RNA Pseudouridylation in Physiology and Medicine: For Better and for Worse




Abstract
1. Introduction
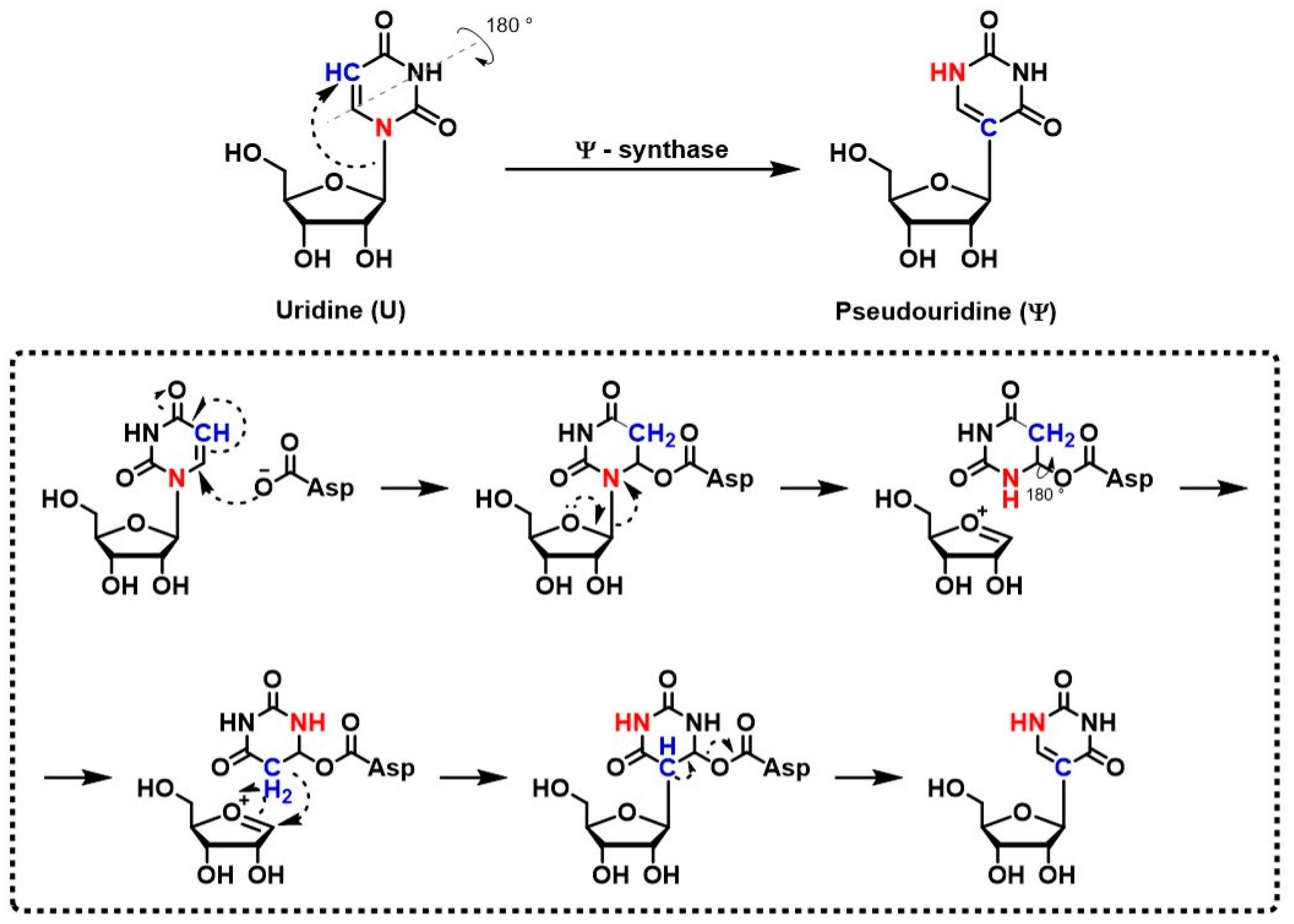
2. Biogenesis and Molecular Impact of Pseudouridine
3. Pseudouridine in Physiology
4. Pseudouridine in Disease
5. Perspectives
Acknowledgments
Conflicts of Interest
References
- Machnicka, M.A.; Milanowska, K.; Osman Oglou, O.; Purta, E.; Kurkowska, M.; Olchowik, A.; Januszewski, W.; Kalinowski, S.; Dunin-Horkawicz, S.; Rother, K.M.; et al. MODOMICS: A database of RNA modification pathways—2013 update. Nucleic Acids Res. 2013, 41, D262–D267. [Google Scholar] [CrossRef] [PubMed]
- Cohn, W.E.; Volkin, E. Nucleoside-5′-Phosphates from Ribonucleic Acid. Nature 1951, 167, 483–484. [Google Scholar] [CrossRef]
- Davis, F.F.; Allen, F.W. Ribonucleic acids from yeast which contain a fifth nucleotide. J. Biol. Chem. 1957, 227, 907–915. [Google Scholar] [PubMed]
- Reddy, R.; Ro-Choi, T.S.; Henning, D.; Shibata, H.; Choi, Y.C.; Busch, H. Modified nucleosides of nuclear and nucleolar low molecular weight ribonucleic acid. J. Biol. Chem. 1972, 247, 7245–7250. [Google Scholar] [PubMed]
- Patton, J.R. Pseudouridine modification of U5 RNA in ribonucleoprotein particles assembled in vitro. Mol. Cell. Biol. 1991, 11, 5998–6006. [Google Scholar] [CrossRef] [PubMed]
- Hall, K.B.; McLaughlin, L.W. Properties of a U1/mRNA 5′ splice site duplex containing pseudouridine as measured by thermodynamic and NMR methods. Biochemistry 1991, 30, 1795–1801. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.-J.; Li, J.-H.; Liu, S.; Wu, J.; Zhou, H.; Qu, L.-H.; Yang, J.-H. RMBase: A resource for decoding the landscape of RNA modifications from high-throughput sequencing data. Nucleic Acids Res. 2016, 44, D259–D265. [Google Scholar] [CrossRef] [PubMed]
- Spenkuch, F.; Motorin, Y.; Helm, M. Pseudouridine: Still mysterious, but never a fake (uridine)! RNA Biol. 2014, 11, 1540–1554. [Google Scholar] [CrossRef] [PubMed]
- Rintala-Dempsey, A.C.; Kothe, U. Eukaryotic stand-alone pseudouridine synthases—RNA modifying enzymes and emerging regulators of gene expression? RNA Biol. 2017, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Motorin, Y.; Keith, G.; Simon, C.; Foiret, D.; Simos, G.; Hurt, E.; Grosjean, H. The yeast tRNA:pseudouridine synthase Pus1p displays a multisite substrate specificity. RNA 1998, 4, 856–869. [Google Scholar] [CrossRef] [PubMed]
- Lecointe, F.; Simos, G.; Sauer, A.; Hurt, E.C.; Motorin, Y.; Grosjean, H. Characterization of yeast protein Deg1 as pseudouridine synthase (Pus3) catalyzing the formation of Ψ38 and Ψ39 in tRNA anticodon loop. J. Biol. Chem. 1998, 273, 1316–1323. [Google Scholar] [CrossRef] [PubMed]
- Becker, H.F.; Motorin, Y.; Planta, R.J.; Grosjean, H. The yeast gene YNL292w encodes a pseudouridine synthase (Pus4) catalyzing the formation of Ψ55 in both mitochondrial and cytoplasmic tRNAs. Nucleic Acids Res. 1997, 25, 4493–4499. [Google Scholar] [CrossRef] [PubMed]
- Ansmant, I.; Motorin, Y.; Massenet, S.; Grosjean, H.; Branlant, C. Identification and characterization of the tRNA: Ψ31-synthase (Pus6p) of Saccharomyces cerevisiae. J. Biol. Chem. 2001, 276, 34934–34940. [Google Scholar] [CrossRef] [PubMed]
- Behm-Ansmant, I.; Urban, A.; Ma, X.; Yu, Y.-T.; Motorin, Y.; Branlant, C. The Saccharomyces cerevisiae U2 snRNA:pseudouridine-synthase Pus7p is a novel multisite-multisubstrate RNA: Ψ-synthase also acting on tRNAs. RNA 2003, 9, 1371–1382. [Google Scholar] [CrossRef] [PubMed]
