Abstract
Salt stress is a significant environmental factor affecting plant growth and development, with NaCl stress being one of the most common types of salt stress. The halophyte, Tamarix ramosissima Ledeb (T. ramosissima), is frequently utilized for the afforestation of saline-alkali soils. Indeed, there has been limited research and reports by experts and scholars on the regulatory mechanisms of basic leucine zipper (bZIP) genes in T. ramosissima when treated with exogenous potassium (K+) to alleviate the effects of NaCl stress. This study focused on the bZIP genes in T. ramosissima roots under NaCl stress with additional KCl applied. We identified key candidate genes and metabolic pathways related to bZIP and validated them through quantitative real-time PCR (qRT-PCR). The results revealed that under NaCl stress with additional KCl applied treatments at 0 h, 48 h, and 168 h, based on Pfam protein domain prediction and physicochemical property analysis, we identified 20 related bZIP genes. Notably, four bZIP genes (bZIP_2, bZIP_6, bZIP_16, and bZIP_18) were labeled with the plant hormone signal transduction pathway, showing a predominant up-regulation in expression levels. The results suggest that these genes may mediate multiple physiological pathways under NaCl stress with additional KCl applied at 48 h and 168 h, enhancing signal transduction, reducing the accumulation of ROS, and decreasing oxidative damage, thereby enhancing the tolerance of T. ramosissima to NaCl stress. This study provides gene resources and a theoretical basis for further breeding of salt-tolerant Tamarix species and the involvement of bZIP transcription factors in mitigating NaCl toxicity.
1. Introduction
In the context of the global climate instability and deteriorating ecological environment, an increasing number of vegetation, including crops and trees, are subjected to multiple abiotic stresses such as drought, cold, and salinity [1,2,3]. Among these, salt stress is a primary factor affecting plant growth and development [4]. Under salt stress, plant roots become less vigorous, cell membrane permeability decreases, and antioxidant enzyme activities decline, all of which inhibit plant growth [5]. To cope with these stresses, plants have evolved various regulatory mechanisms at the molecular, cellular, physiological, and biochemical levels. This includes a myriad of transcription factors (TFs) to regulate stress-related gene expression, acting as a critical defense against abiotic stresses [6]. Based on existing research, 64 types of transcription factors have been discovered in plants [7], encompassing various families that participate in stress responses and regulate plant resistance. Examples include basic leucine zipper (bZIP) [8], basic helix-loop-helix (bHLH) [9], NAC [10], MYB [11], and WRKY [12].
bZIP transcription factors are among the most widely distributed and conserved gene families in higher plants and play a crucial role in many biotic and abiotic stresses [13,14]. The quantity of bZIP transcription factors differs among various plant species. In Arabidopsis thaliana [15], there are 78 identified members. In contrast, maize (Zea mays L.) [15], wheat (Triticum aestivum L.) [16], soybean (Glycine max L.) [17], and cotton (Gossypium hirsutum L.) [18] have a higher number of these factors, with 125, 187, 160, and 207 members, respectively. Research has shown that bZIP transcription factors participate in various biological processes, including regulating plant growth and development from seed germination [19], the growth and development of various organs like roots and stems [20,21,22,23], plant aging [23], light signal transduction [24], and responses to biotic [25,26,27] and abiotic stresses [28,29]. Under salt stress, bZIP transcription factors play a pivotal role in modulating the salt stress response through several pathways: First, they can either directly or indirectly activate responsive genes. For instance, in Arabidopsis thaliana, at least one TGACG motif exists within the upstream 1 kb of salt-responsive genes. The AtbZIP17 protein in Arabidopsis has a high sequence homology with the TGA1b protein in tobacco (Nicotiana tabacum), which belongs to the TGA/OBF subfamily within the bZIP transcription factors. Both proteins can specifically bind to the TGACG motif, thereby regulating the expression of downstream genes [30]. Second, these transcription factors regulate genes that play a role in intracellular osmotic adjustment. The OsbZIP71 protein in rice (Oryza sativa) specifically binds to the promoter of the OsNHX1 gene (an antiporter protein in rice). The OsNHX1 protein can transport excessive Na+ and K+ from the plant cytoplasm to the vacuoles. This enhances rice’s resistance to salt stress by adjusting cellular osmolarity [31]. Third, they participate in the removal of reactive oxygen species (ROS) within plants. The accumulation of ROS in plants is a clear indicator of oxidative stress when exposed to salt stress. In the Chlamydomonas reinhardtii, the bZIP transcription factor BLZ8 induces the carbon-concentrating mechanism (CCM) under oxidative stress. This increases the CO2 supply for photosynthesis, subsequently inhibiting ROS accumulation [32]. Moreover, when plants are under salt stress, apart from modulating intracellular K+/Na+ concentrations, the expression of plant bZIP transcription factors can also be influenced by plant hormones such as salicylic acid (SA), ethylene (ET), gibberellin (GA), methyl jasmonate/jasmonic acid (MeJA), and abscisic acid (ABA) [33].
Potassium (K+) is an essential macronutrient that plays significant roles in osmoregulation, membrane potential regulation, sugar cotransport, stress adaptation, and growth in plants [34,35]. K+ is very mobile within plant cells and can be reused repeatedly, meaning its efficiency relies heavily on a series of coordinated processes like absorption, transportation, storage, and redistribution [36]. Plants absorb K+ mainly through K+ channels and transport proteins located on the root cell plasma membranes [37]. The absorption of K+ is closely linked to root growth and structure, influencing cell expansion and root hair growth [38]. After absorption from the environment, K+ is transported and distributed to various tissues and organs via the plant’s vascular system [39]. K+ provides abiotic stress resistance. Under saline conditions, K+ maintains ionic balance and osmotic equilibrium, enhancing plant resilience to abiotic stresses [40,41]. Moreover, increasing K+ intake in plants activates the antioxidant defense system and reduces ROS accumulation, thus helping defend against abiotic stresses and significantly improving plant tolerance to such stresses [42]. Chen et al. reported that in Tamarix species, the uptake of K+ in roots under NaCl stress can elevate the activities of SOD, POD, and CAT, reducing the accumulation of ROS and enhancing the salt tolerance of the plant [43]. Similarly, studies by Song and Su demonstrated that in Alternanthera philoxeroides, the enrichment of K+ uptake under drought stress effectively alleviates the inhibitory effects of drought on the growth of the plant [44].
Tamarix ramosissima Ledeb (T. ramosissima) is one of the most widely distributed Tamarix species in the extreme arid regions of northwestern China, encompassing nearly all habitats occupied by Tamarix species [45]. Tamarix species are typical salt-secreting plants, possessing ‘salt glands’ made up of six secretory cells and two collecting cells [46]. These glands transport salts from within the plant to the surface, excreting them and thus protecting the plant from ion toxicity [47]. Studies by Lu et al. have shown that T. ramosissima thrives under salt concentrations less than 100 mM, but concentrations above 200 mM hinder its normal growth and development. This research aims to understand the response of bZIP transcription factors in T. ramosissima under a salt concentration of 200 mM and the application of KCl at the transcriptomic level. The goal is to identify key candidate genes regulated by bZIP transcription factors that mediate K+’s alleviation of salt stress and their critical metabolic pathways. This will provide a theoretical foundation for salt-resistant plant breeding and the role of bZIP transcription factors in mitigating salt damage.
2. Materials and Methods
2.1. Experimental Materials and Treatment
The experiment selected T. ramosissima cuttings with similar growth (5 months old) and placed them in a 24-hole hydroponic container filled with 1/2-strength Hoagland’s nutrient solution (size: 40 cm × 30 cm × 16 cm). The nutrient solution was replaced every 3 days. The cuttings were placed in a greenhouse with controlled conditions (temperature: 26 ± 2 °C, relative humidity: 40–55%) and grown for 2 months before use. The experiment was conducted in the laboratory of the College of Forestry at Nanjing Forestry University, with tamarisk seed sources coming from the Xinjiang Institute of Ecology and Geography, Chinese Academy of Sciences. Each experiment set up a control group and a treatment group, with 8 plants in each group and 3 replicates, totaling 24 plants. Plants were cultivated in 1/2-strength Hoagland’s nutrient solution as the control group. The treatment groups were cultivated in 1/2-strength Hoagland’s nutrient solution supplemented with 200 mM NaCl and another with 1/2-strength Hoagland’s nutrient solution supplemented with both 200 mM NaCl and 10 mM KCl. The nutrient solution was replaced every 3 days for all groups. The root samples were then collected at 0 h, 48 h, and 168 h after treatment for transcriptome sequencing.
