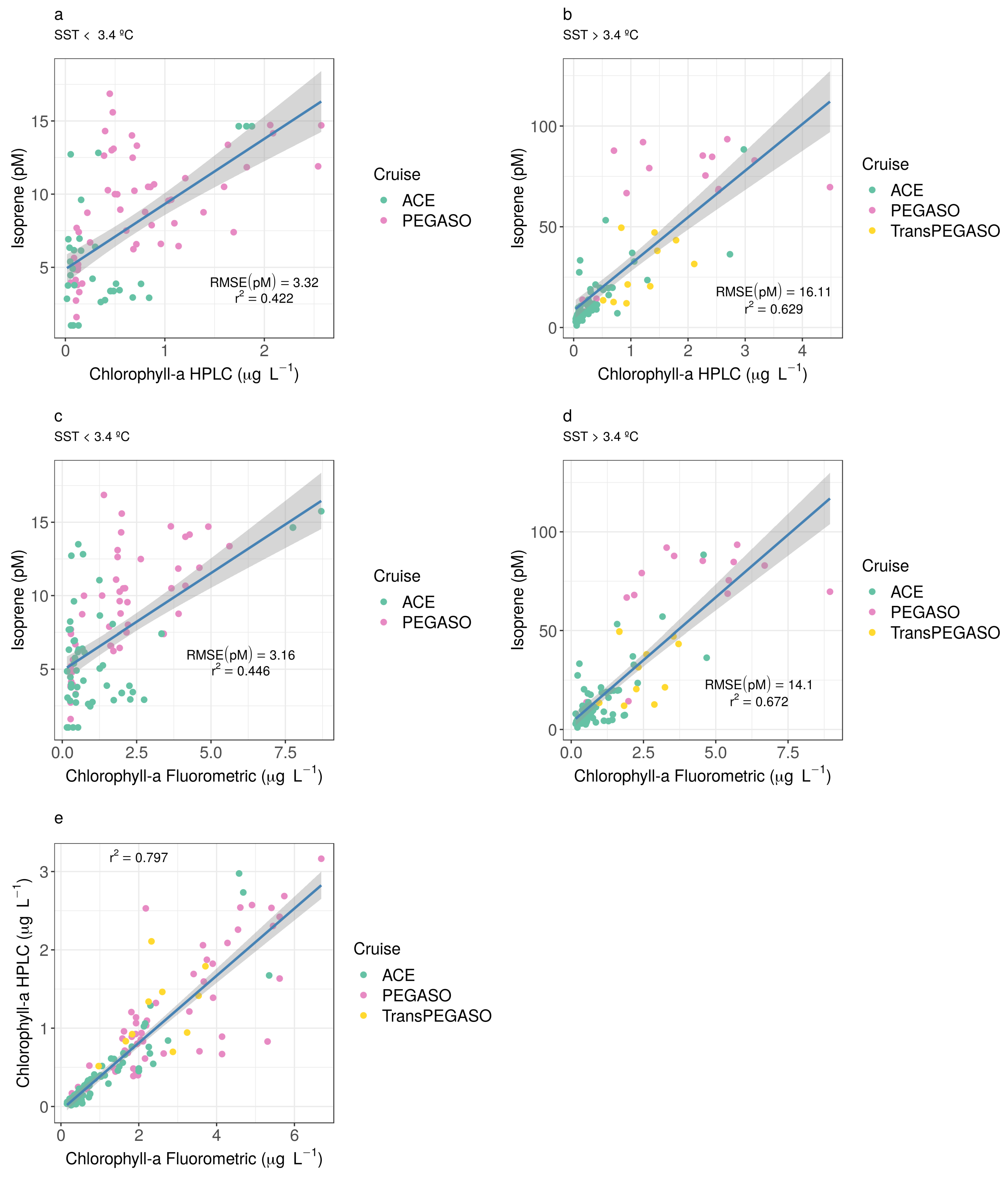
3.3. The Predictive Capacity of Chlorophyll-a and Sea Surface Temperature to Isoprene Concentration in the Southern Ocean
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A. Summary of Compiled Cruise Variables

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isoprene (pM) | Chlorophyll-a (g ) | Sea Surface Temperature (C) | Southern Ocean Area | Cruise |
|---|---|---|---|---|
| Mean (Min–Max) | Mean (Min–Max) | Mean (Min–Max) | ||
| 10.7 (2.1–88.4) | 1.46 (0.15–8.70 ) | 4.16 (−0.18–13.06) | SO Circumnavigation | ACE |
| 22.4 (1.6–93.5) | 2.42 (0.28–8.95) | 1.45 (−0.87–5.38) | SO and Weddell Sea | PEGASO |
| 25.3 (12.0–49.5) | 2.59 (0.97–3.71) | 9.97 (7.73–12.28) | Southwestern Atlantic Self | TransPEGASO |
| 29.0 (13.1–57.1) | 0.97 (0.55–1.67) | 12.68 (10.46–14.40) | SO + South Atlantic Ocean | AMT23 & AMT22 |
| 9.5 (2.3–39.0) | 0.49 (0.34–0.56) | 1.82 (−1.45–14.2) | SO + South Indian Ocean | KH-09-5 |
| 13.5 (4.8–39.1) | 0.33 (0.02–1.27) | 0.91 (−1.68–10.63) | SO + South Atlantic Ocean | ANDREXII |
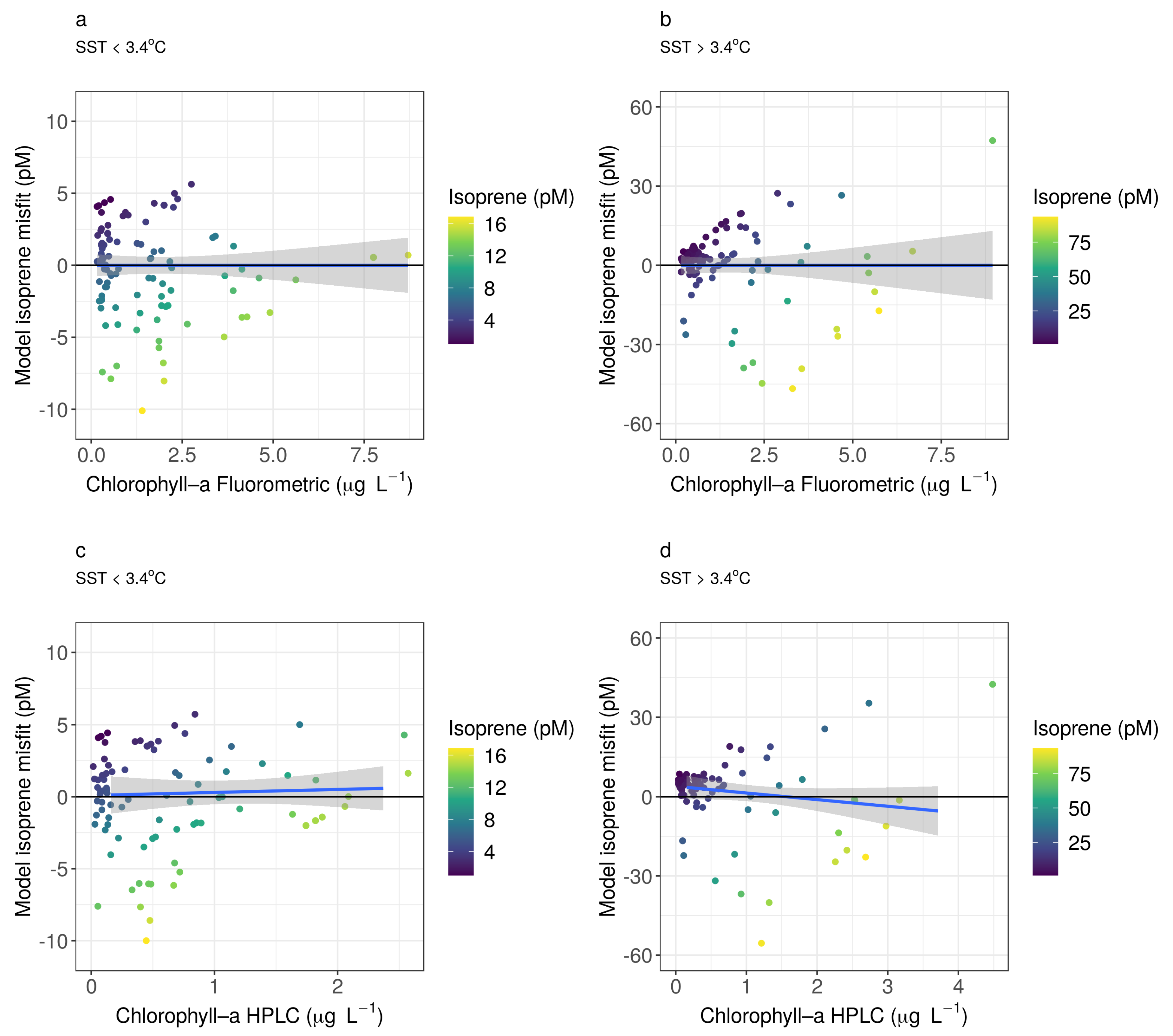
Appendix B. Residuals

Appendix C. Statistical Relationships between Isoprene Concentration and Chlorophyll-a Concentration and Sea Surface Temperature

References
- Carpenter, L.J.; Archer, S.D.; Beale, R. Ocean-atmosphere trace gas exchange. Chem. Soc. Rev. 2012, 41, 6473–6506. [Google Scholar] [CrossRef] [PubMed]
- Shaw, S.L.; Chisholm, S.W.; Prinn, R.G. Isoprene production by Prochlorococcus, a marine cyanobacterium, and other phytoplankton. Mar. Chem. 2003, 80, 227–245. [Google Scholar] [CrossRef]
- Exton, D.; Suggett, D.J.; McGenity, T.J.; Steinke, M. Chlorophyll-normalized isoprene production in laboratory cultures of marine microalgae and implications for global models. Limnol. Oceanogr. 2013, 58, 1301–1311. [Google Scholar] [CrossRef]
- Meskhidze, N.; Nenes, A. Phytoplankton and cloudiness in the Southern Ocean. Science 2006, 314, 1419–1423. [Google Scholar] [CrossRef] [PubMed]
- Arnold, S.; Spracklen, D.; Williams, J.; Yassaa, N.; Sciare, J.; Bonsang, B.; Gros, V.; Peeken, I.; Lewis, A.; Alvain, S.; et al. Evaluation of the global oceanic isoprene source and its impacts on marine organic carbon aerosol. Atmos. Chem. Phys. 2009, 9, 1253–1262. [Google Scholar] [CrossRef]
- Vallina, S.; Simó, R.; Gassó, S.; de Boyer-Montégut, C.; Del Río, E.; Jurado, E.; Dachs, J. Analysis of a potential “solar radiation dose–dimethylsulfide–cloud condensation nuclei” link from globally mapped seasonal correlations. Glob. Biogeochem. Cycles 2007, 21, 0886–6236. [Google Scholar]
