Transcriptional Regulation of HMOX1 Gene in Hezuo Tibetan Pigs: Roles of WT1, Sp1, and C/EBPα

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Quantitative Real-Time PCR Analysis of Gene Expression Patterns
2.3. HMOX1 Gene Promoter Region Cloning and Bioinformatics Analysis
2.4. Cell Culture, Transfection, and Dual-Luciferase Reporter Assay
2.5. Site-Directed Mutagenesis
2.6. RNA Interference
2.7. Statistical Analysis
3. Results
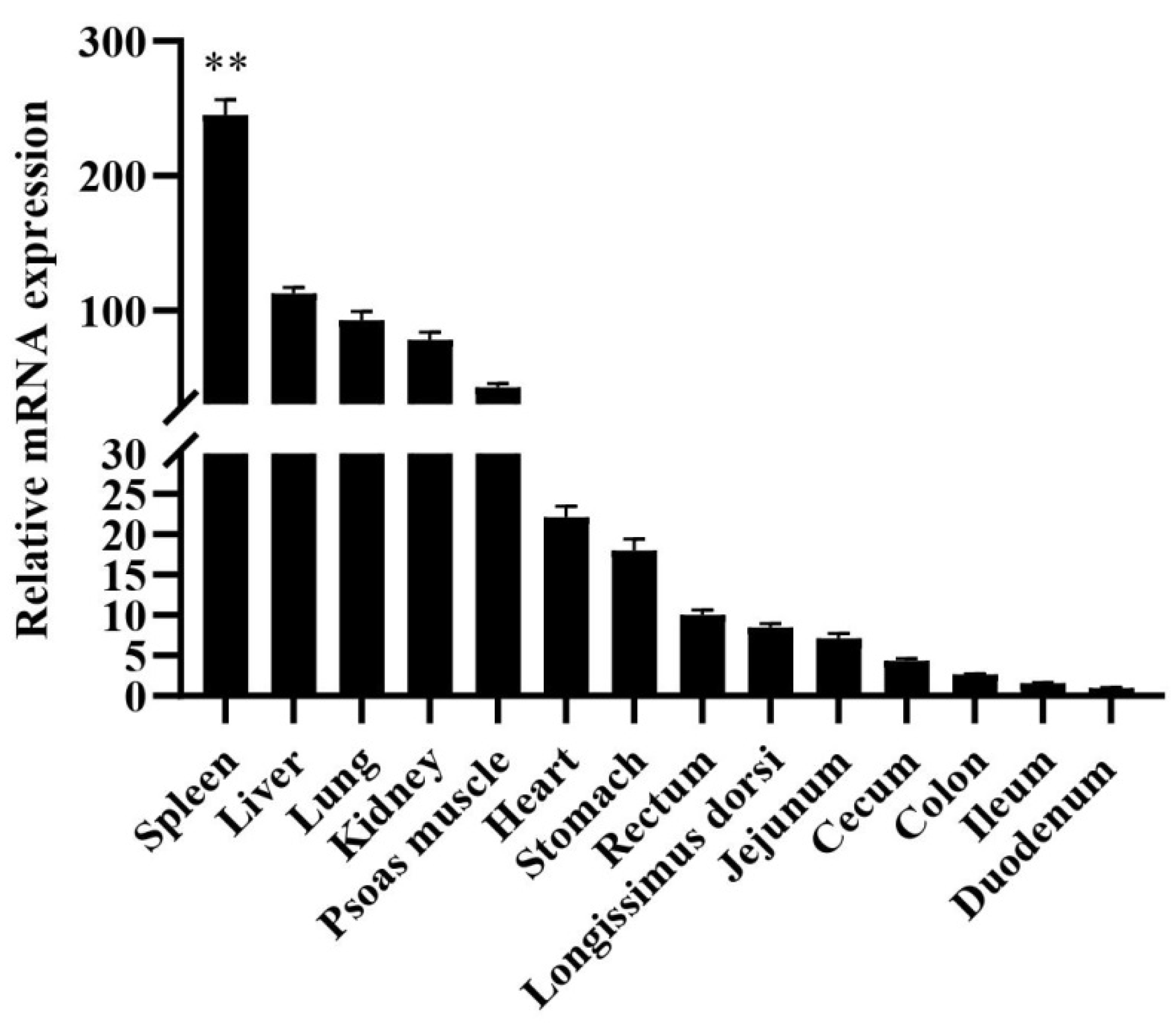
3.1. Tissue Expression Analysis of mRNA
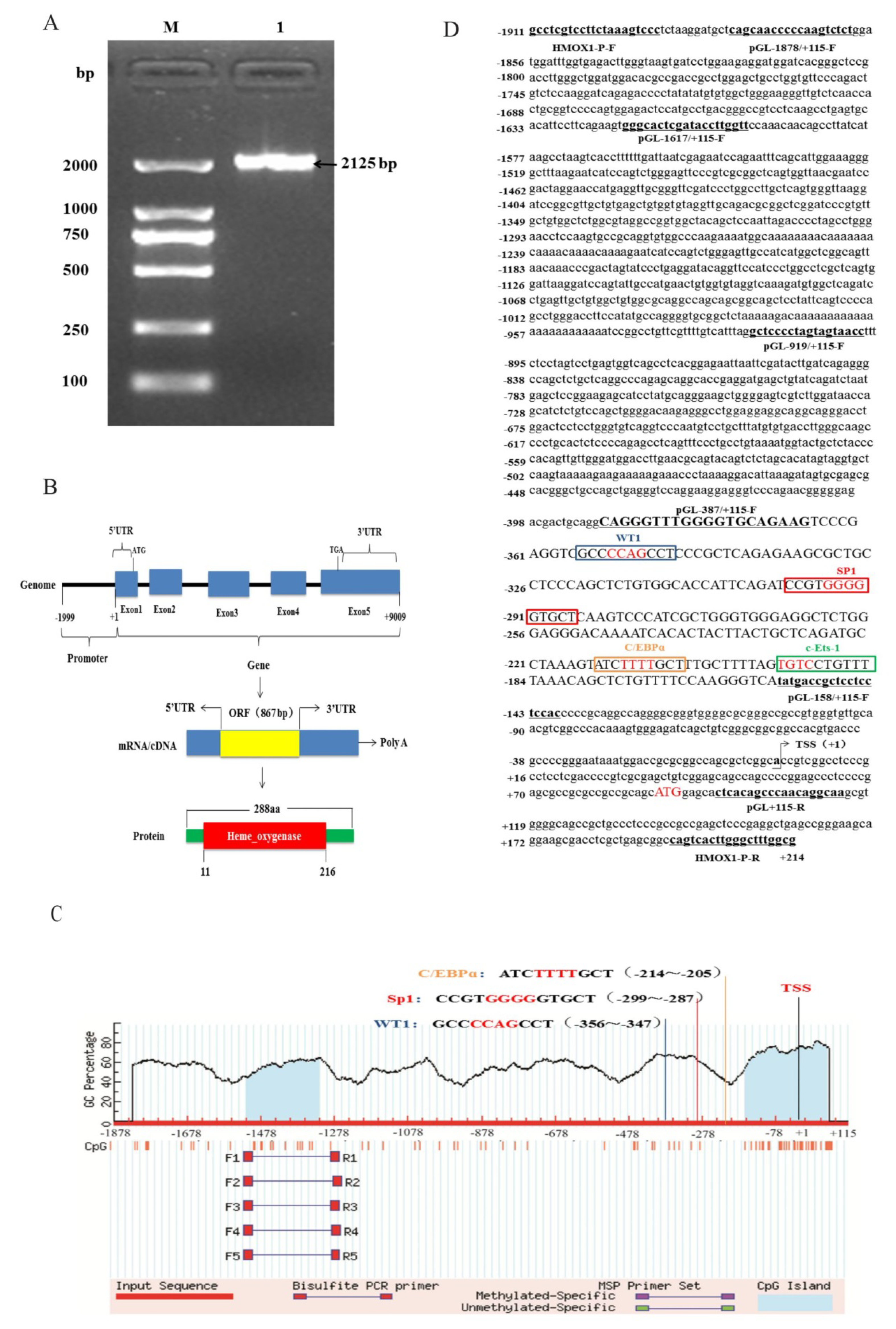
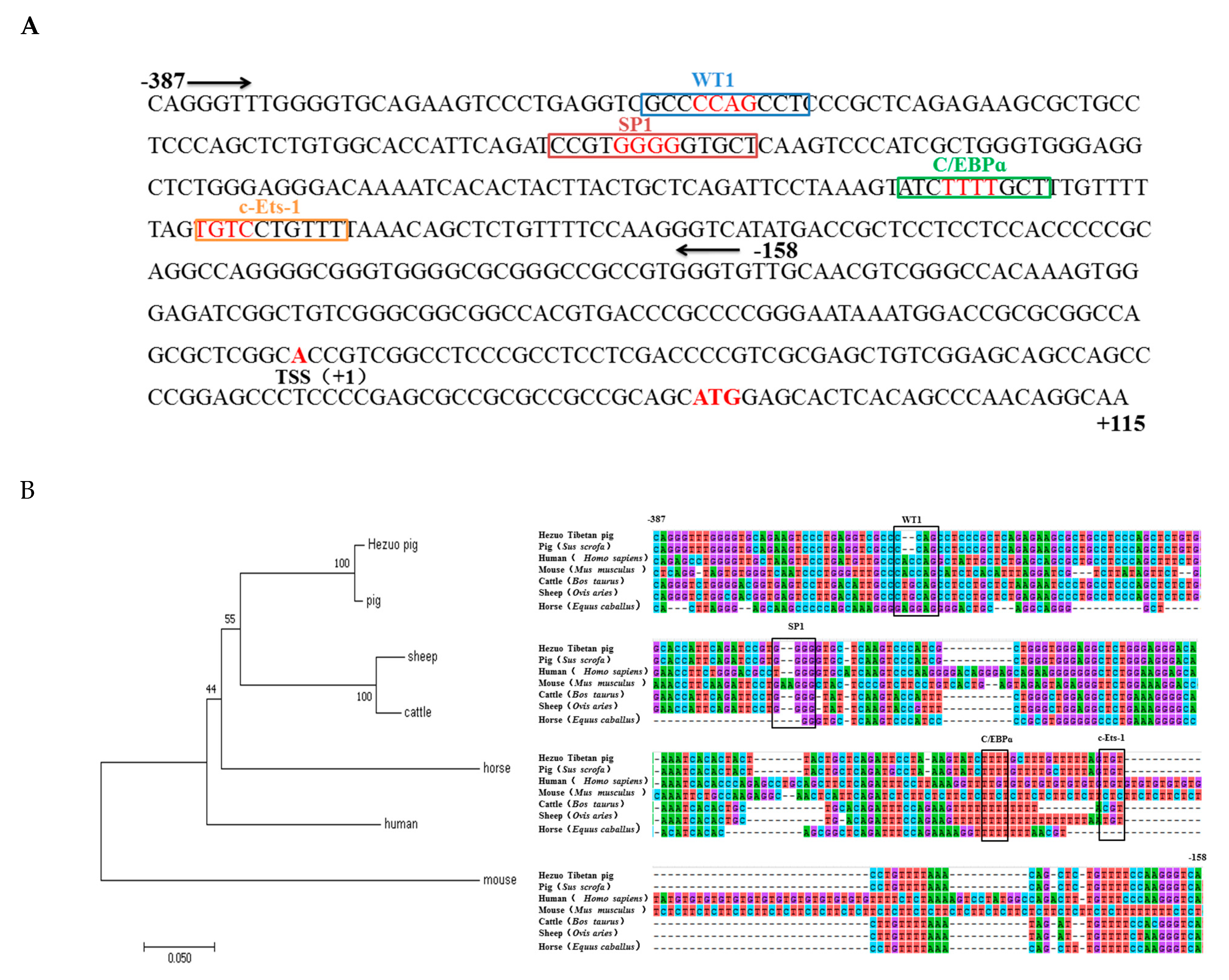
3.2. Promoter Region Cloning and Bioinformatics Analysis
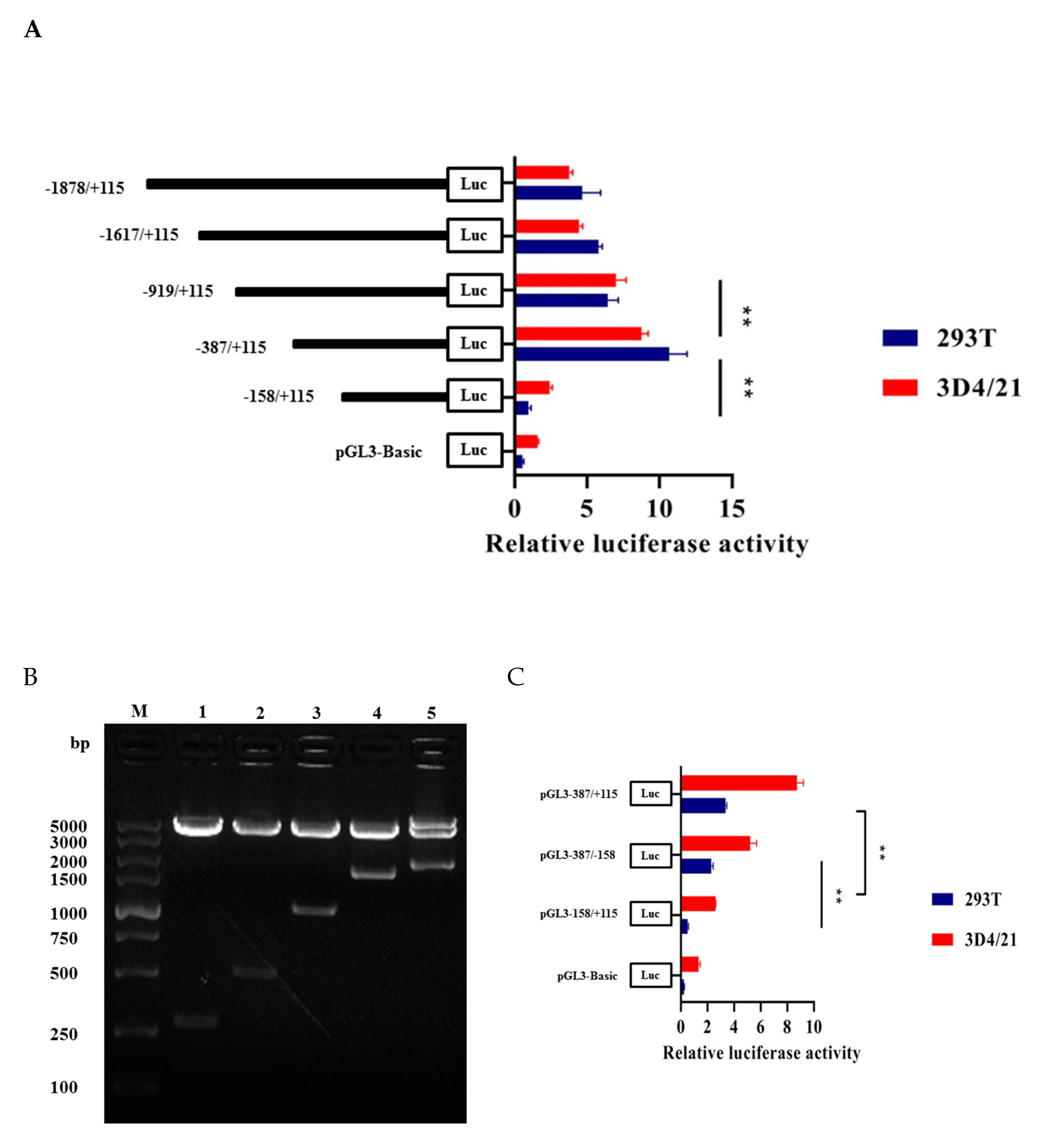
3.3. Promoter Activity Analysis and Core Promoter Region Identification
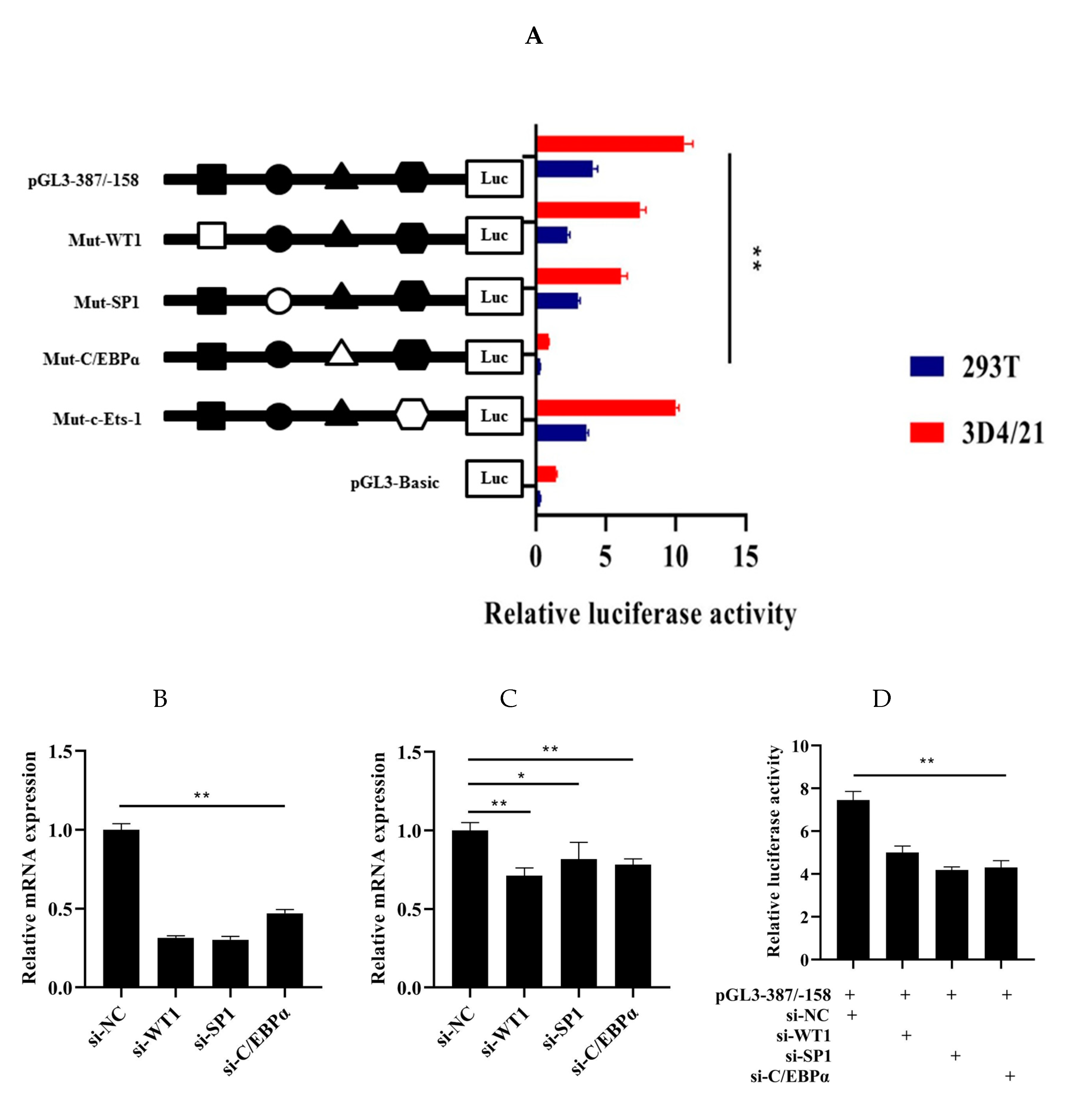
3.4. Roles of WT1, Sp1, C/EBPα, and c-Ets-1 in Transcriptional Regulation of HMOX1 Gene
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gan, M.L.; Shen, L.Y.; Fan, Y.; Guo, Z.X.; Liu, B.; Chen, L.; Tang, G.Q.; Jiang, Y.Z.; Li, X.W.; Zhang, S.H.; et al. High altitude adaptability and meat quality in Tibetan pigs: A reference for local pork processing and genetic improvement. Animals 2019, 9, 1080. [Google Scholar] [CrossRef]
- Zhang, B.; Qiangba, Y.Z.; Shang, P.; Wang, Z.X.; Ma, J.; Wang, L.Y.; Zhang, H. A comprehensive microRNA expression profile related to hypoxia adaptation in the Tibetan pig. PLoS ONE 2015, 10, e0143260. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.F.; Han, X.M.; Huang, C.P.; Zhong, L.; Adeola, A.C.; Irwin, D.M.; Xie, H.B.; Zhang, Y.P. Population genomics analysis revealed origin and high-altitude adaptation of Tibetan pigs. Sci. Rep. 2019, 9, 11463. [Google Scholar] [CrossRef] [PubMed]
- Li, M.Z.; Tian, S.L.; Jin, L.; Zhou, G.Y.; Li, Y.; Zhang, Y.; Wang, T.; Yeung, C.K.; Chen, L.; Ma, J.D.; et al. Genomic analyses identify distinct patterns of selection in domesticated pigs and Tibetan wild boars. Nat. Genet. 2013, 45, 1431–1438. [Google Scholar] [CrossRef] [PubMed]