- Behm-Ansmant, I.; Grosjean, H.; Massenet, S.; Motorin, Y.; Branlant, C. Pseudouridylation at position 32 of mitochondrial and cytoplasmic tRNAs requires two distinct enzymes in Saccharomyces cerevisiae. J. Biol. Chem. 2004, 279, 52998–53006. [Google Scholar] [CrossRef] [PubMed]
- Ofengand, J.; Fournier, M.J. The pseudouridine residues of rRNA: Number, location, biosynthesis, and function. In Modification and Editing of RNA; Grosjean, H., Benne, R., Eds.; ASM Press: Washington, DC, USA, 1998; pp. 229–253. [Google Scholar]
- Ge, J.; Yu, Y.-T. RNA pseudouridylation: New insights into an old modification. Trends Biochem. Sci. 2013, 38, 210–218. [Google Scholar] [CrossRef] [PubMed]
- Deryusheva, S.; Gall, J.G. Dual nature of pseudouridylation in U2 snRNA: Pus1p-dependent and Pus1p-independent activities in yeasts and higher eukaryotes. RNA 2017, 23, 1060–1067. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, S.; Bernstein, D.A.; Mumbach, M.R.; Jovanovic, M.; Herbst, R.H.; León-Ricardo, B.X.; Engreitz, J.M.; Guttman, M.; Satija, R.; Lander, E.S.; et al. Transcriptome-wide mapping reveals widespread dynamic-regulated pseudouridylation of ncRNA and mRNA. Cell 2014, 159, 148–162. [Google Scholar] [CrossRef] [PubMed]
- Carlile, T.M.; Rojas-Duran, M.F.; Zinshteyn, B.; Shin, H.; Bartoli, K.M.; Gilbert, W.V. Pseudouridine profiling reveals regulated mRNA pseudouridylation in yeast and human cells. Nature 2014, 515, 143–146. [Google Scholar] [CrossRef] [PubMed]
- Lovejoy, A.F.; Riordan, D.P.; Brown, P.O. Transcriptome-wide mapping of pseudouridines: Pseudouridine synthases modify specific mRNAs in S. cerevisiae. PLoS ONE 2014, 9, e110799. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhu, P.; Ma, S.; Song, J.; Bai, J.; Sun, F.; Yi, C. Chemical pulldown reveals dynamic pseudouridylation of the mammalian transcriptome. Nat. Chem. Biol. 2015, 11, 592–597. [Google Scholar] [CrossRef] [PubMed]
- Charette, M.; Gray, M.W. Pseudouridine in RNA: What, where, how, and why. IUBMB Life 2000, 49, 341–351. [Google Scholar] [CrossRef] [PubMed]
- Hayrapetyan, A.; Seidu-larry, S.; Helm, M. Function of Modified Nucleosides in RNA Stabilization. In Structure Mechanism, Functions, Cellular Interactions and Evolution; Grosjean, H., Ed.; Landes Bioscience: Austin, TX, USA, 2009; pp. 550–563. [Google Scholar]
- Davis, D.R. Stabilization of RNA stacking by pseudouridine. Nucleic Acids Res. 1995, 23, 5020–5026. [Google Scholar] [CrossRef] [PubMed]
- Roovers, M.; Hale, C.; Tricot, C.; Terns, M.; Terns, R.; Grosjean, H.; Droogmans, L. Formation of the conserved pseudouridine at position 55 in archaeal tRNA. Nucleic Acids Res. 2006, 34, 4293–4301. [Google Scholar] [CrossRef] [PubMed]
- Bekaert, M.; Rousset, J.-P. An extended signal involved in eucaryotic-1 frameshifting operates through modification of the E site tRNA. Mol. Cell 2005, 17, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Penzo, M.; Galbiati, A.; Treré, D.; Montanaro, L. The importance of being (slightly) modified: The role of rRNA editing on gene expression control and its connections with cancer. Biochim. Biophys. Acta Rev. Cancer 2016, 1866. [Google Scholar] [CrossRef] [PubMed]
- King, T.H.; Liu, B.; McCully, R.R.; Fournier, M.J. Ribosome structure and activity are altered in cells lacking snoRNPs that form pseudouridines in the peptidyl transferase center. Mol. Cell 2003, 11, 425–435. [Google Scholar] [CrossRef]