2.2. Transcriptome Sequencing and Differential Gene Expression Screening
We selected root samples from treated T. ramosissima and sent them to a biotechnology company (GENE Denovo, Guangzhou, China) for transcriptome sequencing. RNA was isolated using the Invitrogen kit (Beinuo Bio, Shanghai, China). Following isolation, mRNA was enriched with oligo (dT) cellulose and then segmented to an average length of 200 nt with a fragmentation buffer from New England Biolabs (#E7530). Utilizing random hexamer primers, the first-strand cDNA synthesis occurred, succeeded by the creation of the second-strand cDNA using DNA polymerase I and RNase H. Subsequent steps included the purification of the cDNA fragments, the addition of a terminal “A” base, and their ligation to Illumina adapters. Size selection was conducted on an agarose gel, from which appropriate fragments were isolated and subjected to PCR amplification. The sequenced reads of these fragments were generated on the Illumina HiSeq system by a biotechnology company (GENE Denovo, Guangzhou, China) [48]. The raw data obtained from transcriptome sequencing was submitted to the Sequence Read Archive (SRA) database of the National Center for Biotechnology Information (NCBI) with an SRP number of SRP356215.
The read count data from sequencing were analyzed using DESeq2 software version 1.42.0 [49] to calculate the accurate False Discovery Rate (FDR) values, which are the p-values adjusted by the BH correction. Genes were considered significantly enriched if they had a corrected p-value of less than 0.05. Based on the differential analysis results, genes with an FDR < 0.05 and an absolute log2 fold-change (|log2FC|) greater than 1 (|log2FC| > 1) were identified as significant DEGs. We conducted DEG screening on the obtained data [50] and labeled them for the Gene Ontology (GO) [51] and Kyoto Encyclopedia of Genes and Genomes (KEGG) [52] for enrichment analysis.
2.3. Prediction of Pfam Protein Structure Domains
We aligned the protein sequences of the identified bZIP candidate genes in the Pfam database [53] to obtain labels for their protein domains. The basic physicochemical properties of the members of the bZIP gene family were analyzed and predicted online using Protparam (https://web.expasy.org/protparam/, accessed on 15 September 2023), including the molecular weight, theoretical pI, grand average of hydropathicity (GRAVY), instability index, aliphatic index, etc. Finally, the obtained bZIP proteins were subjected to subcellular localization prediction analysis using the website CELLOv.2.5 (http://cello.life.nctu.edu.tw/, accessed on 15 September 2023).
2.4. Validation of Candidate Genes with Quantitative Real-Time PCR (qRT-PCR)
We chose nine genes at random to validate the precision of our RNA-Seq results. We extracted the total RNA from T. ramosissima root samples using the RNAprep pure kit (Tiangeng, Beijing, China) and then synthesized cDNA with the Prime Script™ II 1st Strand cDNA Synthesis Kit (Takara, Beijing, China). The primers for this process were designed using the Primer Premier 5 software (Supplementary Table S1). For analysis, we utilized the SYBR Green Realtime PCR Master Mix (TOYBO, Jinan, China) and ran the samples on the ABI ViiA™ 7 Real-time PCR system (ABI, Carlsbad, CA, USA). The PCR amplification protocol included an initial denaturation at 95 °C for 30 s, followed by 40 cycles of denaturation at 95 °C for 5 s and annealing at 60 °C for 30 s. The melting curve process involved heating at 95 °C for 5 s, then 60 °C for 1 min, gradually increasing to 95 °C, and finally cooling to 50 °C for 30 s. Each gene underwent three biological replications, with Tubulin serving as the internal reference gene. We determined the relative expression levels using the 2−ΔΔCt method [54].
2.5. Data Processing
We processed the data using Microsoft Excel 2016 (Microsoft, Washington, DC, USA) to compute the mean, standard deviation, and log2 fold change. Phylogenetic trees were constructed using MEGA 11 software (MEGA software, State College, PA, USA). Statistical analyses, including ANOVA with the LSD post hoc test, were performed using SPSS 26.0 software (SPSS, New York, NY, USA). Graphical representations of the data were created with Origin 2019 software (OriginLab Corporation, Northampton, MA, USA).
3. Results
3.1. Transcriptome Data Quality Analysis
The obtained transcriptome data (Supplementary Table S2) show that RawDatas are within the range of 37,641,876–44,483,216. The percentage of CleanData is above 98%, Adapters are in the range of 0.25%–1.00%, and LowQuality is between 0.68% and 1.03%. The number of polyA reads (polyA) is 0, and the ratio of reads with N in high filtration (N) is between 0 and 0.01%. In summary, the quality of the transcriptome data in this study is relatively high and is suitable for subsequent analysis.
3.2. Identification and Physicochemical Property Analysis of bZIP Genes in the Roots of T. ramosissima
Based on the gene label results from the transcriptome data in the NCBI database, we constructed a clustered heat map (Figure 1) for the bZIP-related genes and performed Pfam protein domain prediction and filtering (Supplementary Table S3). Ultimately, we identified bZIP transcription factors in the roots of the T. ramosissima. We used the Protparam online tool to analyze the physicochemical properties of the 20 identified bZIP transcription factors (Table 1). In this study, the clustered heat map clearly displays the distribution of these 20 bZIP-related genes across different samples. The results of the physicochemical property analysis show that the number of amino acids ranged from 75 (bZIP_13) to 749 (bZIP_9), showing a significant variation. The molecular weight ranged between 8828.02 (bZIP_13) and 80,805.18 (bZIP_9) Da, proportional to the number of amino acids. The theoretical pI ranged from 4.48 (bZIP_12) to 9.57 (bZIP_13). Among them, 10 members were basic proteins (pI > 7), and 10 members were acidic proteins (pI < 7). Only one bZIP (bZIP_20) had an instability index below 40, indicating that most of them are unstable proteins. The hydrophobicity results for all 20 bZIP proteins were less than 0, indicating that these bZIP proteins are hydrophilic. Subcellular localization predictions revealed that seven bZIP proteins (bZIP_1, bZIP_2, bZIP_11, bZIP_12, bZIP_13, bZIP_17, and bZIP_20) are located in the Cytoplasmic region, seven bZIP proteins (bZIP_4, bZIP_7, bZIP_8, bZIP_10, bZIP_14, bZIP_15, and bZIP_18) are located in the Extracellular region, three bZIP proteins (bZIP_6, bZIP_9, and bZIP_16) are in the Outer Membrane, and three bZIP proteins (bZIP_2, bZIP_5, and bZIP_19) are in the Periplasmic region.
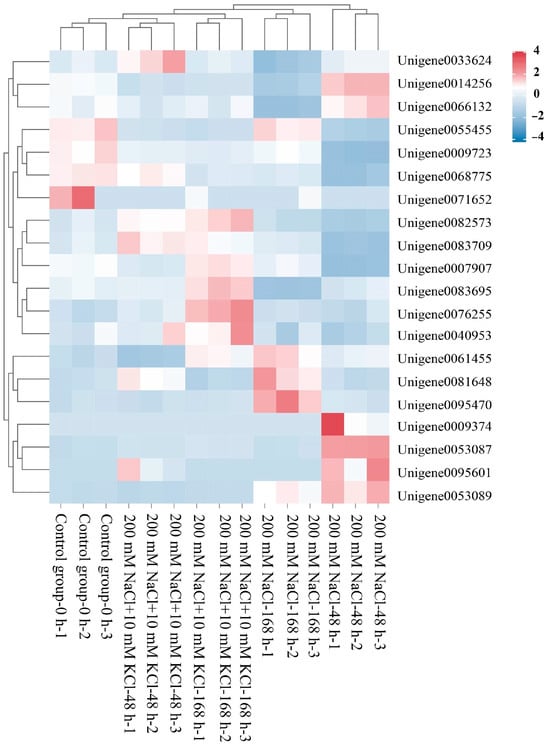
Figure 1.
