
Integrative Taxonomy within Eremias multiocellata Complex (Sauria, Lacertidae) from the Western Part of Range: Evidence from Historical DNA


Abstract
:
1. Introduction
2. Materials and Methods
3. Results
3.1. Sequence Characteristics
3.2. Molecular Phylogeny
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Uetz, P.; Freed, P.; Aguilar, R.; Hošek, J. The Reptile Database. Available online: https://reptile-database.reptarium.cz/ (accessed on 17 April 2022).
- Szczerbak, N.N. The Palearctic Desert Lizards (Eremias); Naukova Dumka: Kiev, Ukraine, 1974. (In Russian) [Google Scholar]
- Mayer, W.; Pavlicev, M. The Phylogeny of the Family Lacertidae (Reptilia) Based on Nuclear DNA Sequences: Convergent Adaptations to Arid Habitats within the Subfamily Eremiainae. Mol. Phylogenet. Evol. 2007, 44, 1155–1163. [Google Scholar] [CrossRef] [PubMed]
- Sindaco, R.; Eremchenko, V.K. The Reptiles of the Western Palearctic. 1. Annotated Checklist and Distributional Atlas of the Turtles, Crocodiles, Amphisbaenians and Lizards of Europe, North Africa, Middle East and Central Asia. In Monografie della Societas Herpetologica Italica; Edizioni Belvedere: Latina, Italy, 2008. [Google Scholar]
- Bannikov, A.G.; Darevsky, I.S.; Ishchenko, V.G.; Rustamov, A.K.; Szczerbak, N.N. Guide to Amphibians and Reptiles of the USSR Fauna; Prosvetscheniye: Moscow, Russia, 1977. (In Russian) [Google Scholar]
- Guo, X.; Dai, X.; Chen, D.; Papenfuss, T.J.; Ananjeva, N.B.; Melnikov, D.A.; Wang, Y. Phylogeny and Divergence Times of Some Racerunner Lizards (Lacertidae: Eremias) Inferred from Mitochondrial 16S RRNA Gene Segments. Mol. Phylogenet. Evol. 2011, 61, 400–412. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Dujsebayeva, T.; Chirikova, M.; Song, Q.; Gong, X.; Guo, X. The Stummer’s Racerunner (Eremias stummeri Wettstein, 1940) Does Occur in Northwest China. Amphib. Reptil. 2021, 42, 93–105. [Google Scholar] [CrossRef]
- Liu, J.-L.; Dujsebayeva, T.N.; Chirikova, M.A.; Gong, X.; Li, D.-J.; Guo, X.-G. Does the Dzungarian Racerunner (Eremias dzungarica Orlova, Poyarkov, Chirikova, Nazarov, Munkhbaatar, Munkhbayar & Terbish, 2017) Occur in China? Species Delimitation and Identification with DNA Barcoding and Morphometric Analyses. Zool. Res. 2021, 42, 287–293. [Google Scholar] [CrossRef]
- Yakovleva, I.D. Reptiles of Kirgyzia; Academy of Sciences of Kirgyzia SSR: Frunze, Kyrgyzstan, 1964. (In Russian) [Google Scholar]