- Ai, H.S.; Yang, B.; Li, J.; Xie, X.H.; Chen, H.; Ren, J. Population history and genomic signatures for high-altitude adaptation in Tibetan pigs. BMC Genom. 2014, 15, 834. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.Y.; Dong, X.X.; Yang, S.L.; Qian, J.H.; Yang, J.F.; Jiang, Q.; Li, X.R.; Wang, B.; Yan, D.W.; Lu, S.X.; et al. Natural selection on TMPRSS6 associated with the blunted erythropoiesis and improved blood viscosity in Tibetan pigs. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2019, 233, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Otterbein, L.E.; Soares, M.P.; Yamashita, K.; Bach, F.H. Heme oxygenase-1: Unleashing the protective properties of heme. Trends Immunol. 2003, 24, 449–455. [Google Scholar] [CrossRef]
- Kishimoto, Y.; Kondo, K.; Momiyama, Y. The Protective Role of Heme Oxygenase-1 in Atherosclerotic Diseases. Int. J. Mol. Sci. 2019, 20, 3628. [Google Scholar] [CrossRef]
- Lin, H.H.; Chen, Y.H.; Chang, P.F.; Lee, Y.T.; Yet, S.F.; Chau, L.Y. Heme oxygenase-1 promotes neovascularization in ischemic heart by coinduction of VEGF and SDF-1. J. Mol. Cell. Cardiol. 2008, 45, 44–55. [Google Scholar] [CrossRef]
- Singh, S.P.; Greenberg, M.; Glick, Y.; Bellner, L.; Favero, G.; Rezzani, R.; Rodella, L.F.; Agostinucci, K.; Shapiro, J.I.; Abraham, N.G. Adipocyte specific HO-1 gene therapy is effective in antioxidant treatment of insulin resistance and vascular function in an obese mice model. Antioxidants 2020, 9, 40. [Google Scholar] [CrossRef]
- Inguaggiato, P.; Gonzalez-Michaca, L.; Croatt, A.J.; Haggard, J.J.; Alam, J.; Nath, K.A. Cellular overexpression of heme oxygenase-1 up-regulates p21 and confers resistance to apoptosis. Kidney Int. 2001, 60, 2181–2191. [Google Scholar] [CrossRef] [PubMed]
- Takao, M.; Okinaga, T.; Ariyoshi, W.; Iwanaga, K.; Nakamichi, I.; Yoshioka, I.; Tominaga, K.; Nishihara, T. Role of heme oxygenase-1 in inflammatory response induced by mechanical stretch in synovial cells. Inflamm. Res. 2011, 60, 861–867. [Google Scholar] [CrossRef] [PubMed]