Clustering heatmap of the bZIP candidate genes (heatmap clustering analysis was performed on the 20 identified bZIP candidate genes across various samples).

Table 1.
Physicochemical properties of bZIP protein in Tamarix ramosissima.
3.3. Analysis of the Expression Levels of bZIP Genes in the Roots of T. ramosissima
Based on the expression level changes of these 20 bZIP genes (Supplementary Figure S1), three bZIP genes (bZIP_4, bZIP_8, and bZIP_11) showed an initial increase and then a decrease under 48 h and 168 h of NaCl stress. However, when additional KCl was added under NaCl stress, their expression levels first decreased and then increased. Two bZIP genes (bZIP_6 and bZIP_16) displayed a pattern of first decreasing and then increasing under 48 h and 168 h of NaCl stress, while their expression levels consistently rose when additional KCl was added under NaCl stress. Notably, the expression level of bZIP_17 consistently declined under 48 h and 168 h of NaCl stress but increased after an initial drop when additional KCl was added. In addition, bZIP_14 showed a constant increase in its expression level under both conditions, with a notably higher level when exogenous K+ was added compared to the control group.
3.4. Analysis of bZIP Genes Enriched in Gene Ontology and the Kyoto Encyclopedia of Genes and Genomes Pathway
Based on the GO enrichment of 20 bZIP genes obtained, the results (Supplementary Table S4) indicate that 5 bZIP genes are not enriched in GO, specifically bZIP_10, bZIP_13, bZIP_15, bZIP_19, and bZIP_20. Nine genes (bZIP_1, bZIP_2, bZIP_3, bZIP_6, bZIP_8, bZIP_11, bZIP_12, bZIP_14, and bZIP_17) are present in the GO Component. Fourteen genes (bZIP_1, bZIP_2, bZIP_3, bZIP_5, bZIP_6, bZIP_7, bZIP_8, bZIP_11, bZIP_12, bZIP_14, bZIP_16, bZIP_17, and bZIP_18) are found in the GO Function. Fifteen genes (bZIP_1, bZIP_2, bZIP_3, bZIP_4, bZIP_5, bZIP_6, bZIP_7, bZIP_8, bZIP_9, bZIP_11, bZIP_12, bZIP_14, bZIP_16, bZIP_17, and bZIP_18) are present in the GO Process.
Four bZIP genes (bZIP_2, bZIP_6, bZIP_16, and bZIP_18) are labeled in the plant hormone signal transduction pathway (Supplementary Table S4). In the N-48 h vs. N + K-48 h comparison group, bZIP_2 is downregulated, whereas bZIP_6, bZIP_16 and bZIP_18 are upregulated. In the same comparison group, only bZIP_6, bZIP_16, and bZIP_18 continue to be upregulated. Additionally, these four bZIP genes (bZIP_2, bZIP_6, bZIP_16, and bZIP_18) belong to the environmental information processing category in KEGG_A_class and to the signal transduction category in KEGG_B_class.
3.5. Analysis of the Plant Hormone Signal Transduction Pathway
Based on the differentially expressed genes labeled for the plant hormone signal transduction pathway when additional KCl was added under NaCl stress in the roots of T. ramosissima (Supplementary Table S5), 74 differentially expressed genes were observed in the 200 mM NaCl-48 h vs. 200 mM NaCl + 10 mM KCl-48 h (N-48 h vs. N + K-48 h) comparison group: 40 genes were upregulated, and 34 genes were downregulated. In the 200 mM NaCl-168 h vs. 200 mM NaCl + 10 mM KCl-168 h (N-168 h vs. N + K-168 h) comparison group, 70 differentially expressed genes were identified, with 40 genes upregulated and 30 genes downregulated. The plant hormone signal transduction pathway is classified under environmental information processing and was significantly enriched in differentially expressed genes in the N-48 h vs. N + K-48 h comparison group.
According to Supplementary Figure S1 and Figure 2, four bZIP genes (bZIP_2, bZIP_6, bZIP_16, and bZIP_18) were labeled for the plant hormone signal transduction pathway, and their expression levels varied. Notably, in the N-48 h vs. N + K-48 h comparison group, bZIP_2 was downregulated, while bZIP_6, bZIP_16, and bZIP_18 were upregulated. In the N-168 h vs. N + K-168 h comparison group, the latter three genes continued to be upregulated. Based on the expression patterns of these four genes, we observed that under 200 mM NaCl for 48 h and 168 h, apart from bZIP_2, which first increased and then decreased, the other three genes’ expression levels first decreased and then increased. Interestingly, two genes (bZIP_6 and bZIP_16) consistently showed an upward trend under 48 h and 168 h of 200 mM NaCl + 10 mM KCl. However, bZIP_2 remained unchanged, while bZIP_18 first increased and then decreased.
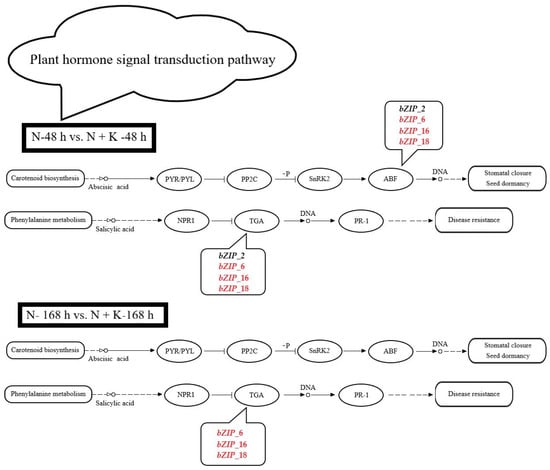
Figure 2.
Label of bZIP-related differentially expressed genes in the plant hormone signal transduction pathway (changes in the bZIP-related DEGs labeled for the plant hormone signal transduction pathway in the roots of T. ramosissima under NaCl stress with the application of additional KCl for 48 h and 168 h. Red indicates differentially expressed genes that are upregulated, and black indicates differentially expressed genes that are downregulated).
3.6. Phylogenetic Analysis of Key Candidate bZIP Genes
Based on the bZIP-related genes obtained from the T. ramosissima under 48 h and 168 h with additional KCl applied during NaCl stress and their labels in the KEGG pathway regarding expression changes, we noticed that at a concentration of 200 mM NaCl for both 48 h and 168 h, the expression levels of two bZIP-related genes (bZIP_6 and bZIP_16) first decreased and then increased. Under 48 h and 168 h of 200 mM NaCl + 10 mM KCl, the expression levels of these two genes continually rose. Therefore, we speculate that these two genes (bZIP_6 and bZIP_16) are key candidate genes for the bZIP transcription factor.
We selected these two key candidate genes (bZIP_6 and bZIP_16) and used their protein amino acid sequences to perform a BLAST search on the NCBI website. We then chose ten homologous species for each gene (Supplementary Table S6) and constructed a phylogenetic tree. The results showed that bZIP_6 is closely related to Tamarix hispida (T. hispida), while bZIP_16 has a close phylogenetic relationship with Fagopyrum tataricum (Figure 3).