- Eremchenko, V.K.; Shukurov, E.D.; Szczerbak, N.N. A new species for the USSR fauna—Kashgar racerunner (Eremias buechneri, Sauria Reptilia). Vestn. Zool. 1985, 4, 79–80. (In Russian) [Google Scholar]
- Orlova, V.F.; Terbish, K. Materials on herpetofauna of Dzungarian Gobi. In Proceedings of the Gerpetologicheskiye Issledovaniya v; Mongolskoy Narodnoy Respublike: Moscow, Russia, 1986; pp. 91–108. (In Russian) [Google Scholar]
- Eremchenko, V.K.; Panfilov, A.M.; Tsarinenko, E.I. Synopsis of Research in Cytogenetic and Taxonomy of Some Asian Species of Scincidae and Lacertidae; Ilym: Bishkek, Kyrgyzstan, 1992. (In Russian) [Google Scholar]
- Eremchenko, V.K.; Panfilov, A.M. Taxonomic situation of multiocellated racerunner of the “multiocellata” complex of Kyrgyzstan and neighbor China (Sauria: Lacertidae: Eremias). Nauka I Novye Tekhnologii 1999, 4, 112–124. (In Russian) [Google Scholar]
- Orlova, V.F.; Poyarkov, N.A.; Chirikova, M.A.; Nazarov, R.A.; Munkhbaatar, M.; Munkhbayar, K.; Terbish, K. MtDNA Differentiation and Taxonomy of Central Asian Racerunners of Eremias multiocellata–E. przewalskii species complex (Squamata, Lacertidae). Zootaxa 2017, 1, 1–42. [Google Scholar] [CrossRef] [Green Version]
- Parham, J.F.; Stuart, B.L.; Danilov, I.G.; Ananjeva, N.B. A Genetic Characterization of CITES-Listed Iranian Tortoises (Testudo graeca) through the Sequencing of Topotypic Samples and a 19th Century Holotype. Herpetol. J. 2012, 22, 73–78. [Google Scholar]
- Hykin, S.M.; Bi, K.; McGuire, J.A. Fixing Formalin: A Method to Recover Genomic-Scale DNA Sequence Data from Formalin-Fixed Museum Specimens Using High-Throughput Sequencing. PLoS ONE 2015, 10, e0141579. [Google Scholar] [CrossRef] [Green Version]
- Mamani, L.; Cruz, R.; Mallqui, S.; Catenazzi, A. Molecular Phylogenetics and Comparative Examination of Voucher Museums Reveal Two New Species of Gymnophthalmid Lizards (Squamata, Gymnophthalmidae) from the Peruvian Andes, with Comments on Proctoporus guentheri (Boettger, 1891). Diversity 2022, 14, 215. [Google Scholar] [CrossRef]
- Mulcahy, D.G.; Ibáñez, R.; Jaramillo, C.A.; Crawford, A.J.; Ray, J.M.; Gotte, S.W.; Jacobs, J.F.; Wynn, A.H.; Gonzalez-Porter, G.P.; McDiarmid, R.W.; et al. DNA Barcoding of the National Museum of Natural History Reptile Tissue Holdings Raises Concerns about the Use of Natural History Collections and the Responsibilities of Scientists in the Molecular Age. PLoS ONE 2022, 17, e0264930. [Google Scholar] [CrossRef] [PubMed]