- Yachie, A.; Niida, Y.; Wada, T.; Igarashi, N.; Kaneda, H.; Toma, T.; Ohta, K.; Kasahara, Y.; Koizumi, S. Oxidative stress causes enhanced endothelial cell injury in human heme oxygenase-1 deficiency. J. Clin. Investig. 1999, 103, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Brouard, S.; Otterbein, L.E.; Anrather, J.; Tobiasch, E.; Bach, F.H.; Choi, A.M.K.; Soares, M.P. Carbon monoxide generated by heme oxygenase 1 suppresses endothelial cell apoptosis. J. Exp. Med. 2000, 192, 1015–1026. [Google Scholar] [CrossRef]
- Kawamura, K.; Ishikawa, K.; Wada, Y.; Kimura, S.; Matsumoto, H.; Kohro, T.; Itabe, H.; Kodama, T.; Maruyama, Y. Bilirubin from heme oxygenase-1 attenuates vascular endothelial activation and dysfunction. Arter. Thromb Vasc Biol. 2005, 25, 155–160. [Google Scholar] [CrossRef]
- Clark, J.E.; Foresti, R.; Sarathchandra, P.; Kaur, H.; Green, C.J.; Motterlini, R. Heme oxygenase-1-derived bilirubin ameliorates postischemic myocardial dysfunction. Am. J. Physiol. Heart Circ. Physiol. 2000, 278, H643–H651. [Google Scholar] [CrossRef]
- Fujiwara, A.; Hatayama, N.; Matsuura, N.; Yokota, N.; Fukushige, K.; Yakura, T.; Tarumi, S.; Go, T.; Hirai, S.; Naito, M.; et al. High-pressure carbon monoxide and oxygen mixture is effective for lung preservation. Int. J. Mol. Sci. 2019, 20, 2719. [Google Scholar] [CrossRef]
- Rashid, I.; Baisvar, V.S.; Singh, M.; Kumar, R.; Srivastava, P.; Kushwaha, B.; Pathak, A.K. Isolation and characterization of hypoxia inducible heme oxygenase 1 (HMOX1) gene in Labeo rohita. Genomics 2020, 112, 2327–2333. [Google Scholar] [CrossRef]
- Taylor, J.L.; Carraway, M.S.; Piantadosi, C.A. Lung-specific induction of heme oxygenase-1 and hyperoxic lung injury. Am. J. Physiol. 1998, 274, L582–L590. [Google Scholar] [CrossRef]
