
Role of Different Members of the AGPAT Gene Family in Milk Fat Synthesis in Bubalus bubalis

 ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Animals and Sampling
2.2. Extraction of Total RNA from Milk Fat Globules
2.3. Single-Strand cDNA Synthesis
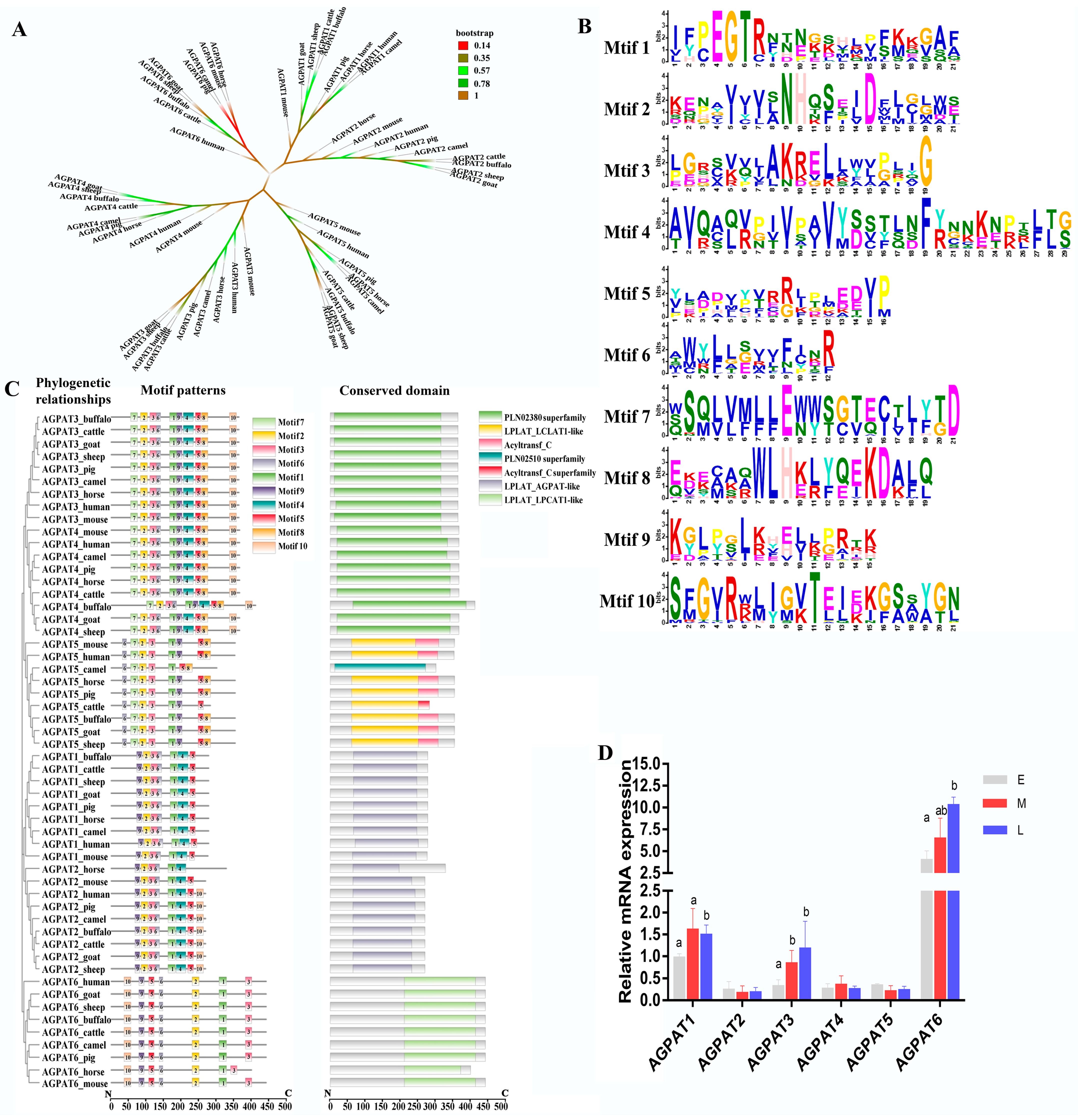
2.4. Phylogenetic, Secondary Structure, and Multiple Sequence Alignment Analyses
2.5. Isolation, Culture, and Purification of Buffalo Mammary Epithelial Cells
2.6. siRNA Synthesis and Overexpression Vector Construction of Buffalo AGPAT1, AGPAT3, and AGPAT4
2.7. Transfection of BMECs
2.8. Extraction and Component Analysis of Fatty Acids
2.9. qRT-PCR Analysis
2.10. Statistical Analysis
3. Results
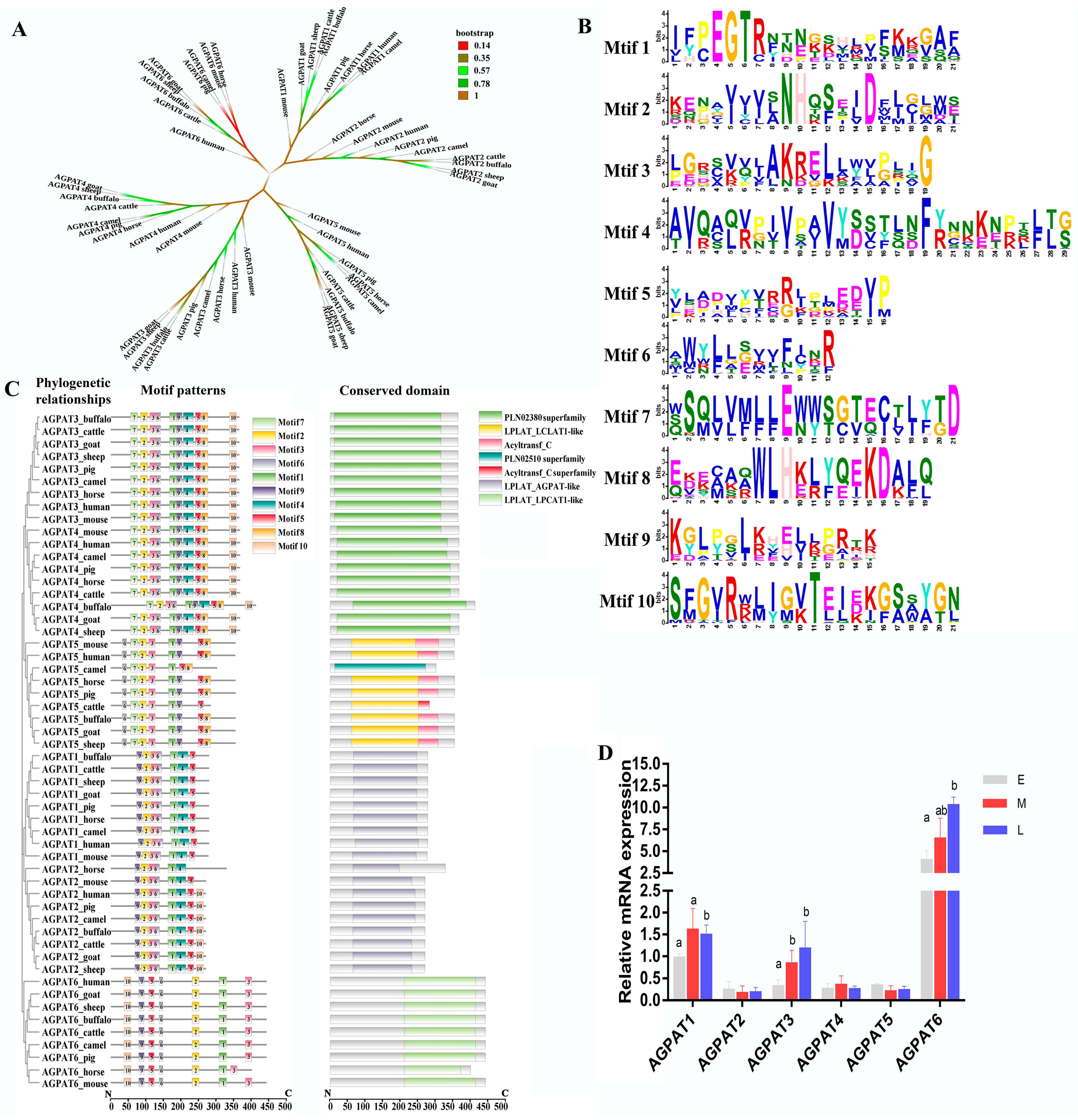
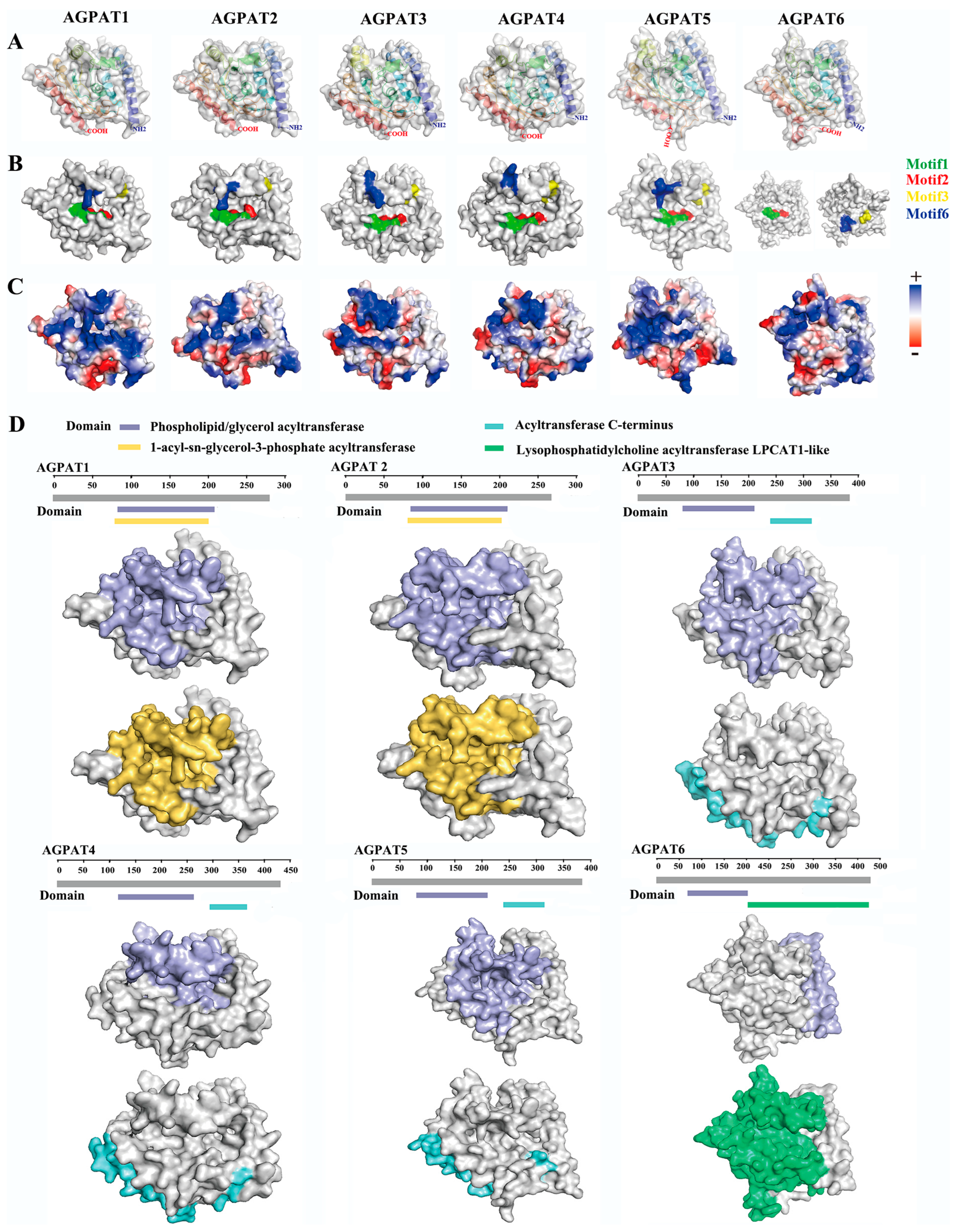
3.1. Analysis of Amino Acid and Protein Domain Conservation of Buffalo AGPAT Protein
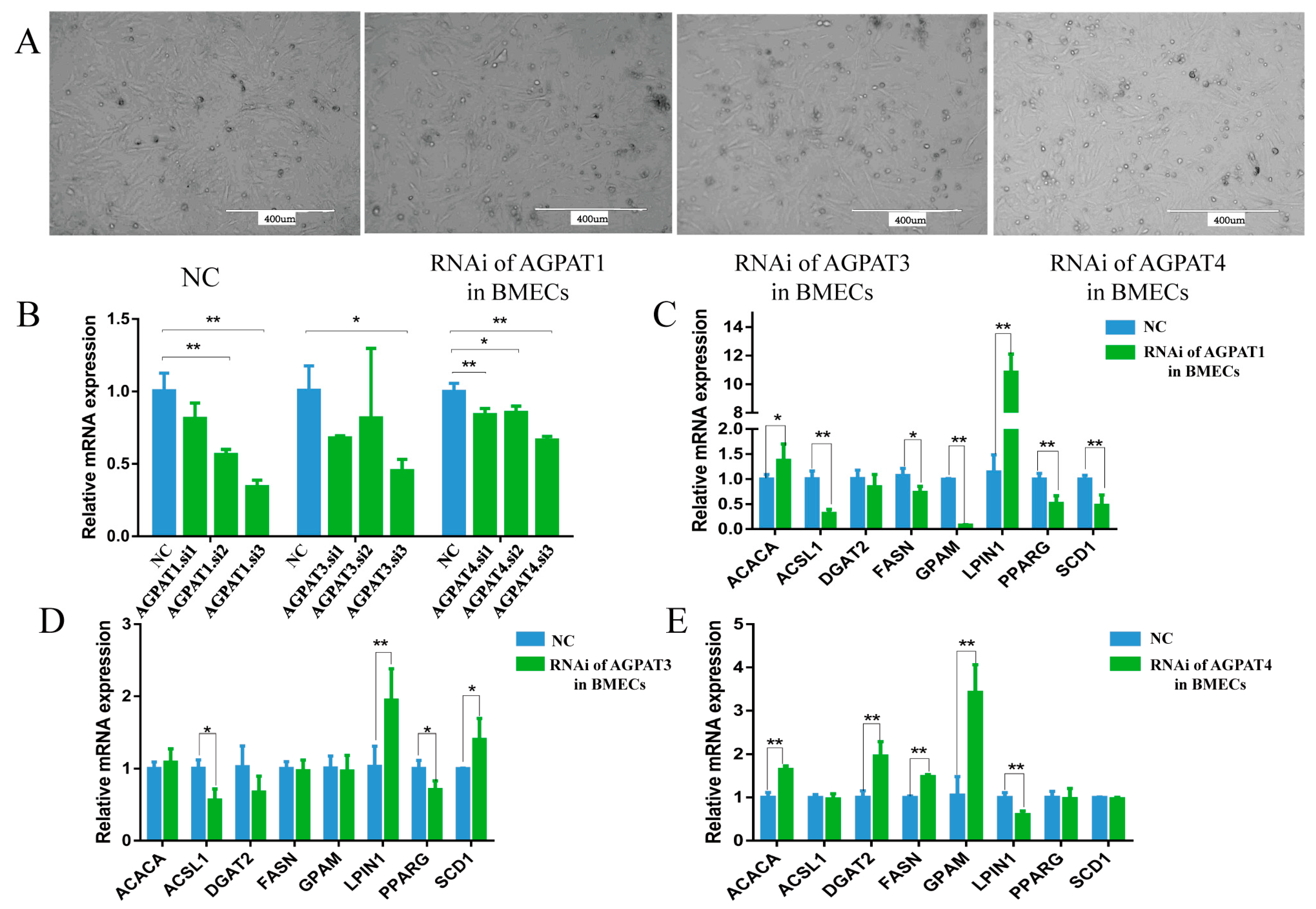
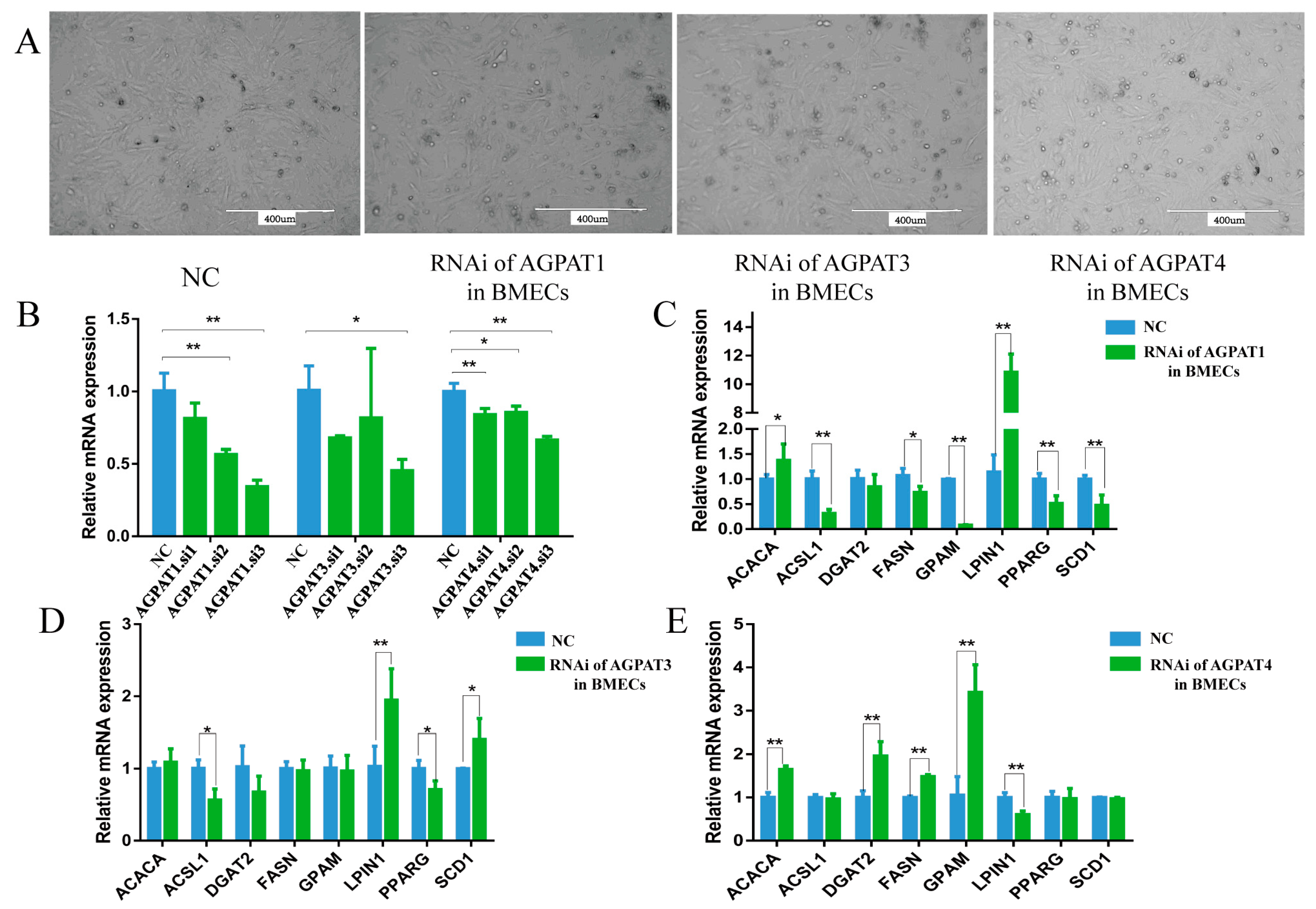
3.2. Effect of AGPAT Interference on Gene Expression and Fat Synthesis in BMECs
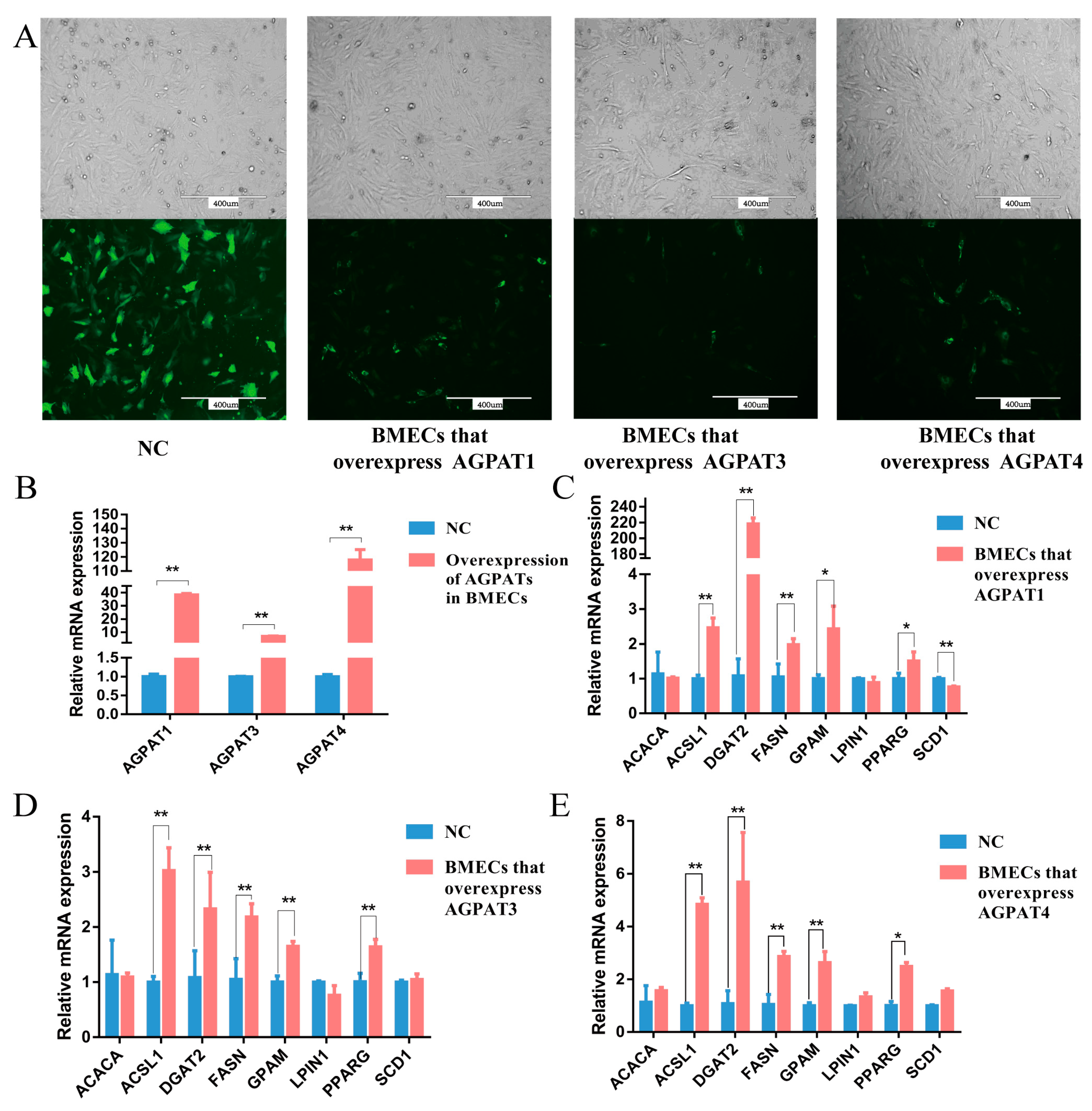
3.3. Effects of AGPATs Overexpression on Gene Expression and Fat Synthesis in BMECs
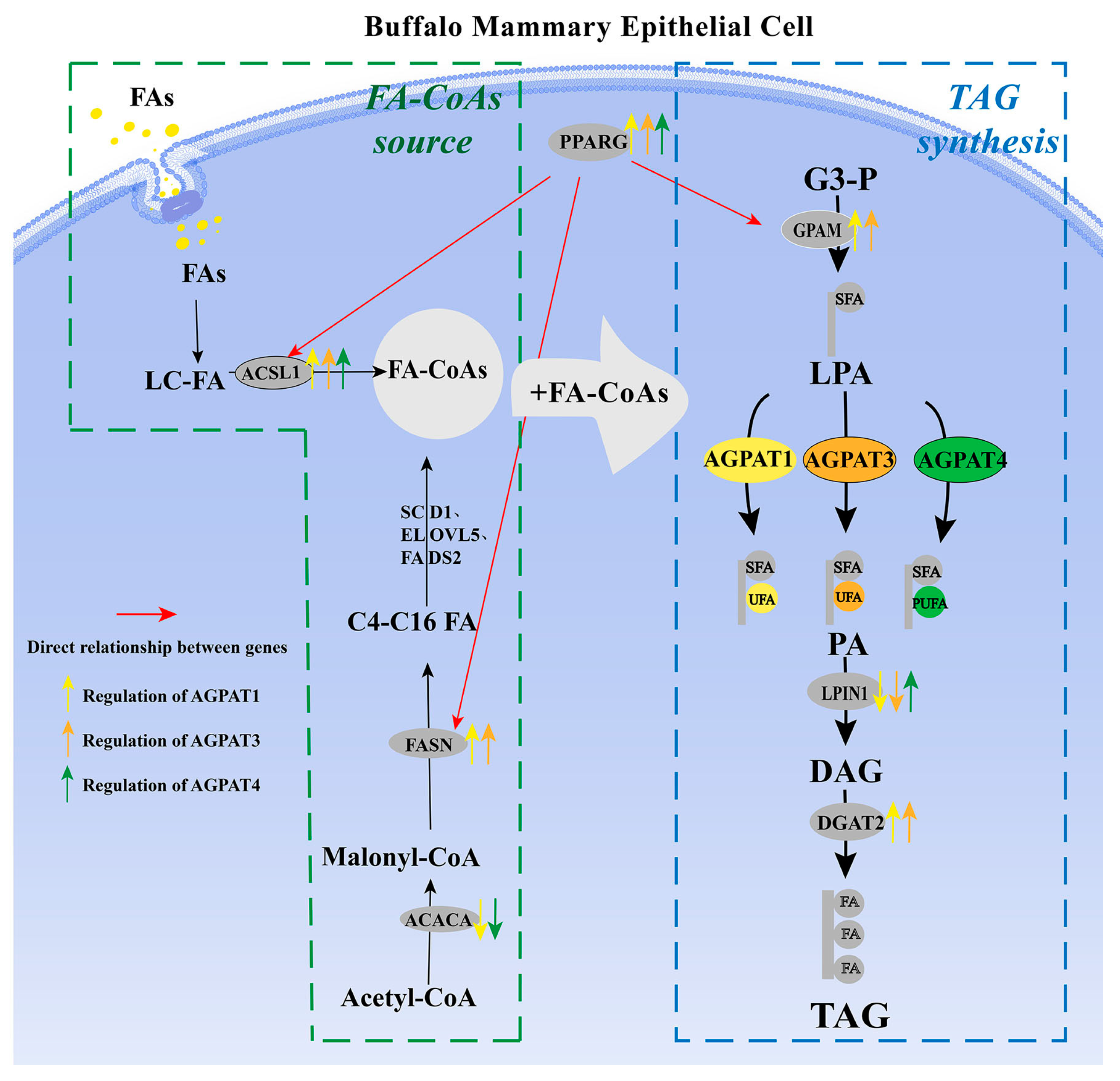
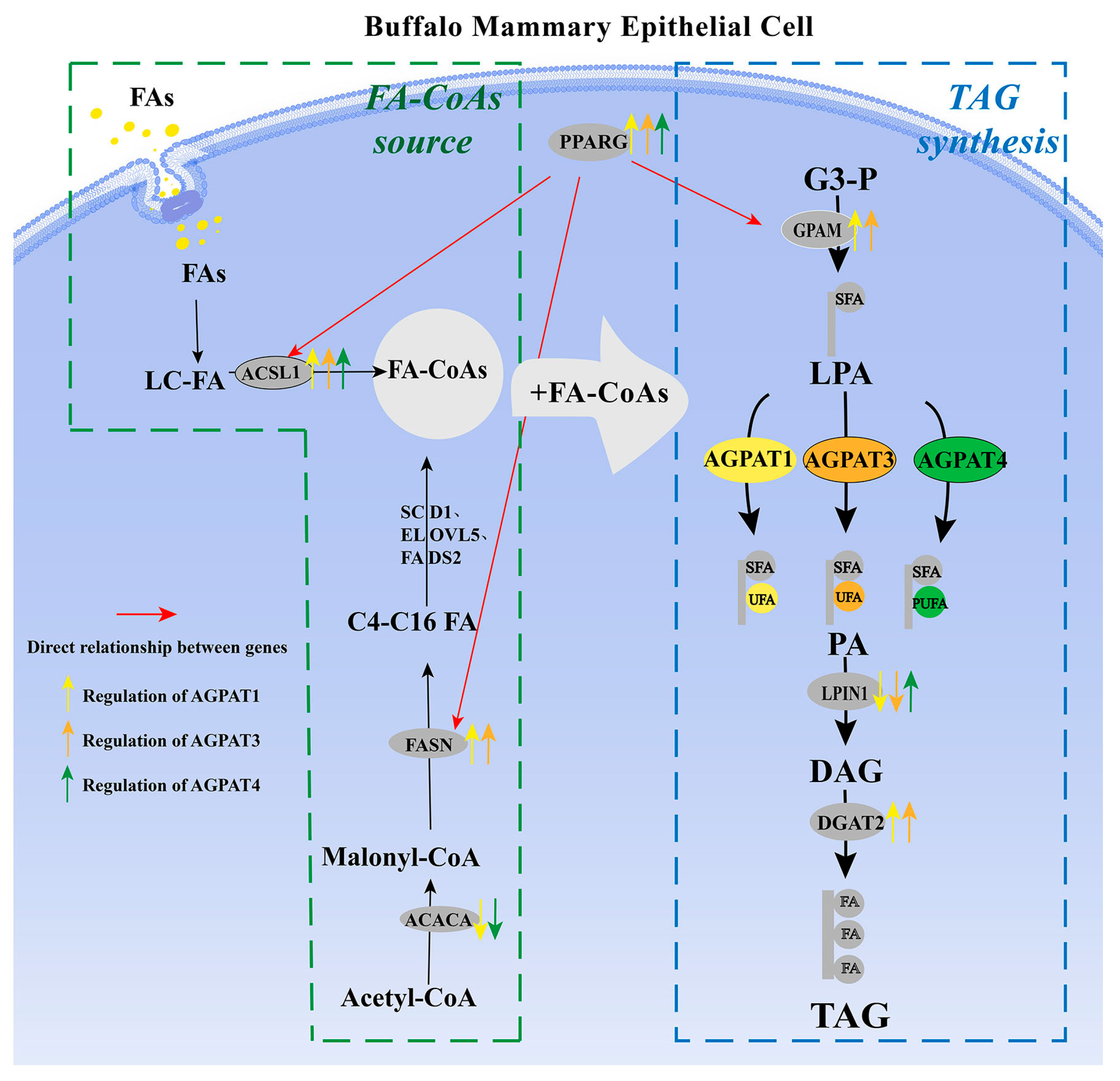
3.4. Potential Molecular Mechanism of AGPAT Gene Family Regulating Fat Synthesis in BMECs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, Z.; Lu, S.; Cui, K.; Shafique, L.; Luo, C.; Wang, Z.; Ruan, J.; Qian, Q.; Liu, Q. Fatty acid biosynthesis and transcriptional regulation of Stearoyl-CoA Desaturase 1 (SCD1) in buffalo milk. BMC Genet. 2020, 21, 23. [Google Scholar] [CrossRef] [PubMed]
- Minervino, A.H.H.; Zava, M.; Vecchio, D.; Borghese, A. Bubalus bubalis: A short story. Front. Vet. Sci. 2020, 7, 570413. [Google Scholar] [CrossRef] [PubMed]
- Medhammar, E.; Wijesinha Bettoni, R.; Stadlmayr, B.; Nilsson, E.; Charrondiere, U.R.; Burlingame, B. Composition of milk from minor dairy animals and buffalo breeds: A biodiversity perspective. J. Sci. Food Agric. 2012, 92, 445–474. [Google Scholar] [CrossRef]
- Žáček, P.; Bukowski, M.; Mehus, A.; Johnson, L.; Zeng, H.; Raatz, S.; Idso, J.P.; Picklo, M. Dietary saturated fatty acid type impacts obesity-induced metabolic dysfunction and plasma lipidomic signatures in mice. J. Nutr. Biochem. 2019, 64, 32–44. [Google Scholar] [CrossRef] [PubMed]
