

Isolation and Characterization of Articular Cartilage-Derived Cells Obtained by Arthroscopic Cartilage Biopsy from Non-Osteoarthritic Patients

 , , ,
, , ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Samples
2.2. Enzymatic Digestion
2.3. Explant Cell Migration
2.4. Cell Characterization
2.5. Protein Detection by Indirect Immunofluorescence
2.6. Gene Expression
2.7. Evaluation of Genes Involved in Cell Differentiation
2.8. Data Analysis
3. Results
3.1. Cell Morphology
3.2. Cell Characterization
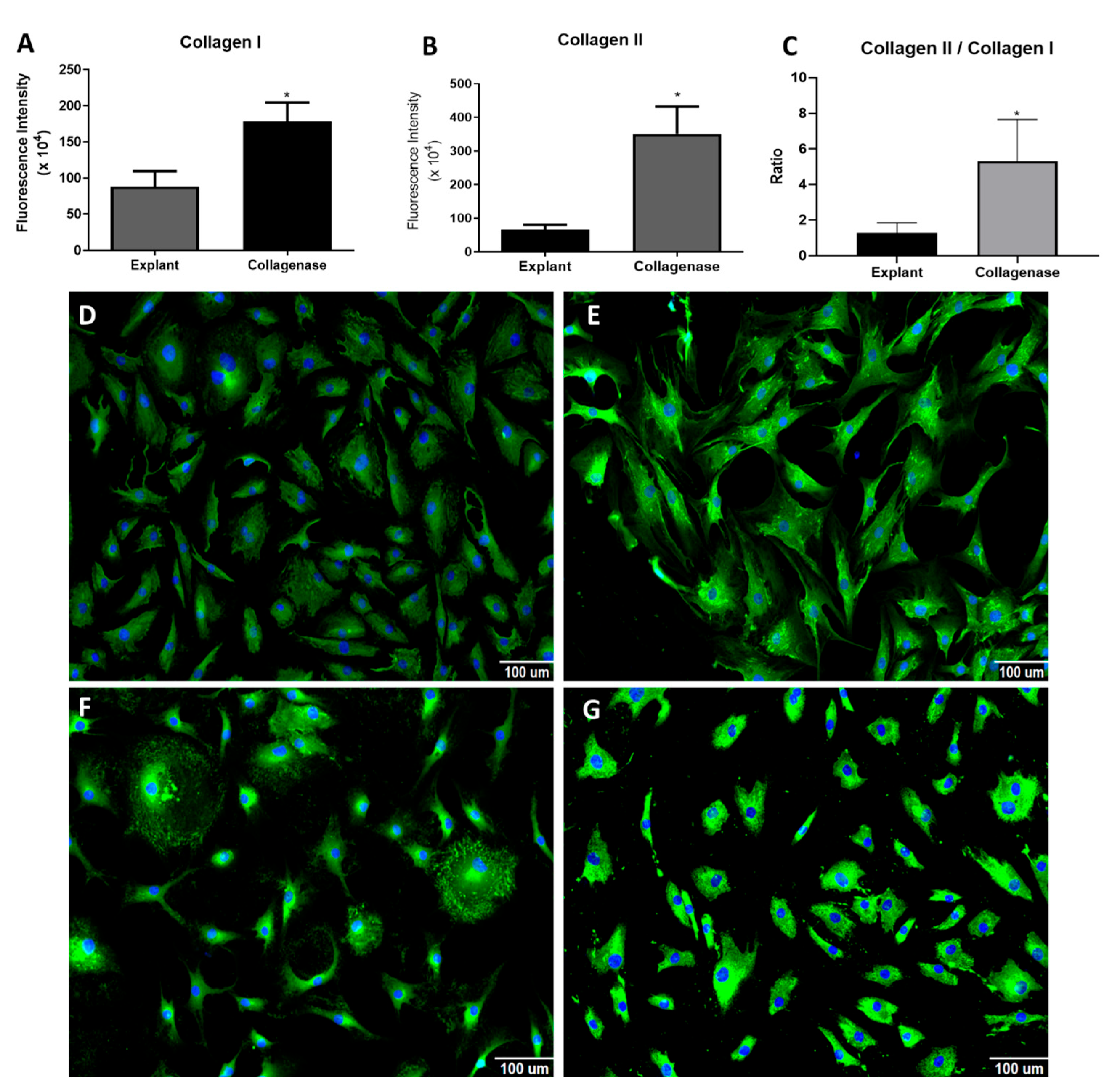
3.3. Analysis of Proteins Analysis: Collagen I and II, RUNX2, SOX9 and PPARγ
3.4. Gene Expression of Cell Differentiation Markers
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hinckel, B.B.; Thomas, D.; Vellios, E.E.; Hancock, K.J.; Calcei, J.G.; Sherman, S.L.; Eliasberg, C.D.; Fernandes, T.L.; Farr, J.; Lattermann, C.; et al. Algorithm for Treatment of Focal Cartilage Defects of the Knee: Classic and New Procedures. Cartilage 2021, 13, 473S–495S. [Google Scholar] [CrossRef] [PubMed]
- Makarczyk, M.J. Cell Therapy Approaches for Articular Cartilage Regeneration. Organogenesis 2023, 19, 2278235. [Google Scholar] [CrossRef] [PubMed]
- Giglio, P.N.; Lizier, N.F.; Levy, D.; Sobrado, M.F.; Gobbi, R.G.; Pécora, J.R.; Bydlowski, D.P.; Demange, M.K. Autologous chondrocyte implantation in Brazil. Acta Ortop. Bras. 2020, 28, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Vinod, E.; Parameswaran, R.; Ramasamy, B.; Kachroo, U. Pondering the Potential of Hyaline Cartilage-Derived Chondroprogenitors for Tissue Regeneration: A Systematic Review. Cartilage 2020, 13, 34S–52S. [Google Scholar] [CrossRef]
- Matta, C.; Boocock, D.J.; Fellows, C.R.; Miosge, N.; Dixon, J.E.; Liddell, S.; Smith, J.; Mobasheri, A. Molecular phenotyping of the surfaceome of migratory chondroprogenitors and mesenchymal stem cells using biotinylation, glycocapture and quantitative LC-MS/MS proteomic analysis. Sci. Rep. 2019, 9, 9018. [Google Scholar] [CrossRef] [PubMed]
- Batschkus, S.; Atanassov, I.; Lenz, C.; Meyer-Marcotty, P.; Cingöz, G.; Kirschneck, C.; Urlaub, H.; Miosge, N. Mapping the secretome of human chondrogenic progenitor cells with mass spectrometry. Ann. Anat. 2017, 212, 4–10. [Google Scholar] [CrossRef]
- Nguyen, V.T.; Cancedda, R.; Descalzi, F. Platelet lysate activates quiescent cell proliferation and reprogramming in human articular cartilage: Involvement of hypoxia inducible factor 1. J. Tissue Eng. Regen. Med. 2018, 12, e1691–e1703. [Google Scholar] [CrossRef]