- Dani, K.S.; Loreto, F. Trade-off between dimethyl sulfide and isoprene emissions from marine phytoplankton. Trends Plant Sci. 2017, 22, 361–372. [Google Scholar]
- Luo, G.; Yu, F. A numerical evaluation of global oceanic emissions of α-pinene and isoprene. Atmos. Chem. Phys. 2010, 10, 2007–2015. [Google Scholar] [CrossRef]
- Booge, D.; Schlundt, C.; Bracher, A.; Endres, S.; Zäncker, B.; Marandino, C.A. Marine isoprene production and consumption in the mixed layer of the surface ocean—A field study over two oceanic regions. Biogeosciences (BG) 2018, 15, 649–667. [Google Scholar]
- Alvarez, L.A.; Exton, D.A.; Timmis, K.N.; Suggett, D.J.; McGenity, T.J. Characterization of marine isoprene-degrading communities. Environ. Microbiol. 2009, 11, 3280–3291. [Google Scholar] [CrossRef]
- Ciuraru, R.; Fine, L.; Pinxteren, M.v.; D’Anna, B.; Herrmann, H.; George, C. Unravelling new processes at interfaces: Photochemical isoprene production at the sea surface. Environ. Sci. Technol. 2015, 49, 13199–13205. [Google Scholar] [CrossRef] [PubMed]
- Brüggemann, M.; Hayeck, N.; George, C. Interfacial photochemistry at the ocean surface is a global source of organic vapors and aerosols. Nat. Commun. 2018, 9, 2101. [Google Scholar] [CrossRef] [PubMed]
- Ooki, A.; Nomura, D.; Nishino, S.; Kikuchi, T.; Yokouchi, Y. A global-scale map of isoprene and volatile organic iodine in surface seawater of the Arctic, Northwest Pacific, Indian, and Southern Oceans. J. Geophys. Res. Ocean. 2015, 120, 4108–4128. [Google Scholar] [CrossRef]
- Booge, D.; Marandino, C.A.; Schlundt, C.; Palmer, P.I.; Schlundt, M.; Atlas, E.L.; Bracher, A.; Saltzman, E.S.; Wallace, D.W. Can simple models predict large-scale surface ocean isoprene concentrations? Atmos. Chem. Phys. 2016, 16, 11807–11821. [Google Scholar] [CrossRef]
- Hackenberg, S.; Andrews, S.; Airs, R.; Arnold, S.; Bouman, H.; Brewin, R.; Chance, R.; Cummings, D.; Dall’Olmo, G.; Lewis, A.; et al. Potential controls of isoprene in the surface ocean. Glob. Biogeochem. Cycles 2017, 31, 644–662. [Google Scholar] [CrossRef]
- Palmer, P.I.; Shaw, S.L. Quantifying global marine isoprene fluxes using MODIS chlorophyll observations. Geophys. Res. Lett. 2005, 32. [Google Scholar] [CrossRef]
- Dani, K.S.; Benavides, A.M.S.; Michelozzi, M.; Peluso, G.; Torzillo, G.; Loreto, F. Relationship between isoprene emission and photosynthesis in diatoms, and its implications for global marine isoprene estimates. Mar. Chem. 2017, 189, 17–24. [Google Scholar] [CrossRef]