- Nie, P.; Pan, B.; Ahmad, M.J.; Zhang, X.; Chen, C.; Yao, Z.; Lv, H.; Wei, K.; Yang, L. Summer buffalo milk produced in China: A desirable diet enriched in polyunsaturated fatty acids and amino acids. Foods 2022, 11, 3475. [Google Scholar] [CrossRef]
- Bionaz, M.; Loor, J.J. Gene networks driving bovine milk fat synthesis during the lactation cycle. BMC Genom. 2008, 9, 366. [Google Scholar] [CrossRef] [PubMed]
- Mu, T.; Hu, H.; Ma, Y.; Feng, X.; Zhang, J.; Gu, Y. Regulation of key genes for milk fat synthesis in ruminants. Front. Nutr. 2021, 8, 765147. [Google Scholar] [CrossRef]
- Takeuchi, K.; Reue, K. Biochemistry, physiology, and genetics of GPAT, AGPAT, and lipin enzymes in triglyceride synthesis. Am. J. Physiol. Endocrinol. Metab. 2009, 296, E1195–E1209. [Google Scholar] [CrossRef]
- Ma, X.; Duan, A.; Lu, X.; Liang, S.; Sun, P.; Sohel, M.M.H.; Abdel-Shafy, H.; Amin, A.; Liang, A.; Deng, T. Novel Insight into the Potential Role of Acylglycerophosphate Acyltransferases Family Members on Triacylglycerols Synthesis in Buffalo. Int. J. Mol. Sci. 2022, 23, 6561. [Google Scholar] [CrossRef]
- Dircks, L.K.; Sul, H.S. Mammalian mitochondrial glycerol-3-phosphate acyltransferase. Biochim. Biophys. Acta (BBA)-Lipids Lipid Metab. 1997, 1348, 17–26. [Google Scholar] [CrossRef]
- Mardian, E.B.; Bradley, R.M.; Henao, J.J.A.; Marvyn, P.M.; Moes, K.A.; Bombardier, E.; Tupling, A.R.; Stark, K.D.; Duncan, R.E. Agpat4/Lpaatδ deficiency highlights the molecular heterogeneity of epididymal and perirenal white adipose depots. J. Lipid Res. 2017, 58, 2037–2050. [Google Scholar] [CrossRef] [PubMed]
- Koeberle, A.; Shindou, H.; Harayama, T.; Yuki, K.; Shimizu, T. Polyunsaturated fatty acids are incorporated into maturating male mouse germ cells by lysophosphatidic acid acyltransferase 3. FASEB J. 2012, 26, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Arora, R.; Sharma, A.; Sharma, U.; Girdhar, Y.; Kaur, M.; Kapoor, P.; Ahlawat, S.; Vijh, R.K. Buffalo milk transcriptome: A comparative analysis of early, mid and late lactation. Sci. Rep. 2019, 9, 5993. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef]
