
Genome-Wide Identification and Characterization of HSP70 Gene Family in Aquilaria sinensis (Lour.) Gilg



Abstract
:
1. Introduction
2. Materials and Methods
2.1. Identification of HSP70s in Plant Species
2.2. Phylogenetic Analyses
2.3. Gene Structure Analysis, Conserved Motif Analysis and Chromosomal Location Analysis
2.4. Plant Materials and Stress Treatments
2.5. Subcellular Localization in Arabidopsis Protoplasts
2.6. RNA Isolation and Real-Time qRT-PCR
3. Results
3.1. Identification of the HSP70 Gene Family in A. sinensis
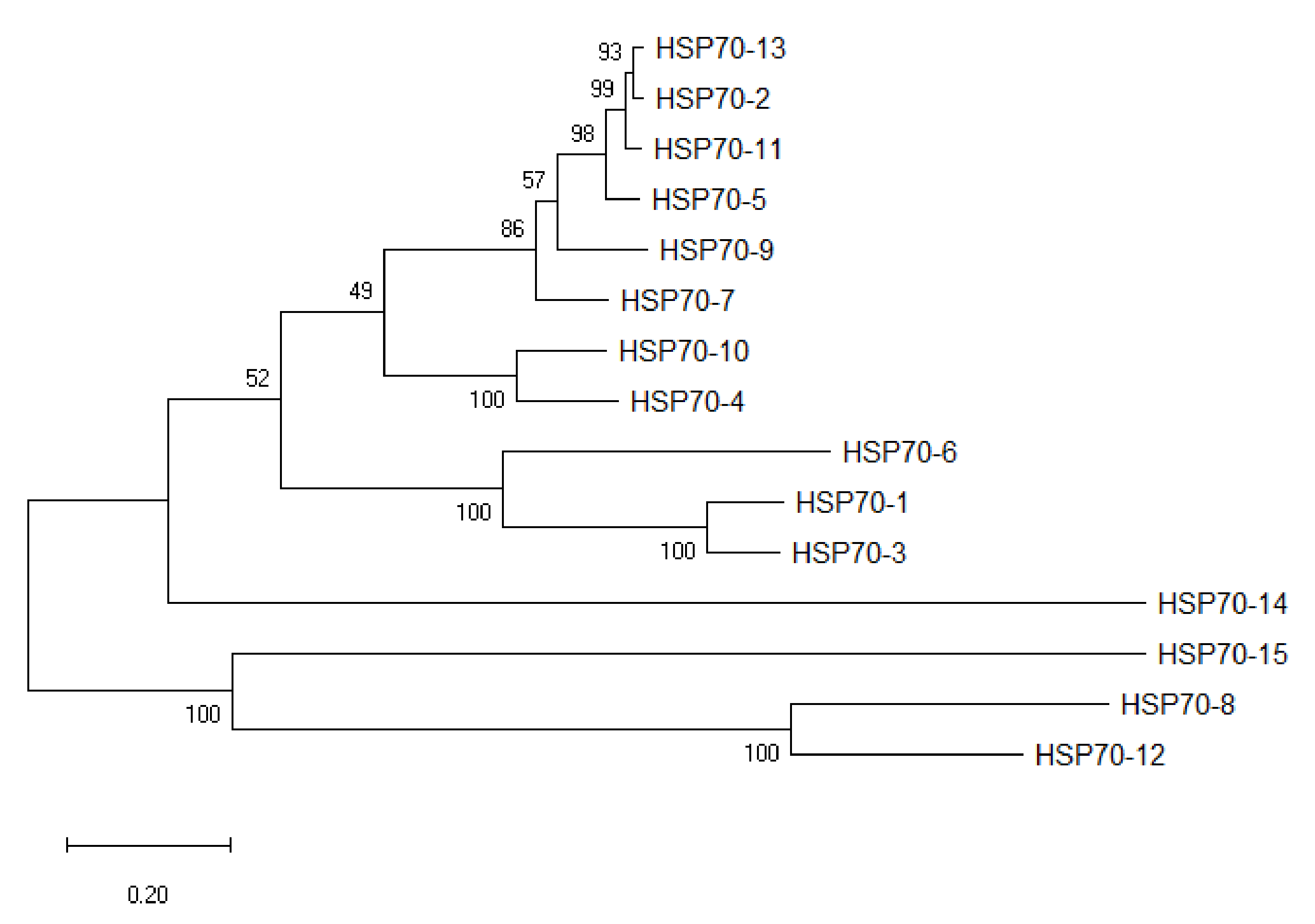
3.2. Phylogenetic Relationship of the HSP70 Genes in A. sinensis
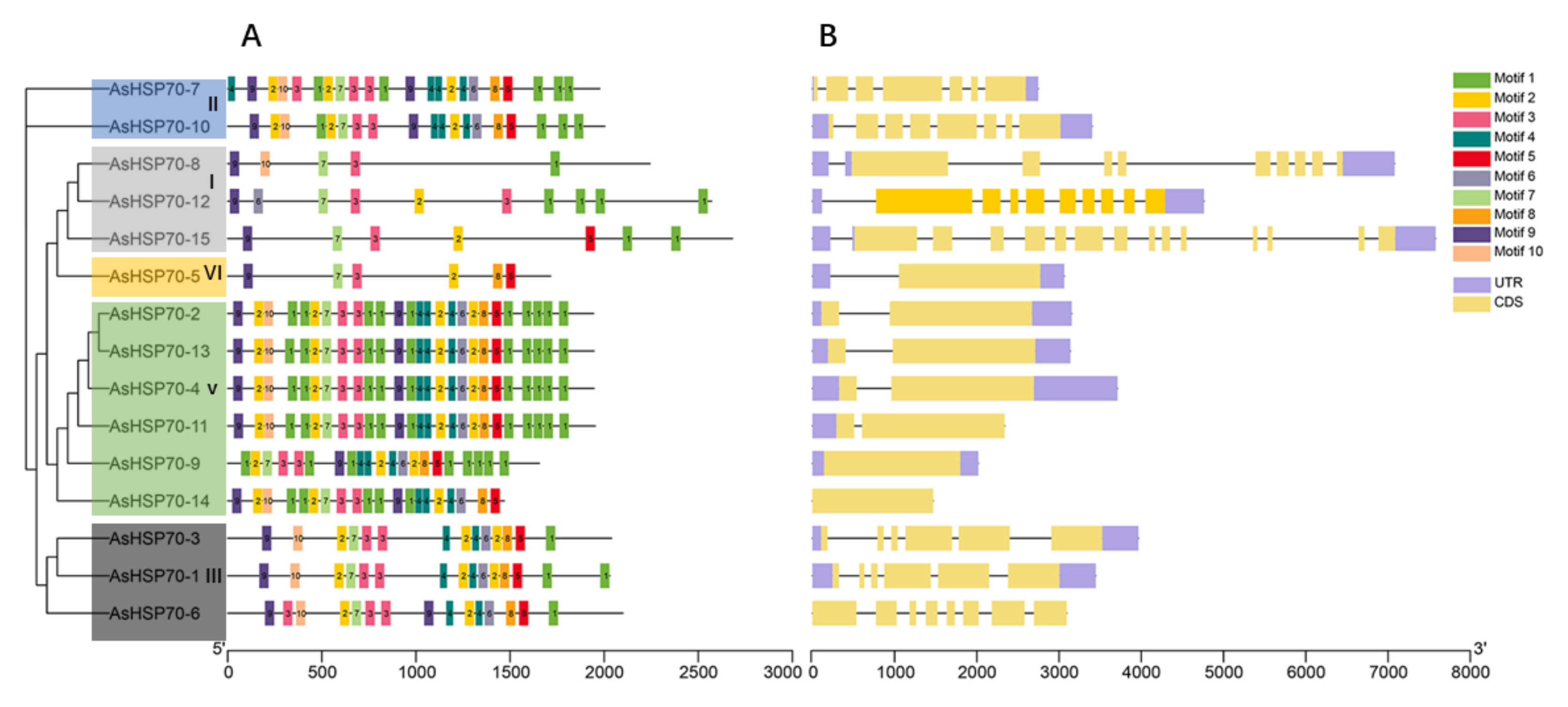
3.3. Conserved Motif and Gene Structure Analyses of HSP70 Genes in A. sinensis
3.4. Expression Profiles of AsHSP70 Genes in Response to Heat Stress
3.5. Subcellular Localization of AsHSP70s
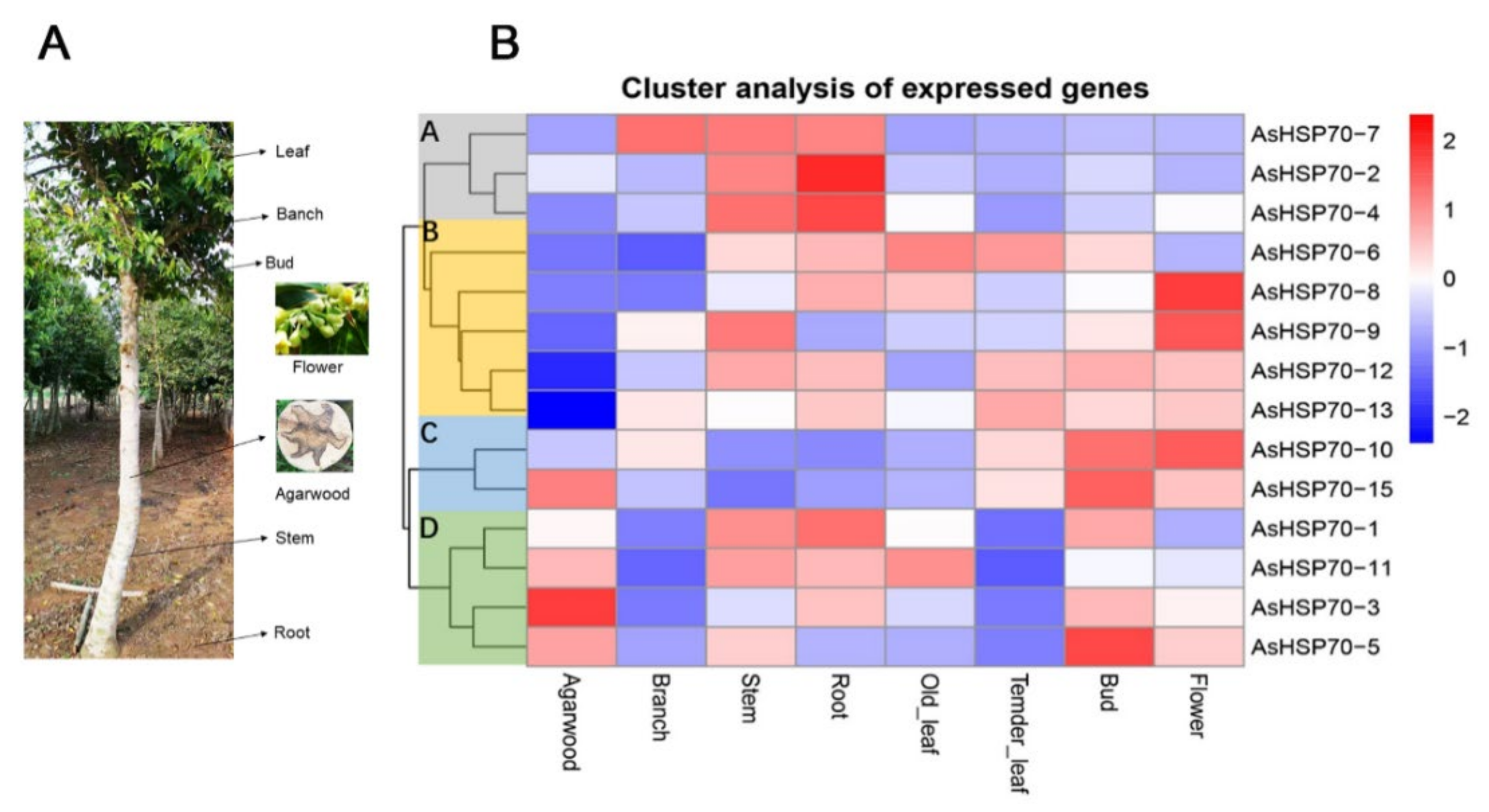
3.6. Expression Profiles of AsHSP70 in Different Tissues and Different Layers during the Whole-Tree-Inducing Agarwood Formation Process in A. sinensis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, W.-X.; Vinocur, B.; Altman, A. Plant responses to drought, salinity and extreme temperatures: Towards genetic engineering for stress tolerance. Planta 2003, 218, 1–14. [Google Scholar] [CrossRef]
- Wang, W.-X.; Vinocur, B.; Shoseyov, O.; Altman, A. Role of plant heat-shock proteins and molecular chaperones in the abiotic stress response. Trends Plant Sci. 2004, 9, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Swindell, W.R.; Huebner, M.; Weber, A.P. Transcriptional profiling of Arabidopsis heat shock proteins and transcription factors reveals extensive overlap between heat and non-heat stress response pathways. BMC Genom. 2007, 8, 125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, E.K.; Choi, Y.J. A nuclear-localized HSP70 confers thermoprotective activity and drought-stress tolerance on plants. Biotechnol. Lett. 2008, 31, 597–606. [Google Scholar] [CrossRef]
