
Intermittent Hypoxia Alters the Circadian Expression of Clock Genes in Mouse Brain and Liver

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design and Sample Collection
2.2. Gene Expression Analysis
2.3. Analysis of Circadian Rhythms
3. Results
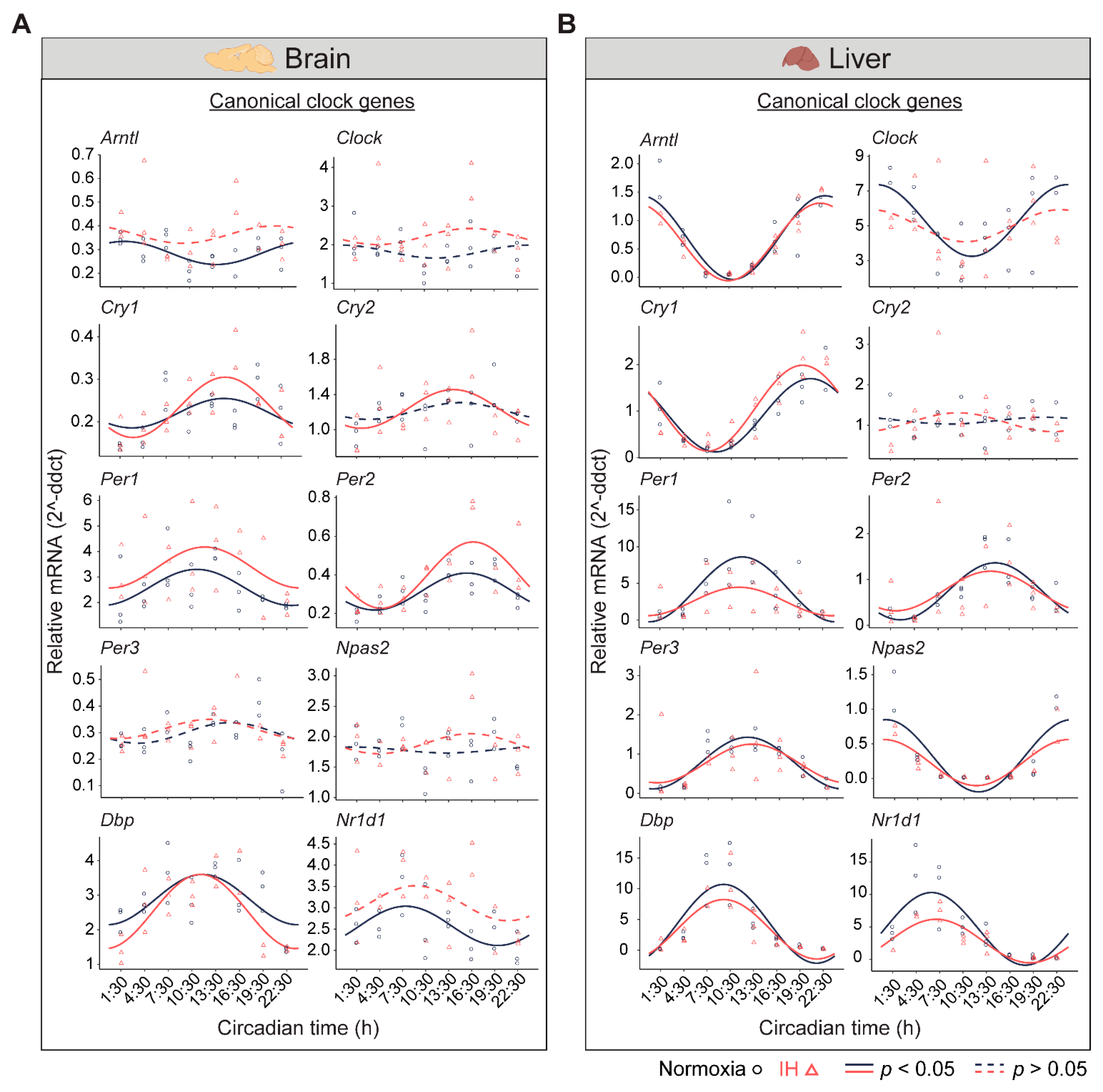
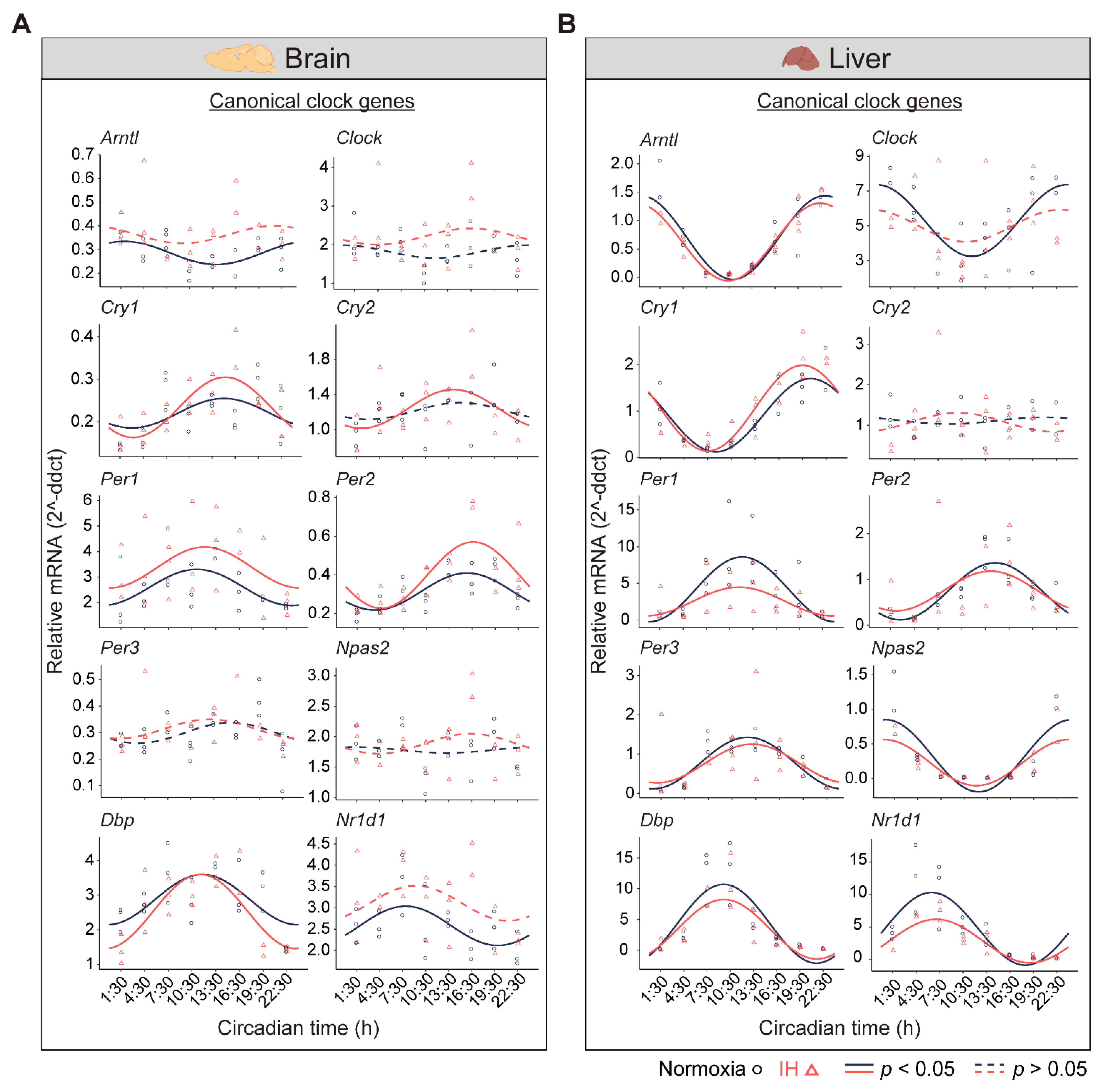
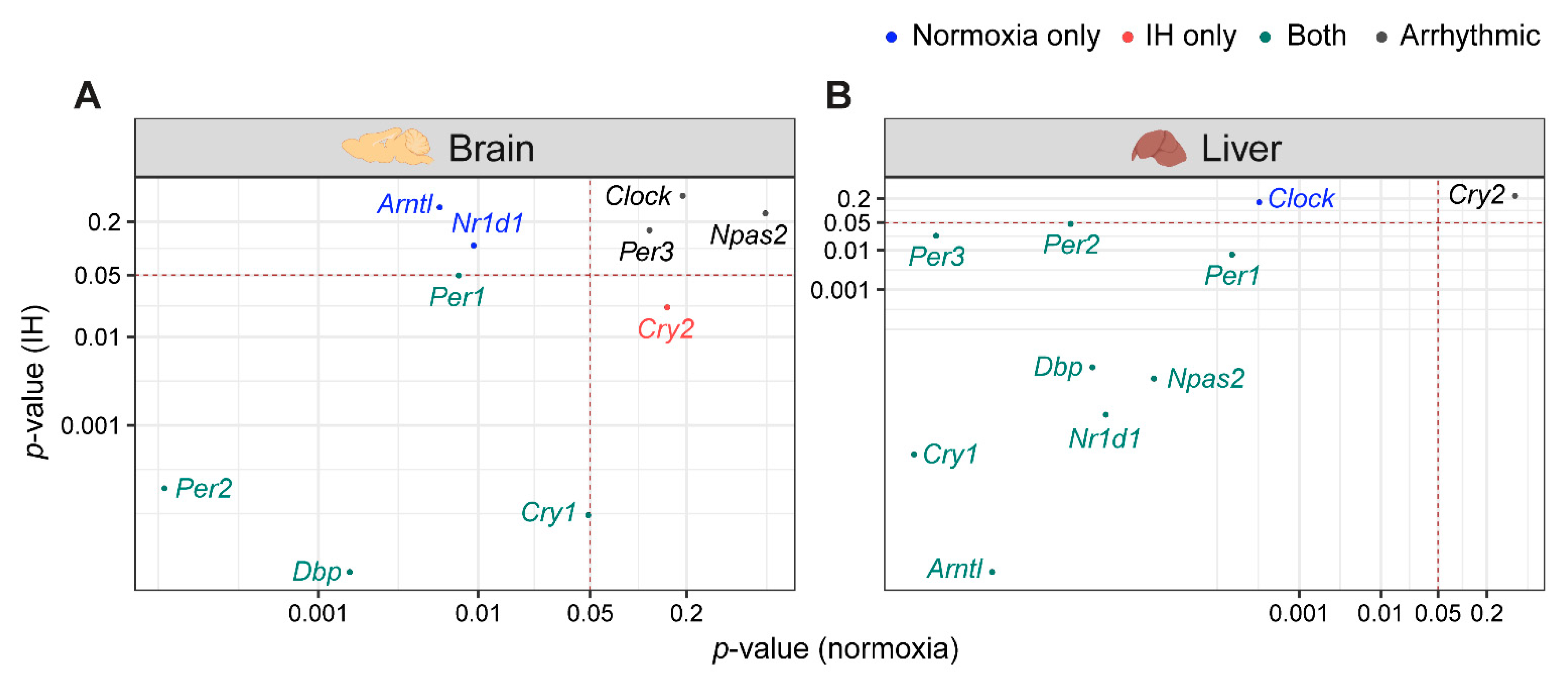
3.1. IH Alters Tissue-Specific Circadian Rhythms of Canonical Clock Genes
3.2. The Clock in the Liver Is More Stable against IH Compared to the Clock in the Brain
4. Discussion
5. Limitations
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wagner, P.D. The biology of oxygen. Eur. Respir. J. 2008, 31, 887–890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tripathi, A.; Melnik, A.V.; Xue, J.; Poulsen, O.; Meehan, M.J.; Humphrey, G.; Jiang, L.; Ackermann, G.; McDonald, D.; Zhou, D.; et al. Intermittent hypoxia and hypercapnia, a hallmark of obstructive sleep apnea, alters the gut microbiome and