Abstract
Synthetic hexaploid wheat (SHW) is a useful genetic resource that can be used to improve the performance of common wheat by transferring favorable genes from a wide range of tetraploid or diploid donors. From the perspectives of physiology, cultivation, and molecular genetics, the use of SHW has the potential to increase wheat yield. Moreover, genomic variation and recombination were enhanced in newly formed SHW, which could generate more genovariation or new gene combinations compared to ancestral genomes. Accordingly, we presented a breeding strategy for the application of SHW—the ‘large population with limited backcrossing method’—and we pyramided stripe rust resistance and big-spike-related QTLs/genes from SHW into new high-yield cultivars, which represents an important genetic basis of big-spike wheat in southwestern China. For further breeding applications of SHW-derived cultivars, we used the ‘recombinant inbred line-based breeding method’ that combines both phenotypic and genotypic evaluations to pyramid multi-spike and pre-harvest sprouting resistance QTLs/genes from other germplasms to SHW-derived cultivars; consequently, we created record-breaking high-yield wheat in southwestern China. To meet upcoming environmental challenges and continuous global demand for wheat production, SHW with broad genetic resources from wild donor species will play a major role in wheat breeding.
1. Introduction
Even though wheat (Triticum aestivum L.) is the most widely consumed food in the world, its global production still needs to be enhanced to meet the growing demand in the coming decades [1]. To improve the genetic yield potential of wheat, the introduction of alien genomic components with superior alleles and elite genes from wild/related species is considered an effective conventional method that could broaden the narrow genetic basis of modern common wheat, especially the D genome [2]. However, homologous recombination between alien chromatin and wheat chromosomes is often repressed [3,4,5]; alien chromatin also leads to chromosomal aberrations [6], which is disadvantageous for breeding.
To simulate and study the origin process of the common wheat, synthetic hexaploid wheat (SHW) was created by crossing tetraploid T. turgidum with Aegilops tauschii and subsequently doubling chromosomes [7,8,9,10]. Fortunately, SHW makes it easy to transfer both T. turgidum and Ae. tauschii genes into common wheat, and homologous recombination could break the undesirable gene linkages when crossing SHW with common wheat and fix desirable haplotypes. Therefore, the number of released elite commercial wheat cultivars derived from SHW has continually increased [11,12,13,14,15].
2. High Breeding Potential of Synthetic Hexaploid Wheat
It is thought that the use of SHW can increase wheat yield from the perspectives of physiology and cultivation [16,17,18] because SHW is thought to be a source of genetic diversity for important physiological traits such as enhanced photosynthetic rate [19]. Among the most representative and widely planted cultivars released from 1969 to 2012 in southwestern China, the SHW-derived cultivars possessed higher values of physiological traits, including dry matter accumulation at maturity, harvest index, N utilization efficiency, soil and plant analyzer development (SPAD) value, and canopy/net photosynthetic rate, compared to non-SHW-derived cultivars [18]. This was especially true for SPAD value, canopy apparent photosynthesis rates, and harvest index [16,17], which showed an 11.5% yield increase [16]. For phosphorus use efficiency, SHW-derived cultivars were rated as efficient compared to non-SHW-derived cultivars, which were determined to be moderately efficient or inefficient according to the phosphorus efficiency index inferred from principal component analysis and cluster analysis [20]. Moreover, the use of SHW has the potential to improve a range of stress-adaptive traits of modern bread wheat, such as increased water use efficiency under drought conditions [21].
From the perspective of genetic improvement, favorable genes resistant to biotic/abiotic stresses and related to yield and yield components in T. turgidum or Ae. tauschii are normally expressed in a hexaploid genetic background, and many reported QTLs/genes have been mapped to the AB and D genomes of SHW. The genes that were reported to be resistant to rust [22,23,24], powdery mildew (Pm34 [25]; Pm58 [26,27]), pests (H26 resistant to Hessian fly [28]; greenbug resistance genes [29]), aluminum [30], and drought [31] from Ae. tauschii were also transferred to common wheat, and serve as a gene reservoir for modern wheat adaptation [32,33].
For yield-related traits, using the common wheat ‘Flair’ [34], ‘Opata85’ [35], and ‘Chuanmai 32’ [36] as controls, favorable QTL alleles were also detected in the AB and D genomes of SHW lines ‘XX86’, ‘W7984’, and ‘SHW-L1’; the alleles are mostly involved in grain weight, grain number per spike, spike length, and tiller number. Wan et al. [37] detected a major QTL for leaf sheath hairiness (LSH) that was also associated with grain yield in the common wheat chromosome 4DL; the favorable QTL allele was introgressed from Ae. tauschii, which enhanced grain yield by increasing grain weight. This finding indicated that SHWs might carry yield-related QTL alleles superior to those of modern common wheat.
3. Enhanced Genomic Variation and Recombination in Synthetic Hexaploid Wheat
Approximately 9000 years ago, an accidental hybridization between domesticated emmer wheat (T. turgidum conv. turgidum, 2n = 4x = 28) and goat grass (Ae. tauschii spp. strangulate, 2n = 2x = 14) with chromosome doubling naturally resulted in the generation of a free-threshing hexaploid common wheat (T. aestivum, 2n = 6x = 42) (Figure 1). This hybrid accounts for approximately 95% of current global wheat production, with tetraploid durum wheat (T. turgidum ssp. durum) representing the remaining 5% [38]. Allohexaploidization added the Ae. tauschii D genome into tetraploid wheat, and the allohexaploid wheat was more adaptive to changing environments and then spreading more rapidly around the world than the tetraploid wheat.
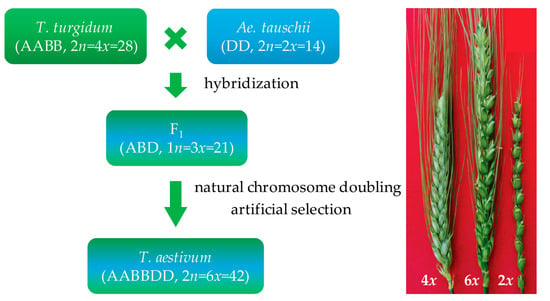
Figure 1.