- Bailey, T.L.; Johnson, J.; Grant, C.E.; Noble, W.S. The MEME suite. Nucleic Acids Res. 2015, 43, W39–W49. [Google Scholar] [CrossRef]
- Sayers, E.W.; Bolton, E.E.; Brister, J.R.; Canese, K.; Chan, J.; Comeau, D.C.; Connor, R.; Funk, K.; Kelly, C.; Kim, S. Database resources of the national center for biotechnology information. Nucleic Acids Res. 2022, 50, D20. [Google Scholar] [CrossRef]
- Wang, J.; Chitsaz, F.; Derbyshire, M.K.; Gonzales, N.R.; Gwadz, M.; Lu, S.; Marchler, G.H.; Song, J.S.; Thanki, N.; Yamashita, R.A. The conserved domain database in 2023. Nucleic Acids Res. 2023, 51, D384–D388. [Google Scholar] [CrossRef]
- Chen, C.; Wu, Y.; Xia, R. A painless way to customize Circos plot: From data preparation to visualization using TBtools. iMeta 2022, 1, e35. [Google Scholar] [CrossRef]
- Kelley, L.A.; Mezulis, S.; Yates, C.M.; Wass, M.N.; Sternberg, M.J. The Phyre2 web portal for protein modeling, prediction and analysis. Nat. Protoc. 2015, 10, 845–858. [Google Scholar] [CrossRef] [PubMed]
- DeLano Scientific LLC. The PyMOL Molecular Graphics System, version 2.5.2 ed.; DeLano Scientific LLC: San Francisco, CA, USA, 2022. [Google Scholar]
- Paysan-Lafosse, T.; Blum, M.; Chuguransky, S.; Grego, T.; Pinto, B.L.; Salazar, G.A.; Bileschi, M.L.; Bork, P.; Bridge, A.; Colwell, L. InterPro in 2022. Nucleic Acids Res. 2023, 51, D418–D427. [Google Scholar] [CrossRef]
- Zhou, F.; Fan, X.; Miao, Y. LPIN1 promotes triglycerides synthesis and is transcriptionally regulated by PPARG in buffalo mammary epithelial cells. Sci. Rep. 2022, 12, 2390. [Google Scholar] [CrossRef]
- Benson, D.A.; Cavanaugh, M.; Clark, K.; Karsch-Mizrachi, I.; Ostell, J.; Pruitt, K.D.; Sayers, E.W. GenBank. Nucleic Acids Res. 2018, 46, D41. [Google Scholar] [CrossRef]
- Rychlik, W. OLIGO 7 primer analysis software. PCR Primer Des. 2007, 402, 35–59. [Google Scholar]
- Zheng, M.; Chen, X.; Cui, Y.; Li, W.; Dai, H.; Yue, Q.; Zhang, H.; Zheng, Y.; Guo, X.; Zhu, H. TULP2, a new RNA-binding protein, is required for mouse spermatid differentiation and male fertility. Front. Cell. Dev. Biol. 2021, 9, 623738. [Google Scholar] [CrossRef] [PubMed]
- Rao, X.; Huang, X.; Zhou, Z.; Lin, X. An improvement of the 2ˆ(–delta delta CT) method for quantitative real-time polymerase chain reaction data analysis. Biostat. Bioinform. Biomath. 2013, 3, 71. [Google Scholar]