metabolome. mSystems 2018, 3, e00020-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zieliński, J. Effects of intermittent hypoxia on pulmonary haemodynamics: Animal models versus studies in humans. Eur. Respir. J. 2005, 25, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Neubauer, J.A. Invited review: Physiological and pathophysiological responses to intermittent hypoxia. J. Appl. Physiol. 2001, 90, 1593–1599. [Google Scholar] [CrossRef] [PubMed]
- Adamovich, Y.; Ladeuix, B.; Sobel, J.; Manella, G.; Neufeld-Cohen, A.; Assadi, M.H.; Golik, M.; Kuperman, Y.; Tarasiuk, A.; Koeners, M.P.; et al. Oxygen and carbon dioxide rhythms are circadian clock controlled and differentially directed by behavioral signals. Cell Metab. 2019, 29, 1092–1103.e3. [Google Scholar] [CrossRef] [PubMed]
- Adamovich, Y.; Ladeuix, B.; Golik, M.; Koeners, M.P.; Asher, G. Rhythmic oxygen levels reset circadian clocks through HIF1α. Cell Metab. 2017, 25, 93–101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bersten, D.C.; Sullivan, A.E.; Peet, D.J.; Whitelaw, M.L. bHLH-PAS proteins in cancer. Nat. Rev. Cancer 2013, 13, 827–841. [Google Scholar] [CrossRef] [PubMed]
- O’Connell, E.J.; Martinez, C.-A.; Liang, Y.G.; Cistulli, P.A.; Cook, K.M. Out of breath, out of time: Interactions between HIF and circadian rhythms. Am. J. Physiol. Cell Physiol. 2020, 319, C533–C540. [Google Scholar] [CrossRef] [PubMed]
- Manella, G.; Aviram, R.; Bolshette, N.; Muvkadi, S.; Golik, M.; Smith, D.F.; Asher, G. Hypoxia induces a time- and tissue-specific response that elicits intertissue circadian clock misalignment. Proc. Natl. Acad. Sci. USA 2020, 117, 779–786. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Lee, Y.Y.; Gulla, E.M.; Potter, A.; Kitzmiller, J.; Ruben, M.D.; Salomonis, N.; Whitsett, J.A.; Francey, L.J.; Hogenesch, J.B.; et al. Short-term exposure to intermittent hypoxia leads to changes in gene expression seen in chronic pulmonary disease. Elife 2021, 10, e63003. [Google Scholar] [CrossRef] [PubMed]