Hexaploidization and artificial synthesis of wheat.
However, the D genome of the first bread wheat originated from only a small number of wild Ae. tauschii ssp. strangulata plants [2,39], and the AB genome was suggested to be from a free-threshing form of tetraploid wheat [39,40]. As the AB and D genomes originated from limited sources, the individuals from tetraploid and diploid parents involved in the hexaploidization of wheat did not possess all superior characteristics in a few totipotent plants to make them sufficiently adaptable to changing environments worldwide. Therefore, in addition to the advantages of heterosis and gene redundancy [41], there are likely other forces in hexaploidization that accelerate wheat evolution and spread.
Our lab simulated the evolutionary hexaploidization process and generated SHW using different Ae. tauschii and tetraploid wheat with the ability to automatically double chromosomes, which occurs by unreduced gamete formation controlled by genetic factors (QTug.sau-3B [42]). With the 10× resequencing data of SHWs and their parents from next-generation sequencing (NGS) technology, we found that the sequences of the tetraploid AB and diploid D genomes were altered in SHW (Table 1: unpublished data provided by H.W.), and kept changing in subsequent generations of SHW.

Table 1.
Numbers of detected SNP/Indel between different generations of SHW and their parents ¶.
Genomic change by DNA elimination and interchromosomal exchange often occurs in newly formed hexaploid wheat [43,44,45]. Wan et al. [9] found that approximately 10% of the SNP loci of Ae. tauschii were eliminated in derived SHW using diversity array technology (DArT)-Seq technology. Moreover, in the co-dominant genotypes of F2 individuals from a diploid population (SQ665 × SQ783, 2x, D1D1 × D2D2) and a new SHW-derived population (Langdon/SQ665 × Langdon/SQ783, 6x, AABBD1D1 × AABBD2D2), the recombination frequency of Ae. tauschii was found to be enhanced 2.3-fold by hexaploidization with T. turgidum [9].
The changes of ancestral genomes during hexaploidization could generate more genovariation or new alleles, and the gene redundancy that contributed to polyploidization could shield polyploids from the deleterious effect of unfavorable genomic variations. Additionally, the increased genetic recombination in new SHW-derived cultivars could produce more new allelic combinations subject to natural or artificial selection; the evolution of wheat could be accelerated via hexaploidization, and this could help wheat to rapidly spread and increase its role as a major global crop. Therefore, SHW have the potential to enhance variation and adaptive evolution of bread wheat in the breeding process [13].
4. A Case of Successful Direct Application of SHW: Chuanmai 42 from Southwestern China
SHW enhanced genomic variation and recombination, and had high breeding potential. However, primary SHWs also have many unfavorable traits, such as late maturity, taller plants, and difficulty in threshing, which made direct application of SHW difficult. Therefore, our team presented a breeding strategy using a large population with limited backcrossing to common wheat (Figure 2). This breeding strategy involved three core aspects: (1) limited backcrossing with common wheat (2–3 times), reserving more favorable genes and genetic diversity from SHW; (2) using a population of more than 1000 individuals to select favorable gene recombination events between SHW and the backcrossed common wheat; and (3) selecting agronomic traits under multiple environments and testing the candidates’ yield potential using high-yield cultivation methods.
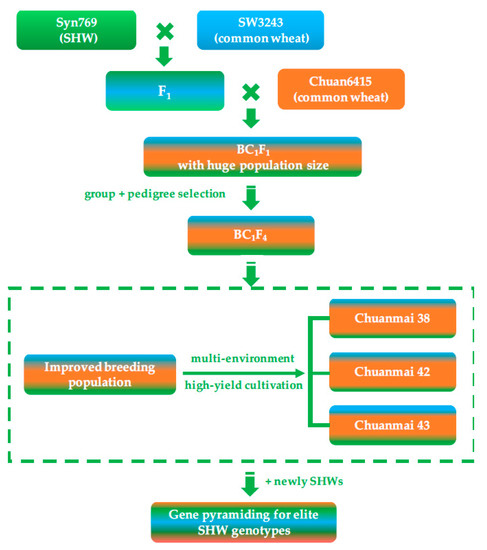
Figure 2.
Direct application of SHW and breeding procedure of Chuanmai 42.
Under this breeding strategy, our team used one SHW line introduced from the International Maize and Wheat Improvement Center (CIMMYT), Syn769, successively crossed with two local common wheat lines, SW3243 and Chuan 6415, and three cultivars were bred: Chuanmai 38, Chuanmai 42, and Chuanmai 43. Among them, Chuanmai 42 was released in 2003 and was the first commercial SHW derivative in the world. In the Sichuan regional trials of wheat cultivars in 2002 and 2003, the average grain yield of Chuanmai 42 increased by 70.2% and 28.3%, respectively, compared to the check cultivars Chuanmai 28 and Chuanmai 107 (Table 2). Overall, grain yield increased by 35%, which broke the yield record of commercial cultivars in southwestern China [46,47].
Table 2.
Yield performance of Chuanmai 42 in Sichuan and national regional cultivar trials.
Since 2003, the use of Chuanmai 42 in wheat production has increased grain output by approximately 1,000,000,000 Kg compared to the old cultivars [47]. Moreover, as a leading wheat cultivar, Chuanmai 42 has become a foundation breeding parent for wheat improvement in southwestern China. From 2008 to 2021, a total of 26 cultivars were selected from crosses containing Chuanmai 42; among them, 17 cultivars were from the first generation of Chuanmai 42 crossed with another parental line with high-yield potential, such as Chuanmai 104 and Chuanmai 602 (Table 3). The application of the SHW-derived Chuanmai 42 received a Second-class Prize of the State Scientific and Technological Progress Award in 2010 and a First-class Prize of the Sichuan Province Scientific and Technological Progress Award in 2009.