- Zou, J.; Liu, C.; Liu, A.; Zou, D.; Chen, X. Overexpression of OsHsp17.0 and OsHsp23.7 enhances drought and salt tolerance in rice. J. Plant Physiol. 2012, 169, 628–635. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.-M.; Rhee, J.-S.; Jeong, C.-B.; Seo, J.S.; Park, G.S.; Lee, Y.-M.; Lee, J.-S. Heavy metals induce oxidative stress and trigger oxidative stress-mediated heat shock protein (hsp) modulation in the intertidal copepod Tigriopus japonicus. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2014, 166, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Heckathorn, S.A.; Mueller, J.K.; LaGuidice, S.; Zhu, B.; Barrett, T.; Blair, B.; Dong, Y. Chloroplast small heat-shock proteins protect photosynthesis during heavy metal stress. Am. J. Bot. 2004, 91, 1312–1318. [Google Scholar] [CrossRef] [PubMed]
- Neumann, D.; Lichtenberger, O.; Tschiersch, K.; Nover, L. Heat-shock proteins induce heavy-metal tolerance in higher plants. Planta 1994, 194, 360–367. [Google Scholar] [CrossRef]
- Vierling, E. The Roles of Heat Shock Proteins in Plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1991, 42, 579–620. [Google Scholar] [CrossRef]
- Mayer, M.P.; Bukau, B. Hsp70 chaperones: Cellular functions and molecular mechanism. Cell. Mol. Life Sci. 2005, 62, 670–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dragovic, Z.; Broadley, S.A.; Shomura, Y.; Bracher, A.; Hartl, F.U. Molecular chaperones of the Hsp110 family act as nucleotide exchange factors of Hsp70s. EMBO J. 2006, 25, 2519–2528. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.; Zhao, X.; Burkholder, W.F.; Gragerov, A.; Ogata, C.M.; Gottesman, M.E.; Hendrickson, W.A. Structural Analysis of Substrate Binding by the Molecular Chaperone DnaK. Science 1996, 272, 1606–1614. [Google Scholar] [CrossRef] [Green Version]
- Flaherty, K.M.; DeLuca-Flaherty, C.; McKay, D.B. Three-dimensional structure of the ATPase fragment of a 70K heat-shock cognate protein. Nat. Cell Biol. 1990, 346, 623–628. [Google Scholar] [CrossRef]
- Daugaard, M.; Rohde, M.; Jäättelä, M. The heat shock protein 70 family: Highly homologous proteins with overlapping and distinct functions. FEBS Lett. 2007, 581, 3702–3710. [Google Scholar] [CrossRef] [Green Version]
- Alvim, F.C.; Carolino, S.M.B.; Cascardo, J.C.M.; Nunes, C.C.; Martinez, C.A.; Otoni, W.C.; Fontes, E.P.B. Enhanced Accumulation of BiP in Transgenic Plants Confers Tolerance to Water Stress. Plant Physiol. 2001, 126, 1042–1054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, E.K.; Hong, C.B. Over-expression of tobacco NtHSP70-1 contributes to drought-stress tolerance in plants. Plant Cell Rep. 2006, 25, 349–358. [Google Scholar] [CrossRef]