- Janssen, J.N.; Batschkus, S.; Schimmel, S.; Bode, C.; Schminke, B.; Miosge, N. The Influence of TGF-β3, EGF, and BGN on SOX9 and RUNX2 Expression in Human Chondrogenic Progenitor Cells. J. Histochem. Cytochem. 2019, 67, 117–127. [Google Scholar] [CrossRef] [PubMed]
- Wagner, G.; Lehmann, C.; Bode, C.; Miosge, N.; Schubert, A. High Mobility Group Box 1 Protein in Osteoarthritic Knee Tissue and Chondrogenic Progenitor Cells: An Ex Vivo and In Vitro Study. Cartilage 2021, 12, 484–495. [Google Scholar] [CrossRef]
- Wang, Y.-X.; Zhao, Z.-D.; Wang, Q.; Li, Z.-L.; Huang, Y.; Zhao, S.; Hu, W.; Liang, J.-W.; Li, P.-L.; Wang, H.; et al. Biological potential alterations of migratory chondrogenic progenitor cells during knee osteoarthritic progression. Arthritis Res. Ther. 2020, 22, 62. [Google Scholar] [CrossRef]
- Wang, K.; Li, J.; Li, Z.; Wang, B.; Qin, Y.; Zhang, N.; Zhang, H.; Su, X.; Wang, Y.; Zhu, H. Chondrogenic Progenitor Cells Exhibit Superiority over Mesenchymal Stem Cells and Chondrocytes in Platelet-Rich Plasma Scaffold-Based Cartilage Regeneration. Am. J. Sports Med. 2019, 47, 2200–2215. [Google Scholar] [CrossRef] [PubMed]
- Levy, D.; Calllera, A.F.; Moreira, A.R.; Tibério, I.d.F.L.C.; Giglio, P.N.; Demange, M.K.; Bydlowski, S.P.; Lopes, F.D.T.Q.D.S. TNF-Alpha Inhibitor Prevents Cigarette Smoke Extract-Induced Cell Death in Osteoarthritis-Derived Chondrocytes in Culture. Cells 2025, 14, 489. [Google Scholar] [CrossRef]
- Janz, F.d.L.; Debes, A.d.A.; Cavaglieri, R.d.C.; Duarte, S.A.; Romão, C.M.; Morón, A.F.; Zugaib, M.; Bydlowski, S.P. Evaluation of distinct freezing methods and cryoprotectants for human amniotic fluid stem cells cryopreservation. J. Biomed. Biotechnol. 2012, 14, 649353. [Google Scholar] [CrossRef]
- Welton, K.L.; Logterman, S.; Bartley, J.H.; Vidal, A.F.; McCarty, E.C. Knee Cartilage Repair and Restoration: Common Problems and Solutions. Clin. Sports Med. 2018, 37, 307–330. [Google Scholar] [CrossRef] [PubMed]
- Seol, D.; McCabe, D.J.; Choe, H.; Zheng, H.J.; Yu, Y.; Jang, K.; Walter, M.W.; Lehman, A.D.; Ding, L.; Buckwalter, J.A.; et al. Chondrogenic progenitor cells respond to cartilage injury. Arthritis Rheum. 2012, 64, 3626–3637. [Google Scholar] [CrossRef]
- Wang, S.; Zhou, C.; Zheng, H.; Zhang, Z.; Mei, Y.; Martin, J.A. Chondrogenic progenitor cells promote vascular endothelial growth factor expression through stromal-derived factor-1. Osteoarthr. Cartil. 2017, 25, 742–749. [Google Scholar] [CrossRef]
- Zhou, C.; Zheng, H.; Buckwalter, J.A.; Martin, J.A. Enhanced phagocytic capacity endows chondrogenic progenitor cells with a novel scavenger function within injured cartilage. Osteoarthr. Cartil. 2016, 24, 1648–1655. [Google Scholar] [CrossRef]
