Abstract
The 52 members of the Teosinte-Branched 1/Cycloidea/Proliferating Cell Factors (TCP) Transcription Factor gene family in Malus × domestica (M. × domestica) were identified in 2014 on the first genome assembly, which was released in 2010. In 2017, a higher quality genome assembly for apple was released and is now considered to be the reference genome. Moreover, as in several other species, the identified TCP genes were named based on the relative position of the genes on the chromosomes. The present work consists of an update of the TCP gene family based on the latest genome assembly of M. × domestica. Compared to the previous classification, the number of TCP genes decreased from 52 to 40 as a result of the addition of three sequences and the deduction of 15. An analysis of the intragenic identity led to the identification of 15 pairs of orthologs, shedding light on the forces that shaped the evolution of this gene family. Furthermore, a revised nomenclature system is proposed that is based both on the intragenic identity and the homology with Arabidopsis thaliana (A. thaliana) TCPs in an effort to set a common standard for the TCP classification that will facilitate any future interspecific analysis.
1. Introduction
The first complete genome of a plant, the model organism Arabidopsis thaliana (A. thaliana), was released at the very beginning of the sequencing era [1], and only ten years later, the milestone of the tenth plant genome being sequenced was achieved. The recent huge advances in genome sequencing technologies and the drastic decrease in sequencing costs have led to the generation of enormous amounts of primary and derived genomic data, with over a thousand plant genomes published in the past ten years (https://www.plabipd.de/index.ep, accessed 15 March 2022).
The availability and ease of access to vast amounts of genomic sequences have enabled the proliferation of works aiming to identify gene families in a broad range of plant species, including members of the Teosinte-Branched 1/Cycloidea/Proliferating Cell Factors (TCP) gene family. TCPs consist of a group of genes found in all the land plants, from mosses to eudicots, that encode transcription factors acting as key regulators of a large number of biological processes related to several aspects of plant growth and development and, possibly, in biotic and abiotic stresses responses [2,3,4,5,6,7].
TCP proteins are characterized by the presence of a non-canonical basic helix–loop–helix (bHLH) motif known as the TCP domain, which mediates nuclear localization, DNA binding and protein–protein interactions [8]. Based on conserved differences in the TCP domain sequence that reflect functional differences, TCPs can be divided in Class I and II, the latter being further subdivided into Cincinnata (CIN) and Cycloidea/Teosinte Branched 1 (CYC/TB1) subclasses [3].
The evolution of the TCP gene family was shaped by several duplication events and a rapid expansion from lower to higher plants [2,9,10]. Consequently, a relatively small number of TCP genes have been found in the basal groups of land plants (e.g., seven in the moss Physcomitrella patens and six in the lycophyte Selaginella moellendorffi [11]), while higher plants exhibit the presence of a much larger number (e.g., 26 in Oryza sativa, 24 in A. thaliana, 46 in Zea mays) [9,12,13,14].
The identification of the TCP gene family in Malus × domestica (M. × domestica) dates to 2014, when Xu and collaborators [15] revealed 52 TCPs (MdTCPs) on the first apple genome assembly released in 2010 [16] (http://www.rosaceae.org; accessed 10 January 2022). The 52 genes were numbered from 1 to 52 based on the relative position on chromosomes and subdivided as follows: 22 in Class I, 26 in Class II-CIN and four in Class II–CYC/TB1. Additionally, the authors described 12 pairs of paralogs, of which seven were linked to potential chromosomal duplications associated with a genome-wide duplication, and two more pairs were tightly collocated in the apple genome. Therefore, it was proposed that the evolution of the TCP gene family in M. × domestica was shaped by both segmental duplication and transposition events [15].
In 2017, a new assembly of M. × domestica genome, GDDH13v1.1, was released [17] and is now considered to be the reference genome for apple. In fact, despite having the lowest number of predicted genes among all the previous M. × domestica assemblies, GDDH13v1.1 is characterized by an overall higher quality, making it the most complete among apple genome sequences published to date [18].
This study aimed to perform a novel identification of MdTCP genes based on the high-quality genome assembly GDDH13v1.1, which is followed by an analysis of the gene sequences to identify genes that originated from genome duplication events. Moreover, an AtTCP-homology-based nomenclature system for the MdTCP genes is proposed.