- Lebedev, V.S.; Kovalskaya, Y.; Solovyeva, E.N.; Zemlemerova, E.D.; Bannikova, A.A.; Rusin, M.Y.; Matrosova, V.A. Molecular Systematics of the Sicista tianschanica Species Complex: A Contribution from Historical DNA Analysis. PeerJ 2021, 9, e10759. [Google Scholar] [CrossRef] [PubMed]
- de Abreu, E.F., Jr.; Pavan, S.E.; Tsuchiya, M.T.N.; Wilson, D.E.; Percequillo, A.R.; Maldonado, J.E. Museomics of Tree Squirrels: A Dense Taxon Sampling of Mitogenomes Reveals Hidden Diversity, Phenotypic Convergence, and the Need of a Taxonomic Overhaul. BMC Evol. Biol. 2020, 20, 77. [Google Scholar] [CrossRef] [PubMed]
- Rancilhac, L.; Bruy, T.; Scherz, M.D.; Pereira, E.A.; Preick, M.; Straube, N.; Lyra, M.L.; Ohler, A.; Streicher, J.W.; Andreone, F.; et al. Target-Enriched DNA Sequencing from Historical Type Material Enables a Partial Revision of the Madagascar Giant Stream Frogs (Genus Mantidactylus). J. Nat. Hist. 2020, 54, 87–118. [Google Scholar] [CrossRef]
- Reyes-Velasco, J.; Goutte, S.; Freilich, X.; Boissinot, S. Mitogenomics of Historical Type Specimens Clarifies the Taxonomy of Ethiopian Ptychadena Boulenger, 1917 (Anura, Ptychadenidae). ZooKeys 2021, 1070, 135–149. [Google Scholar] [CrossRef]
- Solovyeva, E.N.; Lebedev, V.S.; Dunayev, E.A.; Nazarov, R.A.; Bannikova, A.A.; Che, J.; Murphy, R.W.; Poyarkov, N.A. Cenozoic Aridization in Central Eurasia Shaped Diversification of Toad-Headed Agamas (Phrynocephalus; Agamidae, Reptilia). PeerJ 2018, 6, e4543. [Google Scholar] [CrossRef] [Green Version]
- Lebedev, V.S.; Bannikova, A.A.; Lu, L.; Snytnikov, E.A.; Adiya, Y.; Solovyeva, E.N.; Abramov, A.V.; Surov, A.V.; Shenbrot, G.I. Phylogeographical Study Reveals High Genetic Diversity in a Widespread Desert Rodent, Dipus Sagitta (Dipodidae: Rodentia). Biol. J. Linn. Soc. 2018, 123, 445–462. [Google Scholar] [CrossRef]
- Bannikova, A.A.; Jenkins, P.D.; Solovyeva, E.N.; Pavlova, S.V.; Demidova, T.B.; Simanovsky, S.A.; Sheftel, B.I.; Lebedev, V.S.; Fang, Y.; Dalen, L.; et al. Who Are You, Griselda? A Replacement Name for a New Genus of the Asiatic Short-Tailed Shrews (Mammalia, Eulipotyphla, Soricidae): Molecular and Morphological Analyses with the Discussion of Tribal Affinities. ZooKeys 2019, 888, 133–158. [Google Scholar] [CrossRef]