- Augustine, S.M.; Cherian, A.V.; Syamaladevi, D.P.; Subramonian, N. Erianthus arundinaceus HSP70 (EaHSP70) acts as a key regulator in the formation of anisotropic interdigitation in sugarcane (Saccharum spp. hybrid) in response to drought stress. Plant Cell Physiol. 2015, 56, 2368–2380. [Google Scholar] [CrossRef] [Green Version]
- Mosaviazam, B.; Ramezani, A.; Morowvat, M.H.; Niazi, A.; Mousavi, P.; Moghadam, A.; Zarrini, G.; Ghasemi, Y. HSP70 Gene Expression Analysis in Dunaliella salina Under Salt Stress. Int. J. Pharm. Phytochem. Res. 2016, 8, 767–770. [Google Scholar]
- Ndimba, B.K.; Thomas, L.A.; Ngara, R. Sorghum 2-Dimensional Proteome Profiles and Analysis of HSP70 Expression Under Salinity Stress. Agric. Nat. Resour. 2010, 44, 768–775. [Google Scholar]
- Su, P.-H.; Li, H.-M. Arabidopsis Stromal 70-kD Heat Shock Proteins Are Essential for Plant Development and Important for Thermotolerance of Germinating Seeds. Plant Physiol. 2008, 146, 1231–1241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Latijnhouwers, M.; Xu, X.-M.; Møller, S.G. Arabidopsis stromal 70-kDa heat shock proteins are essential for chloroplast development. Planta 2010, 232, 567–578. [Google Scholar] [CrossRef] [PubMed]
- Lin, B.-L.; Wang, J.-S.; Barakat, A.; Liu, H.-C.; Delseny, M. Genomic analysis of the Hsp70 superfamily in Arabidopsis thaliana. Cell Stress Chaperones 2001, 6, 201. [Google Scholar] [CrossRef]
- Liu, J.; Wang, R.; Liu, W.; Zhang, H.; Guo, Y.; Wen, R. Genome-Wide Characterization of Heat-Shock Protein 70s from Chenopodium quinoa and Expression Analyses of Cqhsp70s in Response to Drought Stress. Genes 2018, 9, 35. [Google Scholar] [CrossRef] [Green Version]
- Divya, K.; Kishor, P.B.K.; Singam, P.; Maraka, N.; Vadez, V.; Reddy, P.S.; Bhatnagar-Mathur, P. Genome-wide Identification and Characterization of Hsp70 gene family in Pearl millet (Pennisetumglaucum). Curr. Trends Biotechnol. Pharm. 2019, 13, 102–111. [Google Scholar]