- Parsons, R.; Parsons, R.; Garner, N.; Oster, H.; Rawashdeh, O. CircaCompare: A method to estimate and statistically support differences in mesor, amplitude and phase, between circadian rhythms. Bioinformatics 2020, 36, 1208–1212. [Google Scholar] [CrossRef] [PubMed]
- Casas, A.I.; Geuss, E.; Kleikers, P.W.M.; Mencl, S.; Herrmann, A.M.; Buendia, I.; Egea, J.; Meuth, S.G.; Lopez, M.G.; Kleinschnitz, C.; et al. NOX4-dependent neuronal autotoxicity and BBB breakdown explain the superior sensitivity of the brain to ischemic damage. Proc. Natl. Acad. Sci. USA 2017, 114, 12315–12320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leach, R.M.; Treacher, D.F. Oxygen transport-2. Tissue hypoxia. BMJ 1998, 317, 1370–1373. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Lahens, N.F.; Ballance, H.I.; Hughes, M.E.; Hogenesch, J.B. A circadian gene expression atlas in mammals: Implications for biology and medicine. Proc. Natl. Acad. Sci. USA 2014, 111, 16219–16224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manella, G.; Sabath, E.; Aviram, R.; Dandavate, V.; Ezagouri, S.; Golik, M.; Adamovich, Y.; Asher, G. The liver-clock coordinates rhythmicity of peripheral tissues in response to feeding. Nat. Metab. 2021, 3, 829–842. [Google Scholar] [CrossRef] [PubMed]
- Pizarro, A.; Hayer, K.; Lahens, N.F.; Hogenesch, J.B. CircaDB: A da-tabase of mammalian circadian gene expression profiles. Nucleic Acids Res. 2013, 41, D1009–D1013. [Google Scholar]
- Albrecht, U. Timing to perfection: The biology of central and peripheral circadian clocks. Neuron 2012, 74, 246–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benjafield, A.V.; Ayas, N.T.; Eastwood, P.R.; Heinzer, R.; Ip, M.S.M.; Morrell, M.J.; Nunez, C.M.; Patel, S.R.; Penzel, T.; Pépin, J.-L.; et al. Estimation of the global prevalence and burden of obstructive sleep apnoea: A literature-based analysis. Lancet Respir. Med. 2019, 7, 687–698. [Google Scholar] [CrossRef] [Green Version]
