
The Identification of a New Gene KRTAP 6-3 in Capra hircus and Its Potential for the Diameter Improvement of Cashmere Fibers

Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Species and Sources
2.2. Collection of Cashmere and Blood Samples
2.3. Collection of Skin Tissues
2.4. Identification of the KRTAP6-3 Gene
2.5. DNA Extraction and Amplification of Target Fragments
2.6. Characterization of Genetic Variation in the Goat KRTAP6-3 Gene
2.7. In Situ Hybridization and RT-qPCR
2.8. Data Collation and Analysis
3. Results
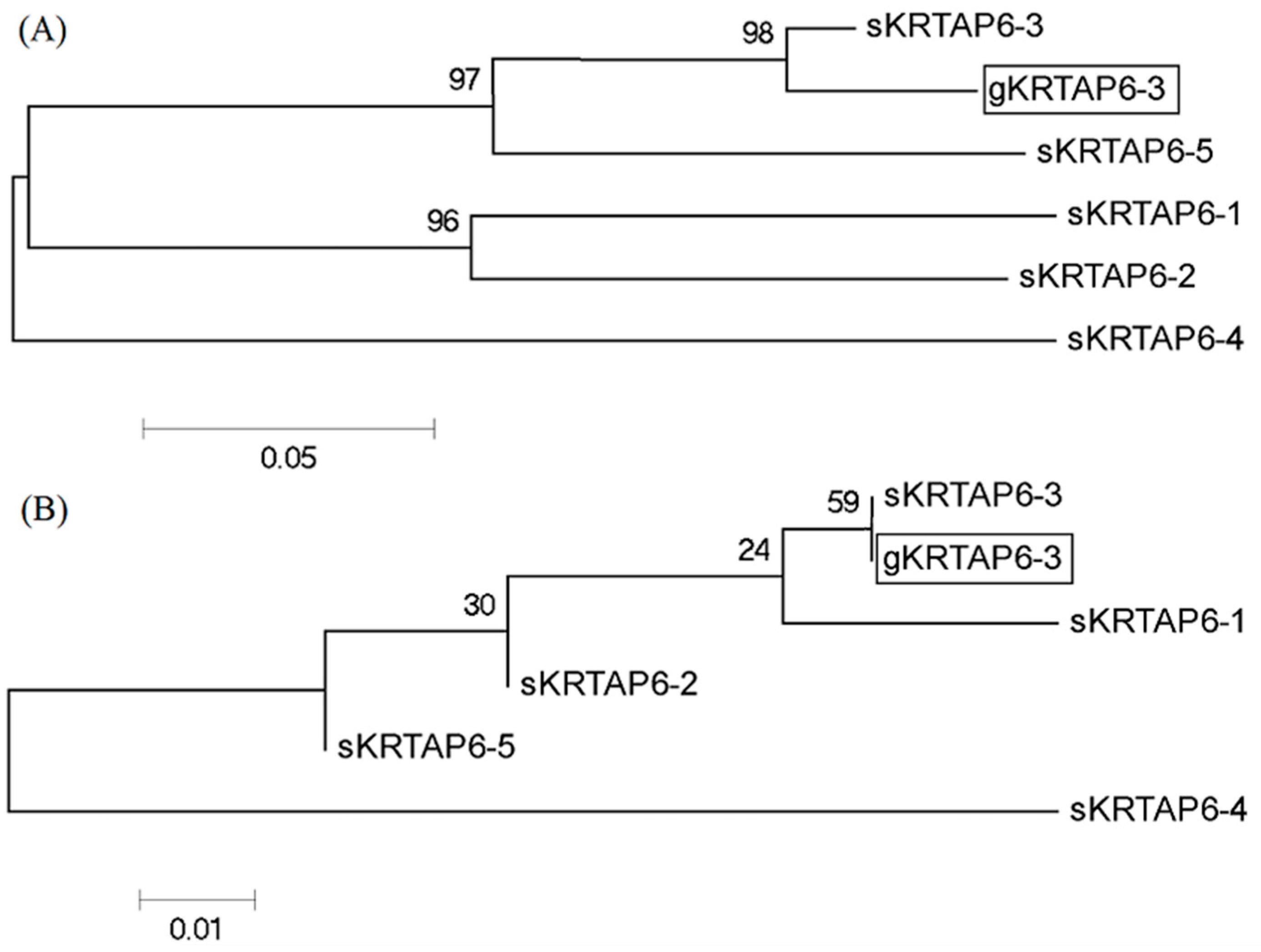
3.1. The Result of KRTAP6-3 Gene Identification
3.2. Genetic Characterization of the KRTAP6-3 Gene
3.3. Localization and Relative Expression Levels of KRTAP6-3 mRNA at Different Periods of Time
3.4. Association Analysis of KRTAP6-3 Gene with Cashmere Traits
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wu, C.; Qin, C.; Fu, X.; Huang, X.; Tian, K. Integrated analysis of lncRNAs and mRNAs by RNA-Seq in secondary hair follicle development and cycling (anagen, catagen and telogen) of Jiangnan cashmere goat (Capra hircus). BMC Vet. Res. 2022, 18, 167. [Google Scholar] [CrossRef] [PubMed]
- Ge, W.; Zhang, W.; Zhang, Y.; Zheng, Y.; Li, F.; Wang, S.; Liu, J.; Tan, S.; Yan, Z.; Wang, L.; et al. A Single-cell Transcriptome Atlas of Cashmere Goat Hair Follicle Morphogenesis. Genom. Proteom. Bioinform. 2021, 19, 437–451. [Google Scholar] [CrossRef]
- Wu, C.; Li, J.; Xu, X.; Xu, Q.; Qin, C.; Liu, G.; Wei, C.; Zhang, G.; Tian, K.; Fu, X. Effect of the FA2H Gene on cashmere fineness of Jiangnan cashmere goats based on transcriptome sequencing. BMC Genom. 2022, 23, 527. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.; Wang, L.; Li, S.; Xing, M.X.; Zhang, X. Characterization and expression analysis of KAP7.1, KAP8.2 gene in Liaoning new-breeding cashmere goat hair follicle. Mol. Biol. Rep. 2011, 38, 3023–3028. [Google Scholar] [CrossRef]
- Gong, H.; Zhou, H.; Bai, L.; Li, W.; Li, S.; Wang, J.; Luo, Y.; Hickford, J.G.H. Associations between variation in the ovine high glycine-tyrosine keratin-associated protein gene KRTAP20-1 and wool traits. J. Anim. Sci. 2019, 97, 587–595. [Google Scholar] [CrossRef]
- Bai, L.; Wang, J.; Zhou, H.; Gong, H.; Tao, J.; Hickford, J.G.H. Identification of Ovine KRTAP28-1 and Its Association with Wool Fibre Diameter. Animals 2019, 9, 142. [Google Scholar] [CrossRef]