- Zhang, L.; Zhao, H.-K.; Dong, Q.-L.; Zhang, Y.-Y.; Wang, Y.-M.; Li, H.-Y.; Xing, G.-J.; Li, Q.-Y.; Dong, Y.-S. Genome-wide analysis and expression profiling under heat and drought treatments of HSP70 gene family in soybean (Glycine max L.). Front. Plant Sci. 2015, 6, 773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, P.W.; Xu, Y.H.; Yu, C.C.; Lv, F.F.; Tang, X.L.; Gao, Z.H.; Zhang, Z.; Wang, H.; Liu, Y.; Wei, J.H. WRKY44 represses expression of the wound-induced sesquiterpene biosynthetic gene ASS1 in Aquilaria sinensis. J. Exp. Bot. 2020, 71, 1128–1138. [Google Scholar]
- Takemoto, H.; Ito, M.; Shiraki, T.; Yagura, T.; Honda, G. Sedative effects of vapor inhalation of agarwood oil and spikenard extract and identification of their active components. J. Nat. Med. 2007, 62, 41–46. [Google Scholar] [CrossRef]
- Kakino, M.; Tazawa, S.; Maruyama, H.; Tsuruma, K.; Araki, Y.; Shimazawa, M.; Hara, H. Laxative effects of agarwood on low-fiber diet-induced constipation in rats. BMC Complement. Altern. Med. 2010, 10, 68. [Google Scholar] [CrossRef] [Green Version]
- Lancaster, C.; Espinoza, E. Evaluating agarwood products for 2-(2-phenylethyl)chromones using direct analysis in real time time-of-flight mass spectrometry. Rapid Commun. Mass Spectrom. 2012, 26, 2649–2656. [Google Scholar] [CrossRef]
- Mei, W.-L.; Yang, D.-L.; Wang, H.; Yang, J.-L.; Zeng, Y.-B.; Guo, Z.-K.; Dong, W.-H.; Li, W.; Dai, H.-F. Characterization and Determination of 2-(2-Phenylethyl)chromones in Agarwood by GC-MS. Molecules 2013, 18, 12324–12345. [Google Scholar] [CrossRef] [PubMed]
- China Pharmacopoeia Committee. The Pharmacopoeia of People’s Republic of China (I); Chemical Industry Press: Beijing, China, 2020. [Google Scholar]
- Zhang, Z.; Wei, J.; Han, X.; Liang, L.; Yang, Y.; Meng, H.; Xu, Y.; Gao, Z. The Sesquiterpene Biosynthesis and Vessel-Occlusion Formation in Stems of Aquilaria sinensis (Lour.) Gilg Trees Induced by Wounding Treatments without Variation of Microbial Communities. Int. J. Mol. Sci. 2014, 15, 23589–23603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Zhang, X.; Yang, Y.; Wei, J.-H.; Meng, H.; Gao, Z.-H.; Xu, Y.-H. Hydrogen peroxide induces vessel occlusions and stimulates sesquiterpenes accumulation in stems of Aquilaria sinensis. Plant Growth Regul. 2013, 72, 81–87. [Google Scholar] [CrossRef]