- Mitchell, P.J.; Tjian, R. Transcriptional regulation in mammalian cells by sequence-specific DNA binding proteins. Science 1989, 245, 371–378. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-△△Ct method. Methods 2000, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Wingender, E.; Dietze, P.; Karas, H.; Knüppel, R. TRANSFAC: A database on transcription factors and their DNA binding sites. Nucleic Acids Res. 1996, 24, 238–241. [Google Scholar] [CrossRef] [PubMed]
- Fornes, O.; Castro-Mondragon, J.A.; Khan, A.; van der Lee, R.; Zhang, X.; Richmond, P.A.; Modi, B.P.; Correard, S.; Gheorghe, M.; Baranašić, D.; et al. JASPAR 2020: Update of the open-access database of transcription factor binding profiles. Nucleic Acids Res. 2019, 48, D87–D92. [Google Scholar] [CrossRef] [PubMed]
- Li, L.C.; Dahiya, R. MethPrimer: Designing primers for methylation PCRs. Bioinformatics 2002, 11, 1427–1431. [Google Scholar] [CrossRef]
- Mccoubrey, W.K.; Ewing, J.F.; Maines, M.D. Human heme oxygenase-2: Characterization and expression of a full-length cDNA and evidence suggesting that the two HO-2 transcripts may differ by choice of polyadenylation signal. Arch. Biochem. Biophys. 1992, 295, 13–20. [Google Scholar] [CrossRef]
- Mccoubrey, W.K.; Huang, T.J.; Maines, M.D. Isolation and Characterization of a cDNA from the Rat Brain that Encodes Hemoprotein Heme Oxygenase-3. Eur. J. Biochem. 1997, 247, 725–732. [Google Scholar] [CrossRef]
- Li, L.; Li, C.M.; Wu, J.; Huang, S.; Wang, G.L. Heat shock protein 32/heme oxygenase-1 protects mouse Sertoli cells from hyperthermia-induced apoptosis by CO activation of sGC signalling pathways. Cell Biol. Int. 2014, 38, 64–71. [Google Scholar] [CrossRef]
- Stuhlmeier, K.M. Activation and regulation of Hsp32 and Hsp70. Eur. J. Biochem. 2000, 267, 1161–1167. [Google Scholar] [CrossRef]
- Taylor, L.; Hillman, A.R.; Midgley, A.W.; Peart, D.J.; Chrismas, B.; McNaughton, L.R. Hypoxia-mediated prior induction of monocyte-expressed HSP72 and HSP32 provides protection to the disturbances to redox balance associated with human sub-maximal aerobic exercise. Amino Acids 2012, 43, 1933–1944. [Google Scholar] [CrossRef]