- Liang, X.-H.; Liu, Q.; Fournier, M.J. Loss of rRNA modifications in the decoding center of the ribosome impairs translation and strongly delays pre-rRNA processing. RNA 2009, 15, 1716–1728. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Liu, Q.; Fournier, M.J. rRNA modifications in an intersubunit bridge of the ribosome strongly affect both ribosome biogenesis and activity. Mol. Cell 2007, 28, 965–977. [Google Scholar] [CrossRef] [PubMed]
- Piekna-Przybylska, D.; Przybylski, P.; Baudin-Baillieu, A.; Rousset, J.-P.; Fournier, M.J. Ribosome Performance Is Enhanced by a Rich Cluster of Pseudouridines in the A-site Finger Region of the Large Subunit. J. Biol. Chem. 2008, 283, 26026–26036. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, S.; Loh, Y.-H.; McLoughlin, E.M.; Huang, J.; Park, I.-H.; Miller, J.D.; Huo, H.; Okuka, M.; dos Reis, R.M.; Loewer, S.; et al. Telomere elongation in induced pluripotent stem cells from Dyskeratosis Congenita patients. Nature 2010, 464, 292–296. [Google Scholar] [CrossRef] [PubMed]
- Fong, Y.W.; Ho, J.J.; Inouye, C.; Tjian, R. The dyskerin ribonucleoprotein complex as an OCT4/SOX2 coactivator in embryonic stem cells. Elife 2014, 3. [Google Scholar] [CrossRef] [PubMed]
- Jády, B.E.; Darzacq, X.; Tucker, K.E.; Matera, A.G.; Bertrand, E.; Kiss, T. Modification of Sm small nuclear RNAs occurs in the nucleoplasmic Cajal body following import from the cytoplasm. EMBO J. 2003, 22, 1878–1888. [Google Scholar] [CrossRef]
- Donmez, G.; Hartmuth, K.; Lührmann, R. Modified nucleotides at the 5′ end of human U2 snRNA are required for spliceosomal E-complex formation. RNA 2004, 10, 1925–1933. [Google Scholar] [CrossRef] [PubMed]
- Arnez, J.G.; Steitz, T.A. Crystal structure of unmodified tRNA(Gln) complexed with glutaminyl-tRNA synthetase and ATP suggests a possible role for pseudo-uridines in stabilization of RNA structure. Biochemistry 1994, 33, 7560–7567. [Google Scholar] [CrossRef] [PubMed]
- Karijolich, J.; Yu, Y.-T. Spliceosomal snRNA modifications and their function. RNA Biol. 2010, 7, 192–204. [Google Scholar] [CrossRef] [PubMed]
- Basak, A.; Query, C.C. A Pseudouridine Residue in the Spliceosome Core is Part of the Filamentous Growth Program in Yeast. Cell Rep. 2014, 8, 966–973. [Google Scholar] [CrossRef] [PubMed]
- Karijolich, J.; Yi, C.; Yu, Y.-T. Transcriptome-wide dynamics of RNA pseudouridylation. Nat. Rev. Mol. Cell Biol. 2015, 16, 581–585. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Xiao, M.; Yang, C.; Yu, Y.-T. U2 snRNA is inducibly pseudouridylated at novel sites by Pus7p and snR81 RNP. EMBO J. 2011, 30, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.-L.; Opperman, K.K.; Greider, C.W. A critical stem-loop structure in the CR4-CR5 domain of mammalian telomerase RNA. Nucleic Acids Res. 2002, 30, 592–597. [Google Scholar] [CrossRef] [PubMed]
- Karikó, K.; Muramatsu, H.; Welsh, F.A.; Ludwig, J.; Kato, H.; Akira, S.; Weissman, D. Incorporation of Pseudouridine Into mRNA Yields Superior Nonimmunogenic Vector With Increased Translational Capacity and Biological Stability. Mol. Ther. 2008, 16, 1833–1840. [Google Scholar] [CrossRef]
- Hoernes, T.P.; Clementi, N.; Faserl, K.; Glasner, H.; Breuker, K.; Lindner, H.; Hüttenhofer, A.; Erlacher, M.D. Nucleotide modifications within bacterial messenger RNAs regulate their translation and are able to rewire the genetic code. Nucleic Acids Res. 2016, 44, 852–862. [Google Scholar] [CrossRef] [PubMed]
- Karijolich, J.; Yu, Y.-T. Converting nonsense codons into sense codons by targeted pseudouridylation. Nature 2011, 474, 395–398. [Google Scholar] [CrossRef] [PubMed]