- Liu, P.; Zhang, X.; Yang, Y.; Sui, C.; Xu, Y.; Wei, J. Interxylary phloem and xylem rays are the structural foundation of agarwood resin formation in the stems of Aquilaria sinensis. Trees 2018, 33, 533–542. [Google Scholar] [CrossRef]
- Xu, Y.; Zhang, Z.; Wang, M.; Wei, J.; Chen, H.; Gao, Z.; Sui, C.; Luo, H.; Zhang, X.; Yang, Y.; et al. Identification of genes related to agarwood formation: Transcriptome analysis of healthy and wounded tissues of Aquilaria sinensis. BMC Genom. 2013, 14, 227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Yang, Y.; Xue, J.; Wei, J.; Zhang, Z.; Chen, H. Comparison of Compositions and Antimicrobial Activities of Essential Oils from Chemically Stimulated Agarwood, Wild Agarwood and Healthy Aquilaria sinensis (Lour.) Gilg Trees. Molecules 2011, 16, 4884–4896. [Google Scholar] [CrossRef] [Green Version]
- Hu, B.; Jin, J.; Guo, A.-Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef] [Green Version]
- Voorrips, R.E. MapChart: Software for the graphical presentation of linkage maps and QTLs. J. Hered. 2002, 93, 77–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gasteiger, E.; Gattiker, A.; Hoogland, C.; Ivanyi, I.; Appel, R.D.; Bairoch, A. ExPASy: The proteomics server for in-depth protein knowledge and analysis. Nucl. Acids Res. 2003, 31, 3784–3788. [Google Scholar] [CrossRef] [Green Version]
- Wei, J.H.; Yang, Y.; Zhang, Z.; Meng, H.; Feng, J.D.; Gan, B.C. Production of agarwood in Aquilaria sinensis trees via transfusion technique. China Patent CN101755629 B, 1 February 2010. [Google Scholar]
- Kim, J.; Somers, D.E. Rapid assessment of gene function in the circadian clock using artificial microRNA in Arabidopsis mesophyll protoplasts. Plant Physiol. 2010, 154, 611–621. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.; Pan, F.; Lou, X.; Wang, D.; Yang, C.; Zhang, B.; Zhang, H. Genome-wide identification and characterization of Hsp70 gene family in Nicotiana tabacum. Mol. Biol. Rep. 2019, 46, 1941–1954. [Google Scholar] [CrossRef]