- Bussolati, B.; Mason, J.C. Dual role of VEGF-induced heme-oxygenase-1 in angiogenesis. Antioxid. Redox Signal. 2006, 8, 1153–1163. [Google Scholar] [CrossRef] [PubMed]
- Dulak, J.; Loboda, A.; Zagórska, A.; Józkowicz, A. Complex role of heme oxygenase-1 in angiogenesis. Antioxid. Redox Signal. 2004, 6, 858–866. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Zhong, X.P.; Qiao, Z.X.; Gui, J.F. Inductive transcription and protective role of fish heme oxygenase-1 under hypoxic stress. J. Exp. Biol. 2008, 211, 2700–2706. [Google Scholar] [CrossRef]
- Gou, X.; Li, N.; Lian, L.S.; Yan, D.W.; Zhang, H.; Wei, Z.H.; Wu, C.X. Hypoxic adaptations of hemoglobin in Tibetan chick embryo: High oxygen-affinity mutation and selective expression. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2007, 147, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Kovtunovych, G.; Eckhaus, M.A.; Ghosh, M.C.; Ollivierrewilson, H.; Rouault, T.A. Dysfunction of the heme recycling system in heme oxygenase 1–deficient mice: Effects on macrophage viability and tissue iron distribution. Blood 2010, 116, 6054–6062. [Google Scholar] [CrossRef] [PubMed]
- Park, S.J.; Cho, B.; Koo, O.J.; Kim, H.; Kang, J.T.; Hurh, S.; Kim, S.J.; Yeom, H.J.; Moon, J.; Lee, E.M.; et al. Production and characterization of soluble human TNFRI-Fc and human HO-1 (HMOX1) transgenic pigs by using the F2A peptide. Transgenic Res. 2014, 23, 407–419. [Google Scholar] [CrossRef] [PubMed]
- Alam, J. Functional analysis of the heme oxygenase-1 gene promoter. Curr. Protoc. Toxicol. 2000, 6. [Google Scholar] [CrossRef] [PubMed]
- Elbashir, S.M.; Harborth, J.; Lendeckel, W.; Yalcin, A.; Weber, K.; Tuschl, T. Duplexes of 21-nucleotide RNAs mediate RNA interference in cultured mammalian cells. Nature 2001, 411, 494–498. [Google Scholar] [CrossRef]