2. Materials and Methods
2.1. Revision and De Novo Identification of the MdTCP Gene Family Set
Each of the 52 deduced amino acid MdTCP sequences identified by Xu et al. [15] on the 2010 assembly was used as a query for a tblastn search on the RefSeq database (http://www.ncbi.nlm.nih.gov/refseq/; accessed 10 January 2022) [19]. Additional M. × domestica TCP-containing putative protein sequences were obtained on Pfam (https://pfam.xfam.org/; accessed 10 January 2022) [20] and PlantTFDB (http://planttfdb.gao-lab.org/; accessed 10 January 2022) [21] databases. Alignments were performed on MEGA X software [22] (Mega X, version 10.2.5) with the Clustal Omega Multiple Alignment Tool [23] applying the following parameters: a gap-opening penalty of 5.00 and a gap extension penalty of 0.1 for the pairwise alignments; a gap-opening penalty of 10.00 and a gap extension penalty of 0.05 for multiple alignments; BLOSUM substitution weight matrix and a delayed divergent cutoff of 30% (sequences diverging by more than 30% were aligned later). All alignments were manually inspected and edited, if necessary. Selected sequences were subsequently used as queries to identify the gene positions on the GDDH13v1.1 assembly [17] (IRHS database; https://iris.angers.inra.fr/gddh13/; accessed 19 January 2022). The presence of an Open Reading Frame (ORF) and a TCP domain was determined on InterPro (http://www.ebi.ac.uk/Tools/pfa/iprscan/; accessed 25 January 2022) and Pfam databases. The GDDH13v1.1 Genome assembly and corresponding mRNA prediction list were accessed through https://www.rosaceae.org/analysis/242 (accessed 19 January 2022) and visualized using Blast and JBrowse functions.
2.2. Identification of Potential Sister Genes among the MdTCP Gene Family
Reciprocal identities among MdTCP and AtTCP amino acid sequences were calculated on BioEdit software [24] on the alignment of amino acid sequences. The cutoffs for high, moderate and low-identity MdTCPs were set based on the Percentage of Identity (PID) values observed in AtTCPs.
To correlate the identity values with the pattern of chromosome duplication of apple, a Circos Plot [25] was generated on Galaxy servers (https://usegalaxy.eu/; accessed 7 February 2022) [26]. The Karyotype file of the GDDH13v1.1 assembly was downloaded from the IRHS database, while the BED file containing the relative chromosome positions of the putative paralogous MdTCPs was produced manually.
2.3. A Novel MdTCPs Nomenclature System Based on the Homology with AtTCPs
The A. thaliana homolog for each of the putative MdTCP was inferred based on DNA sequence similarity. A BLAST search restricted to A. thaliana sequences was performed using each deduced MdTCP amino acid sequence as query with the following cutoffs: query cover > 75% and Evalue < 1^ (−50). The same parameters were used to perform a Blast search restricted to M. × domestica sequences using AtTCP amino acid sequences as queries.
The MdTCP neighbor-joining [27] phylogenetic tree was inferred on amino acid sequences with 1000 bootstrap reiterations. Evolutionary distances were computed using the Poisson correction method [28] in units of number of amino acid substitutions per site. The rate variation among sites was modeled with a γ distribution with a γ parameter of 1.00 and a homogeneous pattern, with the partial deletion option (i.e., fewer than 40% alignment gaps, missing data, and ambiguous bases were allowed at any position). Analyses were conducted with MEGA X software.
To calculate the maximum likelihood tree on A. thaliana and M. × domestica TCPs, the amino acid sequences of 40 MdTCPs and 24 AtTCPs were aligned on MEGA X as described above, with subsequent manual revision. The Smart Model Selection algorithm [29] with the Akaike Information Criterion (AIC) was applied to calculate the appropriate evolution model. The maximum likelihood phylogenetic tree was inferred in PhyML 3.0 software [30] (Guindon; accessed from Bolzano/Bozen, Italy) using the Jones–Taylor–Thornton substitution model, discrete γ distribution with three parameters, proportion of invariable site, and empirical amino acid frequencies count (JTT + G + I + F). Initial trees for the heuristic search were obtained by automatically applying the Neighbor-Join and BioNJ algorithms to a matrix of pairwise distances estimated using the JTT model and finally selecting the topology with the superior log-likelihood value.
3. Results
3.1. Revision and De Novo Identification of the MdTCP Gene Family Set
All the putative TCP sequences of M. × domestica found on public databases were aligned with the 52 MdTCP sequences published by Xu and colleagues [15]. The alignment showed that sequences retrieved from these databases correspond to the published MdTCP gene set, constitute fragments of TCPs, or they do not contain a TCP domain at all. The database search did not reveal the presence of other TCP-containing sequences besides those already mentioned in the MdTCP set [15], which were then employed for the subsequent analyses.
The manual evaluation of the 52 MdTCP sequences highlighted that four pairs (MdTCP5/MdTCP6, MdTCP13/MdTCP14, MdTCP19/MdTCP20, MdTCP43/MdTCP44) and one trio (MdTCP8/MdTCP9/MdTCP10) exhibit a reciprocal 100% identity. The accessions of these 11 sequences were aligned to the M. × domestica double haploid genome GDDH13v1.1 (IRHS database; https://iris.angers.inra.fr/gddh13/; accessed 19 January 2022) [17], and the members within each pair or trio mapped to single positions on the genome. Six of these redundant sequences, one for each pair and two for the trio, were thus excluded from the gene set.