- Li, S.; Zhou, H.; Gong, H.; Zhao, F.; Hu, J.; Luo, Y.; Hickford, J.G.H. Identification of the Ovine Keratin-Associated Protein 26-1 Gene and Its Association with Variation in Wool Traits. Genes 2017, 8, 225. [Google Scholar] [CrossRef]
- Liu, B.; Zhao, R.; Wu, T.; Ma, Y.; Gao, Y.; Wu, Y.; Hao, B.; Yin, J.; Li, Y. Transcriptomes reveal microRNAs and mRNAs in different photoperiods influencing cashmere growth in goat. PLoS ONE 2023, 18, e0282772. [Google Scholar] [CrossRef] [PubMed]
- Rile, N.; Liu, Z.; Gao, L.; Qi, J.; Zhao, M.; Xie, Y.; Su, R.; Zhang, Y.; Wang, R.; Li, J.; et al. Expression of Vimentin in hair follicle growth cycle of inner Mongolian Cashmere goats. BMC Genom. 2018, 19, 38. [Google Scholar] [CrossRef]
- Dai, B.; Liang, H.; Guo, D.D.; Bi, Z.W.; Yuan, J.L.; Jin, Y.; Huan, L.; Guo, X.D.; Cang, M.; Liu, D.J. The Overexpression of Tβ4 in the Hair Follicle Tissue of Alpas Cashmere Goats Increases Cashmere Yield and Promotes Hair Follicle Development. Animals 2019, 10, 75. [Google Scholar] [CrossRef]
- Hui, T.; Zheng, Y.; Yue, C.; Wang, Y.; Bai, Z.; Sun, J.; Cai, W.; Zhang, X.; Bai, W.; Wang, Z. Screening of cashmere fineness-related genes and their ceRNA network construction in cashmere goats. Sci. Rep. 2021, 11, 21977. [Google Scholar] [CrossRef] [PubMed]
- Kiktev, D.A.; Sheng, Z.; Lobachev, K.S.; Petes, T.D. GC content elevates mutation and recombination rates in the yeast Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 2018, 115, E7109–E7118. [Google Scholar] [CrossRef]
- Tian, D.; Wang, Q.; Zhang, P.; Araki, H.; Yang, S.; Kreitman, M.; Nagylaki, T.; Hudson, R.; Bergelson, J.; Chen, J.Q. Single-nucleotide mutation rate increases close to insertions/deletions in eukaryotes. Nature 2008, 455, 105–108. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.Q.; Wu, Y.; Yang, H.; Bergelson, J.; Kreitman, M.; Tian, D. Variation in the ratio of nucleotide substitution and indel rates across genomes in mammals and bacteria. Mol. Biol. Evol. 2009, 26, 1523–1531. [Google Scholar] [CrossRef]
- McDonald, M.J.; Wang, W.C.; Huang, H.D.; Leu, J.Y. Clusters of nucleotide substitutions and insertion/deletion mutations are associated with repeat sequences. PLoS Biol. 2011, 9, e1000622. [Google Scholar] [CrossRef]
- Powell, B.C.; Rogers, G.E. The role of keratin proteins and their genes in the growth, structure and properties of hair. Exs 1997, 78, 59–148. [Google Scholar] [CrossRef] [PubMed]