Table 3.
Cultivars released from the cross involving Chuanmai 42 in southwestern China.
Since 2003, why is Chuanmai 42 so widely applied in wheat production and breeding in southwestern China? Additionally, what is the role of the SHW germplasm in Chuanmai 42 production?
Unlike the common hexaploid wheat with glabrous leaf sheaths, most SHW accessions have a hairy leaf sheath, which is mostly present in wild species of Triticeae, such as Ae. tauschii and T. turgidum var. dicoccoides. Chuanmai 42 has a hairy leaf sheath that was derived from the SHW line Syn769 (Figure 3). Genetic analysis showed that the LSH is from Ae. tauschii with its controlling gene on chromosome 4DL [37]. Interestingly, almost all cultivars derived from Chuanmai 42 inherited the hairy leaf sheath character, even in the 2nd generation, which was caused by the tight linkage between the LSH gene and the QTLs associated with grain weight and yield in Syn769 (Figure 3E; [37]). In breeding programs, the leaf sheath hairiness can be used as a morphological marker for high grain yield QTL selection from Chuanmai 42 derivatives.

Figure 3.
Hairy leaf sheaths from SHW and its association with grain yield. Leaf sheaths of (A) Chuanmai 42, (B) Syn769, (C) common wheat, and (D) Ae. tauschii; (E) QTL mapping for LSH, and grain yield (GY) and weight (GW).
With a total of 1029 simple sequence repeat (SSR) and 2268 DArT markers detecting polymorphisms among three parents (Syn769, SW3243, and Chuan6415) of Chuanmai 42, the frequency of SHW alleles introgressed to Chuanmai 42 was 15.14%, which was significantly less than the expected 25% assuming random gene assortment. The distribution of introgressed alleles over the A, B, and D genomes was not uniform (B > A > D); introgression occurred most frequently on chromosomes 1A, 1B, 2B, 3A, 4D, 6A, and 6B, whereas none were detected on chromosomes 1D and 7A [13,48,49]. On chromosome 1B, the stripe rust resistance gene YrCH42 from Chuanmai 42 (Figure 4A; [50]) and the QTL allele for increased grain number per spike (Figure 4B; [51]) originated from its SHW parent Syn769 (Figure 4C; [48]).
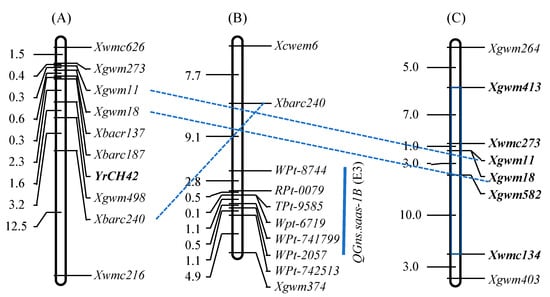
Figure 4.
Molecular mapping of (A) YrCH42, (B) QTL for grain number per spike (GNS), and (C) the SHW alleles introgressed to Chuanmai 42 on chromosome 1B. Note: the interval in gray and the loci in bold italics were from Syn769.
5. Further Application Using SHW-Derived Cultivars: Chuanmai 104
In 2020, the grain yield of Chuanmai 104 reached 729.8 Kg/mu in the high-yield cultivation field in Jiangyou, Sichuan, which was a record for the highest yield of wheat in southwestern China [52,53]. It was selected from the 127 F7 recombinant inbred lines (RILs) of an SHW-derived Chuanmai 42 crossed with Chuannong 16 (Figure 5), which increased the grain yield by approximately 8.42% more than Chuanmai 42 in 2010–2012 national regional cultivar trials.
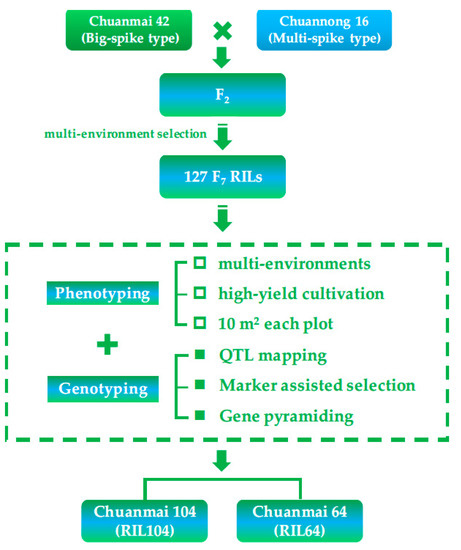
Figure 5.
Application of SHW derivatives and breeding procedure of Chuanmai 104.
We genotyped each F7 RIL using SSR and DArT markers, and evaluated yield-related traits of each RIL (10 m2/plot) in multiple environments with high-yield cultivation. The F2 population of Chuanmai 42 × Chuannong 16 was exposed to different environmental stress to generate more gene recombination [54,55], and 127 RILs were finally obtained. Additionally, we combined phenotypic and genotypic evaluations to artificially select cultivars with high yield potential. Our team referred to this as the ‘RIL-based breeding method’ (Figure 5). Their obvious differences with the traditional phenotypic evaluation for QTL analysis are reflected in the following aspects: (1) replacing single-seed sowing with planting density of high-yield wheat production in China, considering the trade-off between yield components and interplant competition; (2) expanding the planting area of each plot from 2–3 rows to at least 10 m2, simulating field conditions of wheat production, with the aim to bridge the gaps between individual and population performances. In addition, evaluating the contributions of high-yielding QTLs to grain yield on the population level rather than in the individual plant or panicle level was also highly recommended by Xiong et al. [56] recently.