- Kaur, R.; Sodhi, M.; Sharma, A.; Sharma, V.L.; Verma, P.; Swami, S.K.; Kumari, P.; Mukesh, M. Selection of suitable reference genes for normalization of quantitative RT-PCR (RT-qPCR) expression data across twelve tissues of riverine buffaloes (Bubalus bubalis). PLoS ONE 2018, 13, e0191558. [Google Scholar] [CrossRef]
- Chen, Q.; Wu, Y.; Zhang, M.; Xu, W.; Guo, X.; Yan, X.; Deng, H.; Jiang, Q.; Yang, X.; Lan, G. Milk fat globule is an alternative to mammary epithelial cells for gene expression analysis in buffalo. J. Dairy Res. 2016, 83, 202–208. [Google Scholar] [CrossRef]
- Valentine, W.J.; Yanagida, K.; Kawana, H.; Kono, N.; Noda, N.N.; Aoki, J.; Shindou, H. Update and nomenclature proposal for mammalian lysophospholipid acyltransferases, which create membrane phospholipid diversity. J. Biol. Chem. 2022, 298, 101470. [Google Scholar] [CrossRef]
- Lv, Y.; Guan, W.; Qiao, H.; Wang, C.; Chen, F.; Zhang, Y.; Liao, Z. Veterinary medicine and omics (veterinomics): Metabolic transition of milk triacylglycerol synthesis in sows from late pregnancy to lactation. Omics A J. Integr. Biol. 2015, 19, 602–616. [Google Scholar] [CrossRef]
- Bionaz, M.; Loor, J.J. ACSL1, AGPAT6, FABP3, LPIN1, and SLC27A6 are the most abundant isoforms in bovine mammary tissue and their expression is affected by stage of lactation. J. Nutr. 2008, 138, 1019–1024. [Google Scholar] [CrossRef]
- Zhou, F.; Xue, J.; Shan, X.; Qiu, L.; Miao, Y. Functional roles for AGPAT6 in milk fat synthesis of buffalo mammary epithelial cells. Anim. Biotechnol. 2022, 34, 2120–2131. [Google Scholar] [CrossRef] [PubMed]
- Zhukovsky, M.A.; Filograna, A.; Luini, A.; Corda, D.; Valente, C. The Structure and Function of Acylglycerophosphate Acyltransferase 4/Lysophosphatidic Acid Acyltransferase Delta (AGPAT4/LPAATδ). Front. Cell. Dev. Biol. 2019, 7, 147. [Google Scholar] [CrossRef]