- Schileyko, A.A.; Solovyeva, E.N. On the Taxonomic Position of the Enigmatic Genus Tonkinodentus Schileyko, 1992 (Chilopoda, Scolopendromorpha): The First Molecular Data. ZooKeys 2019, 840, 133–155. [Google Scholar] [CrossRef] [PubMed]
- GenBank. Available online: https://www.ncbi.nlm.nih.gov/genbank/ (accessed on 13 May 2022).
- Hall, T. BioEdit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Burland, T.G. DNASTAR’s Lasergene Sequence Analysis Software. Methods Mol. Biol. 2000, 132, 71–91. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanfear, R.; Calcott, B.; Ho, S.Y.W.; Guindon, S. Partitionfinder: Combined Selection of Partitioning Schemes and Substitution Models for Phylogenetic Analyses. Mol. Biol. Evol. 2012, 29, 1695–1701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian Phylogenetic Inference under Mixed Models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior Summarization in Bayesian Phylogenetics Using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [Green Version]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian Inference of Phylogenetic Trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.-T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Minh, B.Q.; Nguyen, M.A.T.; von Haeseler, A. Ultrafast Approximation for Phylogenetic Bootstrap. Mol. Biol. Evol. 2013, 30, 1188–1195. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.; Wong, T.K.F. ModelFinder: Fast Model Selection for Accurate Phylogenetic Estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [Green Version]
- Swofford, D. PAUP*. Phylogenetic Analysis Using Parsimony (*and Other Methods); Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Müller, K. PRAP–Computation of Bremer Support for Large Data Sets. Mol. Phylogenet. Evol. 2004, 31, 780–782. [Google Scholar] [CrossRef]
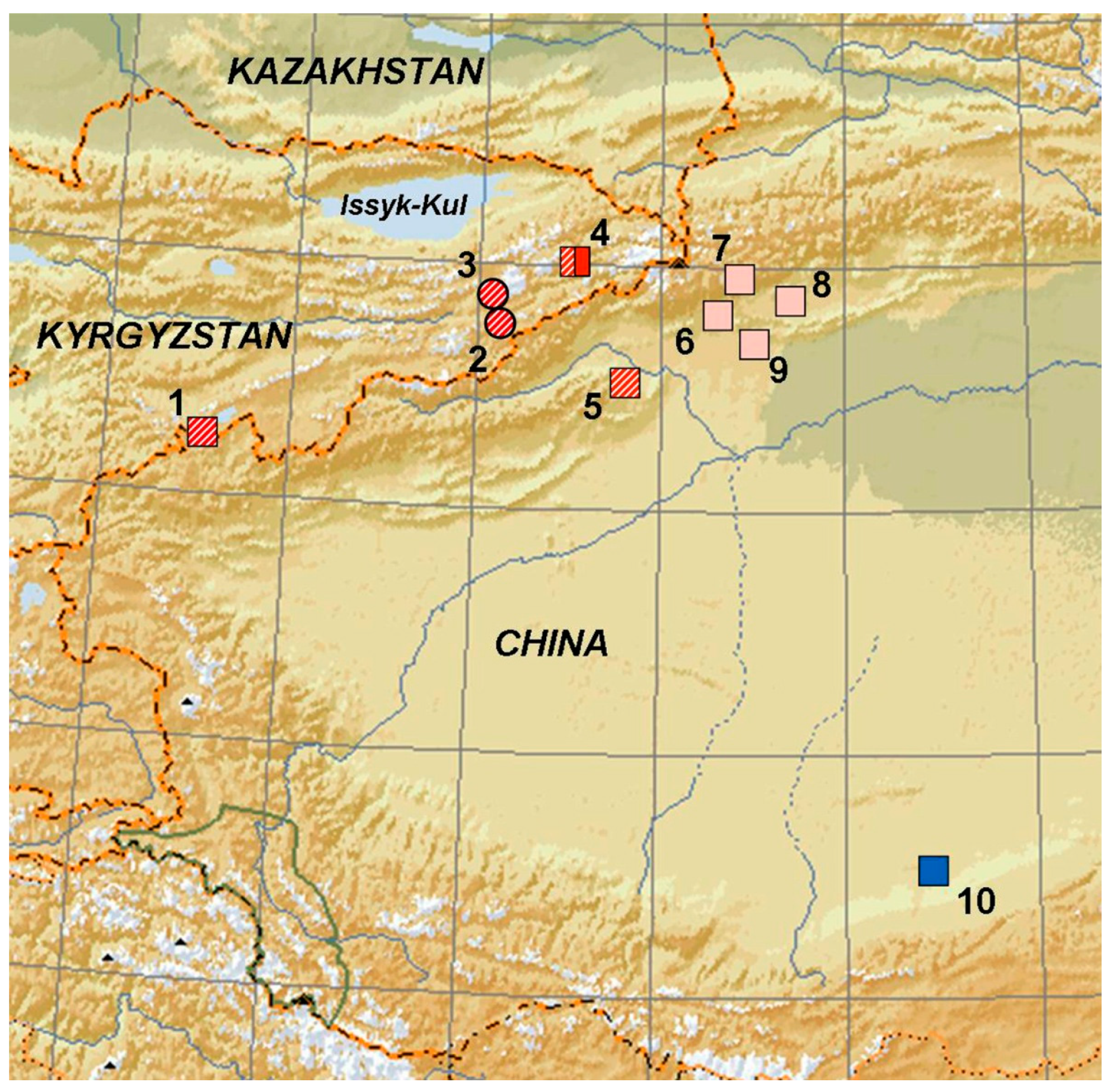
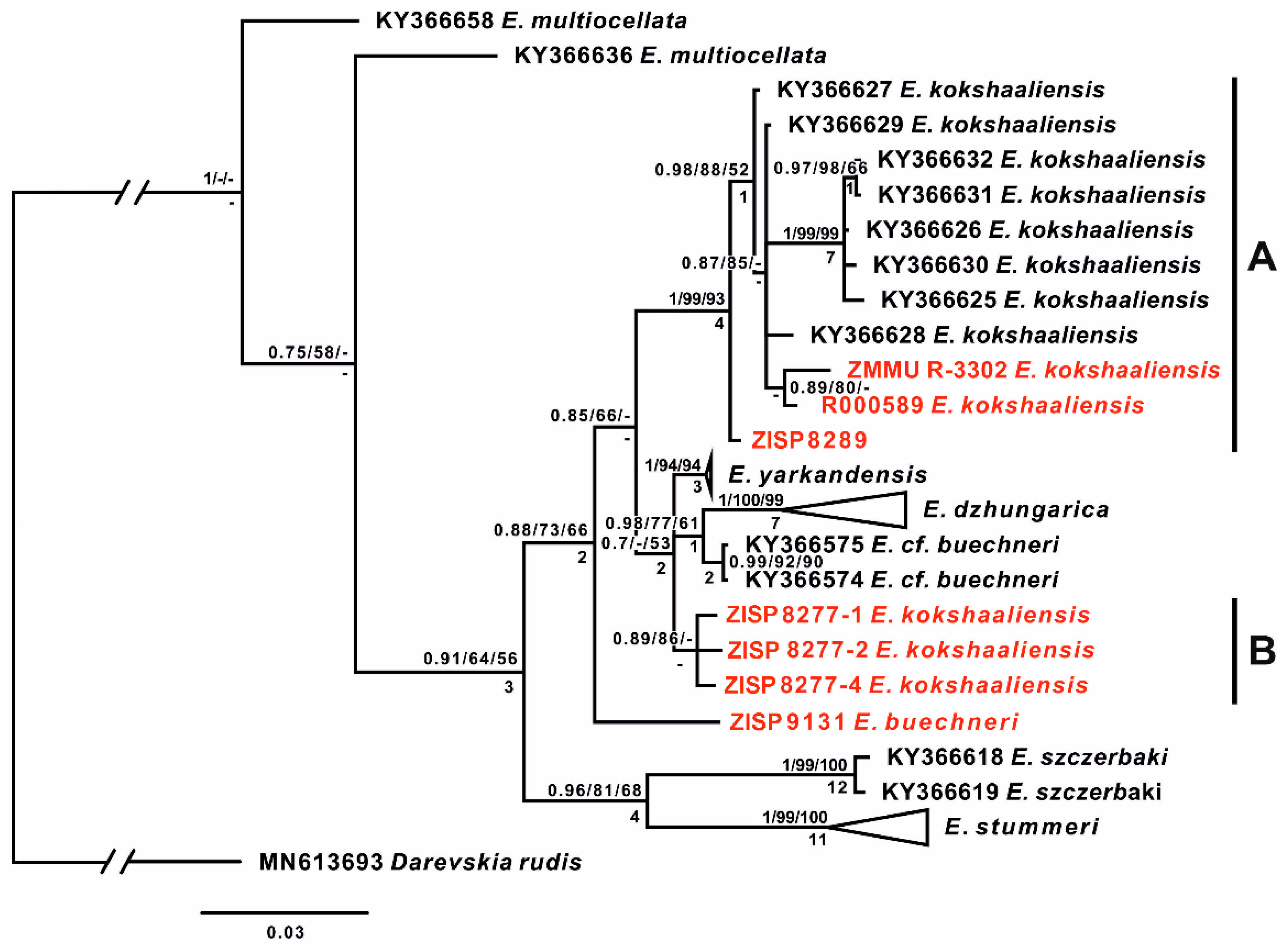




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genbank Assession Number | Species | Specimen Voucher Number | Country | Locality | Coordinates |
|---|---|---|---|---|---|
| Sequences obtained in this study | |||||
| OK624438 | Eremias kokshaaliensis | BSI R000589 | Paratype, Kyrgyzstan | Sary-Dhaz, vicinity of Engylchek (Enylchek) | - |
| OK624437 | Eremias kokshaaliensis | ZMMU R-3302 | Kyrgyzstan | Kyrgyzstan, central Tien Shan, Sary-Dhaz River at the confluence with the Inylchek River | - |