- Scholz, H.; Wagner, K.D.; Wagner, N. Role of the Wilms’ tumour transcription factor, Wt1, in blood vessel formation. Pflug. Arch. 2009, 458, 315–323. [Google Scholar] [CrossRef]
- Panuzzo, C.; Signorino, E.; Calabrese, C.; Ali, M.S.; Petiti, J.; Bracco, E.; Cilloni, D. Landscape of Tumor Suppressor Mutations in Acute Myeloid Leukemia. J. Clin. Med. 2020, 9, 802. [Google Scholar] [CrossRef]
- Wang, S.J.; Wang, C.; Li, T.; Wang, W.Q.; Hao, Q.Q.; Xie, X.S.; Wang, D.M.; Jiang, Z.X.; Liu, Y.F. WT1 overexpression predicted good outcomes in adult B-cell acute lymphoblastic leukemia patients receiving chemotherapy. Hematology 2020, 25, 118–124. [Google Scholar] [CrossRef]
- Bouwman, P.; Philipsen, S. Regulation of the activity of Sp1-related transcription factors. Mol. Cell. Endocrinol. 2002, 195, 27–38. [Google Scholar] [CrossRef]
- Ryu, H.; Lee, J.; Zaman, K.; Kubilis, J.; Ferrante, R.J.; Ross, B.D.; Neve, R.; Ratan, R.R. Sp1 and Sp3 are oxidative stress-inducible, antideath transcription factors in cortical neurons. J. Neurosci. 2003, 23, 3597–3606. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.H.; Lu, S.J.; Wu, P.; Kong, L.C.; Ning, C.; Li, H.Y. Molecular mechanism whereby paraoxonase-2 regulates coagulation activation through endothelial tissue factor in rat haemorrhagic shock model. Int. Wound J. 2020, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Ding, A.; Bian, Y.Y.; Zhang, Z.H. SP1/TGF-β1/SMAD2 pathway is involved in angiogenesis during osteogenesis. Mol. Med. Rep. 2020, 21, 1581–1589. [Google Scholar] [CrossRef] [PubMed]
- Deshane, J.; Kim, J.; Bolisetty, S.; Hock, T.D.; Hill-Kapturczak, N.; Agarwal, A. Sp1 regulates chromatin looping between an intronic enhancer and distal promoter of the human heme oxygenase-1 gene in renal cells. J. Biol. Chem. 2010, 285, 16476–16486. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.Y.; Khan, R.; Raza, S.H.A.; Li, A.N.; Zhang, Y.; Liang, C.C.; Yang, W.C.; Wu, S.; Zan, L. Molecular characterization of ABHD5 gene promoter in intramuscular preadipocytes of Qinchuan cattle: Roles of Evi1 and C/EBPα. Gene 2019, 690, 38–47. [Google Scholar] [CrossRef]