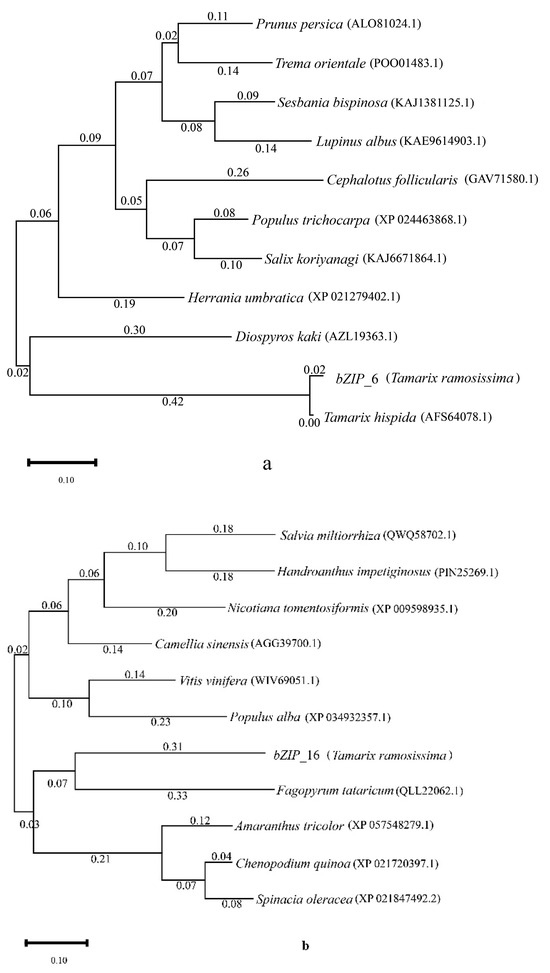
Figure 3.
Phylogenetic tree analysis of T. ramosissima bZIP genes and bZIP genes of other species. (Analysis of the phylogenetic tree constructed from the protein amino acid sequences of bZIP_6 and bZIP_16 from T. ramosissima roots, along with those of 10 other homologous gene species. (a) Evolutionary tree constructed for the bZIP_6 protein. (b) Evolutionary tree constructed for the bZIP_16 protein.)
3.7. qRT-PCR Validation of bZIP Candidate Genes
Nine principal bZIP genes, chosen at random from a group of 20, underwent qRT-PCR analysis. The data from this qRT-PCR were employed to corroborate the precision of the transcriptome sequencing findings in our research. It was observed that the expression patterns of these nine genes, as determined by qRT-PCR, aligned with the trends noted in the transcriptome sequencing analysis (refer to Figure 4). This alignment offers ample proof of the dependability and exactness of the transcriptome sequencing outcomes in this investigation.
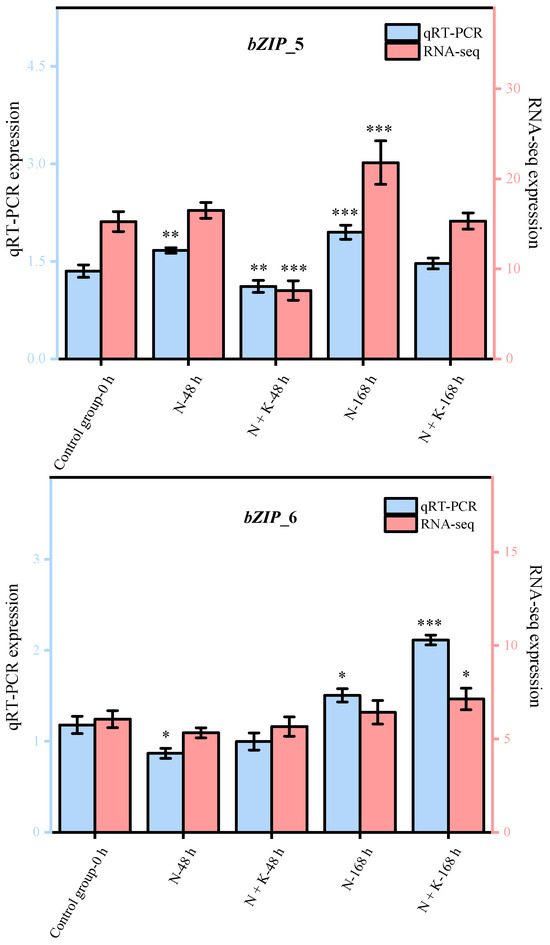
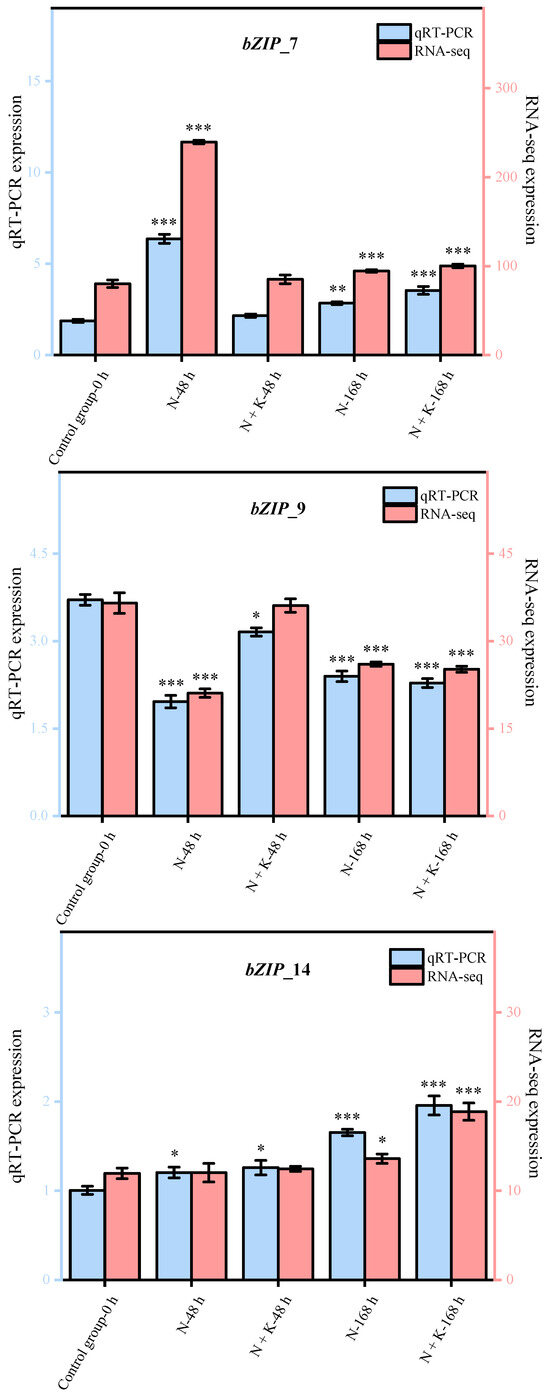
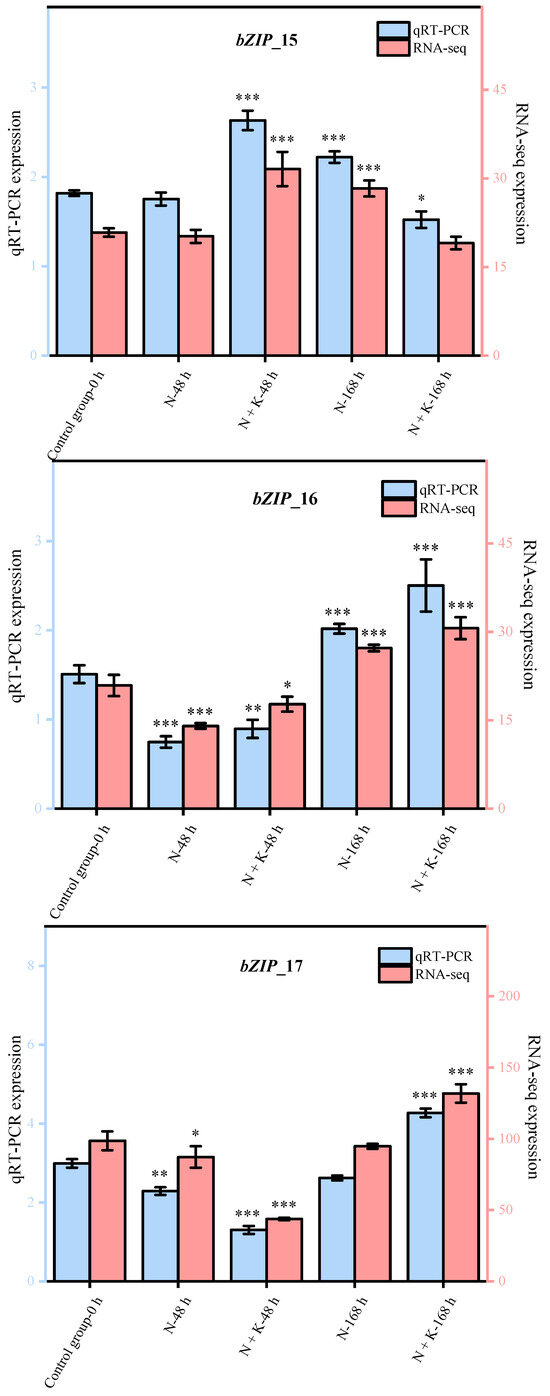
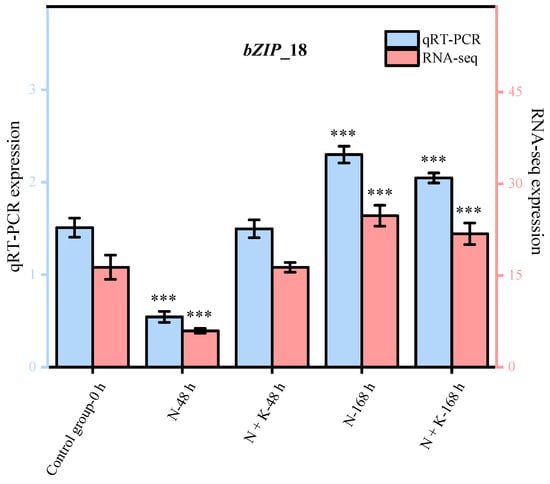
Figure 4.