The alignment of the 46 remaining MdTCP sequences on the GDDH13v1.1 assembly resulted in 33 sequences matching with predicted protein-coding genes and 13 either mapping onto intergenic regions or partially overlapped non-TCP predicted gene regions. The corresponding GDDH13v1.1 nucleotide sequences of the 13 non-TCP mapped genes were therefore retrieved and verified for the presence of the TCP domain: seven deduced amino acid sequences (MdTCP3, MdTCP4, MdTCP19, MdTCP36, MdTCP37, MdTCP41, MdTCP50) display premature stop codons, and two more (MdTCP7, MdTCP42) constitute fragments of non-TCP gene sequences. The nine sequences were thus removed from the set. On the contrary, the remaining four non-TCP mapping sequences (MdTCP2, MdTCP17, MdTCP23, MdTCP52) contain a full-open reading frame (ORF) and a TCP domain and were thus kept in the set.
Overall, 15 out of the 52 starting sequences were excluded from the set during the revision process: six due to redundancy, seven due to the presence of premature stop codons and two for constituting fragments of non-TCP genes. Table 1 summarizes the details regarding excluded and confirmed sequences, including previous and updated accessions.

Table 1.
MdTCP genes identified by Xu et al. [15] and relative accession numbers determined on the 2010 genome assembly [16] and in the present work on the most recent genome assembly GDDH13v1.1 (IRHS database; https://iris.angers.inra.fr/gddh13/; accessed 19 January 2022) [17]. From this original set, six sequences were excluded due to redundancy with other sequences, seven due to the presence of premature stop codons and two for constituting fragments of non-TCP genes. A new GDDH13v1.1 accession was successfully determined for 33 MdTCPs, while for four additional sequences, an accession was not found.
Thirty out of the 37 remaining sequences were found to be listed as TCP-containing in the mRNA gene list predicted from GDDH13v1.1. Additionally, three novel putative TCP genes that were not reported in the original set by Xu et al. [15] were identified in this predicted mRNA set. Three sequences (MdTCP33, MdTCP40, MdTCP45) previously observed to match with protein-coding genes on the GDDH13v1.1 assembly appeared to be automatically annotated as uncharacterized proteins.
The three novel sequences include a start and a stop codon as well as a TCP domain. The sequences were provisionally named “nc” (“not classified”) 1, 2 and 3 (MdTCPnc1, MdTCPnc2 and MdTCPnc3) and added to the set. The alignment of the deduced amino acid sequences of the three novel MdTCPs, displayed in Figure 1, shows that these three sequences are similar. In particular, MdTCPnc3 and MdTCPnc2 align, respectively, with the 3′ (positions 140–393) and 5′ end (1–265) of MdTCPnc1, which is the longest of the three. Consequently, the two shorter sequences MdTCPnc3 and MdTCPnc2 share an almost 100% conserved region of 124 amino acids, corresponding to the central region of MdTCPnc1. Interestingly, the two shorter sequences map on physically close regions of the same chromosome.

Figure 1.
Alignment of the deduced amino acid sequences of the three novel putative MdTCPs, provisionally named MdTCPnc1/2/3. Numbers indicate the alignment positions, including gaps. The alignment was performed and visualized on MEGA X software. A colored background (MEGA X default color code) indicates identical positions in at least two sequences.
Each TCP-mRNA predicted from the GDDH13v1.1 database was used as query for a BLAST search against the A. thaliana gene database “The Arabidopsis Information Resource” (TAIR; www.arabidopsis.org/aboutarabidopsis; accessed 22 February 2022) to retrieve all the GDDH13v1.1-predicted mRNA sequences displaying similarity to AtTCP genes. Out of a total of 24 AtTCP genes, only 14 were best hits for the 33 predicted MdTCP mRNAs. No additional mRNA sequence from the entire set of all mRNAs predicted from GDDH13v1.1 was found to be displaying similarity with any AtTCP, further confirming the absence of additional non-identified TCP-containing sequences in the GDDH13v1.1 genome.
3.2. Identification of Potential Sister Genes among the MdTCP Gene Family
Potential sister MdTCPs were inferred based on the reciprocal identity values, namely the proportion of invariant sites for each pair of deduced amino acid sequences. Since recently duplicated MdTCPs are expected to show a higher percentage of identity (PID) than those that are non-recently duplicated, the AtTCPs PID was calculated to determine the threshold to define two genes as potential sister genes. AtTCPs exhibiting a notable PID have been identified based on the PID cutoff beyond which sequence alignment algorithms are able to distinguish between protein pairs of similar and non-similar structure (i.e., 35%) [31]. Among eight couples with a PID higher than 35%, the highest observed value was 62% and the mean was 47%. Percentages of 47% and 62% were thus selected as thresholds to define MdTCP sequences with moderate and high PID values, respectively. The identity matrix calculated on the deduced amino acid sequences of the 40 MdTCPs (Supplementary Table S2) is graphically represented in Figure 2. Results show that 14 pairs of MdTCPs share high PID (red squares), and one pair shows a moderate degree (light red). One trio (MdTCPnc1, MdTCPnc2, MdTCPnc3) exhibits a moderate PID: two sequences do not show a PID higher than the 47% threshold, but both show a PID higher than 47% with the third sequence, MdTCPnc1. Four sequences (MdTCP2, MdTCP17, MdTCP23 and MdTCP52) appear to constitute a group with a moderate-high PID (mean 62%), making the relationships between each of the sequences in this cluster challenging to disentangle. Finally, three sequences do not display a moderate nor high PID with any other MdTCP. These results suggest that overall, at least 15 pairs of MdTCPs can be considered paralogous genes originated through the whole genome duplication event, although hints of a recent duplication appear in the three-member group, MdTCPnc1, MdTCPnc2 and MdTCPnc3, as well as in the four-member group (MdTCP2, MdTCP17, MdTCP23 and MdTCP52).