- Reckzeh, K.; Cammenga, J. Molecular mechanisms underlying deregulation of C/EBPα in acute myeloid leukemia. Int. J. Hematol. 2010, 91, 557–568. [Google Scholar] [CrossRef]
- Liu, X.H.; Jia, X.E.; Yuan, H.; Ma, K.; Chen, Y.; Jin, Y.; Deng, M.; Pan, W.J.; Chen, S.J.; Chen, Z.; et al. DNA methyltransferase 1 functions through C/ebpa to maintain hematopoietic stem and progenitor cells in zebrafish. J. Hematol. Oncol. 2015, 8, 15. [Google Scholar] [CrossRef]
- Hung, C.C.; Liu, X.L.; Kwon, M.Y.; Kang, Y.H.; Chung, S.W.; Perrella, M.A. Regulation of heme oxygenase-1 gene by peptidoglycan involves the interaction of Elk-1 and C/EBPα to increase expression. Am. J. Physiol. Lung Cell Mol. Physiol. 2010, 298, 870–879. [Google Scholar] [CrossRef][Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Primer Sequence(5’–3’) | Tm (°C) | Length | Region |
|---|---|---|---|---|
| HMOX1-P | F: GCCTCGTCCTTCTAAAGTCCC | 56 | 2125 bp | −1911/−1892 |
| R: CGCCAAAGCCCAAGTGACTG | +214/+195 | |||
| HMOX1-RT | F: GACATGGCCTTCTGGTATGGG | 60 | 141 bp | 348–368 |
| R: CATGTAGCGGGTGTAGGCGT | 488–469 | |||
| WT1-RT | F: GGTGTCTTCAGGGGCATTCA | 60 | 102 bp | 1127–1146 |
| R: ACACATGAAGGGGCGTTTCT | 1228–1209 | |||
| Sp1-RT | F: TGTCTCTGGTGGGCAGTATG | 60 | 133 bp | 601–620 |
| R: TTGCCCATCAACCGTCTGG | 733–715 | |||
| C/EBPα-RT | F: GGCAAAGCCAAGAAGTCGGT | 60 | 124 bp | 976–995 |
| R: TCTGTTGAGTCTCCACGTTGC | 1099–1079 | |||
| β-actin-RT | F: ATATTGCTGCGCTCGTGGT | 60 | 148 bp | 142–160 |
| R: TAGGAGTCCTTCTGGCCCAT | 289–270 | |||
| HMOX1-P1 | F: CTAGCTAGCTATGACCGCTCCTCCTCCAC | 60 | 273 bp | −158/+115 |
| HMOX1-P2 | F: CTAGCTAGCCAGGGTTTGGGGTGCAGAAG | 60 | 502 bp | −387/+115 |
| HMOX1-P3 | F: CTAGCTAGCGCTCCCCTAGTAGTAACCTGC | 64 | 1034 bp | −919/+115 |
| HMOX1-P4 | F: CTAGCTAGCGGGCACTCGATACCTTGGTT | 60 | 1732 bp | −1617/+115 |
| HMOX1-P5 | F: CTAGCTAGCCAGCAACCCCCAAGTCTCT | 62 | 1993 bp | −1878/+115 |