- Carluccio, S.; Martinelli, D.; Palamà, M.E.F.; Pereira, R.C.; Benelli, R.; Guijarro, A.; Cancedda, R.; Gentili, C. Progenitor Cells Activated by Platelet Lysate in Human Articular Cartilage as a Tool for Future Cartilage Engineering and Reparative Strategies. Cells 2020, 9, 1052. [Google Scholar] [CrossRef] [PubMed]
- de la Fuente, R.; Abad, J.L.; García-Castro, J.; Fernández-Miguel, G.; Petriz, J.; Rubio, D.; Vicario-Abejón, C.; Guillén, P.; González, M.A.; Bernad, A. Dedifferentiated adult articular chondrocytes: A population of human multipotent primitive cells. Exp. Cell Res. 2004, 297, 313–328. [Google Scholar] [CrossRef]
- Benz, K.; Stippich, C.; Freudigmann, C.; Mollenhauer, J.A.; Aicher, W.K. Maintenance of “stem cell” features of cartilage cell sub-populations during in vitro propagation. J. Transl. Med. 2013, 11, 27. [Google Scholar] [CrossRef]
- Sandrasaigaran, P.; Algraittee, S.J.R.; Ahmad, A.R.; Vidyadaran, S.; Ramasamy, R. Characterisation and immunosuppressive activity of human cartilage-derived mesenchymal stem cells. Cytotechnology 2018, 70, 1037–1050. [Google Scholar] [CrossRef] [PubMed]
- De Luca, P.; Kouroupis, D.; Viganò, M.; Perucca-Orfei, C.; Kaplan, L.; Zagra, L.; de Girolamo, L.; Correa, D.; Colombini, A. Human Diseased Articular Cartilage Contains a Mesenchymal Stem Cell-Like Population of Chondroprogenitors with Strong Immunomodulatory Responses. J. Clin. Med. Res. 2019, 8, 423. [Google Scholar] [CrossRef] [PubMed]
- Barbero, A.; Ploegert, S.; Heberer, M.; Martin, I. Plasticity of clonal populations of dedifferentiated adult human articular chondrocytes. Arthritis Rheum. 2003, 48, 1315–1325. [Google Scholar] [CrossRef] [PubMed]
- Dell’Accio, F.; De Bari, C.; Luyten, F.P. Microenvironment and phenotypic stability specify tissue formation by human articular cartilage-derived cells in vivo. Exp. Cell Res. 2003, 287, 16–27. [Google Scholar] [CrossRef]
- Tallheden, T.; Dennis, J.E.; Lennon, D.P.; Sjögren-Jansson, E.; Caplan, A.I.; Lindahl, A. Phenotypic plasticity of human articular chondrocytes. J. Bone Jt. Surg. Am. 2003, 85 (Suppl. S2), 93–100. [Google Scholar] [CrossRef]
- Thornemo, M.; Tallheden, T.; Jansson, E.S.; Larsson, A.; Lövstedt, K.; Nannmark, U.; Brittberg, M.; Lindahl, A. Clonal populations of chondrocytes with progenitor properties identified within human articular cartilage. Cells Tissues Organs 2005, 180, 141–150. [Google Scholar] [CrossRef]