The yield potential of Chuanmai 104 was enhanced by its pre-harvest sprouting resistance and low-temperature tolerance at the flowering stage (Figure 6: unpublished data provided by Y.T.); it is more resistant than its parent, Chuanmai 42 (Figure 6B; [57]). Moreover, Chuanmai 104 had stripe rust and powdery mildew resistance, inheriting the resistance loci of YrCH42 [50], Qyr.saas-7B [58], and QPm.saas-4AS [59] from its parent Chuanmai 42 [51]. The actual grain yield over five continuous years’ field production ranged from 650 kg/mu to 700 kg/mu.
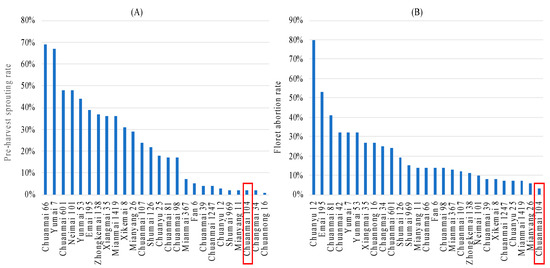
Figure 6.
Pre-harvest sprouting (A) and low temperature (B) tolerance of Chuanmai 104.
Chuanmai 104 was listed as the leading commercial cultivar for wheat production by the Sichuan Province government from 2013 to 2016 and the Ministry of Agriculture and Rural Affairs of China from 2015 to 2016. From 2015 to present, a total of 11 cultivars were selected from crosses involving Chuanmai 104, such as the big-spike cultivar Chuanmai 93 and tight-plant-type cultivar Chuanmai 98; generally, Chuanmai 104 has also been used as a foundation parent for recent wheat breeding (Table 4). Since 2012, the planting acreage of Chuanmai 104 has totaled 21.5 million mu and generated an additional economic output of 2.4 billion Chinese yuan [53]. The breeding and application of Chuanmai 104 obtained the First-class Prize of the Sichuan Province Scientific and Technological Progress Award in 2020.
Table 4.
Cultivars released from the cross involving Chuanmai 42 in southwestern China.
The genetic components of Chuanmai 104 were dissected by QTL mapping for yield-related traits [51,60,61], pre-harvest sprouting resistance [57], and powdery mildew resistance [59]. On chromosome 1B, the majority of genomic regions of Chuanmai 104 were from the female parent Chuanmai 42, and among the detected QTLs or genes, the QTL allele of Chuanmai 42 increased the resistance of stripe rust and grain weight (Figure 7). On chromosome 1DS, the genomic region of Chuanmai 104 associated with increasing spike number per m2 and grain yield was from Chuannong 16. Chromosome 2BS of Chuanmai 104 was from Chuannong 16, and is related to enhanced grain yield and pre-harvest sprouting resistance, whereas its chromosome 2BL was from Chuanmai 42 and was associated with increased thousand-kernel weight and plant height. The chromosome arm 4AS of Chuanmai 104 was from Chuanmai 42, and QTL alleles from Chuanmai 42 increased the grain number per spike and powdery mildew resistance (Figure 7; [59]). For 4DS, the QTL alleles of Chuanmai 42 increased thousand-kernel weight, grain number per spike, and spike number per plant, finally enhancing grain yield (Figure 7; [37,49,51,60]), and Chuanmai 104 inherited the favorable QTL alleles from Chuanmai 42. For the yield-related QTLs on chromosome 5B and 5D, Chuanmai 104 inherited the QTL alleles from Chuanmai 42 with increased thousand-kernel weight and grain yield, respectively (Figure 7; [51]).
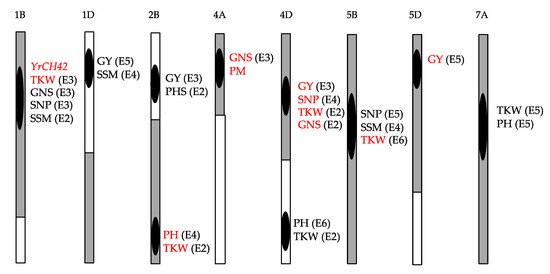
Figure 7.
Genotypic patterns of Chuanmai 104 and yield-related QTLs on seven chromosomes. GY, grain yield; SSM, spike number per square meter; TKW, thousand-kernel weight; GNS, grain number per spike; SNP, spike number per plant; PH, plant height; PHS, pre-harvest sprouting resistance; PM, powdery mildew resistance. Red capital letters indicate that a QTL allele-increased phenotypic value was from the parent Chuanmai 42, and chromosome intervals in gray indicate the genomic regions from Chuanmai 42. The numbers in parentheses represent the number of environments in which each QTL could be detected.
Chuanmai 104 inherited the big-spike properties of higher grain number per spike and grain weight from SHW-derived Chuanmai 42, multiple spike properties including spike number per square meter from Chuannong 16, and grain yield from both parents contributed. Additionally, Chuanmai 42 provided stripe rust [50,58] and powdery mildew resistance [59], whereas Chuannong 16 provided pre-harvest sprouting resistance [57].
6. Conclusions and Perspectives
SHW is a useful genetic resource that can be used to improve the performance of common wheat by transferring favorable genes from a wide range of tetraploid or diploid donors because many QTLs/genes can be expressed under a genetic background of common hexaploid breeding lines [34,35,36,62,63,64,65]. The use of SHW could also bypass the disadvantages of using alien chromatin such as from rye, Agropyron elongatum, and Haynaldia villosa, which undergo reduced homologous recombination with wheat chromosomes. Importantly, genomic variation and recombination were enhanced in newly formed SHW, which could generate more genovariation or new gene combinations compared to ancestral genomes. Additionally, the genomic instability of newly formed SHW can also enhance the genetic variation and recombination when they are crossed with common breeding lines, as well as abundant variation in agronomic traits [66].