- Stephenson, R. Circadian rhythms and sleep-related breathing disorders. Sleep Med. 2007, 8, 681–687. [Google Scholar] [CrossRef] [PubMed]
- Lyons, M.M.; Bhatt, N.Y.; Pack, A.I.; Magalang, U.J. Global burden of sleep-disordered breathing and its implications. Respirology 2020, 25, 690–702. [Google Scholar] [CrossRef] [PubMed]
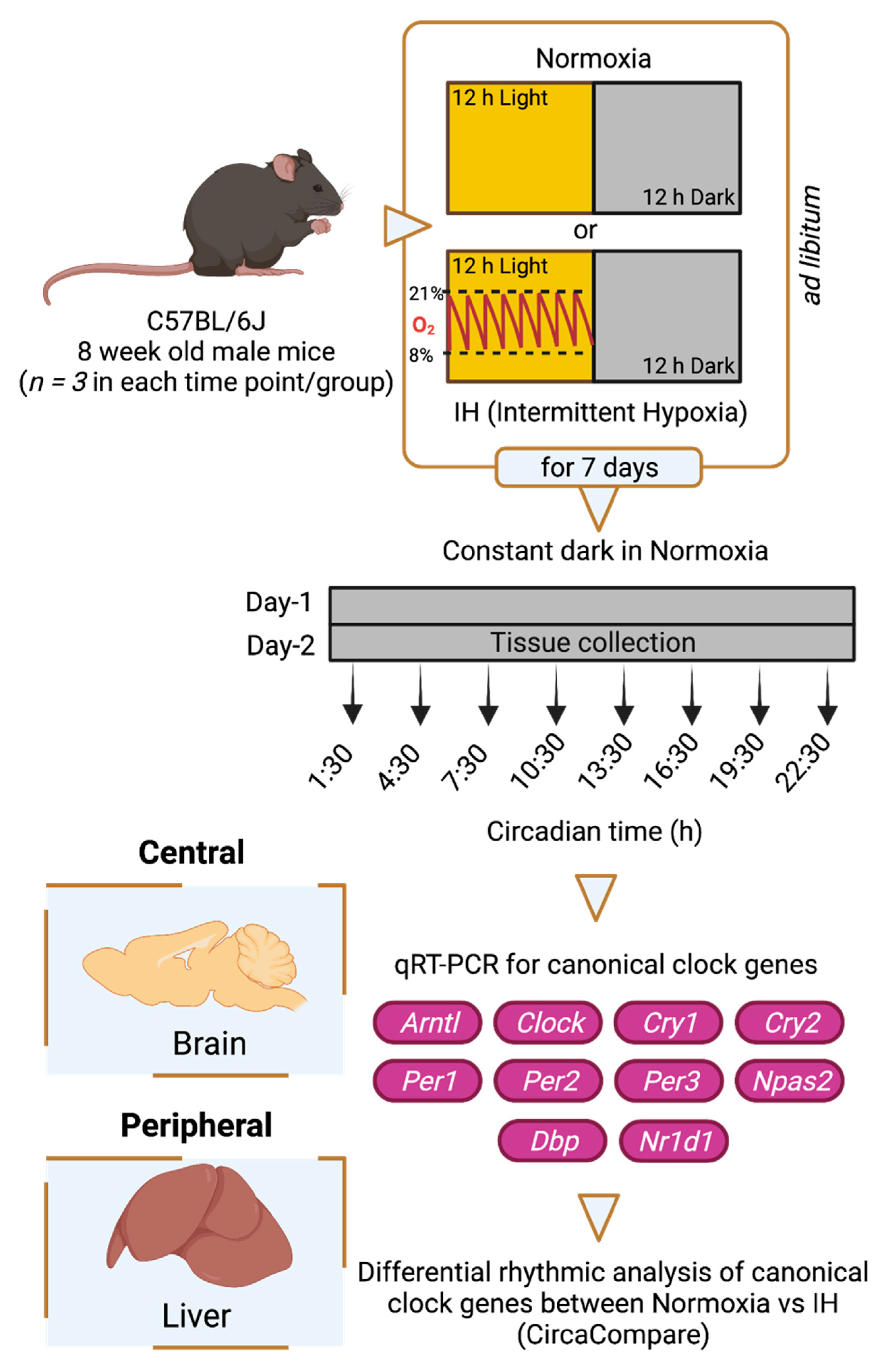


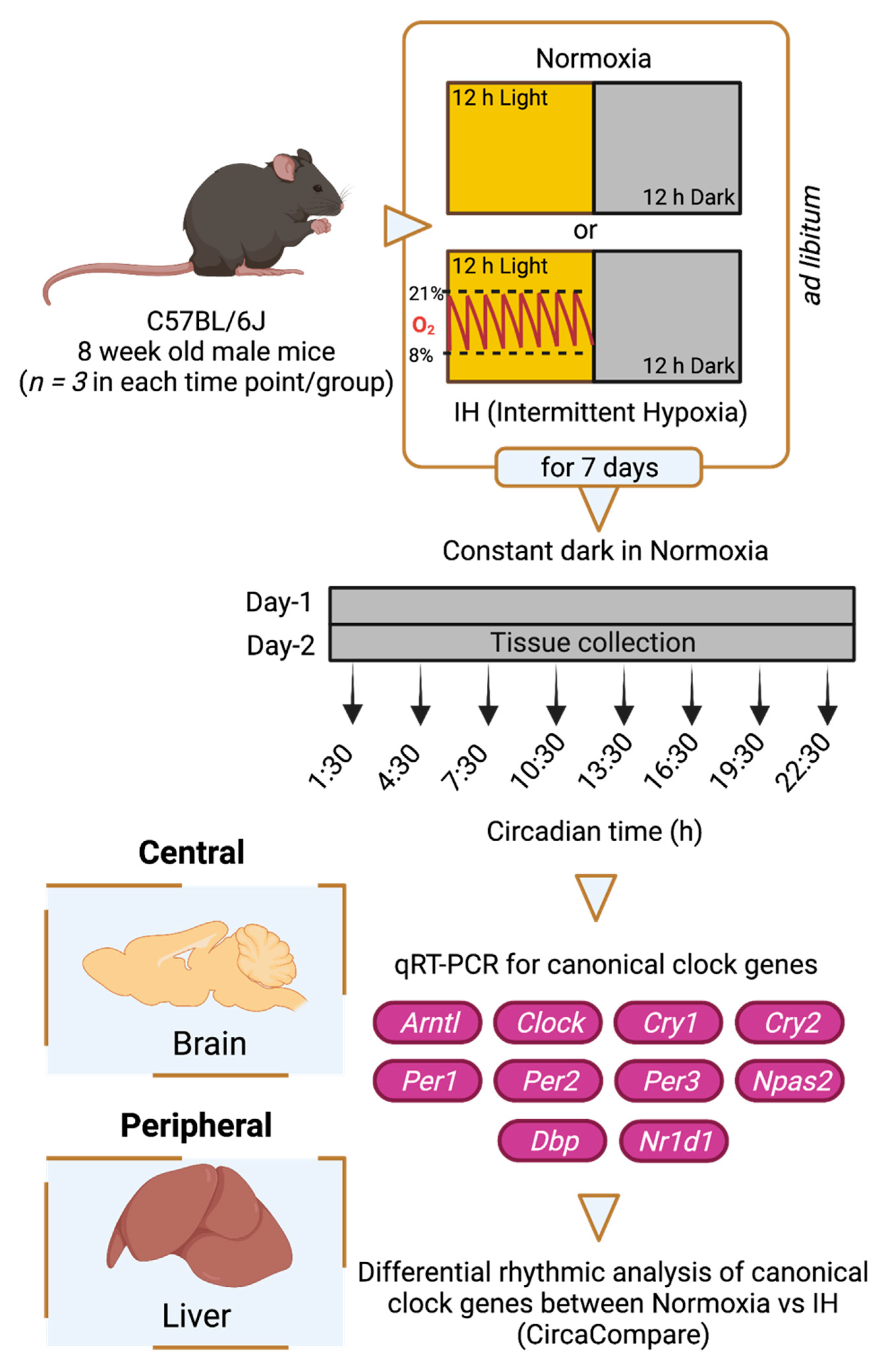{kind=link}
{kind=link}
{kind=link}
| Gene Name | Assay ID |
|---|---|
| Arntl | Mm00500226_m1 |
| Clock | Mm00455950_m1 |
| Cry1 | Mm00514392_m1 |
| Cry2 | Mm01331539_m1 |
| Dbp | Mm00497539_m1 |
| Npas2 | Mm00500848_m1 |
| Nr1d1 | Mm00520708_m1 |
| Per1 | Mm00501813_m1 |
| Per2 | Mm00478113_m1 |
| Per3 | Mm00478120_m1 |
| Gapdh | Mm99999915_g1 |
| Ppib | Mm00478295_m1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koritala, B.S.C.; Lee, Y.Y.; Bhadri, S.S.; Gaspar, L.S.; Stanforth, C.; Wu, G.; Ruben, M.D.; Francey, L.J.; Smith, D.F. Intermittent Hypoxia Alters the Circadian Expression of Clock Genes in Mouse Brain and Liver. Genes 2021, 12, 1627. https://doi.org/10.3390/genes12101627
Koritala BSC, Lee YY, Bhadri SS, Gaspar LS, Stanforth C, Wu G, Ruben MD, Francey LJ, Smith DF. Intermittent Hypoxia Alters the Circadian Expression of Clock Genes in Mouse Brain and Liver. Genes. 2021; 12(10):1627. https://doi.org/10.3390/genes12101627
Chicago/Turabian StyleKoritala, Bala S. C., Yin Yeng Lee, Shweta S. Bhadri, Laetitia S. Gaspar, Corinne Stanforth, Gang Wu, Marc D. Ruben, Lauren J. Francey, and David F. Smith. 2021. "Intermittent Hypoxia Alters the Circadian Expression of Clock Genes in Mouse Brain and Liver" Genes 12, no. 10: 1627. https://doi.org/10.3390/genes12101627
APA StyleKoritala, B. S. C., Lee, Y. Y., Bhadri, S. S., Gaspar, L. S., Stanforth, C., Wu, G., Ruben, M. D., Francey, L. J., & Smith, D. F. (2021). Intermittent Hypoxia Alters the Circadian Expression of Clock Genes in Mouse Brain and Liver. Genes, 12(10), 1627. https://doi.org/10.3390/genes12101627