- Costa, S.; Sampaio, L.; Berta Sousa, A.; Xing, C.; Agarwal, A.K.; Garg, A. Face-sparing congenital generalized lipodystrophy type 1 associated with nonclassical congenital adrenal hyperplasia. J. Clin. Endocrinol. Metab. 2022, 107, 2433–2438. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, A.; Nakanishi, H.; Suzuki, H.; Kamata, R.; Tanaka, K.; Waku, K.; Sugiura, T. Topology of acyltransferase motifs and substrate specificity and accessibility in 1-acyl-sn-glycero-3-phosphate acyltransferase 1. Biochim. Et Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2007, 1771, 1202–1215. [Google Scholar] [CrossRef] [PubMed]
- Hishikawa, D.; Shindou, H.; Kobayashi, S.; Nakanishi, H.; Taguchi, R.; Shimizu, T. Discovery of a lysophospholipid acyltransferase family essential for membrane asymmetry and diversity. Proc. Natl. Acad. Sci. USA 2008, 105, 2830–2835. [Google Scholar] [CrossRef]
- Ding, L.; Shen, Y.; Jawad, M.; Wu, T.; Maloney, S.K.; Wang, M.; Chen, N.; Blache, D. Effect of arginine supplementation on the production of milk fat in dairy cows. J. Dairy Sci. 2022, 105, 8115–8129. [Google Scholar] [CrossRef]
- Yoon, H.; Shaw, J.L.; Haigis, M.C.; Greka, A. Lipid metabolism in sickness and in health: Emerging regulators of lipotoxicity. Mol. Cell 2021, 81, 3708–3730. [Google Scholar] [CrossRef]
- Demmelmair, H.; Koletzko, B. Lipids in human milk. Best Pract. Res. Clin. Endoc. Metab. 2018, 32, 57–68. [Google Scholar] [CrossRef]
- Wang, H.; Airola, M.V.; Reue, K. How lipid droplets “TAG” along: Glycerolipid synthetic enzymes and lipid storage. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2017, 1862, 1131–1145. [Google Scholar] [CrossRef]
- Bradley, R.M.; Duncan, R.E. The lysophosphatidic acid acyltransferases (acylglycerophosphate acyltransferases) family: One reaction, five enzymes, many roles. Curr. Opin. Lipidol. 2018, 29, 110–115. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.K.; Sukumaran, S.; Cortés, V.A.; Tunison, K.; Mizrachi, D.; Sankella, S.; Gerard, R.D.; Horton, J.D.; Garg, A. Human 1-acylglycerol-3-phosphate O-acyltransferase isoforms 1 and 2: Biochemical characterization and inability to rescue hepatic steatosis in Agpat2−/− gene lipodystrophic mice. J. Biol. Chem. 2011, 286, 37676–37691. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Wang, C.; Chang, Z.H.; Guo, B.L.; Li, R.; Yue, X.P.; Lan, X.Y.; Chen, H.; Lei, C.Z. AGPAT6 polymorphism and its association with milk traits of dairy goats. Genet. Mol. Res. 2011, 10, 2747–2756. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Li, S.; Ge, G. Apatinib promotes ferroptosis in colorectal cancer cells by targeting ELOVL6/ACSL4 signaling. Cancer Manag. Res. 2021, 13, 1333–1342. [Google Scholar] [CrossRef]