| OK624441 | Eremias kokshaaliensis | ZISP 8277-1 | Paratype, China | China, Xinjiang, Kara-Teke, | - |
| OK624442 | Eremias kokshaaliensis | ZISP 8277-2 | Paratype, China | China, Xinjiang, Kara-Teke, | - |
| OK624443 | Eremias kokshaaliensis | ZISP 8277-4 | Paratype, China | China, Xinjiang, Kara-Teke, | - |
| OK624439 | Eremias kokshaaliensis | ZISP 8289 | Paratype | China, Xinjiang, Taushkan –Darya | - |
| OK624440 | Eremias buechneri | ZISP 9131 | China | Kashgaria | - |
| Sequences from (Orlova et al., 2017, Figure 1, Appendix II) | |||||
| KY366618 | Eremias szczerbaki | ZMMU R-14342-1 | Kyrgyzstan | Naryn Prov., Naryn Distr., N from Naryn | 41.48 N; 75.98 E |
| KY366619 | Eremias szczerbaki | ZMMU R-14342-2 | Kyrgyzstan | Naryn Prov., Naryn Distr., N from Naryn | 41.48 N; 75.98 E |
| KY366589 | Eremias stummeri | ZMMU R-12551-1a | Kazakhstan | Almaty Prov., Rayimbek Distr., Ketmen Mts. foothills, 7–8 km E from Kegen | 42.97 N; 79.32 E |
| KY366590 | Eremias stummeri | ZMMU R-12551-2 | Kazakhstan | Almaty Prov., Rayimbek Distr., Ketmen Mts. foothills, 7–8 km E from Kegen | 42.97 N; 79.32 E |
| KY366591 | Eremias stummeri | ZMMU R-12551-2a | Kazakhstan | Almaty Prov., Rayimbek Distr., Ketmen Mts. foothills, 7–8 km E from Kegen | 42.97 N; 79.32 E |
| KY366592 | Eremias stummeri | ZMMU R-12551-3 | Kazakhstan | Almaty Prov., Rayimbek Distr., Ketmen Mts. foothills, 7–8 km E from Kegen | 42.97 N; 79.32 E |
| KY366593 | Eremias stummeri | ZMMU R-12551-4 | Kazakhstan | Almaty Prov., Rayimbek Distr., Ketmen Mts. foothills, 7–8 km E from Kegen | 42.97 N; 79.32 E |
| KY366594 | Eremias stummeri | ZMMU R-12551-5 | Kazakhstan | Almaty Prov., Rayimbek Distr., Ketmen Mts. foothills, 7–8 km E from Kegen | 42.97 N; 79.32 E |
| KY366601 | Eremias stummeri | ZMMU R-12552-1 | Kazakhstan | Almaty Prov., Rayimbek Distr., central Tian Shan Mts., 15 km S from Tuzkol Lake, Zhabyrtau Mt. | 42.92 N; 80.08 E |
| KY366604 | Eremias stummeri | ZMMU R-14335-2 | Kyrgyzstan | Issyk-Kul Prov., E bank of Issyk-Kul lake, env. of Karakol | 42.46 N; 78.36 E |
| KY366607 | Eremias stummeri | ZMMU R-14338-1 | Kyrgyzstan | Issyk-Kul Prov., 100 km SW from Karakol, Kaji-Say env. | 42.16 N; 77.18 E |
| KY366609 | Eremias stummeri | ZMMU R-12427-1 | Kyrgyzstan | Issyk-Kul Prov., NW bank of Issyk-Kul lake, 20–25 km N from Toru Aygyr, Kungei-Alatau Mts. | 42.58 N; 76.41 E |