- Parisien, M.; Yi, C.; Pan, T. Rationalization and prediction of selective decoding of pseudouridine-modified nonsense and sense codons. RNA 2012, 18, 355–367. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Zhao, X.; Kierzek, R.; Yu, Y.-T. A flexible RNA backbone within the polypyrimidine tract is required for U2AF65 binding and pre-mRNA splicing in vivo. Mol. Cell. Biol. 2010, 30, 4108–4119. [Google Scholar] [CrossRef] [PubMed]
- Fedorov, N.A.; Bogomazov, M.J. Urinary excretion of purine bases and pseudouridine normal human and in cancer patients before and after radiotherapy. Radiobiol. Radiother. 1969, 10, 605–608. [Google Scholar]
- Waalkes, T.P.; Dinsmore, S.R.; Mrochek, J.E. Urinary excretion by cancer patients of the nucleosides N-dimethylguanosine, 1-methylinosine, and pseudouridine. J. Natl. Cancer Inst. 1973, 51, 271–274. [Google Scholar] [CrossRef] [PubMed]
- Gehrke, C.W.; Kuo, K.C.; Waalkes, T.P.; Borek, E. Patterns of urinary excretion of modified nucleosides. Cancer Res. 1979, 39, 1150–1153. [Google Scholar] [PubMed]
- Seidel, A.; Brunner, S.; Seidel, P.; Fritz, G.I.; Herbarth, O. Modified nucleosides: An accurate tumour marker for clinical diagnosis of cancer, early detection and therapy control. Br. J. Cancer 2006, 94, 1726–1733. [Google Scholar] [CrossRef] [PubMed]
- Jansen, L.H. The so-called "Dyskeratosis Congenita" (Cole, Rauschkolb and Toomey) (Pigmentatio parvo-reticularis cum leucoplakia et dystrophia unguium). Dermatologica 1951, 103, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Knight, S.; Vulliamy, T.; Copplestone, A.; Gluckman, E.; Mason, P.; Dokal, I. Dyskeratosis Congenita (DC) Registry: Identification of new features of DC. Br. J. Haematol. 1998, 103, 990–996. [Google Scholar] [CrossRef] [PubMed]
- Dokal, I. Dyskeratosis Congenita in all its forms. Br. J. Haematol. 2000, 110, 768–779. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, J.R.; Wood, E.; Collins, K. A telomerase component is defective in the human disease dyskeratosis congenita. Nature 1999, 402, 551–555. [Google Scholar] [CrossRef] [PubMed]
- Vulliamy, T.; Marrone, A.; Goldman, F.; Dearlove, A.; Bessler, M.; Mason, P.J.; Dokal, I. The RNA component of telomerase is mutated in autosomal dominant dyskeratosis congenita. Nature 2001, 413, 432–435. [Google Scholar] [CrossRef] [PubMed]
- Ruggero, D.; Grisendi, S.; Piazza, F.; Rego, E.; Mari, F.; Rao, P.H.; Cordon-Cardo, C.; Pandolfi, P.P. Dyskeratosis congenita and cancer in mice deficient in ribosomal RNA modification. Science 2003, 299, 259–262. [Google Scholar] [CrossRef] [PubMed]
- Mochizuki, Y.; He, J.; Kulkarni, S.; Bessler, M.; Mason, P.J. Mouse dyskerin mutations affect accumulation of telomerase RNA and small nucleolar RNA, telomerase activity, and ribosomal RNA processing. Proc. Natl. Acad. Sci. USA 2004, 101, 10756–10761. [Google Scholar] [CrossRef] [PubMed]
- Bellodi, C.; McMahon, M.; Contreras, A.; Juliano, D.; Kopmar, N.; Nakamura, T.; Maltby, D.; Burlingame, A.; Savage, S.A.; Shimamura, A.; et al. H/ACA small RNA dysfunctions in disease reveal key roles for noncoding RNA modifications in hematopoietic stem cell differentiation. Cell Rep. 2013, 3, 1493–1502. [Google Scholar] [CrossRef] [PubMed]
- Yoon, A.; Peng, G.; Brandenburger, Y.; Brandenburg, Y.; Zollo, O.; Xu, W.; Rego, E.; Ruggero, D. Impaired control of IRES-mediated translation in X-linked Dyskeratosis Congenita. Science 2006, 312, 902–906. [Google Scholar] [CrossRef] [PubMed]
- Hellen, C.U.; Sarnow, P. Internal ribosome entry sites in eukaryotic mRNA molecules. Genes Dev. 2001, 15, 1593–1612. [Google Scholar] [CrossRef] [PubMed]