- Zindler, C.; Marandino, C.A.; Bange, H.W.; Schütte, F.; Saltzman, E.S. Nutrient availability determines dimethyl sulfide and isoprene distribution in the eastern Atlantic Ocean. Geophys. Res. Lett. 2014, 41, 3181–3188. [Google Scholar] [CrossRef][Green Version]
- Kameyama, S.; Tanimoto, H.; Inomata, S.; Tsunogai, U.; Ooki, A.; Takeda, S.; Obata, H.; Tsuda, A.; Uematsu, M. High-resolution measurement of multiple volatile organic compounds dissolved in seawater using equilibrator inlet–proton transfer reaction-mass spectrometry (EI–PTR-MS). Mar. Chem. 2010, 122, 59–73. [Google Scholar] [CrossRef]
- Wohl, C.; Brown, I.; Kitidis, V.; Jones, A.E.; Sturges, W.T.; Nightingale, P.D.; Yang, M. Underway seawater and atmospheric measurements of volatile organic compounds in the Southern Ocean. Biogeosciences 2020, 2020, 2593–2619. [Google Scholar] [CrossRef]
- Dall’Osto, M.; Ovadnevaite, J.; Paglione, M.; Beddows, D.C.; Ceburnis, D.; Cree, C.; Cortés, P.; Zamanillo, M.; Nunes, S.O.; Pérez, G.L.; et al. Antarctic sea ice region as a source of biogenic organic nitrogen in aerosols. Sci. Rep. 2017, 7, 6047. [Google Scholar] [CrossRef] [PubMed]
- Nunes, S.; Latasa, M.; Delgado, M.; Emelianov, M.; Simó, R.; Estrada, M. Phytoplankton community structure in contrasting ecosystems of the Southern Ocean: South Georgia, South Orkneys and western Antarctic Peninsula. Deep Sea Res. Part I Oceanogr. Res. Pap. 2019, 151, 103059. [Google Scholar] [CrossRef]
- Zamanillo, M.; Ortega-Retuerta, E.; Nunes, S.; Estrada, M.; Sala, M.M.; Royer, S.J.; López-Sandoval, D.C.; Emelianov, M.; Vaqué, D.; Marrasé, C.; et al. Distribution of transparent exopolymer particles (TEP) in distinct regions of the Southern Ocean. Sci. Total Environ. 2019, 691, 736–748. [Google Scholar] [CrossRef] [PubMed]
- Henry, T.; Robinson, C.; Haumman, F.; Thomas, J.; Hitchings, J.; Schuback, N.; Tsukernik, M.; Leonard, K. Physical and biogeochemical oceanography data from Conductivity, Temperature, Depth (CTD) rosette deployments during the Antarctic Circumnavigation Expedition (ACE). ACE Exped. Data-Sets 2020. [Google Scholar] [CrossRef]
- Yentsch, C.S.; Menzel, D.W. A method for the determination of phytoplankton chlorophyll and phaeophytin by fluorescence. In Deep Sea Research and Oceanographic Abstracts; Elsevier: Amsterdam, The Netherlands, 1963; Volume 10, pp. 221–231. [Google Scholar]