- Sierro, N.; Battey, J.N.; Ouadi, S.; Bakaher, N.; Bovet, L.; Willig, A.L.; Goepfert, S.; Peitsch, M.; Ivanov, N.V. The tobacco genome sequence and its comparison with those of tomato and potato. Nat. Commun. 2014, 5, 3833. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, H.; Khurana, N.; Agarwal, P.; Khurana, P. Heat shock factors in rice (Oryza sativa L.): Genome-wide expression analysis during reproductive development and abiotic stress. Mol. Genet. Genom. 2011, 286, 171–187. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Chen, H.; Yang, Y.; Zhang, Z.; Wei, J.; Meng, H.; Chen, W.; Feng, J.; Gan, B.; Chen, X.; et al. Whole-tree Agarwood-Inducing Technique: An Efficient Novel Technique for Producing High-Quality Agarwood in Cultivated Aquilaria sinensis Trees. Molecules 2013, 18, 3086–3106. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.-H.; Liao, Y.-C.; Zhang, Z.; Liu, J.; Sun, P.-W.; Gao, Z.-H.; Sui, C.; Wei, J.-H. Jasmonic acid is a crucial signal transducer in heat shock induced sesquiterpene formation in Aquilaria sinensis. Sci. Rep. 2016, 6, 21843. [Google Scholar] [CrossRef] [PubMed]
- Tamuli, P.; Boruah, P.; Samanta, R. Biochemical changes in agarwood tree (Aquilaria malaccensis Lamk.) during pathogenesis. J. Spices Aromat. Crop. 2004, 13, 87–91. [Google Scholar]
- Su, H.; Xing, M.; Liu, X.; Fang, Z.; Yang, L.; Zhuang, M.; Zhang, Y.; Wang, Y.; Lv, H. Genome-wide analysis of HSP70 family genes in cabbage (Brassica oleracea var. capitata) reveals their involvement in floral development. BMC Genom. 2019, 20, 369. [Google Scholar] [CrossRef]
- Liu, J.; Pang, X.; Cheng, Y.; Yin, Y.; Zhang, Q.; Su, W.; Hu, B.; Guo, Q.; Ha, S.; Zhang, J.; et al. The Hsp70 Gene Family in Solanum tuberosum: Genome-Wide Identification, Phylogeny, and Expression Patterns. Sci. Rep. 2018, 8, 16628. [Google Scholar] [CrossRef]
- Skolnick, J.; Fetrow, J. From genes to protein structure and function: Novel applications of computational approaches in the genomic era. Trends Biotechnol. 2000, 18, 34–39. [Google Scholar] [CrossRef]
- Chen, J.; Gao, T.; Wan, S.; Zhang, Y.; Yang, J.; Yu, Y.; Wang, W. Genome-Wide Identification, Classification and Expression Analysis of the HSP Gene Superfamily in Tea Plant (Camellia sinensis). Int. J. Mol. Sci. 2018, 19, 2633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, Z.; Chandrasekaran, U.; Liu, A. Genome-wide analysis of the Dof transcription factors in castor bean (Ricinus communis L.). Genes Genom. 2014, 36, 527–537. [Google Scholar] [CrossRef]