- Bellodi, C.; Krasnykh, O.; Haynes, N.; Theodoropoulou, M.; Peng, G.; Montanaro, L.; Ruggero, D. Loss of function of the tumor suppressor DKC1 perturbs p27 translation control and contributes to pituitary tumorigenesis. Cancer Res. 2010, 70, 6026–6035. [Google Scholar] [CrossRef] [PubMed]
- Montanaro, L.; Calienni, M.; Bertoni, S.; Rocchi, L.; Sansone, P.; Storci, G.; Santini, D.; Ceccarelli, C.; Taffurelli, M.; Carnicelli, D.; et al. Novel dyskerin-mediated mechanism of p53 inactivation through defective mRNA translation. Cancer Res. 2010, 70, 4767–4777. [Google Scholar] [CrossRef] [PubMed]
- Bellodi, C.; Kopmar, N.; Ruggero, D. Deregulation of oncogene-induced senescence and p53 translational control in X-linked Dyskeratosis Congenita. EMBO J. 2010, 29, 1865–1876. [Google Scholar] [CrossRef] [PubMed]
- Rocchi, L.; Pacilli, A.; Sethi, R.; Penzo, M.; Schneider, R.J.; Treré, D.; Brigotti, M.; Montanaro, L. Dyskerin depletion increases VEGF mRNA internal ribosome entry site-mediated translation. Nucleic Acids Res. 2013, 41. [Google Scholar] [CrossRef] [PubMed]
- Montanaro, L.; Brigotti, M.; Clohessy, J.; Barbieri, S.; Ceccarelli, C.; Santini, D.; Taffurelli, M.; Calienni, M.; Teruya-Feldstein, J.; Trerè, D.; et al. Dyskerin expression influences the level of ribosomal RNA pseudo-uridylation and telomerase RNA component in human breast cancer. J. Pathol. 2006, 210, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Baudin-Baillieu, A.; Fabret, C.; Liang, X.-H.; Piekna-Przybylska, D.; Fournier, M.J.; Rousset, J.-P. Nucleotide modifications in three functionally important regions of the Saccharomyces cerevisiae ribosome affect translation accuracy. Nucleic Acids Res. 2009, 37, 7665–7677. [Google Scholar] [CrossRef] [PubMed]
- Montanaro, L. Dyskerin and cancer: More than telomerase. The defect in mRNA translation helps in explaining how a proliferative defect leads to cancer. J. Pathol. 2010, 222, 345–349. [Google Scholar] [CrossRef] [PubMed]
- Penzo, M.; Rocchi, L.; Brugiere, S.; Carnicelli, D.; Onofrillo, C.; Coute, Y.; Brigotti, M.; Montanaro, L. Human ribosomes from cells with reduced dyskerin levels are intrinsically altered in translation. FASEB J. 2015, 29, 3472–3482. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Zhang, J.; Huang, C.; Liu, H. Dyskerin overexpression in human hepatocellular carcinoma is associated with advanced clinical stage and poor patient prognosis. PLoS ONE 2012, 7, e43147. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Garcia, I.; Marcos, T.; Muñoz-Barrutia, A.; Serrano, D.; Pio, R.; Montuenga, L.M.; Ortiz-de-Solorzano, C. Multiscale in situ analysis of the role of dyskerin in lung cancer cells. Integr. Biol. 2013, 5, 402–413. [Google Scholar] [CrossRef] [PubMed]
- Sieron, P.; Hader, C.; Hatina, J.; Engers, R.; Wlazlinski, A.; Müller, M.; Schulz, W.A. DKC1 overexpression associated with prostate cancer progression. Br. J. Cancer 2009, 101, 1410–1416. [Google Scholar] [CrossRef] [PubMed]
- Penzo, M.; Ludovini, V.; Treré, D.; Siggillino, A.; Vannucci, J.; Bellezza, G.; Crinò, L.; Montanaro, L. Dyskerin and TERC expression may condition survival in lung cancer patients. Oncotarget 2015, 6, 21755. [Google Scholar] [CrossRef] [PubMed]
- Penzo, M.; Casoli, L.; Ceccarelli, C.; Treré, D.; Ludovini, V.; Crinò, L.; Montanaro, L. DKC1 gene mutations in human sporadic cancer. Histol. Histopathol. 2013, 28, 365–372. [Google Scholar] [PubMed]
- Alawi, F.; Lin, P. Dyskerin is required for tumor cell growth through mechanisms that are independent of its role in telomerase and only partially related to its function in precursor rRNA processing. Mol. Carcinog. 2011, 50, 334–345. [Google Scholar] [CrossRef] [PubMed]