- Antoine, D.; Thomalla, S.; Berliner, D.; Little, H.; Moutier, W.; Olivier-Morgan, A.; Robinson, C.; Ryan-Keogh, T.; Schuback, N. Phytoplankton pigment concentrations of seawater sampled during the Antarctic Circumnavigation Expedition (ACE) during the Austral Summer of 2016/2017. Zenodo 2019. [Google Scholar] [CrossRef]
- Rodriguez, F.; Varela, M.; Zapata, M. Phytoplankton assemblages in the Gerlache and Bransfield Straits (Antarctic Peninsula) determined by light microscopy and CHEMTAX analysis of HPLC pigment data. Deep Sea Res. Part II Top. Stud. Oceanogr. 2002, 49, 723–747. [Google Scholar] [CrossRef]
- Zapata, M.; Rodríguez, F.; Garrido, J.L. Separation of chlorophylls and carotenoids from marine phytoplankton: A new HPLC method using a reversed phase C8 column and pyridine-containing mobile phases. Mar. Ecol. Prog. Ser. 2000, 195, 29–45. [Google Scholar] [CrossRef]
- Cook, S.S.; Whittock, L.; Wright, S.W.; Hallegraeff, G.M. Photosynthetic pigment and genetic differences between two Southern Ocean morphotypes of Emiliania huxleyi (Haptophyta) 1. J. Phycol. 2011, 47, 615–626. [Google Scholar] [CrossRef]
- Higgins, H.W.; Wright, S.W.; Schluter, L. Quantitative Interpretation of Chemotaxonomic Pigment Data; Cambridge University Press: Cambridge, UK, 2011. [Google Scholar]
- Cassar, N.; Wright, S.W.; Thomson, P.G.; Trull, T.W.; Westwood, K.J.; de Salas, M.; Davidson, A.; Pearce, I.; Davies, D.M.; Matear, R.J. The relation of mixed-layer net community production to phytoplankton community composition in the Southern Ocean. Glob. Biogeochem. Cycles 2015, 29, 446–462. [Google Scholar] [CrossRef]
- Kolber, Z.S.; Prášil, O.; Falkowski, P.G. Measurements of variable chlorophyll fluorescence using fast repetition rate techniques: Defining methodology and experimental protocols. Biochim. Biophys. Acta (BBA)-Bioenerg. 1998, 1367, 88–106. [Google Scholar] [CrossRef]
- Royer, S.J.; Mahajan, A.; Galí, M.; Saltzman, E.; Simó, R. Small-scale variability patterns of DMS and phytoplankton in surface waters of the tropical and subtropical Atlantic, Indian, and Pacific Oceans. Geophys. Res. Lett. 2015, 42, 475–483. [Google Scholar] [CrossRef]
- Ryan-Keogh, T.; Robinson, C. Phytoplankton Photophysiology Utilities: A Python Toolbox for the standardisation of processing active chlorophyll fluorescence data. Front. Mar. Sci. Aquat. Physiol. 2020, submitted. [Google Scholar]
- Gasol, J.M.; Del Giorgio, P.A. Using flow cytometry for counting natural planktonic bacteria and understanding the structure of planktonic bacterial communities. Sci. Mar. 2000, 64, 197–224. [Google Scholar] [CrossRef]