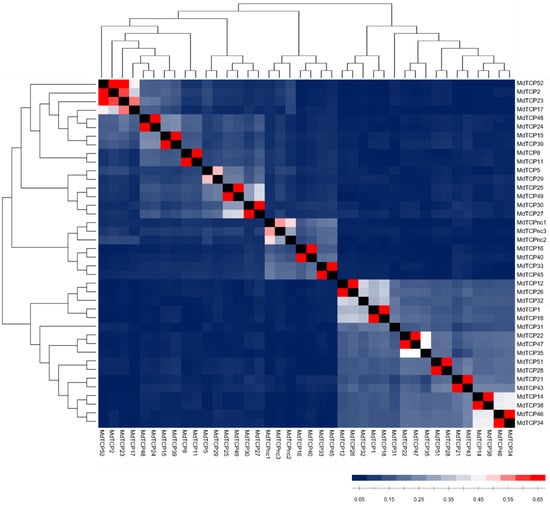
Figure 2.
Graphical representation of the identity matrix calculated on the amino acid sequences of the 40 MdTCPs identified. MdTCP identifiers are reported to the right and bottom of each row/column. The phylogenetic relationships among genes are displayed by a neighbor-joining tree on top and to the left. The color gradient of each square indicates a low (blue), moderate (white), or high (red) percentage of identity between two MdTCPs.
The comparison between potential paralogous MdTCP genes and the apple intragenomic synteny is displayed as an overlapped Circos plot in Figure 3. Two MdTCP genes (MdTCP2 and MdTCP18) map on the pseudochromosome 0, meaning that they were detected on contigs not assembled on any of the 17 apple chromosomes. Thus, the links involving these genes could not be represented. Figure 3 shows that the hypothesis of duplication of 15 pairs of genes is consistent with the results of the synteny analysis. Regarding the group of three, the two sequences MdTCPnc2 and MdTCPnc3 are located on the same chromosome (9), and both links with MdTCPnc1 (indicated with red lines) are consistent with the duplication pattern (chromosomes 9–17). None of the links between members of the group of four genes (dotted lines) appear to be supported. Despite the similarity observed between the four sequences, the ambiguity of these results suggests that the four genes MdTCP2, MdTCP17, MdTCP23 and MdTCP52 cannot be considered as sister genes.
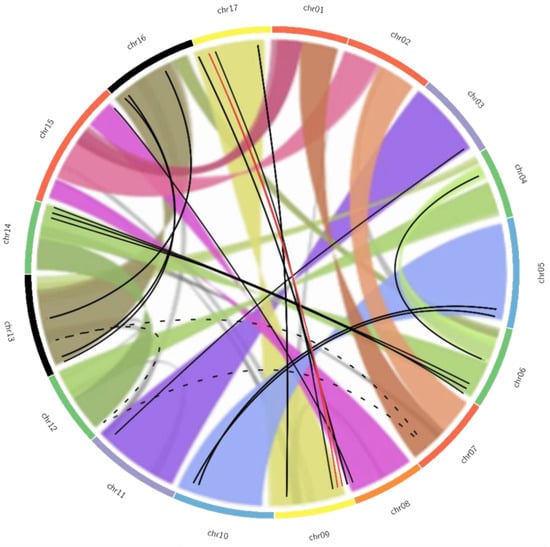
Figure 3.
Graphical representation of the duplication pattern observed in MdTCP genes overlapped to pattern of chromosome duplication observed in the Apple genome. The pattern of apple genome duplication, as determined by Daccord and colleagues [17], is indicated as colored ribbons connecting homologous regions between the chromosomes, as represented by the colored slim boxes on the outside of the circle. Similar chromosomes are indicated with the same color to help visualization. Lines connect the physical positions on the chromosomes of the potentially duplicated MdTCPs in accordance with the previous analysis: the black solid lines link the couples, the red lines link the members of the group of three, while the black dotted lines connect members of the group of four. Adapted from Daccord et al., 2019 [17], under CC BY 4.0 license.