- Rogers, M.A.; Langbein, L.; Praetzel-Wunder, S.; Winter, H.; Schweizer, J. Human hair keratin-associated proteins (KAPs). Int. Rev. Cytol. 2006, 251, 209–263. [Google Scholar] [CrossRef]
- Gong, H.; Zhou, H.; Wang, J.; Li, S.; Luo, Y.; Hickford, J.G.H. Characterisation of an Ovine Keratin Associated Protein (KAP) Gene, Which Would Produce a Protein Rich in Glycine and Tyrosine, but Lacking in Cysteine. Genes 2019, 10, 848. [Google Scholar] [CrossRef]
- McGaughey, G.B.; Gagné, M.; Rappé, A.K. pi-Stacking interactions. Alive and well in proteins. J. Biol. Chem. 1998, 273, 15458–15463. [Google Scholar] [CrossRef]
- Levitt, M.; Perutz, M.F. Aromatic rings act as hydrogen bond acceptors. J. Mol. Biol. 1988, 201, 751–754. [Google Scholar] [CrossRef]
- Fraser, R.D.; Parry, D.A. Filamentous Structure of Hard β-Keratins in the Epidermal Appendages of Birds and Reptiles. Sub-Cell. Biochem. 2017, 82, 231–252. [Google Scholar] [CrossRef]
- Dressler, E.A.; Bormann, J.M.; Weaber, R.L.; Merkel, R.C.; Rolf, M.M. A review of cashmere fiber phenotypes: Production, heritabilities, and genetic correlations. Small Rumin. Res. 2024, 240, 107369. [Google Scholar] [CrossRef]
- Gao, Y.; Duo, L.; Zhe, X.; Hao, L.; Song, W.; Gao, L.; Cai, J.; Liu, D. Developmental Mapping of Hair Follicles in the Embryonic Stages of Cashmere Goats Using Proteomic and Metabolomic Construction. Animals 2023, 13, 3076. [Google Scholar] [CrossRef] [PubMed]
- Toigonbaev, S.; Kerven, C.; Mueller, J.P.; Khaliapov, T.; Abdikarimov, A.; McGregor, B.A. Characterization of the growth, productivity and physical fiber properties of indigenous cashmere goats in southern Kyrgyzstan. Small Rumin. Res. 2023, 226, 107057. [Google Scholar] [CrossRef]
- Zhou, H.; Hickford, J.G.; Fang, Q. A two-step procedure for extracting genomic DNA from dried blood spots on filter paper for polymerase chain reaction amplification. Anal. Biochem. 2006, 354, 159–161. [Google Scholar] [CrossRef]
- Gong, H.; Zhou, H.; Yu, Z.; Dyer, J.; Plowman, J.E.; Hickford, J. Identification of the ovine keratin-associated protein KAP1-2 gene (KRTAP1-2). Exp. Dermatol. 2011, 20, 815–819. [Google Scholar] [CrossRef] [PubMed]