- Hansen, H.; Grasshoff, K. Automated chemical analysis. In Methods of Seawater Analysis; Verlag Chemie Weinheim: Weinheim, Germany, 1983; pp. 347–379. [Google Scholar]
- Wolters, M. Determination of Silicate in Brackish or Seawater by Flow Injection Analysis. QuickChem Method 31-114-27-1-D; Methods Manual; Lachat Instruments: Milwaukee, WI, USA, 2002; 12p. [Google Scholar]
- Egan, L. Determination of Nitrate and/or Nitrite in Brackish or Seawater by Flow Injection Analysis. QuikChem Method 31-107-04-1-C; Lachat Instruments: Milwaukee, WI, USA, 2008. [Google Scholar]
- Grasshoff, K.; Kremling, K.; Ehrhardt, M. Methods of Seawater Analysis; John Wiley & Sons: Hoboken, NJ, USA, 2009. [Google Scholar]
- Schlitzer, R.; Anderson, R.F.; Dodas, E.M.; Lohan, M.; Geibert, W.; Tagliabue, A.; Bowie, A.; Jeandel, C.; Maldonado, M.T.; Landing, W.M.; et al. The GEOTRACES Intermediate Data Product 2017. Chem. Geol. 2018, 493, 210–223. [Google Scholar] [CrossRef]
- Wohl, C.; Capelle, D.; Jones, A.; Sturges, W.T.; Nightingale, P.D.; Else, B.G.; Yang, M. Segmented flow coil equilibrator coupled to a Proton Transfer Reaction Mass Spectrometer for measurements of a broad range of Volatile Organic Compounds in seawater. Ocean Sci. 2019, 15, 925–940. [Google Scholar] [CrossRef]
- RStudio Team. RStudio: Integrated Development Environment for R; RStudio, Inc.: Boston, MA, USA, 2015. [Google Scholar]
- Rodríguez-Ros, P.; Galí, M.; Cortés, P.; Robinson, C.M.; Antoine, D.; Wohl, C.; Yang, M.; Simó, R. Remote sensing retrieval of isoprene concentrations in the Southern Ocean. Under Rev.—Geophys. Res. Lett. 2020, in press. [Google Scholar] [CrossRef]
- Suggett, D.J.; Moore, C.M.; Hickman, A.E.; Geider, R.J. Interpretation of fast repetition rate (FRR) fluorescence: Signatures of phytoplankton community structure versus physiological state. Mar. Ecol. Prog. Ser. 2009, 376, 1–19. [Google Scholar] [CrossRef]
- Behrenfeld, M.J.; Boss, E. Beam attenuation and chlorophyll concentration as alternative optical indices of phytoplankton biomass. J. Mar. Res. 2006, 64, 431–451. [Google Scholar] [CrossRef]
- Gervais, F.; Riebesell, U.; Gorbunov, M.Y. Changes in primary productivity and chlorophyll a in response to iron fertilization in the Southern Polar Frontal Zone. Limnol. Oceanogr. 2002, 47, 1324–1335. [Google Scholar] [CrossRef]
- Holeton, C.L.; Nedelec, F.; Sanders, R.; Brown, L.; Moore, C.M.; Stevens, D.P.; Heywood, K.J.; Statham, P.J.; Lucas, C.H. Physiological state of phytoplankton communities in the Southwest Atlantic sector of the Southern Ocean, as measured by fast repetition rate fluorometry. Polar Biol. 2005, 29, 44–52. [Google Scholar] [CrossRef][Green Version]