How to utilize primary SHW for wheat improvement effectively? Different geneticists and/or breeders summarized various advisable strategies from different perspectives [15,67]. Here, according to our application practices of SHW for decades, using the ‘large population with limited backcrossing method’ was suggested as an effective breeding strategy for direct breeding application of SHW to improve the breeding population and select elite cultivars. These improved SHW derivatives could be used as a basic genetic framework for the next round of pyramiding favorable genes from new SHW lines or other breeding lines. Additionally, for further QTL/gene pyramiding, the RIL-based breeding method could be beneficial by accurately evaluating their breeding value and identifying suitable new cultivars that should be selected.
The global demand for wheat production will keep continuously growing with the world’s increasing population, especially as global warming becomes an increasing worldwide threat that enhances difficulty in crop cultivation. In this situation, a significant increase in yield like during the Green Revolution is desirable to break up the bottleneck of wheat yield and make wheat cultivars tolerant to drought, heat, cold, and flooding stress. SHW along with broad genetic resources from wild donor species will play a big role in the race to meet upcoming environmental challenges and the continuous global demand for wheat production.
Author Contributions
Conceptualization, W.Y.; data curation, J.L., H.W., F.Y., Q.W., Z.L. and Y.T.; visualization, H.W. and Y.T.; writing—original draft preparation, H.W. and F.Y.; supervision, W.Y.; writing—review and editing, W.Y. and J.L. All authors have read and agreed to the published version of the manuscript.
Funding
This study was partially supported by the Sichuan Provincial Finance Department (the Accurate Identification Project of Crop Germplasm; 1+9KJGG001; 2022ZZCX006), the Sichuan Province Science and Technology Department (2023NSFSC1925; 2021YFYZ0020; 2021YFYZ0002).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors extend their appreciation to the support from Key Laboratory of Wheat Biology and Genetic Improvement on Southwestern China (Ministry of agriculture and rural affairs of the PRC).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Food and Agriculture Organization of the United Nations. Statistical Yearbook; FAO: Rome, Italy, 2017. [Google Scholar]
- Wang, J.; Luo, M.C.; Chen, Z.; You, F.M.; Wei, Y.; Zheng, Y.; Dvorak, J. Aegilops tauschii single nucleotide polymorphisms shed light on the origins of wheat D-genome genetic diversity and pinpoint the geographic origin of hexaploid wheat. New Phytol. 2013, 198, 925–937. [Google Scholar] [CrossRef]
- Ji, Y.; Chetelat, R.T. Homoeologous pairing and recombination in Solanum lycopersicoides monosomic addition and substitution lines of tomato. Theor. Appl. Genet. 2003, 106, 979–989. [Google Scholar] [CrossRef]
- Nagy, E.D.; Chu, Y.; Guo, Y.; Khanal, S.; Tang, S.; Li, Y.; Dong, W.B.; Timper, P.; Taylor, C.; Ozias-Akins, P.; et al. Recombination is suppressed in an alien introgression in peanut harboring Rma, a dominant root-knot nematode resistance gene. Mol. Breed. 2010, 26, 357–370. [Google Scholar] [CrossRef]
- Xie, W.; Ben-David, R.; Zeng, B.; Dinoor, A.; Xie, C.; Sun, Q.; Röder, M.S.; Fahoum, A.; Fahima, T. Suppressed recombination rate in 6VS/6AL translocation region carrying the Pm21 locus introgressed from Haynaldia villosa into hexaploid wheat. Mol. Breed. 2012, 29, 399–412. [Google Scholar] [CrossRef]
- Ren, T.; Li, Z.; Yan, B.; Tan, F.; Tang, Z.; Fu, S.; Yang, M.; Ren, Z. Targeted segment transfer from rye chromosome 2R to wheat chromosomes 2A, 2B, and 7B. Cytogenet. Genome Res. 2017, 151, 50–59. [Google Scholar] [CrossRef]
- Mujeeb-Kazi, A.; Rosas, V.; Roldan, S. Conservation of the genetic variation of Triticum tauschii (Coss.) Schmalh. (Aegilops squarrosa auct. non L.) in synthetic hexaploid wheats (T. turgidum L. s.lat. x T. tauschii; 2n = 6x = 42, AABBDD) and its potential utilization for wheat improvement. Genet. Resour. Crop Evol. 1996, 43, 129–134. [Google Scholar] [CrossRef]
- Zhang, L.; Sun, G.; Yan, Z.; Chen, Q.; Yuan, Z.; Lan, X.; Zheng, Y.; Liu, D. Comparison of newly synthetic hexaploid wheat with its donors on SSR products. J. Genet. Genom. 2007, 34, 939–946. [Google Scholar] [CrossRef]
- Wan, H.; Li, J.; Ma, S.; Yang, F.; Chai, L.; Liu, Z.; Wang, Q.; Pu, Z.; Yang, W. Allopolyploidization increases genetic recombination in the ancestral diploid D genome during wheat evolution. Crop J. 2022, 10, 743–753. [Google Scholar] [CrossRef]
- Yang, F.; Wan, H.; Li, J.; Wang, Q.; Yang, N.; Zhu, X.; Liu, Z.; Yang, Y.; Ma, W.; Fan, X.; et al. Pentaploidization enriches the genetic diversity of wheat by enhancing the recombination of AB Genomes. Front. Plant Sci. 2022, 13, 883868. [Google Scholar] [CrossRef]
- Yang, W.; Liu, D.; Li, J.; Zhang, L.; Wei, H.; Hu, X.; Zheng, Y.; He, Z.; Zou, Y. Synthetic hexaploid wheat and its utilization for wheat genetic improvement in China. J. Genet. Genom. 2009, 36, 539–546. [Google Scholar] [CrossRef]