- Zhao, Z.; Raza, S.H.A.; Tian, H.; Shi, B.; Luo, Y.; Wang, J.; Liu, X.; Li, S.; Bai, Y.; Hu, J. Effects of overexpression of ACSL1 gene on the synthesis of unsaturated fatty acids in adipocytes of bovine. Arch. Biochem. Biophys. 2020, 695, 108648. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AGPAT1 | AGPAT2 | AGPAT3 | AGPAT4 | AGPAT5 | AGPAT6 | |
|---|---|---|---|---|---|---|
| Number of amino acids | 287 | 278 | 376 | 425 | 365 | 456 |
| Disordered | 13% | 14% | 12% | 19% | 11% | 14% |
| α helix | 52% | 52% | 56% | 53% | 56% | 58% |
| β strand | 14% | 14% | 12% | 10% | 14% | 11% |
| TM helix | 16% | 17% | 32% | 29% | 24% | 29% |
| Fatty Acid | NC | AGPAT1 RNAi | AGPAT3 RNAi | AGPAT4 RNAi |
|---|---|---|---|---|
| C10:0 | 32.11 ± 1.08 | 12.43 ± 3.01 * | 12.90 ± 2.49 * | 15.03 ± 0.47 * |
| C14:0 | 44.97 ± 1.21 | 23.53 ± 0.58 * | 26.53 ± 1.13 * | 39.13 ± 2.96 |
| C15:0 | 22.77 ± 0.59 | 13.23 ± 0.64 * | 15.00 ± 0.45 * | 20.53 ± 1.51 |
| C16:0 | 120.40 ± 2.27 | 104.87 ± 2.30 * | 119.27 ± 3.14 | 212.00 ± 25.27 * |
| C17:0 | 23.92 ± 0.50 | 16.70 ± 0.60 * | 17.93 ± 0.22 * | 24.27 ± 1.71 |
| C18:0 | 89.91 ± 2.55 | 119.00 ± 5.69 * | 122.03 ± 3.87 * | 173.63 ± 3.55 * |
| C20:0 | 48.98 ± 1.79 | 29.93 ± 1.33 * | 31.83 ± 0.55 * | 41.50 ± 2.99 * |
| C22:0 | 52.44 ± 1.41 | 41.30 ± 1.42 * | 42.50 ± 0.70 * | 54.13 ± 5.06 |
| C23:0 | 23.89 ± 0.57 | 16.67 ± 0.33 * | 15.60 ± 1.17 * | 20.23 ± 1.53 |
| C16:1 | 24.36 ± 0.76 | 17.47 ± 0.47 * | 19.97 ± 0.27 * | 29.20 ± 4.13 |
| C18:1n9c | 90.58 ± 2.46 | 43.87 ± 1.56 * | 44.60 ± 2.14 * | 85.60 ± 5.70 |
| C18:2n6c | 30.86 ± 1.64 | 23.50 ± 0.38 * | 27.23 ± 0.43 | 42.77 ± 3.47 * |
| C18:3n3 | 25.76 ± 0.35 | 9.33 ± 0.89 * | 11.97 ± 1.51 * | 16.93 ± 1.33 * |
| C20:1 | 24.81 ± 0.77 | 25.27 ± 0.67 | 26.93 ± 0.92 | 36.23 ± 2.18 * |
| C20:3n6 | 28.89 ± 1.11 | 18.50 ± 0.35 * | 21.37 ± 0.63 * | 24.47 ± 1.11 * |
| C20:3n3 | 26.00 ± 1.17 | 6.97 ± 1.52 * | 7.50 ± 0.87 * | 11.30 ± 2.12 * |
| C22:1n9 | 79.03 ± 3.05 | 121.03 ± 2.63 * | 114.90 ± 2.77 * | 155.07 ± 9.49 * |