- Morris, P.J.; Sanders, R. A carbon budget for a naturally iron fertilized bloom in the Southern Ocean. Glob. Biogeochem. Cycles 2011, 25. [Google Scholar] [CrossRef]
- Ryan-Keogh, T.J.; Macey, A.I.; Nielsdóttir, M.C.; Lucas, M.I.; Steigenberger, S.S.; Stinchcombe, M.C.; Achterberg, E.P.; Bibby, T.S.; Moore, C.M. Spatial and temporal development of phytoplankton iron stress in relation to bloom dynamics in the high-latitude North Atlantic Ocean. Limnol. Oceanogr. 2013, 58, 533–545. [Google Scholar] [CrossRef]
- Moore, C.; Mills, M.; Arrigo, K.; Berman-Frank, I.; Bopp, L.; Boyd, P.; Galbraith, E.; Geider, R.; Guieu, C.; Jaccard, S.; et al. Processes and patterns of oceanic nutrient limitation. Nat. Geosci. 2013, 6, 701–710. [Google Scholar]
- Hoppe, C.; Klaas, C.; Ossebaar, S.; Soppa, M.A.; Cheah, W.; Laglera, L.; Santos-Echeandia, J.; Rost, B.; Wolf-Gladrow, D.; Bracher, A.; et al. Controls of primary production in two phytoplankton blooms in the Antarctic Circumpolar Current. Deep Sea Res. Part II Top. Stud. Oceanogr. 2017, 138, 63–73. [Google Scholar] [CrossRef]
- Fall, R.; Copley, S.D. Bacterial sources and sinks of isoprene, a reactive atmospheric hydrocarbon. Environ. Microbiol. 2000, 2, 123–130. [Google Scholar] [CrossRef]
- Wingenter, O.W.; Haase, K.B.; Strutton, P.; Friederich, G.; Meinardi, S.; Blake, D.R.; Rowland, F.S. Changing concentrations of CO, CH4, C5H8, CH3Br, CH3I, and dimethyl sulfide during the Southern Ocean Iron Enrichment Experiments. Proc. Natl. Acad. Sci. USA 2004, 101, 8537–8541. [Google Scholar]
- Moore, R.M. Methyl halide production and loss rates in sea water from field incubation experiments. Mar. Chem. 2006, 101, 213–219. [Google Scholar]
- Bonsang, B.; Gros, V.; Peeken, I.; Yassaa, N.; Bluhm, K.; Zöllner, E.; Sarda-Esteve, R.; Williams, J. Isoprene emission from phytoplankton monocultures: The relationship with chlorophyll-a, cell volume and carbon content. Environ. Chem. 2010, 7, 554–563. [Google Scholar] [CrossRef]
- Meskhidze, N.; Sabolis, A.; Reed, R.; Kamykowski, D. Quantifying environmental stress-induced emissions of algal isoprene and monoterpenes using laboratory measurements. Biogeosciences 2015, 12, 637–651. [Google Scholar] [CrossRef]
- Broadgate, W.J.; Liss, P.S.; Penkett, S.A. Seasonal emissions of isoprene and other reactive hydrocarbon gases from the ocean. Geophys. Res. Lett. 1997, 24, 2675–2678. [Google Scholar] [CrossRef]