- Diaz-Romero, J.; Nesic, D.; Grogan, S.P.; Heini, P.; Mainil-Varlet, P. Immunophenotypic changes of human articular chondrocytes during monolayer culture reflect bona fide dedifferentiation rather than amplification of progenitor cells. J. Cell Physiol. 2008, 214, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Komori, T. Runx2, an inducer of osteoblast and chondrocyte differentiation. Histochem. Cell Biol. 2018, 149, 313–323. [Google Scholar] [CrossRef]
- Kamekura, S.; Kawasaki, Y.; Hoshi, K.; Shimoaka, T.; Chikuda, H.; Maruyama, Z.; Komori, T.; Sato, S.; Takeda, S.; Karsenty, G.; et al. Contribution of runt-related transcription factor 2 to the pathogenesis of osteoarthritis in mice after induction of knee joint instability. Arthritis Rheum. 2006, 54, 2462–2470. [Google Scholar] [CrossRef]
- Stokes, D.G.; Liu, G.; Dharmavaram, R.; Hawkins, D.; Piera-Velazquez, S.; Jimenez, S.A. Regulation of type-II collagen gene expression during human chondrocyte de-differentiation and recovery of chondrocyte-specific phenotype in culture involves Sry-type high-mobility-group box (SOX) transcription factors. Biochem. J. 2001, 360, 461–470. [Google Scholar] [CrossRef]
- Brew, C.J.; Andrew, J.G.; Boot-Handford, R.; Hardingham, T.E. Late osteoarthritic cartilage shows down regulation of SOX9 and aggrecan expression but little evidence of chondrocyte hypertrophy. Trans. Orthop. Res. Soc. 2004, 50, 938. [Google Scholar]
- Clarke, S.L.; Robinson, C.E.; Gimble, J.M. CAAT/Enhancer Binding Proteins Directly Modulate Transcription from the Peroxisome Proliferator- Activated Receptor γ2 Promoter. Biochem. Biophys. Res. Commun. 1997, 240, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Moseti, D.; Regassa, A.; Kim, W.-K. Molecular Regulation of Adipogenesis and Potential Anti-Adipogenic Bioactive Molecules. Int. J. Mol. Sci. 2016, 17, 124. [Google Scholar] [CrossRef]
- Marlovits, S.; Hombauer, M.; Truppe, M.; Vècsei, V.; Schlegel, W. Changes in the ratio of type-I and type-II collagen expression during monolayer culture of human chondrocytes. J. Bone Jt. Surg. Br. 2004, 86, 286–295. [Google Scholar] [CrossRef] [PubMed]
- Hamada, T.; Sakai, T.; Hiraiwa, H.; Nakashima, M.; Ono, Y.; Mitsuyama, H.; Ishiguro, N. Surface markers and gene expression to characterize the differentiation of monolayer expanded human articular chondrocytes. Nagoya J. Med. Sci. 2013, 75, 101–111. [Google Scholar]
- Suh, J.-K.; Scherping, S.; Mardi, T.; Richard Steadman, J.; Woo, S.L.Y. Basic science of articular cartilage injury and repair. Oper. Tech. Sports Med. 1995, 3, 78–86. [Google Scholar] [CrossRef]