- Jeffares, D.C.; Penkett, C.J.; Bähler, J. Rapidly regulated genes are intron poor. Trends Genet. 2008, 24, 375–378. [Google Scholar] [CrossRef] [PubMed]
- Sung, D.Y.; Vierling, E.; Guy, C.L. Comprehensive expression profile analysis of the Arabidopsis Hsp70 gene family. Plant Physiol. 2001, 126, 789–800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kose, S.; Furuta, M.; Imamoto, N. Hikeshi, a nuclear import carrier for Hsp70s, protects cells from heat shock-induced nuclear damage. Cell 2012, 149, 578–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Amino Acids | MW (kDa) | Exon Number | pI | GRVAY | Instability Index | Aliphatic Index | Subcellular Localization |
|---|---|---|---|---|---|---|---|---|
| AsHSP70-1 | 676 | 72.71 | 6 | 5.83 | −0.292 | 38.21 | 89.2 | mito: 8; chlo: 6 |
| AsHSP70-2 | 647 | 70.88 | 2 | 5.16 | −0.408 | 35.04 | 82.63 | cyto: 8; cysk: 5, chlo: 1 |
| AsHSP70-3 | 679 | 72.77 | 6 | 5.81 | −0.292 | 38.97 | 88.25 | mito: 11; chlo: 3 |
| AsHSP70-4 | 658 | 73.25 | 2 | 5.22 | −0.487 | 31.09 | 88.34 | vacu: 4, chlo: 3, mito: 3, golg: 2, nucl: 1, extr: 1 |
| AsHSP70-5 | 650 | 71.10 | 1 | 5.21 | −0.415 | 31.54 | 81.65 | cyto: 10; cysk: 2; chlo: 1; nucl: 1 |
| AsHSP70-6 | 699 | 74.66 | 8 | 5.3 | −0.299 | 28.82 | 87.58 | chlo: 14 |
| AsHSP70-7 | 489 | 53.74 | 7 | 6.38 | −0.269 | 37.64 | 86.36 | cyto: 10; cysk: 3; chlo: 1 |
| AsHSP70-8 | 747 | 83.43 | 9 | 5.19 | −0.287 | 40.53 | 89.02 | nucl: 7; vacu: 1; cysk: 2; chlo: 2; cyto: 2 |
| AsHSP70-9 | 551 | 61.37 | 1 | 5.46 | −0.422 | 39.27 | 88.82 | cyto: 9; cysk: 3, chlo: 2 |
| AsHSP70-10 | 667 | 73.54 | 8 | 5.2 | −0.449 | 27.24 | 88.58 | E.R.: 14 |
| AsHSP70-11 | 648 | 70.94 | 2 | 5.16 | −0.422 | 33.08 | 82.19 | cyto: 8, cysk: 5, chlo: |
| AsHSP70-12 | 856 | 94.75 | 9 | 5.12 | −0.472 | 40.68 | 77.94 | cyto: 8; chlo: 3; nucl: 2; cysk: 1 |
| AsHSP70-13 | 648 | 70.98 | 2 | 5.17 | −0.407 | 34.1 | 82.35 | cyto: 9, cysk: 3, chlo: 2 |
| AsHSP70-14 | 571 | 61.90 | 1 | 5.4 | 0.075 | 35.79 | 103.26 | cyto: 9; chlo: 4; vacu: 1 |
| AsHSP70-15 | 893 | 100.00 | 14 | 5.26 | −0.45 | 42.41 | 86.7 | E.R. 14 |
| Plant Species | Phylogenetic Class | Number | |||||