3.3. A Novel MdTCPs Nomenclature System Based on the Homology with AtTCPs
The A. thaliana homolog for each sequence was estimated through a BLAST query of each nucleotide sequence over the A. thaliana gene database to determine the best hits for each gene, which was followed by manual investigation and confirmation. MdTCP genes were thus re-named, appending the letter “a” after the number to differentiate between the current and the proposed nomenclatures. The letter “b” is used to indicate paralogs, which is in accordance with the previous analyses. In one case, three MdTCP sequences (MdTCP12, MdTCP26, and MdTCP32) showed similarity to the same A. thaliana sequence (AtTCP9), although only two of them exhibit a high percentage of reciprocal identity. To simultaneously highlight both the correspondence with the same AtTCP and the peculiar intragenic relationships, the two sister genes were named MdTCP9a/b and the third was named MdTCP9-like. Similarly, the cluster of four genes MdTCP2, MdTCP17, MdTCP23, and MdTCP52, whose evolutionary relationships are not exhaustively explained by whole-genome duplication origin, are similar to one A. thaliana gene only (AtTCP17): the four genes were therefore named MdTCP17-like_a/b/c/d. A unique AtTCP (AtTCP1) appears to be the best hit for the three genes MdTCPnc1, MdTCPnc2, MdTCPnc3, for which the genome duplication hypothesis partially explains the relationship between a gene and the other two. Thus, the three genes were named MdTCP1a/b/c.
In total, 19 AtTCP sequences were identified as homologs for the complete set of 40 MdTCPs. A M. × domestica ortholog was not found for the remaining five AtTCPs, namely AtTCP11, AtTCP16, AtTCP22, AtTCP23 and AtTCP24.
The complete revised set of MdTCP genes, listed in Table 2, comprises 40 unique sequences (full-length CDS in Supplementary Material, File S3), of which:
Table 2.
Details of the 40 MdTCP sequences classified in the present work. The table lists the name of the genes determined on the homology with AtTCPs and the corresponding accession number and gene name as described by Xu and co-authors [15]. See Supplementary Materials (Table S1) for full details.
- 33 were deduced from both 2010 and GDDH13v1.1 assemblies.
- Four were deduced from the 2010 assembly but not annotated on GDDH13v1.1.
- Three were deduced from GDDH13v1.1 but absent on the 2010 assembly and, thus, not been predicted before.
The separation of MdTCPs into the two classes and the subclasses thereof is visible in the neighbor-joining phylogenetic tree displayed in Figure 4, panel A: 17 genes belong to TCP Class I (22 in the previous classification by Xu and co-authors [15]), 16 belong to the subclass CIN of Class II (26 in the previous classification) and seven belong to the Class II subclass CYC/TB1 (four in the previous classification). In A. thaliana, Class I contains 13 genes, Class II-CIN contains eight, and Class II-TB1/CYC contains three.
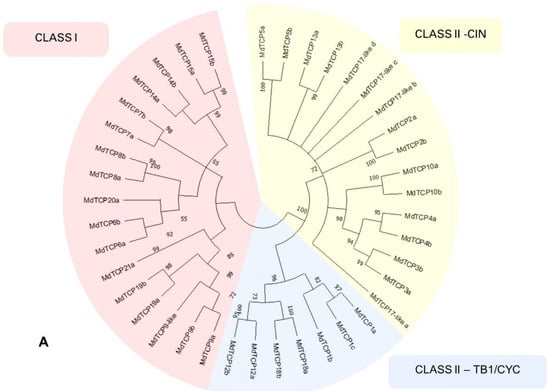
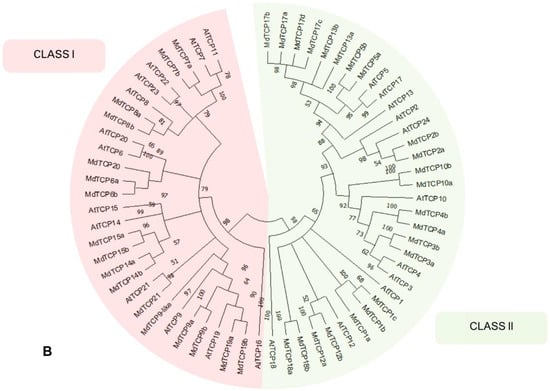
Figure 4.
(A) Phylogenetic tree calculated on the 40 MdTCP genes classified in the present work. The different classes of TCPs are indicated with red (Class I), yellow (Class II-CIN) and blue (Class II–TB1/CYC). The tree was inferred using the neighbor-joining method [27] with 10,000 bootstrap re-iterations. Evolutionary distances were computed using the Poisson correction method [28] and are in the units of the number of amino acid substitutions per site. The rate variation among sites was modeled with a γ distribution. All positions with less than 60% site coverage were eliminated; i.e., fewer than 40% alignment gaps, missing data, and ambiguous bases were allowed at any position (partial deletion option). Node support is indicated as bootstrap values, and nodes with a support lower than 50 are collapsed. Analyses were conducted in MEGA X [22]. (B) Phylogenetic tree calculated from the amino acid sequences of the 40 MdTCP genes identified in the present work, and 24 AtTCPs. Members of the Class I are indicated with a red background, while those in Class II are indicated with a green background. Smart Model Selection algorithm [29] with Akaike Information Criterion (AIC) was applied to calculate the appropriate evolution model. The phylogenetic tree was inferred with the Jones–Taylor–Thornton substitution model, discrete γ distribution with three parameters, proportion of invariable site, and empirical amino acid frequencies count (JTT + G + I + F). Initial trees for the heuristic search were obtained by automatically applying the neighbor-join and BioNJ algorithms to a matrix of pairwise distances estimated using the JTT model and finally selecting the topology with the superior log-likelihood value (−37420,02). Nodes with a support lower than 50 are condensed. Analyses were conducted on PhyML 3.0 software [30] (Guindon; accessed from Bolzano/Bozen, Italy).