- Byun, S.O.; Fang, Q.; Zhou, H.; Hickford, J.G. An effective method for silver-staining DNA in large numbers of polyacrylamide gels. Anal. Biochem. 2009, 385, 174–175. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Zhou, H.; Gong, H.; Wang, J.; Dyer, J.M.; Luo, Y.; Hickford, J.G. Identification of four new gene members of the KAP6 gene family in sheep. Sci. Rep. 2016, 6, 24074. [Google Scholar] [CrossRef]
- Zhou, H.; Gong, H.; Li, S.; Luo, Y.; Hickford, J.G. A 57-bp deletion in the ovine KAP6-1 gene affects wool fibre diameter. Journal of animal breeding and genetics. J. Anim. Breed. Genet. 2015, 132, 301–307. [Google Scholar] [CrossRef]
- Li, S.; Zhou, H.; Gong, H.; Zhao, F.; Wang, J.; Luo, Y.; Hickford, J.G.H. Variation in the Ovine KAP6-3 Gene (KRTAP6-3) Is Associated with Variation in Mean Fibre Diameter-Associated Wool Traits. Genes 2017, 8, 204. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; He, Z.; Zhao, F.; Hu, J.; Wang, J.; Liu, X.; Zhao, Z.; Li, M.; Luo, Y.; Li, S. Spatiotemporal Expression Characterization of KRTAP6 Family Genes and Its Effect on Wool Traits. Genes 2024, 15, 95. [Google Scholar] [CrossRef] [PubMed]
- Bhat, N.A.; Iqbal, Z.; Mir, S.S.; Bhat, B.A.; Ayaz, A.; Bhat, Z.F.; Shah, R.A.; Ganai, N.A.; Andrabi, S.M.; Bhat, H.F. Characterization and Expression Profiling of KRTAP9.2 at Different Developmental Stages of Pashmina Fiber in Changthangi Goat. Indian. J. Anim. Res. 2022, 56, 1214–1220. [Google Scholar] [CrossRef]
- Wu, D.D.; Irwin, D.M. Evolution of Trichocyte Keratin Associated Proteins. Adv. Exp. Med. Biol. 2018, 1054, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Liao, J.; Lowthert, L.A.; Ku, N.O.; Fernandez, R.; Omary, M.B. Dynamics of human keratin 18 phosphorylation: Polarized distribution of phosphorylated keratins in simple epithelial tissues. J. Cell Biol. 1995, 131, 1291–1301. [Google Scholar] [CrossRef]
- Strnad, P.; Windoffer, R.; Leube, R.E. Induction of rapid and reversible cytokeratin filament network remodeling by inhibition of tyrosine phosphatases. J. Cell Sci. 2002, 115, 4133–4148. [Google Scholar] [CrossRef]
- Gallivan, J.P.; Dougherty, D.A. Cation-pi interactions in structural biology. Proc. Natl. Acad. Sci. USA 1999, 96, 9459–9464. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequences | Length of Products (bp) | Annealing Temperature (°C) |
|---|---|---|---|
| KRTAP6-3 | F: 5′-AAGCTATGGCTATGGAGGC-3′ | 99 | 60 |
| R: 5′-CCAGAGCCGTAGCAGGAG-3′ | |||
| β-actin | F: 5′-ACTACAGCAACTACTACGGTG-3′ | 112 | 60 |