| KY366610 | Eremias stummeri | ZMMU R-12427-2 | Kyrgyzstan | Issyk-Kul Prov., NW bank of Issyk-Kul lake, 20–25 km N from Toru Aygyr, Kungei-Alatau Mts. | 42.58 N; 76.41 E |
| KY366612 | Eremias stummeri | ZMMU R-12556-2 | Kyrgyzstan | Issyk-Kul Prov., env. of Balykchy, road to Akolen | 42.35 N; 76.17 E |
| KY366613 | Eremias stummeri | ZMMU R-12557-1 | Kyrgyzstan | Issyk-Kul Prov., env. of Balykchy, road to Akolen | 42.35 N; 76.17 E |
| KY366615 | Eremias stummeri | ZMMU R-14339-1 | Kyrgyzstan | Naryn Prov., Kochkor Distr., Kochkor | 42.22 N; 75.75 E |
| KY366617 | Eremias stummeri | ZMMU R-14341-3 | Kyrgyzstan | Naryn Prov., Kochkor Distr., S from Kochkor | 42.08 N; 75.66 E |
| KY366574 | Eremias cf. buechneri | ZMMU R-8910-1a | China | Xinjiang Prov., Qarqan (Chemo) Distr., Altintag Mt., Chinbulak, 60 km S from Tura | 37.51 N; 86.05 E |
| KY366575 | Eremias cf. buechneri | ZMMU R-8910-1b | China | Xinjiang Prov., Qarqan (Chemo) Distr., Altintag Mt., Chinbulak, 60 km S from Tura | 37.51 N; 86.05 E |
| KY366576 | Eremias dzungarica | ZMMU R-11989-1 | Kazakhstan | east Kazakhstan Prov., Aigyrkum sands, 5–7 km SW from Buran | 47.98 N; 84.89 E |
| KY366577 | Eremias dzungarica | ZMMU R-11989-2 | Kazakhstan | east Kazakhstan Prov., Aigyrkum sands, 5–7 km SW from Buran | 47.98 N; 84.89 E |
| KY366578 | Eremias dzungarica | ZMMU R-12862-1 | Mongolia | Khovd Aimaq, Bulgan Sum, Bayan-Mod, 11 km W from Ikher-Toli | 47.06 N; 92.91 E |
| KY366579 | Eremias dzungarica | ZMMU R-12862-3 | Mongolia | Khovd Aimaq, Bulgan Sum, Bayan-Mod, 11 km W from Ikher-Toli | 47.06 N; 92.91 E |
| KY366580 | Eremias dzungarica | ZMMU R-12845-2 | Mongolia | Khovd Aimaq, 7 km SW from Uyench Sum | 46.93 N; 93.61 E |
| KY366581 | Eremias dzungarica | ZMMU R-12845-4 | Mongolia | Khovd Aimaq, 7 km SW from Uyench Sum | 46.93 N; 93.61 E |
| KY366582 | Eremias dzungarica | ZMMU R-12845-5 | Mongolia | Khovd Aimaq, 7 km SW from Uyench Sum | 46.93 N; 93.61 E |
| KY366583 | Eremias dzungarica | ZMMU R-12845-6 | Mongolia | Khovd Aimaq, 7 km SW from Uyench Sum | 46.93 N; 93.61 E |
| KY366584 | Eremias dzungarica | ZMMU R-12845-7 | Mongolia | Khovd Aimaq, 7 km SW from Uyench Sum | 46.93 N; 93.61 E |
| KY366585 | Eremias dzungarica | ZMMU R-12845-8 | Mongolia | Khovd Aimaq, 7 km SW from Uyench Sum | 46.93 N; 93.61 E |