In the maximum likelihood phylogenetic tree in panel B of Figure 4, including both MdTCP and AtTCP sequences, a general consistency between the MdTCP-AtTCP homologies previously determined can be observed. Even though the phylogenetic tree is unrooted, by implicitly rooting the tree at the split between Class I and Class II TCPs, it is possible to infer the relationships among genes, even though we advise the readers to refer to specific studies on phylogenetics and the evolution of TCP genes ([9,32]). While the Class I and the Class II-CIN subclass appear to be monophyletic, the phylogenetic relationships are not well defined in the subclass CYC/TB1. AtTCP18 does not group with other members of Class II, and, in general, the node support of the CYC/TB1 taxon is low; thus, many nodes are represented as polytomies. This indicates that the intrinsic ambiguity in the alignment of the CYC/TB1 subclass sequences does not allow a confident reconstruction of the phylogenetic relationships in this group.
4. Discussion
In the present work, the MdTCP gene family has been identified from the latest M. × domestica genome assembly GDDH13v1.1. Compared to the classification based on the previous draft genome sequence of 2010, the number of genes decreased from 52 to 40 as a result of the addition of three novel sequences and the exclusion of 15. The motivation for excluding these sequences was because nine did not exhibit a TCP-domain-containing ORF and six showed redundancy with other sequences. Interestingly, all the reciprocally identical sequences were characterized by sequential names, which reflect physical proximity on the chromosomes, since the previous nomenclature was based on the relative position of each gene on the chromosomes. Consequently, the duplications that led to the identification of redundant MdTCP sequences in the 2010 assembly are absent in GDDH13v1.1, and were likely to be artefacts generated during the in silico genome assembly. On the other hand, the analysis of the MdTCP percentage identity values and the comparison with the chromosome duplication pattern within the M. × domestica genome suggest that the origin of 15 pairs, and probably one more pair, is associated to chromosome duplication. As expected, the definition of the evolutionary relationship in the cluster of four genes is elusive and is further complicated by the fact that one member, MdTCP2, is mapped on the pseudochromosome 0. These findings suggest that the chromosomal or segmental duplication had a significant role in the generation of the TCP set in M. × domestica, while transposition events likely had a marginal impact.
Furthermore, in the present work, 19 AtTCPs were identified as homologs of the 40 MdTCPs genes. For comparison, in the classification carried out on the previous draft of the M. × domestica genome assembly, only 15 AtTCPs resulted as homologs for the complete set of MdTCP genes. Nonetheless, despite the higher number of AtTCP homolog genes found in the present work, no correspondent MdTCP was found for any of the five remaining AtTCPs. Interestingly, two paralogs have been found for each Class II AtTCP except AtTCP24, while several members of Class I appear to have only one or no ortholog in M. × domestica. In addition to highlighting the significant role of genome-wide duplication in the generation of the MdTCP gene family, these results suggest that further members of the MdTCP gene family, and especially Class I members, may have been subjected to a process of gene loss or have still to be identified.
The TCP nomenclature is usually based on a numbering system associated with the homology of the sequences with the corresponding A. thaliana TCP, but more often, TCPs (as well as other gene families) are named after the relative order of the sequences on the chromosomes. The adoption of two different standards to classify newly discovered TCP sequences is prone to ambiguities where identical TCP-containing sequences are indicated with two or even three different numbers. Such a condition constitutes a major obstacle for research: more and more genes are being characterized at a functional level in physiological pathways, interaction partners or expression patterns; however, the transferability of data among different species is severely hindered. An effort should be made to establish a common standard for the nomenclature of TCPs, also considering the exponential growth of data generation. In the present study, the identified TCP genes have been renamed after the homology with the AtTCP counterparts. In our view, such classification will facilitate future research by reducing ambiguities associated with a random numbering of TCPs in different species.
In the future, the availability of genomes of even higher assembly quality or pangenomes, transcriptome analyses and the integration of functional data will probably provide further details for a better understanding of the evolution and the function of the members of this important transcription factor family.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/genes13101696/s1, Table S1: Details of the MdTCP sequences identified in the present work, including exon number, coding sequences lengths and details of the deduced amino acid sequences; Table S2: raw PID values calculated on the 40 MdTCP deduced amino acid sequences. File S3: coding sequences of the 40 MdTCPs identified in the present work.
Author Contributions
Conceptualization, M.T., M.M. and K.J.; methodology, M.T.; validation, M.T.; formal analysis, M.T.; investigation, M.T.; resources, M.T., M.M. and K.J.; data curation, M.T.; writing—original draft preparation, M.T.; writing—review and editing, M.T., M.M. and K.J.; visualization, M.T.; supervision, M.M. and K.J.; project administration, M.M. and K.J.; funding acquisition, M.M. and K.J. All authors have read and agreed to the published version of the manuscript.