| C20:4n6 | 30.55 ± 1.94 | 17.53 ± 2.22 * | 21.77 ± 1.55 * | 28.73 ± 3.24 |
| C22:2 | 23.98 ± 0.57 | 19.20 ± 0.29 * | 19.67 ± 1.41 * | 27.90 ± 1.36 |
| C20:5n3 | 26.07 ± 1.04 | — | — | 4.40 ± 0.35 |
| C24:1 | 26.93 ± 0.82 | 30.63 ± 1.07 | 31.23 ± 0.69 * | 42.67 ± 3.50 * |
| C22:6n3 | 22.53 ± 0.64 | 14.10 ± 1.75 * | 20.10 ± 1.90 | 25.50 ± 3.48 |
| LCFA | 887.63 ± 12.78 | 713.97 ± 11.72 * | 759.17 ± 4.39 * | 1116.20 ± 25.56 * |
| Total SFA | 459.39 ± 2.72 | 377.67 ± 10.27 * | 403.60 ± 5.99 | 600.47 ± 11.98 * |
| Total MUFA | 245.72 ± 5.67 | 238.27 ± 5.47 | 237.63 ± 1.87 | 348.77 ± 21.07 * |
| ω-3 PUFA | 100.36 ± 3.06 | 31.73 ± 3.76 * | 40.80 ± 1.64 * | 58.13 ± 6.03 * |
| ω-6 PUFA | 114.28 ± 4.48 | 78.73 ± 1.72 * | 90.03 ± 2.19 * | 123.87 ± 2.82 |
| Total PUFA | 214.64 ± 7.52 | 110.47 ± 5.36 * | 130.83 ± 2.10 * | 182.00 ± 8.63 * |
| Total UFA | 460.35 ± 11.92 | 348.73 ± 2.91 * | 368.47 ± 0.87 * | 530.77 ± 29.34 |
| Total FA | 919.74 ± 13.86 | 764.50 ± 11.17 * | 810.67 ± 6.73 * | 1182.10 ± 27.78 * |
| Fat Acid | NC | AGPAT1 | AGPAT3 | AGPAT4 |
|---|---|---|---|---|
| C14:0 | 8.87 ± 0.75 | 33.40 ± 2.07 * | 53.83 ± 1.98 * | 33.40 ± 1.21 * |
| C15:0 | 4.20 ± 0.45 | 18.27 ± 1.36 * | 28.10 ± 2.28 * | 18.27 ± 0.82 * |
| C16:0 | 103.20 ± 8.59 | 133.90 ± 9.89 | 222.97 ± 36.02 * | 116.80 ± 7.02 |
| C17:0 | 8.47 ± 0.74 | 26.40 ± 1.54 * | 39.47 ± 3.18 * | 26.00 ± 0.95 * |
| C18:0 | 99.03 ± 11.17 | 131.63 ± 8.92 | 183.03 ± 6.68 * | 121.87 ± 10.59 |
| C20:0 | 17.60 ± 2.00 | 73.17 ± 4.65 * | 107.13 ± 9.15 * | 71.63 ± 2.93 * |
| C22:0 | 19.83 ± 2.09 | 83.40 ± 5.03 * | 122.23 ± 10.13 * | 81.33 ± 2.84 * |
| C23:0 | 10.40 ± 0.96 | 39.53 ± 2.20 * | 59.00 ± 5.26 * | 39.13 ± 1.56 * |
| C24:0 | 21.47 ± 2.34 | 90.90 ± 5.19 * | 133.40 ± 11.62 * | 88.20 ± 3.06 * |
| C16:1 | 5.77 ± 0.50 | 20.90 ± 1.27 * | 31.43 ± 2.75 * | 20.73 ± 0.92 * |
| C18:1n9c | 32.50 ± 2.76 | 28.03 ± 2.85 | 42.27 ± 9.32 | 20.30 ± 4.59 |
| C18:2n6c | 22.20 ± 2.67 | 30.53 ± 2.97 | 42.37 ± 4.09 * | 21.20 ± 1.24 |
| C18:3n3 | 8.30 ± 0.87 | 33.10 ± 1.83 * | 49.03 ± 4.37 * | 33.50 ± 1.41 * |
| C20:1 | 11.27 ± 1.09 | 32.70 ± 1.94 * | 48.20 ± 4.45 * | 31.80 ± 1.26 * |
| C20:3n6 | 15.30 ± 1.78 | 35.63 ± 2.02 * | 58.27 ± 4.74 * | 37.37 ± 2.19 * |
| C20:3n3 | 9.47 ± 1.02 | 42.43 ± 2.37 * | 63.33 ± 5.84 * | 41.57 ± 2.12 * |