Quantitative real-time PCR validation of bZIP candidate key genes. (Nine DEGs were randomly selected for qRT-PCR validation, and the error bars were obtained from multiple replicates of qRT-PCR. Note: p ≥ 0.05 is not marked; 0.01 < p < 0.05 is marked as *; 0.001 < p < 0.01 is marked as **; p ≤ 0.001 is marked as ***;  : Numerical value has been shown on the left side of the Y axis;
: Numerical value has been shown on the left side of the Y axis;  : Numerical value has been shown on the right side of the Y axis).
: Numerical value has been shown on the right side of the Y axis).
: Numerical value has been shown on the left side of the Y axis; : Numerical value has been shown on the right side of the Y axis).
4. Discussion
bZIP transcription factors constitute one of the largest and most diverse families in the plant kingdom, with a wide distribution across eukaryotes [55,56]. They are integral to numerous processes in plant biology, including growth, seed development, secondary metabolism, and responses to various stresses [57,58]. Notably, they play a pivotal regulatory role in enhancing plant resistance to abiotic stresses [59,60]. Across different species, bZIP gene family members have been identified in varying numbers, ranging from 62 in the watermelon [61] to 160 in the soybean [17], with counts such as 64 in the yellowhorn [62], 55 in the grapevine [63], and 112 in the apple [64].
However, due to the incomplete genome sequencing of T. ramosissima, there has been limited research on the identification of bZIP gene family members within this plant species. In this study, we conducted transcriptome sequencing to identify 20 members of the bZIP gene family in T. ramosissima in response to NaCl stress with additional KCl applied. Our comprehensive analysis of these 20 bZIP genes contributes to a deeper understanding of the salt-tolerant genes associated with bZIP in T. ramosissima under the influence of additional KCl supplementation during NaCl stress. This knowledge provides a robust scientific basis for the breeding of salt-tolerant crop varieties tailored for saline-alkali areas. The non-biological stress response network mediated by bZIP transcription factors is a crucial mechanism for plants to respond to various abiotic stresses [65]. Under NaCl conditions, ThbZIP1 in T. hispida might regulate the ROS scavenging capacity and stress tolerance-related physiological changes by activating stress-tolerant genes, thus achieving a non-biological stress tolerance and playing a crucial role in ABA-mediated T. hispida stress responses [66]. Additionally, under salt stress, overexpression of ThbZIP1 can enhance the activity of peroxidase (POD) and superoxide dismutase (SOD), and ThbZIP1 enhances ROS scavenging, promotes the accumulation of compatible solutes, induces and strengthens the biosynthesis of soluble proteins, granting plants resistance [67]. In this study, under 200 mM NaCl conditions for 48 h and 168 h, the expression levels of two bZIP-related genes (bZIP_6 and bZIP_16) first decreased and then increased, and the expression level of one bZIP-related gene (bZIP_14) has been rising. Notably, the activities of SOD, POD, and CAT in T. ramosissima under 200 mM NaCl conditions with exogenous K+ for 48 h and 168 h increased, and the antioxidant enzyme system was activated to remove the ROS produced by high salt stress to resist NaCl stress, enhancing plant salt tolerance [43]. Under the influence of additional KCl under the NaCl conditions in this study, the expression levels of bZIP_6, bZIP_16, bZIP_7, and bZIP_14 under exogenous K+ for 48 h and 168 h under NaCl stress were always found to be on the rise. The results suggest that these bZIP-related genes in T. ramosissima may improve plant antioxidant enzyme activity by upregulating expression levels under exogenous K+ in response to NaCl stress and clear ROS accumulation, building an antioxidant defense system and enhancing plant salt tolerance.
ABF transcription factors (ABA-responsive element binding factors) are a type of bZIP protein that specifically recognizes ABRE and belongs to subgroup A of the bZIP family. They mainly participate in the regulation of ABA and stress responses and are also known as ABA-responsive element binding proteins (AREBs) or ABRE binding factors (ABFs) [68,69,70]. ABRE is an ABA response element, and bZIP transcription factors can bind to ABRE elements, known as ABA response element-binding factors, which play a significant role in the ABA-dependent signaling pathway [71]. bZIP is a typical ABRE binding transcription factor, which binds to the ABRE element and trans-activates downstream gene expression [72]. In Tartary buckwheat (Fagopyrum tataricum), the FtbZIP5 gene homologous to ABF was induced by salt stress, and its overexpression in Arabidopsis reduced oxidative damage to transgenic plants under salt stress. Moreover, FtbZIP5, as a positive regulator, mediated plant tolerance to salt and drought through ABA-dependent signaling pathways [73]. In this study, four bZIP-related genes were labeled for the plant hormone signal transduction pathway and activated gene expression. These bZIP genes, induced by ABA, participated in the regulation of NaCl stress. Among them, in the comparison group of N-48 h vs. N + K-48 h, bZIP_2 was downregulated, while bZIP_6, bZIP_16, and bZIP_18 were upregulated. According to the GO classification system, the bZIP genes bZIP_2, bZIP_6, bZIP_16, and bZIP_18 are enriched in GO categories, indicating their involvement in biological processes, molecular functions, and cellular components. From this, it can be inferred that these bZIP-related genes in the roots of T. ramosissima mediate the plant’s tolerance to NaCl through the ABA signaling pathway, thereby enhancing the plant’s inherent salt resistance.
Furthermore, Salicylic acid (SA) is an important plant hormone that participates in the plant’s response to stress through complex signal transduction networks [74]. Plant bZIP-type transcription factors are induced in their expression by SA [75]. In the plant hormone signal transduction pathway, SA primarily mediates the signaling pathway to establish an efficient defense response mechanism against the invasion of pathogens [76]. SA-mediated disease resistance pathways in plants are closely associated with TGA [77]. In this study, three bZIP-related genes (bZIP_6, bZIP_16, and bZIP_18) consistently showed upregulated expression levels in response to SA induction after 48 h and 168 h of exposure to exogenous potassium under NaCl stress.
In summary, as mentioned above, both bZIP_6 and bZIP_16, as bZIP-related genes, consistently showed a trend of upregulation in their expression levels after 48 h and 168 h of exogenous potassium addition under NaCl stress. Furthermore, they are induced by both ABA and SA in the plant hormone signal transduction pathway. Their upregulated expression plays a crucial role when their signal pathways are activated. From this, it is suggested that bZIP_6 and bZIP_16 can be regarded as key candidate genes related to bZIP, and their functions can be further verified. In conclusion, the random selection of nine key candidate bZIP genes for validation has confirmed the reliability and accuracy of the transcriptome data in this study.