- Mujeeb-Kazi, A.; Kazi, A.G.; Dundas, I.; Rasheed, A.; Ogbonnaya, F.; Kishii, M.; Bonnett, D.; Wang, R.R.C.; Xu, S.; Chen, P.; et al. Genetic diversity for wheat improvement as a conduit to food security. Adv. Agron. 2013, 122, 179–257. [Google Scholar]
- Li, J.; Wan, H.S.; Yang, W.Y. Synthetic hexaploid wheat enhances variation and adaptive evolution of bread wheat in breeding processes. J. Syst. Evol. 2014, 52, 735–742. [Google Scholar] [CrossRef]
- Li, A.; Liu, D.; Yang, W.; Kishii, M.; Mao, L. Synthetic hexaploid wheat: Yesterday, today and tomorrow. Engineering 2018, 4, 552–558. [Google Scholar] [CrossRef]
- Hao, M.; Zhang, L.; Zhao, L.; Dai, S.; Li, A.; Yang, W.; Xie, D.; Li, Q.; Ning, S.; Yan, Z.; et al. A breeding strategy targeting the secondary gene pool of bread wheat: Introgression from a synthetic hexaploid wheat. Theor. Appl. Genet. 2019, 132, 2285–2294. [Google Scholar] [CrossRef]
- Tang, Y.; Rosewarne, G.M.; Li, C.; Wu, X.; Yang, W.; Wu, C. Physiological factors underpinning grain yield improvements of synthetic-derived wheat in southwestern China. Crop Sci. 2015, 55, 98–112. [Google Scholar] [CrossRef]
- Tang, Y.; Wu, X.; Li, C.; Yang, W.; Huang, M.; Ma, X.; Li, S. Yield, growth, canopy traits and photosynthesis in high-yielding, synthetic hexaploid-derived wheats cultivars compared with non-synthetic wheats. Crop Pasture Sci. 2017, 68, 115–125. [Google Scholar] [CrossRef]
- Liu, M.; Tong, H.; Liu, Y.; Li, C.; Wu, X.; Li, M.; Li, X.; Tang, Y. Genetic progress in grain yield and the associated physiological traits of popular wheat in southwestern China from 1969 to 2012. Crop Sci. 2021, 61, 1971–1986. [Google Scholar] [CrossRef]
- del Blanco, I.A.; Rajaram, S.; Kronstad, W.E.; Reynolds, M.P. Physiological performance of synthetic hexaploid wheat-derived populations. Crop Sci. 2000, 40, 1257–1263. [Google Scholar] [CrossRef]
- Yang, H.; Chen, R.; Chen, Y.; Li, H.; Wei, T.; Xie, W.; Fan, G. Agronomic and physiological traits associated with genetic improvement of phosphorus use efficiency of wheat grown in a purple lithomorphic soil. Crop J. 2022, 10, 1151–1164. [Google Scholar] [CrossRef]
- Lopes, M.S.; Reynolds, M.P. Drought adaptive traits and wide adaptation in elite lines derived from resynthesized hexaploid wheat. Crop Sci. 2011, 51, 1617–1626. [Google Scholar] [CrossRef]
- Innes, R.L.; Kerber, E.R. Resistance to wheat leaf rust and stem rust in Triticum tauschii and inheritance in hexaploid wheat of resistance transferred from T. tauschii. Genome 1994, 37, 813–822. [Google Scholar] [CrossRef]
- Yang, W.Y.; Yu, Y.; Zhang, Y.; Hu, X.R.; Wang, Y.; Zhou, Y.C.; Lu, B.R. Inheritance and expression of stripe rust resistance in common wheat (Triticum aestivum) transferred from Aegilops tauschii and its utilization. Hereditas 2003, 139, 49–55. [Google Scholar] [CrossRef]
- Olson, E.L.; Rouse, M.N.; Pumphrey, M.O.; Bowden, R.L.; Gill, B.S.; Poland, J.A. Simultaneous transfer, introgression and genomic localization of genes for resistance to stem rust race TTKSK (Ug99) from Aegilops tauschii to wheat. Theor. Appl. Genet. 2013, 126, 1179–1188. [Google Scholar] [CrossRef]
- Miranda, L.M.; Murphy, J.P.; Marshall, D.; Leath, S. Pm34: A new powdery mildew resistance gene transferred from Aegilops tauschii Coss. to common wheat (Triticum aestivum L.). Theor. Appl. Genet. 2006, 113, 1497–1504. [Google Scholar] [CrossRef]
- Wiersma, A.T.; Pulman, J.A.; Brown, L.K.; Cowger, C.; Olson, E.L. Identification of Pm58 from Aegilops tauschii. Theor. Appl. Genet. 2017, 130, 1123–1133. [Google Scholar] [CrossRef]
- Xue, S.; Hu, S.; Chen, X.; Ma, Y.; Lu, M.; Bai, S.; Wang, X.; Sun, T.; Wang, Y.; Wan, H.; et al. Fine mapping of Pm58 from Aegilops tauschii conferring powdery mildew resistance. Theor. Appl. Genet. 2022, 135, 1657–1669. [Google Scholar] [CrossRef]
- Cox, T.S.; Hatchett, J.H. Hessian fly-resistance gene H26 transferred from Triticum tauschii to common wheat. Crop Sci. 1994, 34, 958–960. [Google Scholar] [CrossRef]
- Zhu, L.; Smith, C.M.; Fritz, A.; Boyko, E.; Voothuluru, P.; Gill, B.S. Inheritance and molecular mapping of new greenbug resistance genes in wheat germplasms derived from Aegilops tauschii. Theor. Appl. Genet. 2005, 111, 831–837. [Google Scholar] [CrossRef]
- Ryan, P.R.; Raman, H.; Gupta, S.; Sasaki, T.; Yamamoto, Y.; Delhaize, E. The multiple origins of aluminium resistance in hexaploid wheat include Aegilops tauschii and more recent cis mutations to TaALMT1. Plant J. 2010, 64, 446–455. [Google Scholar] [CrossRef]
- Sohail, Q.; Inoue, T.; Tanaka, H.; Eltayeb, A.E.; Matsuoka, Y.; Tsujimoto, H. Applicability of Aegilops tauschii drought tolerance traits to breeding of hexaploid wheat. Breed. Sci. 2011, 61, 347–357. [Google Scholar] [CrossRef]
- Jia, J.; Zhao, S.; Kong, X.; Li, Y.; Zhao, G.; He, W.; Appels, R.; Pfeifer, M.; Tao, Y.; Zhang, X.; et al. Aegilops tauschii draft genome sequence reveals a gene repertoire for wheat adaptation. Nature 2013, 496, 91–95. [Google Scholar] [CrossRef] [PubMed]