Funding
The work was partially performed as part of the projects APPLIII and APPLIV within the framework agreement in the field of invasive species in fruit growing and major pathologies, which is co-funded by the South Tyrolean Apple Consortium and the Autonomous Province of Bozen/Bolzano, Italy. The authors thank the Department of Innovation, Research, University and Museums of the Autonomous Province of Bozen/Bolzano for covering the Open Access publication costs.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All the sequences used and analyzed in the present work can be found on online repositories or databases: RefSeq (http://www.ncbi.nlm.nih.gov/refseq/; accessed 10 January 2022), Pfam (https://pfam.xfam.org/; accessed 10 January 2022), PlantTFDB (http://planttfdb.gao-lab.org/family.php?sp=Mdo&fam=TCP; accessed 10 January 2022), TAIR, (https://www.arabidopsis.org/browse/genefamily/TCP.jsp; accessed 10 February 2022), GDDH13v1.1 (https://iris.angers.inra.fr/gddh13/; accessed 10 February 2022).
Acknowledgments
The authors thank Diego Micheletti for the bioinformatic inputs during data analysis and curation and Cameron Brodrick Cullinan for language proofreading of the article.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- The Arabidopsis Genome Initiative. Analysis of the genome sequence of the flowering plant Arabidopsis thaliana. Nature 2000, 408, 796–815. [Google Scholar] [CrossRef] [PubMed]
- Martín-Trillo, M.; Cubas, P. TCP genes: A family snapshot ten years later. Trends Plant Sci. 2010, 15, 31–39. [Google Scholar] [CrossRef]
- Nicolas, M.; Cubas, P. TCP factors: New kids on the signaling block. Curr. Opin. Plant Biol. 2016, 33, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Mukhtar, M.S.; Carvunis, A.-R.; Dreze, M.; Epple, P.; Steinbrenner, J.; Moore, J.; Tasan, M.; Galli, M.; Hao, T.; Nishimura, M.T.; et al. Independently evolved virulence effectors converge onto hubs in a plant immune system network. Science 2011, 333, 596–601. [Google Scholar] [CrossRef]
- Sugio, A.; Kingdom, H.N.; MacLean, A.M.; Grieve, V.M.; Hogenhout, S.A. Phytoplasma protein effector SAP11 enhances insect vector reproduction by manipulating plant development and defense hormone biosynthesis. Proc. Natl. Acad. Sci. USA 2011, 108, E1254–E1263. [Google Scholar] [CrossRef]
- Janik, K.; Mithöfer, A.; Raffeiner, M.; Stellmach, H.; Hause, B.; Schlink, K. An effector of apple proliferation phytoplasma targets TCP transcription factors-a generalized virulence strategy of phytoplasma? Mol. Plant Pathol. 2017, 18, 435–442. [Google Scholar] [CrossRef]
- Wang, S.; Shen, Y.; Guo, L.; Tan, L.; Ye, X.; Yang, Y.; Zhao, X.; Nie, Y.; Deng, D.; Liu, S.; et al. Innovation and Emerging Roles of Populus trichocarpa Teosinte Branched1/Cycloidea/Proliferating Cell Factor Transcription Factors in Abiotic Stresses by Whole-Genome Duplication. Front. Plant Sci. 2022, 13, 850064. [Google Scholar] [CrossRef]
- Kosugi, S.; Ohashi, Y. DNA binding and dimerization specificity and potential targets for the TCP protein family. Plant J. 2002, 30, 337–348. [Google Scholar] [CrossRef] [PubMed]
- Navaud, O.; Dabos, P.; Carnus, E.; Tremousaygue, D.; Hervé, C. TCP transcription factors predate the emergence of land plants. J. Mol. Evol. 2007, 65, 23–33. [Google Scholar] [CrossRef]
- Liu, M.-M.; Wang, M.-M.; Yang, J.; Wen, J.; Guo, P.-C.; Wu, Y.-W.; Ke, Y.-Z.; Li, P.-F.; Li, J.-N.; Du, H. Evolutionary and Comparative Expression Analyses of TCP Transcription Factor Gene Family in Land Plants. Int. J. Mol. Sci. 2019, 20, 3591. [Google Scholar] [CrossRef]
- Ortiz-Ramírez, C.; Hernandez-Coronado, M.; Thamm, A.; Catarino, B.; Wang, M.; Dolan, L.; Feijó, J.A.; Becker, J.D. A Transcriptome Atlas of Physcomitrella patens Provides Insights into the Evolution and Development of Land Plants. Mol. Plant 2016, 9, 205–220. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Ma, H.; Wang, J.; Zhang, D. Genome-Wide Comparative Analysis and Expression Pattern of TCP Gene Families in Arabidopsis thaliana and Oryza sativa. J. Integr. Plant Biol. 2007, 49, 885–897. [Google Scholar] [CrossRef]