5. Conclusions
The bZIP protein family plays a crucial role in plant growth and resisting adverse environmental stresses. In this study, under NaCl stress, after exogenous K+ was applied to the roots of T. ramosissima for 48 h and 168 h, we found that bZIP transcription factors were involved in constructing defense systems through related metabolic pathways to combat environmental adversity. They likely mediate signal transduction through various physiological pathways, clear the accumulation of ROS, reduce plant oxidative damage, and thus resist NaCl stress. Notably, adding K+ greatly enhances the plant’s K+/Na+ ratio, helping it combat the stress caused by NaCl. Additionally, K+ plays a vital role in maintaining the normal growth of T. ramosissima. Furthermore, bZIP transcription factors, mediated by the ABA signaling pathway in the plant hormone signal transduction pathway, enhance the salt resistance of T. ramosissima. Especially, bZIP_6 and bZIP_16, as bZIP-related genes, were labeled by the plant hormone signal transduction pathway under NaCl stress with the addition of exogenous K+ at 48 h and 168 h, and their expression consistently showed an upward trend. Hence, they might play a vital role as key candidate genes associated with bZIP in resisting NaCl damage. However, the regulatory mechanisms, metabolic products, and functional verification for alleviating NaCl toxicity by these genes still require further investigation.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/genes14122203/s1, Supplementary Table S1: Sequences of specific primers; Supplementary Table S2: Data filtering statistics table; Supplementary Table S3: Pfam prediction of protein domain structures in bZIP candidate genes; Supplementary Table S4: Analysis of bZIP genes enriched in GO and KEGG pathway; Supplementary Table S5: Analysis of plant hormone signal transduction pathway; Supplementary Table S6. Information sheet for the 10 homologous gene species; Supplementary Figure S1: Changes in the expression level of bZIP genes.
Author Contributions
Conceptualization, Y.C.; Data curation, Y.C.; Formal analysis, Y.C.; Funding acquisition, D.S. and L.W.; Investigation, Y.C., J.J. and L.W.; Methodology, Y.C. and J.J.; Project administration, M.Z., D.S. and L.W.; Resources, Y.C., J.J. and L.W.; Software, Y.C.; Supervision, Y.C., D.S. and L.W.; Validation, Y.C.; Visualization, Y.C., M.Z., D.S., J.J. and L.W.; Writing—original draft, Y.C.; Writing—review and editing, Y.C., M.Z., D.S., J.J. and L.W. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the Jiangsu Provincial Forestry Science and Technology Innovation and Promotion Project “Jiangsu Provincial Long-term Research Base for Breeding Salt-tolerant Native Tree Species (LYKJ2021108)” and “Jiangsu Coastal Tidal Spartina alterniflora Loisel Comprehensive Management Technology Innovation and Demonstration (Su [2023]TG10)”.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article and Supplementary Materials.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Gupta, A.; Rico-Medina, A.; Caño-Delgado, A.I. The physiology of plant responses to drought. Science 2020, 368, 266–269. [Google Scholar] [CrossRef] [PubMed]
- Mou, Z.; Fan, W.; Dong, X. Inducers of plant systemic acquired resistance regulate NPR1 function through redox changes. Cell 2003, 113, 935–944. [Google Scholar] [CrossRef] [PubMed]
- Aslam, M.; Fakher, B.; Ashraf, M.A.; Cheng, Y.; Wang, B.; Qin, Y. Plant low-temperature stress: Signaling and response. Agronomy 2022, 12, 702. [Google Scholar] [CrossRef]
- Ismail, A.M.; Horie, T. Genomics, physiology, and molecular breeding approaches for improving salt tolerance. Annu. Rev. Plant Biol. 2017, 68, 405–434. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Zhang, Q.; Liu, M.; Zhou, H.; Ma, C.; Wang, P. Regulation of plant responses to salt stress. Int. J. Mol. Sci. 2021, 22, 4609. [Google Scholar] [CrossRef] [PubMed]
- Ren, B.; Zhang, J.; Li, X.; Fan, X.; Dong, S.; Liu, P.; Zhao, B. Effects of waterlogging on the yield and growth of summer maize under field conditions. Can. J. Plant Sci. 2014, 94, 23–31. [Google Scholar] [CrossRef]
- Badis, G.; Berger, M.F.; Philippakis, A.A.; Talukder, S.; Gehrke, A.R.; Jaeger, S.A.; Chan, E.T.; Metzler, G.; Vedenko, A.; Chen, X.; et al. Diversity and complexity in DNA recognition by transcription factors. Science 2009, 324, 1720–1723. [Google Scholar] [CrossRef]
- Perez-Rodriguez, P.; Riano-Pachon, D.M.; Correa, L.G.; Rensing, S.A.; Kersten, B.; Mueller-Roeber, B. PlnTFDB: Updated content and new features of the plant transcription factor database. Nucleic Acids Res. 2010, 38, D822–D827. [Google Scholar] [CrossRef]
- Jakoby, M.; Weisshaar, B.; Dröge-Laser, W.; Vicente-Carbajosa, J.; Tiedemann, J.; Kroj, T.; Parcy, F.; BZIP, R.G. bZIP transcription factors in Arabidopsis. Trends Plant Sci. 2002, 7, 106–111. [Google Scholar] [CrossRef]
- Teng, Y.; Lv, M.; Zhang, X.; Cai, M.; Chen, T. BEAR1, a bHLH transcription factor, controls salt response genes to regulate rice salt response. J. Plant Biol. 2022, 65, 217–230. [Google Scholar] [CrossRef]
- So, H.; Lee, J. NAC Transcription factors from soybean (Glycine max L.) Differentially Regulated by Abiotic Stress. J. Plant Biol. 2019, 62, 147–160. [Google Scholar] [CrossRef]
- Dubos, C.; Stracke, R.; Grotewold, E.; Weisshaar, B.; Martin, C.; Lepiniec, L. MYB transcription factors in Arabidopsis. Plant Sci. 2010, 15, 573–581. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Liu, F.; Zhu, C.; Li, X.; Dai, X.; Yang, B.; Zou, X.; Ma, Y. Identification, expression, alternative splicing and functional analysis of pepper WRKY gene family in response to biotic and abiotic stresses. PLoS ONE 2019, 14, e219775. [Google Scholar] [CrossRef] [PubMed]
- Landschulz, W.H.; Johnson, P.F.; McKnight, S.L. The leucine zipper: A hypothetical structure common to a new class of DNA binding proteins. Science 1988, 240, 1759–1764. [Google Scholar] [CrossRef] [PubMed]
- Han, H.; Wang, C.; Yang, X.; Wang, L.; Ye, J.; Xu, F.; Liao, Y.; Zhang, W. Role of bZIP transcription factors in the regulation of plant secondary metabolism. Planta 2023, 258, 13. [Google Scholar] [CrossRef] [PubMed]
- Droge-Laser, W.; Snoek, B.L.; Snel, B.; Weiste, C. The Arabidopsis bZIP transcription factor family-an update. Curr. Opin. Plant Biol. 2018, 45, 36–49. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Gao, S.; Tang, Y.; Li, L.; Zhang, F.; Feng, B.; Fang, Z.; Ma, L.; Zhao, C. Genome-wide identification and evolutionary analyses of bZIP transcription factors in wheat and its relatives and expression profiles of anther development related TabZIP genes. BMC Genom. 2015, 16, 976. [Google Scholar] [CrossRef]
- Zhang, M.; Liu, Y.; Shi, H.; Guo, M.; Chai, M.; He, Q.; Yan, M.; Cao, D.; Zhao, L.; Cai, H.; et al. Evolutionary and expression analyses of soybean basic Leucine zipper transcription factor family. BMC Genom. 2018, 19, 159. [Google Scholar] [CrossRef]