- Katunaric, M.; Zamolo, G. Modulating telomerase activity in tumor patients by targeting dyskerin binding site for hTR. Med. Hypotheses 2012, 79, 319–320. [Google Scholar] [CrossRef] [PubMed]
- Rocchi, L.; Barbosa, A.J.M.; Onofrillo, C.; Del Rio, A.; Montanaro, L. Inhibition of Human Dyskerin as a New Approach to Target Ribosome Biogenesis. PLoS ONE 2014, 9, e101971. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Karijolich, J.; Glaunsinger, B.; Zhou, Q. Pseudouridylation of 7SK snRNA promotes 7SK snRNP formation to suppress HIV-1 transcription and escape from latency. EMBO Rep. 2016, 17, 1441–1451. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Liu, H.; Zheng, J.; Chen, B.; Zhou, M.; Fan, W.; Wang, H.; Liang, X.; Zhou, X.; Eriani, G.; et al. A Deafness- and Diabetes-associated tRNA Mutation Causes Deficient Pseudouridinylation at Position 55 in tRNAGlu and Mitochondrial Dysfunction. J. Biol. Chem. 2016, 291, 21029–21041. [Google Scholar] [CrossRef] [PubMed]
- Bykhovskaya, Y.; Casas, K.; Mengesha, E.; Inbal, A.; Fischel-Ghodsian, N. Missense mutation in pseudouridine synthase 1 (PUS1) causes Mitochondrial Myopathy and Sideroblastic Anemia (MLASA). Am. J. Hum. Genet. 2004, 74, 1303–1308. [Google Scholar] [CrossRef] [PubMed]
- Patton, J.R.; Bykhovskaya, Y.; Mengesha, E.; Bertolotto, C.; Fischel-Ghodsian, N. Mitochondrial Myopathy and Sideroblastic Anemia (MLASA): Missense mutation in the pseudouridine synthase 1 (PUS1) gene is associated with the loss of tRNA pseudouridylation. J. Biol. Chem. 2005, 280, 19823–19828. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Vizarra, E.; Berardinelli, A.; Valente, L.; Tiranti, V.; Zeviani, M. Nonsense mutation in pseudouridylate synthase 1 (PUS1) in two brothers affected by ;yopathy, Lactic Acidosis and Sideroblastic Anaemia (MLASA). J. Med. Genet. 2007, 44, 173–180. [Google Scholar] [CrossRef]
- Bykhovskaya, Y.; Mengesha, E.; Fischel-Ghodsian, N. Pleiotropic effects and compensation mechanisms determine tissue specificity in Mitochondrial Myopathy and Sideroblastic Anemia (MLASA). Mol. Genet. Metab. 2007, 91, 148–156. [Google Scholar] [CrossRef] [PubMed]
- Shaheen, R.; Han, L.; Faqeih, E.; Ewida, N.; Alobeid, E.; Phizicky, E.M.; Alkuraya, F.S. A homozygous truncating mutation in PUS3 expands the role of tRNA modification in normal cognition. Hum. Genet. 2016, 135, 707–713. [Google Scholar] [CrossRef] [PubMed]
- Barrett, J.C.; Hansoul, S.; Nicolae, D.L.; Cho, J.H.; Duerr, R.H.; Rioux, J.D.; Brant, S.R.; Silverberg, M.S.; Taylor, K.D.; Barmada, M.M.; et al. Genome-wide association defines more than 30 distinct susceptibility loci for Crohn’s disease. Nat. Genet. 2008, 40, 955–962. [Google Scholar] [CrossRef] [PubMed]
- Festen, E.A.M.; Goyette, P.; Green, T.; Boucher, G.; Beauchamp, C.; Trynka, G.; Dubois, P.C.; Lagacé, C.; Stokkers, P.C.F.; Hommes, D.W.; et al. A meta-analysis of genome-wide association scans identifies IL18RAP, PTPN2, TAGAP, and PUS10 as shared risk loci for Crohn’s disease and celiac disease. PLoS Genet. 2011, 7, e1001283. [Google Scholar] [CrossRef] [PubMed]
- McGovern, D.P.B.; Gardet, A.; Törkvist, L.; Goyette, P.; Essers, J.; Taylor, K.D.; Neale, B.M.; Ong, R.T.H.; Lagacé, C.; Li, C.; et al. Genome-wide association identifies multiple UlcerativeCcolitis susceptibility loci. Nat. Genet. 2010, 42, 332–337. [Google Scholar] [CrossRef] [PubMed]