| HMOX1-R | R: CCGCTCGAGTTGCCTGTTGGGCTGTGAG | |||
| WT1-m | F: GAAGTCCCTGAGGTCGCCGGTCCCTCCCGCTCAGAGAAGC | 229 bp | −387/−158 | |
| R: GCTTCTCTGAGCCAGCGGCCAGGGAGGGCTCAGGGACTTC | ||||
| Sp1-m | F: GGCACCATTCAGATCCGTAAAAGTGCTCAAGTCCCATCGC | 229 bp | −387/−158 | |
| R: GCGATGGGACTTAGGCATTTTCACGAGCTGAATGGTGCC | ||||
| C/EBPα-m | F: TCAGATTCCTAAAGTATCAAAAGCTTTGTTTTTAGTGTCC | 229 bp | −387/−158 | |
| R: GGACACTAAAAATCATAGTTTTCGAAACTTAGGAATCTGA | ||||
| c-Ets-1-m | F: CTTTTGCTTTGTTTTTAGACAGCTGTTTTAAACAGCTCTG | 229 bp | −387/−158 | |
| R: CAGAGCTGTTTAAAAATCTGTCGACAAAACAAAGVAAAAG | ||||
| si-WT1 | F: GGGCUGCAAUAAGAGAUAUTT | |||
| R: AUAUCUCUUAUUGCAGCCCTT | ||||
| si-Sp1 | F: GCGGAUCUGCAGUCCAUUATT | |||
| R: UAAUGGACUGCAGAUCCGCTT | ||||
| si-C/EBPα | F: ACGAGACGUCCAUCGACAUTT | |||
| R: AUGUCGAUGGACGUCUCGUTT | ||||
| si-NC | F: UUCUCCGAACGUGUCACGUTT | |||
| R: UUAACUCAUCGCUUCUUGCTT |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, W.; Yang, Q.; Xie, K.; Wang, P.; Luo, R.; Yan, Z.; Gao, X.; Zhang, B.; Huang, X.; Gun, S. Transcriptional Regulation of HMOX1 Gene in Hezuo Tibetan Pigs: Roles of WT1, Sp1, and C/EBPα. Genes 2020, 11, 352. https://doi.org/10.3390/genes11040352
Wang W, Yang Q, Xie K, Wang P, Luo R, Yan Z, Gao X, Zhang B, Huang X, Gun S. Transcriptional Regulation of HMOX1 Gene in Hezuo Tibetan Pigs: Roles of WT1, Sp1, and C/EBPα. Genes. 2020; 11(4):352. https://doi.org/10.3390/genes11040352
Chicago/Turabian StyleWang, Wei, Qiaoli Yang, Kaihui Xie, Pengfei Wang, Ruirui Luo, Zunqiang Yan, Xiaoli Gao, Bo Zhang, Xiaoyu Huang, and Shuangbao Gun. 2020. "Transcriptional Regulation of HMOX1 Gene in Hezuo Tibetan Pigs: Roles of WT1, Sp1, and C/EBPα" Genes 11, no. 4: 352. https://doi.org/10.3390/genes11040352
APA StyleWang, W., Yang, Q., Xie, K., Wang, P., Luo, R., Yan, Z., Gao, X., Zhang, B., Huang, X., & Gun, S. (2020). Transcriptional Regulation of HMOX1 Gene in Hezuo Tibetan Pigs: Roles of WT1, Sp1, and C/EBPα. Genes, 11(4), 352. https://doi.org/10.3390/genes11040352