- Li, A.; Liu, D.; Wu, J.; Zhao, X.; Hao, M.; Geng, S.; Yan, J.; Jiang, X.; Zhang, L.; Wu, J.; et al. mRNA and Small RNA transcriptomes reveal insights into dynamic homoeolog regulation of allopolyploid heterosis in nascent hexaploid wheat. Plant Cell 2014, 26, 1878–1900. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.Q.; Kempf, H.; Ganal, M.W.; Röder, M.S. Advanced backcross QTL analysis in progenies derived from a cross between a German elite winter wheat variety and a synthetic wheat (Triticum aestivum L.). Theor. Appl. Genet. 2004, 109, 933–943. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Kulwal, P.L.; Balyan, H.S.; Gupta, P.K. QTL mapping for yield and yield contributing traits in two mapping populations of bread wheat. Mol. Breed. 2007, 19, 163–177. [Google Scholar] [CrossRef]
- Yu, M.; Chen, G.; Zhang, L.; Liu, Y.; Liu, D.; Wang, J.; Pu, Z.; Zhang, L.; Lan, X.; Wei, Y.; et al. QTL mapping for important agronomic traits in synthetic hexaploid wheat derived from Aegiliops tauschii ssp. tauschii. J. Integr. Agric. 2014, 13, 1835–1844. [Google Scholar] [CrossRef]
- Wan, H.; Yang, Y.; Li, J.; Zhang, Z.; Yang, W. Mapping a major QTL for hairy leaf sheath introgressed from Aegilops tauschii and its association with enhanced grain yield in bread wheat. Euphytica 2015, 205, 275–285. [Google Scholar] [CrossRef]
- Dvorak, J.; Luo, M.C.; Yang, Z.L.; Zhang, H.B. The structure of the Aegilops tauschii genepool and the evolution of hexaploid wheat. Theor. Appl. Genet. 1998, 97, 657–670. [Google Scholar] [CrossRef]
- Dvorak, J.; Deal, K.R.; Luo, M.C.; You, F.M.; von Borstel, K.; Dehghani, H. The origin of spelt and free-threshing hexaploid wheat. J. Hered. 2012, 103, 426–441. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhao, X.; Li, Y.; Xu, J.; Bi, A.; Kang, L.; Xu, D.; Chen, H.; Wang, Y.; Wang, Y.G.; et al. Triticum population sequencing provides insights into wheat adaptation. Nat. Genet. 2020, 52, 1412–1422. [Google Scholar] [CrossRef]
- Comai, L. The advantages and disadvantages of being polyploidy. Nat. Rev. Genet. 2005, 6, 836–846. [Google Scholar] [CrossRef]
- Hao, M.; Luo, J.; Zeng, D.; Zhang, L.; Ning, S.; Yuan, Z.; Yan, Z.; Zhang, H.; Zheng, Y.; Feuillet, C.; et al. QTug.sau-3B is a major quantitative trait locus for wheat hexaploidization. Genes Genomes Genet. 2014, 4, 1943–1953. [Google Scholar] [CrossRef] [PubMed]
- Han, F.; Fedak, G.; Guo, W.; Liu, B. Rapid and repeatable elimination of a parental genome-specific DNA repeat (pGc1R-1a) in newly synthesized wheat allopolyploids. Genetics 2005, 170, 1239–1245. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.A.; Guan, L.L.; Chen, G.Y.; Pu, Z.E.; Hou, D.B. Allopolyploidy-induced rapid genomic changes in newly generated synthetic hexaploid wheat. Biotechnol. Biotechnol. Equip. 2017, 31, 236–242. [Google Scholar] [CrossRef]
- Luo, J.; Zhao, L.; Zheng, J.; Li, Y.; Zhang, L.; Liu, D.; Pu, Z.; Hao, M. Karyotype mosaicism in early generation synthetic hexaploid wheats. Genome 2020, 63, 329–336. [Google Scholar] [CrossRef] [PubMed]
- The Wheat Yield per mu in Jiangyou Reach a New Record in Sichuan Province (In Chinese). Available online: http://www.moa.gov.cn/ztzl/qnszcgdfs/201006/t20100602_1497968.htm (accessed on 2 June 2010).
- Representative Achievements: Exploration of Superior Gene from Synthetic Hexaploid wheat and Breeding and Popularization of Chuanmai 42-Series Cultivars (In Chinese). Available online: http://www.chinawestagr.com/zwyjs/showcontent.asp?id=49833 (accessed on 1 September 2022).
- Liao, J.; Wei, H.T.; Li, J.; Yang, Y.M.; Zeng, Y.C.; Peng, Z.S.; Yang, W.Y. Detection of the introgression loci of synthetic hexaploid wheat in wheat variety Chuanmai 42 by SSR markers. Acta Agron. Sin. 2007, 33, 703–707, (In Chinese with English Abstract). [Google Scholar]
- Li, J.; Wei, H.T.; Hu, X.R.; Li, C.S.; Tang, Y.L.; Liu, D.C.; Yang, W.Y. Identification of a high-yield introgression locus in Chuanmai 42 inherited from synthetic hexaploid wheat. Acta Agron. Sin. 2011, 37, 255–262. [Google Scholar] [CrossRef]
- Li, G.Q.; Li, Z.F.; Yang, W.Y.; Zhang, Y.; He, Z.H.; Xu, S.C.; Singh, R.P.; Qu, Y.Y.; Xia, X.C. Molecular mapping of stripe rust resistance gene YrCH42 in Chinese wheat cultivar Chuanmai42 and its allelism with Yr24 and Yr26. Theor. Appl. Genet. 2006, 112, 1434–1440. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wan, H.S.; Yang, W.Y.; Wang, Q.; Zhu, X.G.; Hu, X.R.; Wei, H.T.; Tang, Y.L.; Li, C.S.; Peng, Z.S.; et al. Dissection of genetic components in the new high-yielding wheat cultivar Chuanmai 104. Sci. Agric. Sin. 2014, 47, 2281–2291, (In Chinese with English abstract). [Google Scholar]
- Chuanmai 104, Bred by Sichuan Academy of Agricultural Sciences, Reached the Grain Yield of 729.8 kg/mu, Breaking the Record of Wheat Yield per mu in Southwestern China (In Chinese). Available online: http://www.chinawestagr.com/homepage/showcontent.asp?id=40323 (accessed on 19 May 2020).