- Sakata, R.; Iwakura, T.; Reddi, A.H. Regeneration of Articular Cartilage Surface: Morphogens, Cells, and Extracellular Matrix Scaffolds. Tissue Eng. Part B Rev. 2015, 21, 461–473. [Google Scholar] [CrossRef]
- Carballo, C.B.; Nakagawa, Y.; Sekiya, I.; Rodeo, S.A. Basic Science of Articular Cartilage. Clin. Sports Med. 2017, 36, 413–425. [Google Scholar] [CrossRef]
- Kisiday, J.D. Expansion of chondrocytes for cartilage tissue engineering: A review of chondrocyte dedifferentiation and redifferentiation as a function of growth in expansion culture. Regen. Med. Front. 2019, 2, e200002. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Explant | Collagenase | |
|---|---|---|
| CD14 | 0.6 ± 0.06 | 1.2 ± 0.04 |
| CD29 | 96.6 ± 0.22 | 95.4 ± 0.53 |
| CD34 | 0.9 ± 0.10 | 1.3 ± 0.01 |
| CD45 | 3.8 ± 0.07 | 2.1 ± 0.02 |
| CD80 | 3.4 ± 0.05 | 2.9 ± 0.07 |
| CD90 | 99.5 ± 0.16 | 98.7 ± 0.22 |
| CD105 | 97.4 ± 0.36 | 97.8 ± 0.48 |
| CD117 | 1.2 ± 0.04 | 0.3 ± 0.01 |
| HLA-DR | 1.8 ± 0.05 | 0.5 ± 0.02 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giglio, P.N.; Levy, D.; Favaron, P.O.; Melo, L.d.P.; Reichert, C.O.; de Freitas, F.A.; Sampaio Silva, J.; Teodoro, W.P.R.; Bydlowski, S.P.; Demange, M.K. Isolation and Characterization of Articular Cartilage-Derived Cells Obtained by Arthroscopic Cartilage Biopsy from Non-Osteoarthritic Patients. Cells 2025, 14, 830. https://doi.org/10.3390/cells14110830
Giglio PN, Levy D, Favaron PO, Melo LdP, Reichert CO, de Freitas FA, Sampaio Silva J, Teodoro WPR, Bydlowski SP, Demange MK. Isolation and Characterization of Articular Cartilage-Derived Cells Obtained by Arthroscopic Cartilage Biopsy from Non-Osteoarthritic Patients. Cells. 2025; 14(11):830. https://doi.org/10.3390/cells14110830
Chicago/Turabian StyleGiglio, Pedro Nogueira, Débora Levy, Phelipe Oliveira Favaron, Lucas da Ponte Melo, Cadiele Oliana Reichert, Fábio Alessandro de Freitas, Juliana Sampaio Silva, Walcy Paganelli Rosolia Teodoro, Sérgio Paulo Bydlowski, and Marco Kawamura Demange. 2025. "Isolation and Characterization of Articular Cartilage-Derived Cells Obtained by Arthroscopic Cartilage Biopsy from Non-Osteoarthritic Patients" Cells 14, no. 11: 830. https://doi.org/10.3390/cells14110830
APA StyleGiglio, P. N., Levy, D., Favaron, P. O., Melo, L. d. P., Reichert, C. O., de Freitas, F. A., Sampaio Silva, J., Teodoro, W. P. R., Bydlowski, S. P., & Demange, M. K. (2025). Isolation and Characterization of Articular Cartilage-Derived Cells Obtained by Arthroscopic Cartilage Biopsy from Non-Osteoarthritic Patients. Cells, 14(11), 830. https://doi.org/10.3390/cells14110830