| KY366586 | Eremias dzungarica | ZMMU R-12845-9 | Mongolia | Khovd Aimaq, 7 km SW from Uyench Sum | 46.93 N; 93.61 E |
| KY366587 | Eremias dzungarica | ZMMU R-12845-10 | Mongolia | Khovd Aimaq, 7 km SW from Uyench Sum | 46.93 N; 93.61 E |
| KY366588 | Eremias dzungarica | ZMMU R-12550-1 | Mongolia | Khovd Aimaq, 24 km N from Uyench Sum | 46.27 N; 92.05 E |
| KY366620 | Eremias yarkandensis | ZMMU R-14344-1 | Kyrgyzstan | Osh Prov., vicinity of Nura | 39.65 N; 73.87 E |
| KY366621 | Eremias yarkandensis | ZMMU R-14344-2 | Kyrgyzstan | Osh Prov., vicinity of Nura | 39.65 N; 73.87 E |
| KY366622 | Eremias yarkandensis | ZMMU R-14344-3 | Kyrgyzstan | Osh Prov., vicinity of Nura | 39.65 N; 73.87 E |
| KY366623 | Eremias yarkandensis | ZMMU R-14344-4 | Kyrgyzstan | Osh Prov., vicinity of Nura | 39.65 N; 73.87 E |
| KY366624 | Eremias yarkandensis | ZMMU R-14344-5 | Kyrgyzstan | Osh Prov., vicinity of Nura | 39.65 N; 73.87 E |
| KY366625 | Eremias sp. | ZMMU R-14327-1 | China | Xinjiang Prov., 35 km NE from Aksu | 41.40 N; 81.05 E |
| KY366626 | Eremias sp. | ZMMU R-14329-1 | China | Xinjiang Prov., 60 km NE from Aksu | 41.54 N; 80.79 E |
| KY366627 | Eremias sp. | ZMMU R-14330-1 | China | Xinjiang Prov., 75 km NE from Aksu | 41.74 N; 80.83 E |
| KY366628 | Eremias sp. | ZMMU R-14330-2 | China | Xinjiang Prov., 75 km NE from Aksu | 41.74 N; 80.83 E |
| KY366629 | Eremias sp. | ZMMU R-14330-3 | China | Xinjiang Prov., 75 km NE from Aksu | 41.74 N; 80.83 E |
| KY366630 | Eremias sp. | ZMMU R-14328-1 | China | Xinjiang Prov., 89 km NE from Aksu | 41.56 N; 81.21 E |
| KY366631 | Eremias sp. | ZMMU R-14328-2 | China | Xinjiang Prov., 89 km NE from Aksu | 41.56 N; 81.21 E |
| KY366632 | Eremias sp. | ZMMU R-14328-3 | China | Xinjiang Prov., 89 km NE from Aksu | 41.56 N; 81.21 E |
| Outgroups | |||||
| KY366658 | Eremias multiocellata | ZMMU R-12843-1 | Mongolia | Govi-Altai Aimaq, Shargyn-Govi, 2 km SW from Khaliun Sum | 45.93 N; 96.12 E |
| KY366636 | Eremias multiocellata | ZMMU R-13215 | China | Inner Mongolia Prov., 50 km S from Baotou | 40.28 N; 110.00 E |
| MN613693 | Darevskia rudis | DRbi86 | Turkey | Cankurtaran Pass, Hopa, Artvin | 41.390452 N; 41.541104 E |
| Primer Name | Sequence of 3′–5′ | Annealing Temperature |
|---|---|---|
| Er.k-1aF | ACCAACCACAAAGAYATYGGCACTTT | 58 |
| Er.k-1aR | ATGAAGGCGTGGGCTGTTACGACTAC | |
| Er.k-2F | CCMGGCACCCTCCTAGGAGAYGAY | 53 |