- Wang, X.; Lu, X.; Malik, W.A.; Chen, X.; Wang, J.; Wang, D.; Wang, S.; Chen, C.; Guo, L.; Ye, W. Differentially expressed bZIP transcription factors confer multi-tolerances in Gossypium hirsutum L. Int. J. Biol. Macromol. 2020, 146, 569–578. [Google Scholar] [CrossRef]
- Gibalová, A.; Reňák, D.; Matczuk, K.; Dupl’Áková, N.; Cháb, D.; Twell, D.; Honys, D. AtbZIP34 is required for Arabidopsis pollen wall patterning and the control of several metabolic pathways in developing pollen. Plant Mol. Biol. 2009, 70, 581–601. [Google Scholar] [CrossRef]
- Van Leene, J.; Blomme, J.; Kulkarni, S.R.; Cannoot, B.; De Winne, N.; Eeckhout, D.; Persiau, G.; Van De Slijke, E.; Vercruysse, L.; Vanden, B.R.; et al. Functional characterization of the Arabidopsis transcription factor bZIP29 reveals its role in leaf and root development. J. Exp. Bot. 2016, 67, 5825–5840. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Liu, C.; Li, Z.; Ran, Q.; Xie, G.; Wang, B.; Fang, S.; Chu, J.; Zhang, J. ZmbZIP4 contributes to stress resistance in maize by regulating ABA synthesis and root development. Plant Physiol. 2018, 178, 753–770. [Google Scholar] [CrossRef] [PubMed]
- Mason, G.A. A bZIP transcription factor accelerates the transition to reproductive tuber growth and aging in Solanum tuberosum. Plant Physiol. 2022, 189, 1194–1195. [Google Scholar] [CrossRef] [PubMed]
- Hwan Yang, S.; Yamaguchi, Y.; Koizumi, N.; Kusano, T.; Sano, H. Promoter analysis of tbzF, a gene encoding a bZIP-type transcription factor, reveals distinct variation in cis-regions responsible for transcriptional activation between senescing leaves and flower buds in tobacco plants. Plant Sci. 2002, 162, 973–980. [Google Scholar] [CrossRef]
- Chang, C.; Liu, Z.; Wang, Y.; Tang, Z.; Yu, F. A bZIP transcription factor, CaLMF, mediated light-regulated camptothecin biosynthesis in Camptotheca acuminata. Tree Physiol. 2019, 39, 372–380. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.; Nguyen, B.H.; Xie, Y.; Xiao, B.; Tang, N.; Zhu, W.; Mou, T.; Xiong, L. Co-overexpression of the constitutively active form of OsbZIP46 and ABA-Activated protein kinase SAPK6 improves drought and temperature stress resistance in rice. Front. Plant Sci. 2017, 8, 1102. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Liu, W.; Qi, X.; Liu, Z.; Xie, W.; Wang, Y. Genome-wide identification, expression profiling, and SSR marker development of the bZIP transcription factor family in Medicago truncatula. Biochem. Syst. Ecol. 2015, 61, 218–228. [Google Scholar] [CrossRef]
- Ramakrishna, C.; Singh, S.; Raghavendrarao, S.; Padaria, J.C.; Mohanty, S.; Sharma, T.R.; Solanke, A.U. The membrane tethered transcription factor EcbZIP17 from finger millet promotes plant growth and enhances tolerance to abiotic stresses. Sci. Rep. 2018, 8, 2148. [Google Scholar] [CrossRef]
- Hsieh, T.H.; Li, C.W.; Su, R.C.; Cheng, C.P.; Sanjaya; Tsai, Y.C.; Chan, M.T. A tomato bZIP transcription factor, SlAREB, is involved in water deficit and salt stress response. Planta 2010, 231, 1459–1473. [Google Scholar] [CrossRef]
- Noh, M.; Huque, A.K.M.M.; Jung, K.W.; Kim, Y.Y.; Shin, J.S. A stress-responsive CaM-Binding transcription factor, bZIP4, confers abiotic stress resistance in Arabidopsis. J. Plant Biol. 2021, 64, 359–370. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, L.; Xia, C.; Gao, L.; Hao, C.; Zhao, G.; Jia, J.; Kong, X. A novel wheat C-bZIP gene, TabZIP14-B, participates in salt and freezing tolerance in Transgenic Plants. Front. Plant Sci. 2017, 8, 710. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Mao, B.; Ou, S.; Wang, W.; Liu, L.; Wu, Y.; Chu, C.; Wang, X. Correction to: OsbZIP71, a bZIP transcription factor, confers salinity and drought tolerance in rice. Plant Mol. Biol. 2018, 97, 467–468. [Google Scholar] [CrossRef] [PubMed]
- Choi, B.Y.; Kim, H.; Shim, D.; Jang, S.; Yamaoka, Y.; Shin, S.; Yamano, T.; Kajikawa, M.; Jin, E.; Fukuzawa, H.; et al. The Chlamydomonas bZIP transcription factor BLZ8 confers oxidative stress tolerance by inducing the carbon-concentrating mechanism. Plant Cell 2022, 34, 910–926. [Google Scholar] [CrossRef] [PubMed]
- Schlögl, P.S.; Nogueira, F.T.S.; Drummond, R.; Felix, J.M.; De Rosa, V.E.J.; Vicentini, R.; Leite, A.; Ulian, E.C.; Menossi, M. Identification of new ABA- and MEJA-activated sugarcane bZIP genes by data mining in the SUCEST database. Plant Cell Rep. 2008, 27, 335–345. [Google Scholar] [CrossRef] [PubMed]
- Sardans, J.; Penuelas, J. Potassium control of plant functions: Ecological and agricultural implications. Plants 2021, 10, 419. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.; Vishwakarma, K.; Hossen, M.S.; Kumar, V.; Shackira, A.M.; Puthur, J.T.; Abdi, G.; Sarraf, M.; Hasanuzzaman, M. Potassium in plants: Growth regulation, signaling, and environmental stress tolerance. Plant Physiol. Biochem. 2022, 172, 56–69. [Google Scholar] [CrossRef]
- Dreyer, I.; Gomez-Porras, J.L.; Riedelsberger, J. The potassium battery: A mobile energy source for transport processes in plant vascular tissues. New Phytol. 2017, 216, 1049–1053. [Google Scholar] [CrossRef]
- Shabala, S.; Pottosin, I. Regulation of potassium transport in plants under hostile conditions: Implications for abiotic and biotic stress tolerance. Physiol. Plant 2014, 151, 257–279. [Google Scholar] [CrossRef]
- Sustr, M.; Soukup, A.; Tylova, E. Potassium in root growth and development. Plants 2019, 8, 435. [Google Scholar] [CrossRef]
- Luan, M.; Tang, R.; Tang, Y.; Tian, W.; Hou, C.; Zhao, F.; Lan, W.; Luan, S. Transport and homeostasis of potassium and phosphate: Limiting factors for sustainable crop production. J. Exp. Bot. 2017, 68, 3091–3105. [Google Scholar] [CrossRef]
- Assaha, D.V.M.; Ueda, A.; Saneoka, H.; Al-Yahyai, R.; Yaish, M.W. The role of Na+ and K+ transporters in salt stress adaptation in glycophytes. Front. Physiol. 2017, 8, 509. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Kumar, T.; Singh, S.; Tuteja, N.; Prasad, R.; Singh, J. Potassium: A key modulator for cell homeostasis. J. Biotechnol. 2020, 324, 198–210. [Google Scholar] [CrossRef]
- Pandey, G.K.; Mahiwal, S. Potassium in Abiotic Stress; Springer International Publishing AG: Cham, Switzerland, 2020; pp. 45–49. [Google Scholar]
- Chen, Y.; Li, H.; Zhang, S.; Du, S.; Zhang, J.; Song, Z.; Jiang, J. Analysis of the main antioxidant enzymes in the roots of Tamarix ramosissima under NaCl stress by applying exogenous potassium (K+). Front. Plant Sci. 2023, 14, 1114266. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.; Su, Y. Distinctive potassium-accumulation capability of Alligatorweed (Alternanthera philoxeroides) links to high-affinity potassium transport facilitated by K+-Uptake systems. Weed Sci. 2013, 61, 77–84. [Google Scholar] [CrossRef]
- Lei, S.; Wang, W.; Wang, Y.; Chen, L.; Wu, X.; Deng, Z.; Zhang, T.; Du, Z.; Li, J. Effects of Tamarix ramosissima seedlings on Populus euphratica seedlings under various water and salinity conditions. Acta Ecol. Sin. 2020, 40, 7638–7647. (In Chinese) [Google Scholar] [CrossRef]