| R: 5′-GGACAGCACCGTGTTGGCGTAGA-3′ |
| Genotype (n) | Genotype Frequency (%) | Genotype (n) | Genotype Frequency (%) |
|---|---|---|---|
| AA (134) | 39.8% | AE (7) | 2.1% |
| AB (82) | 24.3% | CD (4) | 1.2% |
| AC (60) | 17.8% | AF (3) | 0.9% |
| BC (21) | 6.2% | AD (3) | 0.9% |
| BB (9) | 2.7% | DE (3) | 0.9% |
| CC (9) | 2.7% | BE (2) | 0.6% |
| Traits | Variant | Absent | Present | p Value | ||
|---|---|---|---|---|---|---|
| Mean ± SE | n | Mean ± SE | n | |||
| Cashmere yield (g) | A | 415 ± 8.5 | 39 | 413 ± 5.1 | 276 | 0.805 |
| B | 419 ± 4.5 | 203 | 406 ± 8.3 | 112 | 0.252 | |
| C | 412 ± 4.7 | 225 | 428 ± 13.1 | 90 | 0.313 | |
| Mean fiber diameter (μm) | A | 13.5 ± 0.06 | 39 | 13.5 ± 0.05 | 276 | 0.629 |
| B | 13.6 ± 0.04 | 203 | 13.5 ± 0.08 | 112 | 0.219 | |
| C | 13.5 ± 0.04 | 225 | 13.9 ± 0.12 | 90 | 0.003 | |
| Crimped fiber length (μm) | A | 4.3 ± 0.10 | 39 | 4.2 ± 0.05 | 276 | 0.864 |
| B | 4.2 ± 0.05 | 203 | 4.3 ± 0.09 | 112 | 0.556 | |
| C | 4.3 ± 0.05 | 225 | 4.2 ± 0.14 | 90 | 0.528 | |
| Traits | Mean ± SE | p Value | ||
|---|---|---|---|---|
| AA (n = 134) | AB (n = 82) | AC (n = 60) | ||
| Cashmere yield (g) | 416 ± 8.6 | 407 ± 8.7 | 423 ± 15.1 | 0.314 |
| Mean fiber diameter (μm) | 13.4 ± 0.08 b | 13.6 ± 0.08 b | 14.0 ± 0.14 a | 0.010 |
| Crimped fiber length (μm) | 4.2 ± 0.09 | 4.3 ± 0.09 | 4.2 ± 0.16 | 1.000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, J.; Chen, Z.; Zhang, J.; Cao, L.; Li, S. The Identification of a New Gene KRTAP 6-3 in Capra hircus and Its Potential for the Diameter Improvement of Cashmere Fibers. Genes 2025, 16, 721. https://doi.org/10.3390/genes16060721
Cao J, Chen Z, Zhang J, Cao L, Li S. The Identification of a New Gene KRTAP 6-3 in Capra hircus and Its Potential for the Diameter Improvement of Cashmere Fibers. Genes. 2025; 16(6):721. https://doi.org/10.3390/genes16060721
Chicago/Turabian StyleCao, Jian, Zhanzhao Chen, Jianmin Zhang, Liang Cao, and Shaobin Li. 2025. "The Identification of a New Gene KRTAP 6-3 in Capra hircus and Its Potential for the Diameter Improvement of Cashmere Fibers" Genes 16, no. 6: 721. https://doi.org/10.3390/genes16060721
APA StyleCao, J., Chen, Z., Zhang, J., Cao, L., & Li, S. (2025). The Identification of a New Gene KRTAP 6-3 in Capra hircus and Its Potential for the Diameter Improvement of Cashmere Fibers. Genes, 16(6), 721. https://doi.org/10.3390/genes16060721