| Er.k-2R | CGCCAGCTTCAACTGCTGATGATG | |
| Er.k-3F | GATATGGCATTYCCACGGATAAATAA | 53 |
| Er.k-3R | ACAAGTGGTGATAAAGTTRATTGCYCCTAAG | |
| Er.k-4F | CACGCAGGRGCATCTGTRGACCTAACTAT | 63 |
| Er.k-4R | AAGAGYATARTGATGCCGGCTGCTAGGACA | |
| Er.k-5F | CCCCCAAACATAACKCARTACCAAACC | 56 |
| Er.k-5R | ACTTCAGGGTGACCAAARAATCARAATAG |
| E. buechneri | E. cf. buechneri | E. dzhungarica | E. kokshaaliensis Group A | E. kokshaaliensis Group B | E. stummeri | E. szczerbaki | E. yarkandensis | E. yarkandensis | |
|---|---|---|---|---|---|---|---|---|---|
| E. buechneri | - | 1.22 | 1.30 | 1.21 | 1.00 | 1.51 | 1.65 | 1.21 | 1.30 |
| E. cf. buechneri | 5.09 | 0 | 0.59 | 0.75 | 0.46 | 1.07 | 1.12 | 0.48 | 0.48 |
| E. dzhungarica | 6.54 | 2.63 | 0.45 | 0.81 | 0.70 | 1.14 | 1.18 | 0.68 | 0.73 |
| E. kokshaaliensis group A | 5.70 | 4.36 | 5.45 | 1.41 | 0.74 | 1.09 | 1.10 | 0.69 | 0.77 |
| E. kokshaaliensis group B | 3.93 | 1.51 | 3.75 | 4.22 | 0.84 | 1.06 | 1.11 | 0.50 | 0.53 |
| E. stummeri | 8.93 | 7.85 | 9.37 | 8.91 | 7.53 | 1.12 | 1.07 | 1.06 | 1.27 |
| E. szczerbaki | 9.67 | 8.43 | 9.15 | 8.53 | 8.12 | 7.82 | 0.32 | 1.11 | 1.31 |
| E. yarkandensis | 4.67 | 1.46 | 3.45 | 3.71 | 1.65 | 7.78 | 8.10 | 0 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Orlova, V.F.; Solovyeva, E.N.; Dunayev, E.A.; Ananjeva, N.B. Integrative Taxonomy within Eremias multiocellata Complex (Sauria, Lacertidae) from the Western Part of Range: Evidence from Historical DNA. Genes 2022, 13, 941. https://doi.org/10.3390/genes13060941
Orlova VF, Solovyeva EN, Dunayev EA, Ananjeva NB. Integrative Taxonomy within Eremias multiocellata Complex (Sauria, Lacertidae) from the Western Part of Range: Evidence from Historical DNA. Genes. 2022; 13(6):941. https://doi.org/10.3390/genes13060941
Chicago/Turabian StyleOrlova, Valentina F., Evgeniya N. Solovyeva, Evgenyi A. Dunayev, and Natalia B. Ananjeva. 2022. "Integrative Taxonomy within Eremias multiocellata Complex (Sauria, Lacertidae) from the Western Part of Range: Evidence from Historical DNA" Genes 13, no. 6: 941. https://doi.org/10.3390/genes13060941
APA StyleOrlova, V. F., Solovyeva, E. N., Dunayev, E. A., & Ananjeva, N. B. (2022). Integrative Taxonomy within Eremias multiocellata Complex (Sauria, Lacertidae) from the Western Part of Range: Evidence from Historical DNA. Genes, 13(6), 941. https://doi.org/10.3390/genes13060941