- Duan, Q.; Zhu, Z.; Wang, B.; Chen, M. Recent Progress on the Salt Tolerance Mechanisms and Application of Tamarisk. Int. J. Mol. Sci. 2022, 23, 3325. [Google Scholar] [CrossRef] [PubMed]
- Bosabalidis, A.M. Programmed cell death in salt glands of Tamarix aphylla L.: An electron microscope analysis. Cent. Eur. J. Biol. 2012, 7, 927–930. [Google Scholar] [CrossRef]
- Wang, J.; Zou, A.; Xiang, S.; Liu, C.; Peng, H.; Wen, Y.; Ma, X.; Chen, H.; Ran, M.; Sun, X. Transcriptome analysis reveals the mechanism of zinc ion-mediated plant resistance to TMV in Nicotiana benthamiana. Pestic. Biochem. Phys. 2022, 184, 105100. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, G.; Zhang, H.; Zhang, N.; Jiang, J.; Song, Z. Transcriptome analysis of Tamarix ramosissima leaves in response to NaCl stress. PLoS ONE 2022, 17, e265653. [Google Scholar] [CrossRef]
- Conesa, A.; Gotz, S.; Garcia-Gomez, J.M.; Terol, J.; Talon, M.; Robles, M. Blast2GO: A universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef] [PubMed]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene Ontology: Tool for the unification of biology. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Coggill, P.; Eberhardt, R.Y.; Eddy, S.R.; Mistry, J.; Mitchell, A.L.; Potter, S.C.; Punta, M.; Qureshi, M.; Sangrador-Vegas, A.; et al. The Pfam protein families database: Towards a more sustainable future. Nucleic Acids Res. 2016, 44, D279–D285. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−∆∆CT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Li, L.; ShangGuan, G.; Jia, C.; Deng, S.; Noman, M.; Liu, Y.; Guo, Y.; Han, L.; Zhang, X.; et al. Genome-wide identification and expression analysis of bZIP gene family in Carthamus tinctorius L. Sci. Rep. 2020, 10, 15521. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.; Gan, S.; Jiao, J.; He, Y.; Liu, H.; Yin, X.; Zhu, Q.; Rao, J. Genome-wide analysis of the bZIP gene family and the role of AchnABF1 from postharvest kiwifruit (Actinidia chinensis cv. Hongyang) in osmotic and freezing stress adaptations. Plant Sci. 2021, 308, 110927. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, Y.; Zhou, R.; Dossa, K.; Yu, J.; Li, D.; Liu, A.; Mmadi, M.A.; Zhang, X.; You, J. Identification and characterization of the bZIP transcription factor family and its expression in response to abiotic stresses in sesame. PLoS ONE 2018, 13, e200850. [Google Scholar] [CrossRef]
- Jiang, M.; Wang, Z.; Ren, W.; Yan, S.; Xing, N.; Zhang, Z.; Li, H.; Ma, W. Identification of the bZIP gene family and regulation of metabolites under salt stress in isatis indigotica. Front. Plant Sci. 2022, 13, 1011616. [Google Scholar] [CrossRef]
- Horberg, J.; Reymer, A. Specifically bound BZIP transcription factors modulate DNA supercoiling transitions. Sci. Rep. 2020, 10, 18795. [Google Scholar] [CrossRef]
- Zhao, P.; Ye, M.; Wang, R.; Wang, D.; Chen, Q. Systematic identification and functional analysis of potato (Solanum tuberosum L.) bZIP transcription factors and overexpression of potato bZIP transcription factor StbZIP-65 enhances salt tolerance. Int. J. Biol. Macromol. 2020, 161, 155–167. [Google Scholar] [CrossRef]
- Yang, Y.; Li, J.; Li, H.; Yang, Y.; Guang, Y.; Zhou, Y. The bZIP gene family in watermelon: Genome-wide identification and expression analysis under cold stress and root-knot nematode infection. PeerJ 2019, 7, e7878. [Google Scholar] [CrossRef] [PubMed]
- Chang, Q.; Lu, X.; Liu, Z.; Zheng, Z.; Yu, S. Identification and characterization of the bZIP transcription factor family in yellowhorn. J. For. Res. 2021, 32, 273–284. [Google Scholar] [CrossRef]
- Liu, J.; Chen, N.; Chen, F.; Cai, B.; Dal Santo, S.; Tornielli, G.B.; Pezzotti, M.; Cheng, Z.M. Genome-wide analysis and expression profile of the bZIP transcription factor gene family in grapevine (Vitis vinifera). BMC Genom. 2014, 15, 281. [Google Scholar] [CrossRef]
- Zhao, J.; Guo, R.; Guo, C.; Hou, H.; Wang, X.; Gao, H. Evolutionary and expression analyses of the apple basic leucine zipper transcription factor family. Front. Plant Sci. 2016, 7, 376. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Tang, X.; Zhang, N.; Li, S.; Si, H. Role of bZIP Transcription Factors in Plant Salt Stress. Int. J. Mol. Sci. 2023, 24, 7893. [Google Scholar] [CrossRef]
- Ji, X.; Liu, G.; Liu, Y.; Zheng, L.; Nie, X.; Wang, Y. The bZIP protein from Tamarix hispida, ThbZIP1, is ACGT elements binding factor that enhances abiotic stress signaling in transgenic Arabidopsis. BMC Plant Biol. 2013, 13, 151. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Gao, C.; Liang, Y.; Wang, C.; Yang, C.; Liu, G. A novel bZIP gene from Tamarix hispida mediates physiological responses to salt stress in tobacco plants. J. Plant Physiol. 2010, 167, 222–230. [Google Scholar] [CrossRef]
- Xiong, L.; Schumaker, K.S.; Zhu, J.K. Cell signaling during cold, drought, and salt stress. Plant Cell 2002, 14 (Suppl. 1), S165–S183. [Google Scholar] [CrossRef]
- Chen, K.; Li, G.J.; Bressan, R.A.; Song, C.P.; Zhu, J.K.; Zhao, Y. Abscisic acid dynamics, signaling, and functions in plants. J. Integr. Plant Biol. 2020, 62, 25–54. [Google Scholar] [CrossRef]
- Dar, N.A.; Amin, I.; Wani, W.; Wani, S.A.; Shikari, A.B.; Wani, S.H.; Masoodi, K.Z. Abscisic acid: A key regulator of abiotic stress tolerance in plants. Plant Gene 2017, 11, 106–111. [Google Scholar] [CrossRef]
- Uno, Y.; Furihata, T.; Abe, H.; Yoshida, R.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Arabidopsis basic leucine zipper transcription factors involved in an abscisic acid-dependent signal transduction pathway under drought and high-salinity conditions. Proc. Natl. Acad. Sci. USA 2000, 97, 11632–11637. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi-Shinozaki, K.; Shinozaki, K. Organization of cis-acting regulatory elements in osmotic- and cold-stress-responsive promoters. Trends Plant Sci. 2005, 10, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Zhao, H.; Wang, X.; Kang, J.; Lv, B.; Dong, Q.; Li, C.; Chen, H.; Wu, Q. Tartary buckwheat transcription factor FtbZIP5, Regulated by FtSnRK2.6, Can improve salt/drought resistance in transgenic Arabidopsis. Int. J. Mol. Sci. 2020, 21, 1123. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Zheng, J.; Zhang, X.; Hu, Q.; Qian, R. Salicylic Acid Alleviates the Adverse Effects of Salt Stress on Dianthus superbus (Caryophyllaceae) by Activating Photosynthesis, Protecting Morphological Structure, and Enhancing the Antioxidant System. Front. Plant Sci. 2017, 8, 600. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, G.; Xia, N.; Wang, X.; Huang, L.; Kang, Z. Cloning and characterization of a bZIP transcription factor gene in wheat and its expression in response to stripe rust pathogen infection and abiotic stresses. Physiol. Mol. Plant Pathol. 2008, 73, 88–94. [Google Scholar] [CrossRef]
- Shah, J. The salicylic acid loop in plant defense. Curr. Opin. Plant Biol. 2003, 6, 365–371. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).