|---|---|---|---|---|---|---|---|
| I | II | III | IV | V | VI | ||
| Aquilaria sinensis | 3 | 2 | 3 | 6 | 1 | 15 | |
| Arabidopsis thaliana | 5 | 3 | 3 | 2 | 13 | ||
| Amborella trichopoda | 5 | 3 | 4 | 6 | 1 | 19 | |
| Glycine max | 12 | 4 | 5 | 19 | 1 | 41 | |
| Hordeum vulgare | 3 | 4 | 6 | 2 | 12 | 27 | |
| Marchantia polymorpha | 2 | 1 | 3 | 1 | 2 | 9 | |
| Oryza sativa | 4 | 5 | 5 | 1 | 11 | 1 | 27 |
| Picea abies | 2 | 1 | 4 | 12 | 1 | 20 | |
| Selaginella moellendorffii | 5 | 6 | 6 | 6 | 2 | 25 | |
| Motifs | Width (aa) | Best Possible Match |
|---|---|---|
| Motif 1 | 50 | SRAGRDGRWBVAGVWKGCKGAGRAGKWYNWGDSNRAGGACAAGAAGRASA |
| Motif 2 | 50 | RTHRWCMCYGWKSMDGCYDWYKYYWATGRYGCYSMRRDYCARGCHRCHAW |
| Motif 3 | 50 | RAVGKBRAGRMVAMYRYYGKDGWYWYYSAYCTBGGTGGTGRDRMYTTTGA |
| Motif 4 | 38 | RAVRARGTCCADGANNTKSTKCTKBTSGDTGKKACTHC |
| Motif 5 | 50 | GGCATYCCWCCWGCNCCAAGRGGWGTTCCKCARATHRAWGTBWSMTTYGA |
| Motif 6 | 50 | GGHMTKGARACTGYTGGTGGWGTNATGACHRWRTTGATTCCHAGRAACAC |
| Motif 7 | 50 | ATTGCDGGSYTDAAWGTNVTKAGDATVATYAATGAGCCMACWGCWGCTGC |
| Motif 8 | 50 | ATYMARGTMTWYGAAGGTGARAGAGCRAKRRCMARRGAYAAYAAMTTKCT |
| Motif 9 | 50 | GGDATTGAYTTCKRYWCSACNWWYWCYHGTGYYRGHKTYKAGGAGCDSRA |
| Motif 10 | 50 | AACCCMRMRAACACMRTYTTCGAKGYCAAGMGKYTBATYGGHAGDARATT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, C.; Rong, M.; Liu, Y.; Sun, P.; Xu, Y.; Wei, J. Genome-Wide Identification and Characterization of HSP70 Gene Family in Aquilaria sinensis (Lour.) Gilg. Genes 2022, 13, 8. https://doi.org/10.3390/genes13010008
Yu C, Rong M, Liu Y, Sun P, Xu Y, Wei J. Genome-Wide Identification and Characterization of HSP70 Gene Family in Aquilaria sinensis (Lour.) Gilg. Genes. 2022; 13(1):8. https://doi.org/10.3390/genes13010008
Chicago/Turabian StyleYu, Cuicui, Mei Rong, Yang Liu, Peiwen Sun, Yanhong Xu, and Jianhe Wei. 2022. "Genome-Wide Identification and Characterization of HSP70 Gene Family in Aquilaria sinensis (Lour.) Gilg" Genes 13, no. 1: 8. https://doi.org/10.3390/genes13010008
APA StyleYu, C., Rong, M., Liu, Y., Sun, P., Xu, Y., & Wei, J. (2022). Genome-Wide Identification and Characterization of HSP70 Gene Family in Aquilaria sinensis (Lour.) Gilg. Genes, 13(1), 8. https://doi.org/10.3390/genes13010008