- Sharkey, T.D.; Yeh, S. Isoprene emission from plants. Annu. Rev. Plant Biol. 2001, 52, 407–436. [Google Scholar] [CrossRef] [PubMed]
- Gantt, B.; Meskhidze, N.; Kamykowski, D. A new physically-based quantification of isoprene and primary organic aerosol emissions from the world’s oceans. Atmos. Chem. Phys. Discuss 2009, 9, 2933–2965. [Google Scholar] [CrossRef]
- Kurihara, M.; Kimura, M.; Iwamoto, Y.; Narita, Y.; Ooki, A.; Eum, Y.J.; Tsuda, A.; Suzuki, K.; Tani, Y.; Yokouchi, Y.; et al. Distributions of short-lived iodocarbons and biogenic trace gases in the open ocean and atmosphere in the western North Pacific. Mar. Chem. 2010, 118, 156–170. [Google Scholar]
- Kurihara, M.; Iseda, M.; Ioriya, T.; Horimoto, N.; Kanda, J.; Ishimaru, T.; Yamaguchi, Y.; Hashimoto, S. Brominated methane compounds and isoprene in surface seawater of Sagami Bay: Concentrations, fluxes, and relationships with phytoplankton assemblages. Mar. Chem. 2012, 134, 71–79. [Google Scholar] [CrossRef]
- Ardyna, M.; Claustre, H.; Sallée, J.B.; D’Ovidio, F.; Gentili, B.; Van Dijken, G.; D’Ortenzio, F.; Arrigo, K.R. Delineating environmental control of phytoplankton biomass and phenology in the Southern Ocean. Geophys. Res. Lett. 2017, 44, 5016–5024. [Google Scholar] [CrossRef]
- Ardyna, M.; Lacour, L.; Sergi, S.; d’Ovidio, F.; Sallée, J.B.; Rembauville, M.; Blain, S.; Tagliabue, A.; Schlitzer, R.; Jeandel, C.; et al. Hydrothermal vents trigger massive phytoplankton blooms in the Southern Ocean. Nat. Commun. 2019, 10, 2451. [Google Scholar]





| Variable | Abbreviation | Units | Statistics | |||||
|---|---|---|---|---|---|---|---|---|
| Dependent Variable | Isoprene | ISO | pmol | p-Value | Intercept | Slope | n | |
| Independent Variables | Chlorophyll-a (Fluorometric) | CHL-FLUO | g | 0.34 | < 0.001 | 1.0 | 0.57 | 173 |
| Chlorophyll-a (HPLC) | CHL-HPLC | g | 0.48 | <0.001 | 1.4 | 0.56 | 120 | |
| Chlorophytes | CHLO | g Chl-a | 0.14 | <0.001 | 1.4 | 0.15 | 119 | |
| Cryptophytes | CRYP | g Chl-a | 0.17 | <0.001 | 1.4 | 0.14 | 119 | |
| Dinoflagellates | DINO | g Chl-a | 0.23 | <0.001 | 1.7 | 0.3 | 119 | |
| Diatoms | DIAT | g Chl-a | 0.26 | <0.001 | 1.4 | 0.3 | 119 | |
| Haptophytes | HAPT | g Chl-a | 0.17 | <0.001 | 1.5 | 0.3 | 118 | |
| Pelagophyceae | PELA | g Chl-a | 0.17 | <0.001 | 1.7 | 0.29 | 119 | |
| Phaeocystis | PHAEO | g Chl-a | 0.26 | <0.001 | 1.3 | 0.30 | 119 | |
| Prasinophytes | PRA | g Chl-a | 0.1 | <0.001 | 1.5 | 0.21 | 119 | |