- Dubois, P.C.A.; Trynka, G.; Franke, L.; Hunt, K.A.; Romanos, J.; Curtotti, A.; Zhernakova, A.; Heap, G.A.R.; Adány, R.; Aromaa, A.; et al. Multiple common variants for celiac disease influencing immune gene expression. Nat. Genet. 2010, 42, 295–302. [Google Scholar] [CrossRef] [PubMed]
- Bakin, A.; Ofengand, J. Four newly located pseudouridylate residues in Escherichia coli 23S ribosomal RNA are all at the peptidyltransferase center: Analysis by the application of a new sequencing technique. Biochemistry 1993, 32, 9754–9762. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Karijolich, J.; Yu, Y.-T. Detection and quantification of RNA 2′-O-methylation and pseudouridylation. Methods 2016. [Google Scholar] [CrossRef] [PubMed]
- Zaringhalam, M.; Papavasiliou, F.N. Pseudouridylation meets next-generation sequencing. Methods 2016, 107, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Yu, Y.-T. Detection and quantitation of RNA base modifications. RNA 2004, 10, 996–1002. [Google Scholar] [CrossRef] [PubMed]
- Esmans, E.L.; Geboes, P.; Luyten, Y.; Alderweireldt, F.C. Direct liquid introduction LC/MS microbore experiments for the analysis of nucleoside material present in human urine. Biomed. Mass Spectrom. 1985, 12, 241–245. [Google Scholar] [CrossRef] [PubMed]
- Palmisano, F.; Rotunno, T.; Guerrieri, A.; Zambonin, P.G. Simultaneous determination of pseudouridine and creatinine in untreated urine by ion-pair liquid chromatography with diode-array ultraviolet detection. J. Chromatogr. 1989, 493, 35–43. [Google Scholar] [CrossRef]
- Addepalli, B.; Limbach, P.A. Mass Spectrometry-Based Quantification of Pseudouridine in RNA. J. Am. Soc. Mass Spectrom. 2011, 22, 1363–1372. [Google Scholar] [CrossRef] [PubMed]
- Kuo, K.C.; Smith, C.E.; Shi, Z.; Agris, P.F.; Jehrke, C.W. High performance liquidd chromatography of Cap structures and nucleoside composition of mRNA. J. Chromatogr. Libr. 1990, 45, A297–A316. [Google Scholar]
- Itoh, K.; Mizugaki, M.; Ishida, N. Detection of elevated amounts of urinary pseudouridine in cancer patients by use of a monoclonal antibody. Clin. Chim. Acta 1989, 181, 305–315. [Google Scholar] [CrossRef]
- Hoshino, A.; Honda, I.; Ishimori, A.; Itoh, K.; Mizugaki, M.; Nose, M. Molecular and immunological approach to hematological disease: Detection and analysis of intracellular modified nucleosides by flow cytometry. Jpn. J. Clin. Pathol. 1990, 38, 756–764. [Google Scholar]
- Zhou, J.; Yuan, Y.; Xu, D. Determination of uric acid, creatinine and pseudouridine in human urine by high performance capillary zone electrophoresis. Chin. J. Chromatogr. 1998, 16, 176–177. [Google Scholar]
- Carlile, T.M.; Rojas-Duran, M.F.; Gilbert, W.V. Pseudo-Seq: Genome-Wide Detection of Pseudouridine Modifications in RNA. Methods Enzymol. 2015, 560, 219–245. [Google Scholar] [CrossRef] [PubMed]
- Penzo, M.; Carnicelli, D.; Montanaro, L.; Brigotti, M. A reconstituted cell-free assay for the evaluation of the intrinsic activity of purified human ribosomes. Nat. Protoc. 2016, 11, 1309–1325. [Google Scholar] [CrossRef] [PubMed]
- Patteson, K.G.; Rodicio, L.P.; Limbach, P.A. Identification of the mass-silent post-transcriptionally modified nucleoside pseudouridine in RNA by matrix-assisted laser desorption/ionization mass spectrometry. Nucleic Acids Res. 2001, 29, e49. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Query, C.C.; Meier, U.T. Immunopurified small nucleolar ribonucleoprotein particles pseudouridylate rRNA independently of their association with phosphorylated Nopp140. Mol. Cell. Biol. 2002, 22, 8457–8466. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Gene Name | Enzyme | Catalytic Domain Family | Localization/Predicted Localization | Substrate |