| C22:1n9 | 38.57 ± 3.97 | 120.60 ± 10.13 * | 168.90 ± 18.05 * | 113.27 ± 2.50 * |
| C20:4n6 | 14.17 ± 3.02 | 34.57 ± 1.89 * | 52.93 ± 4.30 * | 37.23 ± 1.34 * |
| C22:2 | 8.83 ± 1.11 | 40.57 ± 3.14 * | 61.00 ± 6.42 * | 39.83 ± 1.56 * |
| C20:5n3 | 6.50 ± 1.18 | 24.43 ± 0.88 * | 36.17 ± 3.14 * | 23.50 ± 0.86 * |
| C24:1 | 12.07 ± 1.18 | 42.90 ± 2.65 * | 60.83 ± 5.28 * | 40.87 ± 1.39 * |
| C22:6n3 | 7.70 ± 0.90 | 29.43 ± 2.58 * | 45.47 ± 6.13 * | 31.70 ± 2.31 * |
| LC-FA | 485.70 ± 43.85 | 1146.43 ± 74.91 * | 1709.37 ± 105.19 * | 1089.50 ± 37.93 * |
| Total SFA | 293.07 ± 28.80 | 655.53 ± 41.89 * | 985.93 ± 38.59 * | 620.83 ± 26.22 * |
| Total MUFA | 100.17 ± 8.11 | 245.13 ± 17.11 * | 351.63 ± 39.59 * | 226.97 ± 4.19 * |
| ω-3 | 31.97 ± 3.71 | 129.40 ± 7.64 * | 194.00 ± 19.35 * | 130.27 ± 6.49 * |
| ω-6 | 60.50 ± 6.97 | 141.30 ± 10.01 * | 214.57 ± 19.44 * | 135.63 ± 3.91 * |
| Total PUFA | 92.47 ± 10.61 | 270.70 ± 17.50 * | 408.57 ± 38.77 * | 265.90 ± 10.19 * |
| Total UFA | 192.63 ± 18.60 | 515.83 ± 34.58 * | 760.20 ± 78.28 * | 492.87 ± 13.69 * |
| Total FA | 507.60 ± 48.42 | 1171.37 ± 76.45 * | 1748.80 ± 110.65 * | 1113.70 ± 38.72 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Z.; Li, R.; Ren, H.; Qin, C.; Su, J.; Song, X.; Wang, S.; Liu, Q.; Liu, Y.; Cui, K. Role of Different Members of the AGPAT Gene Family in Milk Fat Synthesis in Bubalus bubalis. Genes 2023, 14, 2072. https://doi.org/10.3390/genes14112072
Li Z, Li R, Ren H, Qin C, Su J, Song X, Wang S, Liu Q, Liu Y, Cui K. Role of Different Members of the AGPAT Gene Family in Milk Fat Synthesis in Bubalus bubalis. Genes. 2023; 14(11):2072. https://doi.org/10.3390/genes14112072
Chicago/Turabian StyleLi, Zhipeng, Ruijia Li, Honghe Ren, Chaobin Qin, Jie Su, Xinhui Song, Shuwan Wang, Qingyou Liu, Yang Liu, and Kuiqing Cui. 2023. "Role of Different Members of the AGPAT Gene Family in Milk Fat Synthesis in Bubalus bubalis" Genes 14, no. 11: 2072. https://doi.org/10.3390/genes14112072
APA StyleLi, Z., Li, R., Ren, H., Qin, C., Su, J., Song, X., Wang, S., Liu, Q., Liu, Y., & Cui, K. (2023). Role of Different Members of the AGPAT Gene Family in Milk Fat Synthesis in Bubalus bubalis. Genes, 14(11), 2072. https://doi.org/10.3390/genes14112072