- Riechmann, J.L.; Heard, J.; Martin, G.; Reuber, L.; Jiang, C.; Keddie, J.; Adam, L.; Pineda, O.; Ratcliffe, O.J.; Samaha, R.R.; et al. Arabidopsis transcription factors: Genome-wide comparative analysis among eukaryotes. Science 2000, 290, 2105–2110. [Google Scholar] [CrossRef] [PubMed]
- Ding, S.; Cai, Z.; Du, H.; Wang, H. Genome-Wide Analysis of TCP Family Genes in Zea mays L. Identified a Role for ZmTCP42 in Drought Tolerance. Int. J. Mol. Sci. 2019, 20, 2762. [Google Scholar] [CrossRef] [PubMed]
- Xu, R.; Sun, P.; Jia, F.; Lu, L.; Li, Y.; Zhang, S.; Huang, J. Genomewide analysis of TCP transcription factor gene family in Malus domestica. J. Genet. 2014, 93, 733–746. [Google Scholar] [CrossRef]
- Velasco, R.; Zharkikh, A.; Affourtit, J.; Dhingra, A.; Cestaro, A.; Kalyanaraman, A.; Fontana, P.; Bhatnagar, S.K.; Troggio, M.; Pruss, D.; et al. The genome of the domesticated apple (Malus × domestica Borkh.). Nat. Genet. 2010, 42, 833–839. [Google Scholar] [CrossRef]
- Daccord, N.; Celton, J.-M.; Linsmith, G.; Becker, C.; Choisne, N.; Schijlen, E.; van de Geest, H.; Bianco, L.; Micheletti, D.; Velasco, R.; et al. High-quality de novo assembly of the apple genome and methylome dynamics of early fruit development. Nat. Genet. 2017, 49, 1099–1106. [Google Scholar] [CrossRef]
- Peace, C.P.; Bianco, L.; Troggio, M.; van de Weg, E.; Howard, N.P.; Cornille, A.; Durel, C.-E.; Myles, S.; Migicovsky, Z.; Schaffer, R.J.; et al. Apple whole genome sequences: Recent advances and new prospects. Hortic. Res. 2019, 6, 59. [Google Scholar] [CrossRef]
- O’Leary, N.A.; Wright, M.W.; Brister, J.R.; Ciufo, S.; Haddad, D.; McVeigh, R.; Rajput, B.; Robbertse, B.; Smith-White, B.; Ako-Adjei, D.; et al. Reference sequence (RefSeq) database at NCBI: Current status, taxonomic expansion, and functional annotation. Nucleic Acids Res. 2016, 44, D733–D745. [Google Scholar] [CrossRef]
- Mistry, J.; Chuguransky, S.; Williams, L.; Qureshi, M.; Salazar, G.A.; Sonnhammer, E.L.L.; Tosatto, S.C.E.; Paladin, L.; Raj, S.; Richardson, L.J.; et al. Pfam: The protein families database in 2021. Nucleic Acids Res. 2021, 49, D412–D419. [Google Scholar] [CrossRef]
- Jin, J.; Tian, F.; Yang, D.-C.; Meng, Y.-Q.; Kong, L.; Luo, J.; Gao, G. PlantTFDB 4.0: Toward a central hub for transcription factors and regulatory interactions in plants. Nucleic Acids Res. 2017, 45, D1040–D1045. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Sievers, F.; Higgins, D.G. The Clustal Omega Multiple Alignment Package. Methods Mol. Biol. 2021, 2231, 3–16. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids. Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Krzywinski, M.; Schein, J.; Birol, I.; Connors, J.; Gascoyne, R.; Horsman, D.; Jones, S.J.; Marra, M.A. Circos: An information aesthetic for comparative genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar] [CrossRef] [PubMed]
- Afgan, E.; Baker, D.; Batut, B.; van den Beek, M.; Bouvier, D.; Cech, M.; Chilton, J.; Clements, D.; Coraor, N.; Grüning, B.A.; et al. The Galaxy platform for accessible, reproducible and collaborative biomedical analyses: 2018 update. Nucleic Acids Res. 2018, 46, W537–W544. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef] [PubMed]
- Zuckerkandl, E.; Pauling, L. Molecules as documents of evolutionary history. J. Theor. Biol. 1965, 8, 357–366. [Google Scholar] [CrossRef]
- Lefort, V.; Longueville, J.-E.; Gascuel, O. SMS: Smart Model Selection in PhyML. Mol. Biol. Evol. 2017, 34, 2422–2424. [Google Scholar] [CrossRef] [PubMed]
- Guindon, S.; Dufayard, J.-F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef]
- Rost, B. Twilight zone of protein sequence alignments. Protein Eng. 1999, 12, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Mondragón-Palomino, M.; Trontin, C. High time for a roll call: Gene duplication and phylogenetic relationships of TCP-like genes in monocots. Ann. Bot. 2011, 107, 1533–1544. [Google Scholar] [CrossRef] [PubMed]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).