|---|---|---|---|---|
| PUS1 (MLASA1) (y/h) | Pseudouridine synthase 1 tRNA pseudouridine synthase A | TruA | Nucleus | tRNA (c), snRNA, mRNA |
| PUS2 (y) | Pseudouridine synthase 2 | TruA | Mitochondria | tRNA (m) |
| TRUB2 (h) | TruB pseudouridine synthase 2 | TruB | Mitochondria | tRNA(m) |
| PUS3 (y/h) | Pseudouridine synthase 3 | TruA | Nucleus, Cytoplasm | tRNA (c/m), mRNA |
| PUS4 (TruB1) (y/h) | Pseudouridine synthase 4 | TruB | Nucleus, Mitochondria | mRNA |
| PUS5 (y) | Pseudouridine synthase 5 | RluA | Mitochondria | rRNA (m) |
| RPUSD1 (h) | RNA Pseudouridine Synthase Domain Containing 1 | RluA | ? | ? |
| RPUSD2 (h) | RNA Pseudouridine Synthase Domain Containing 2 | RluA | ? | ? |
| PUS6 (y) | Pseudouridine synthase 6 | RluA | Cytoplasm, Mitochondria | tRNA (m) mRNA |
| RPUSD3 (h) | RNA Pseudouridine Synthase Domain Containing 3 | RluA | Mitochondria | rRNA (m) |
| RPUSD4 (h) | RNA Pseudouridine Synthase Domain Containing 4 | RluA | Mitochondria | rRNA (m) |
| PUS7 (y/h) | Pseudouridine synthase 7 | TruD | Nucleus, Cytoplasm | tRNA(c), mRNA |
| PUS7L (h) | Pseudouridine synthase 7 homolog-like protein | TruD | ? | tRNA(c) |
| PUS8 (RIB2) (y) | Pseudouridine synthase 8 | RluA | Cytoplasm | tRNA(c) |
| PUS9 (y) | Pseudouridine synthase 9 | RluA | Mitochondria, Nucleus, Cytoplasm | tRNA (m), mRNA |
| PUS10 (h) | Pseudouridine synthase 10 | Pus10 | Nucleus, Cytoplasm | tRNA(c), ? |
| DKC1 (h/y) | Dyskerin | TruB | Nucleus, Nuceolus. | rRNA (n), sno/scaRNA, snRNA |
| Application | Method | Resolution | High-Throughput | References | |
|---|---|---|---|---|---|
| Pseudouridylation activity | CMC-based assays | Site-specific | No | [77,89,90,91] | |
| snRNAs | TLC-based | Single-nucleotide | No | [92] | |
| Global Ψ | LC/MS | Potentially site-specific | No | [93,94,95] | |
| Global Ψ, tRNAs | HPLC | Single-nucleotide | No | [66,96] | |
| Identification/Quantification of Ψs | Global Ψ | Immunological, antibodies | N/A | No | [97,98] |
| Global Ψ | High performance capillary zone electrophoresis | N/A | No | [99] | |
| snoRNAs (TERC), mRNAs, rRNAs, | Ψ-seq | Single-nucleotide | Yes | [19] | |
| mRNAs, rRNAs, | Pseudo-seq | Single-nucleotide | Yes | [20,100] | |
| mRNAs, rRNAs | Pseudouridine Site Identification sequencing (PSI-seq) | Single-nucleotide | Yes | [21] | |
| mRNAs, rRNAs | CeU-seq | Single-nucleotide | Yes | [22] | |
| Characterization of the functional effects on translation | Fidelity, IRES/Cap dependent translational efficiency | [101] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Penzo, M.; Guerrieri, A.N.; Zacchini, F.; Treré, D.; Montanaro, L. RNA Pseudouridylation in Physiology and Medicine: For Better and for Worse. Genes 2017, 8, 301. https://doi.org/10.3390/genes8110301
Penzo M, Guerrieri AN, Zacchini F, Treré D, Montanaro L. RNA Pseudouridylation in Physiology and Medicine: For Better and for Worse. Genes. 2017; 8(11):301. https://doi.org/10.3390/genes8110301
Chicago/Turabian StylePenzo, Marianna, Ania N. Guerrieri, Federico Zacchini, Davide Treré, and Lorenzo Montanaro. 2017. "RNA Pseudouridylation in Physiology and Medicine: For Better and for Worse" Genes 8, no. 11: 301. https://doi.org/10.3390/genes8110301
APA StylePenzo, M., Guerrieri, A. N., Zacchini, F., Treré, D., & Montanaro, L. (2017). RNA Pseudouridylation in Physiology and Medicine: For Better and for Worse. Genes, 8(11), 301. https://doi.org/10.3390/genes8110301