| Photoprotective carotenoids | PPC | g | 0.45 | <0.001 | 1.6 | 0.41 | 120 | |
| Light harvesting carotenoids | LHC | g | 0.45 | <0.001 | 1.5 | 0.62 | 120 | |
| PPC:LHC | PPC:LHC | n. d. | 0.16 | <0.001 | 1.4 | 0.47 | 120 | |
| : | : | n. d. | 0.31 | <0.001 | 2.5 | 1.9 | 103 | |
| Prokaryotic heterotrophic abundance | PHA | 0.02 | >0.05 | 169 | ||||
| Particulate organic carbon | POC | mol | 0.25 | <0.001 | 0.02 | 1.07 | 117 | |
| Particulate organic nitrogen | PON | mol | 0.34 | <0.001 | 0.9 | 1.03 | 117 | |
| Nitrate | NITRA | mol | 0.01 | >0.05 | 120 | |||
| Nitrite | NITRI | mol | 0.05 | <0.05 | 0.8 | −0.41 | 120 | |
| Phosphate | PHOSP | mol | 0.001 | >0.05 | 120 | |||
| Silicate | SILIC | mol | 0.03 | <0.001 | 1.2 | −0.13 | 120 | |
| Sea surface temperature | SST | Kelvin | 0.002 | >0.05 | 166 | |||
| Mixed layer depth | MLD | 0.01 | >0.05 | 120 | ||||
| Solar radiation dose | SRD | 0.03 | >0.05 | 117 | ||||
| Predictor var. | SST Regime | Equation | p-Value | RMSE (pM) * | n | |
|---|---|---|---|---|---|---|
| CHL-FLUO | >3.4 C | ISO = 3.5 + 12.6 × CHL-FLUO | 0.67 | <0.001 | 14.1 | 106 |
| CHL-FLUO | <3.4 C | ISO = 4.9 + 1.33 × CHL-FLUO | 0.45 | <0.001 | 3.2 | 115 |
| CHL-HPLC | >3.4 C | ISO = 8.5 + 23.12 × CHL-HPLC | 0.63 | <0.001 | 22.6 | 97 |
| CHL-HPLC | <3.4 C | ISO = 4.9 + 4.45 × CHL-HPLC | 0.43 | <0.001 | 4.4 | 79 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodríguez-Ros, P.; Cortés, P.; Robinson, C.M.; Nunes, S.; Hassler, C.; Royer, S.-J.; Estrada, M.; Sala, M.M.; Simó, R. Distribution and Drivers of Marine Isoprene Concentration across the Southern Ocean. Atmosphere 2020, 11, 556. https://doi.org/10.3390/atmos11060556
Rodríguez-Ros P, Cortés P, Robinson CM, Nunes S, Hassler C, Royer S-J, Estrada M, Sala MM, Simó R. Distribution and Drivers of Marine Isoprene Concentration across the Southern Ocean. Atmosphere. 2020; 11(6):556. https://doi.org/10.3390/atmos11060556
Chicago/Turabian StyleRodríguez-Ros, Pablo, Pau Cortés, Charlotte Mary Robinson, Sdena Nunes, Christel Hassler, Sarah-Jeanne Royer, Marta Estrada, M. Montserrat Sala, and Rafel Simó. 2020. "Distribution and Drivers of Marine Isoprene Concentration across the Southern Ocean" Atmosphere 11, no. 6: 556. https://doi.org/10.3390/atmos11060556
APA StyleRodríguez-Ros, P., Cortés, P., Robinson, C. M., Nunes, S., Hassler, C., Royer, S.-J., Estrada, M., Sala, M. M., & Simó, R. (2020). Distribution and Drivers of Marine Isoprene Concentration across the Southern Ocean. Atmosphere, 11(6), 556. https://doi.org/10.3390/atmos11060556