- Decoding Science & Technology of Sichuan II: What Is the New Cultivar Chuanmai 104 with the Maximum Grain Yield 729.8 kg/mu? Let’s Find Out (in Chinese). Available online: https://cbgc.scol.com.cn/news/1005312 (accessed on 17 March 2021).
- De Storme, N.; Geelen, D. The impact of environmental stress on male reproductive development in plants: Biological processes and molecular mechanisms. Plant Cell Environ. 2014, 37, 1–18. [Google Scholar] [CrossRef]
- Phillips, D.; Jenkins, G.; Macaulay, M.; Nibau, C.; Wnetrzak, J.; Fallding, D.; Colas, I.; Oakey, H.; Waugh, R.; Ramsay, L. The effect of temperature on the male and female recombination landscape of barley. New Phytol. 2015, 208, 421–429. [Google Scholar] [CrossRef]
- Xiong, D.; Flexas, J.; Huang, J.; Cui, K.; Wang, F.; Douthe, C.; Lin, M. Why high yield QTLs failed in preventing yield stagnation in rice? Crop Environ. 2022, 1, 103–107. [Google Scholar] [CrossRef]
- Wang, Q.; Liu, Z.H.; Wan, H.S.; Wei, H.T.; Long, H.; Li, T.; Deng, G.B.; Li, J.; Yang, W.Y. Identification and pyramiding of QTLs for traits associated with pre-harvest sprouting resistance in two wheat cultivars Chuanmai 42 and Chuannong 16. Sci. Agric. Sin. 2020, 53, 3421–3431, (In Chinese with English abstract). [Google Scholar]
- Yang, M.; Li, G.; Wan, H.; Li, L.; Li, J.; Yang, W.; Pu, Z.; Yang, Z.; Yang, E. Identification of QTLs for stripe rust resistance in a recombinant inbred line population. Int. J. Mol. Sci. 2019, 20, 3410. [Google Scholar] [CrossRef]
- Liu, Z.; Wang, Q.; Wan, H.; Yang, F.; Wei, H.; Xu, Z.; Ji, H.; Xia, X.; Li, J.; Yang, W. QTL mapping for adult-plant resistance to powdery mildew in Chinese elite common wheat Chuanmai 104. Cereal Res. Commun. 2021, 49, 99–108. [Google Scholar] [CrossRef]
- Tang, Y.L.; Li, J.; Wu, Y.Q.; Wei, H.T.; Li, C.S.; Yang, W.Y.; Chen, F. Identification of QTLs for yield-related traits in the recombinant inbred line population derived from the cross between a synthetic hexaploid wheat derived variety Chuanmai 42 and a Chinese elite variety Chuannong 16. Agric. Sci. Chi. 2011, 10, 1665–1680. [Google Scholar] [CrossRef]
- Lu, B.; Deng, G.; Zhang, H.; Li, J.; Wan, H.; Pan, Z.; Yang, W.; Yu, M.; Long, H. QTL mapping of yield-related traits in the high-yield wheat variety Chuanmai42. Chin. J. Appl. Environ. Biol. 2017, 23, 0183–0192, (In Chinese with English abstract). [Google Scholar]
- Zhu, X.; Wan, H.; Li, J.; Zheng, J.; Tang, Z.; Yang, W. Mixed major-genes plus polygenes inheritance analysis for breeding superiority in synthetic hexaploid wheat. J. Nanjing Agric. Univ. 2018, 41, 625–632, (In Chinese with English abstract). [Google Scholar]
- Yang, Y.; Wan, H.; Yang, F.; Xiao, C.; Li, J.; Ye, M.; Chen, C.; Deng, G.; Wang, Q.; Li, A.; et al. Mapping QTLs for enhancing early biomass derived from Aegilops tauschii in synthetic hexaploid wheat. PloS ONE 2020, 15, e0234882. [Google Scholar] [CrossRef] [PubMed]
- Mokhtari, N.; Majidi, M.M.; Mirlohi, A. Potentials of synthetic hexaploid wheats to improve drought tolerance. Sci. Rep. 2022, 12, 20482. [Google Scholar] [CrossRef] [PubMed]
- Mourad, A.M.I.; Morgounov, A.; Baenziger, P.S.; Esmail, S.M. Genetic variation in common bunt resistance in synthetic hexaploid wheat. Plants 2023, 12, 2. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, S.; Jia, X.; Tian, Z.; Wang, Y.; Wang, C.; Zhang, H.; Liu, X.; Zhao, J.; Deng, P.; et al. Chromosome karyotype and stability of new synthetic hexaploid wheat. Mol. Breeding 2021, 41, 60. [Google Scholar] [CrossRef]
- Gorafi, Y.S.A.; Kim, J.S.; Elbashir, A.A.E.; Tsujimoto, H. A population of wheat multiple synthetic derivatives: An effective platform to explore, harness and utilize genetic diversity of Aegilops tauschii for wheat improvement. Theor. Appl. Genet. 2018, 131, 1615–1626. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).