
Simulated Microgravity Affects Pro-Resolving Properties of Primary Human Monocytes

 ,
,  , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Peripheral Blood Cells Isolation and RCCS-Simulated Microgravity
2.3. RNA Extraction and qRT-PCR
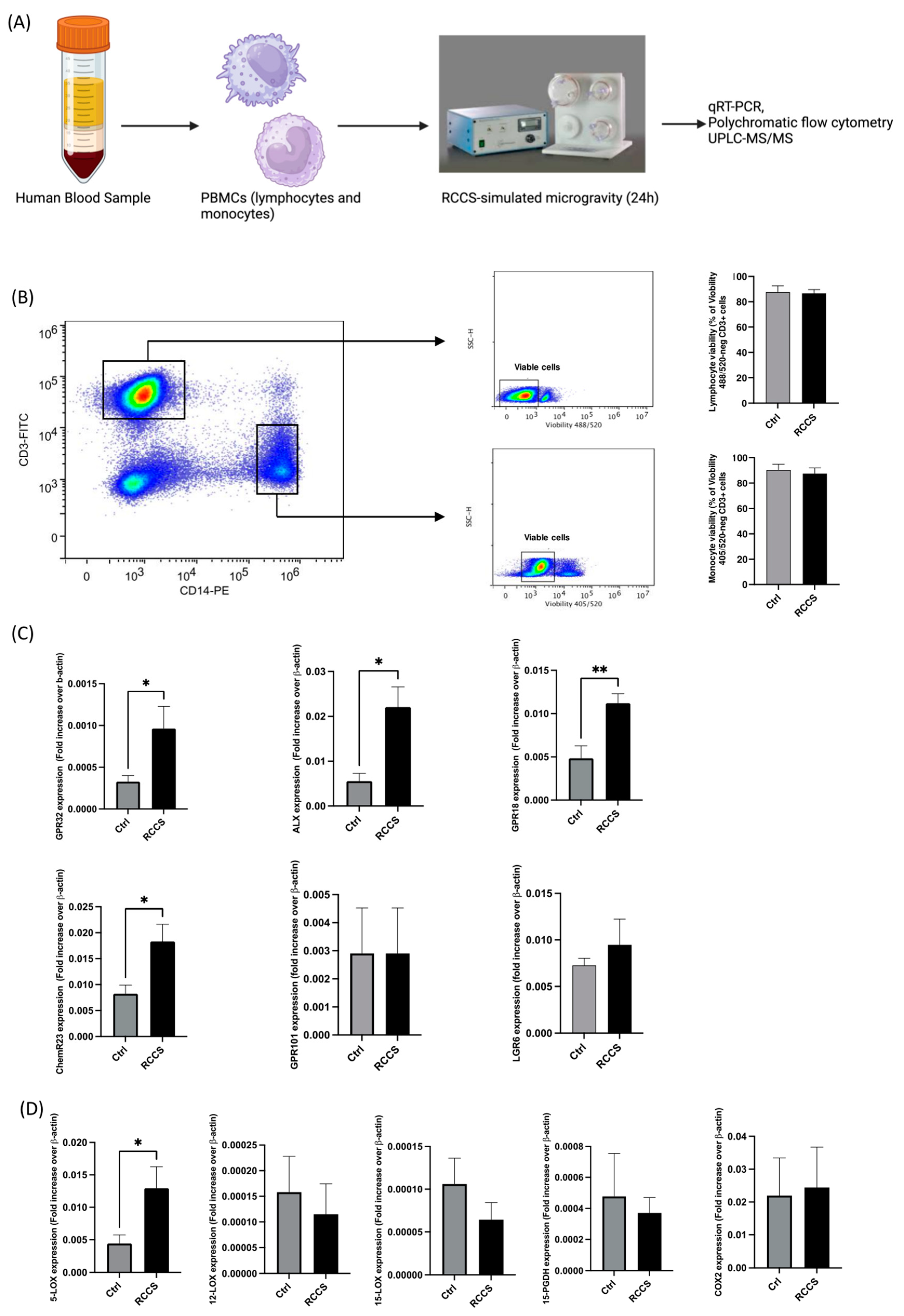
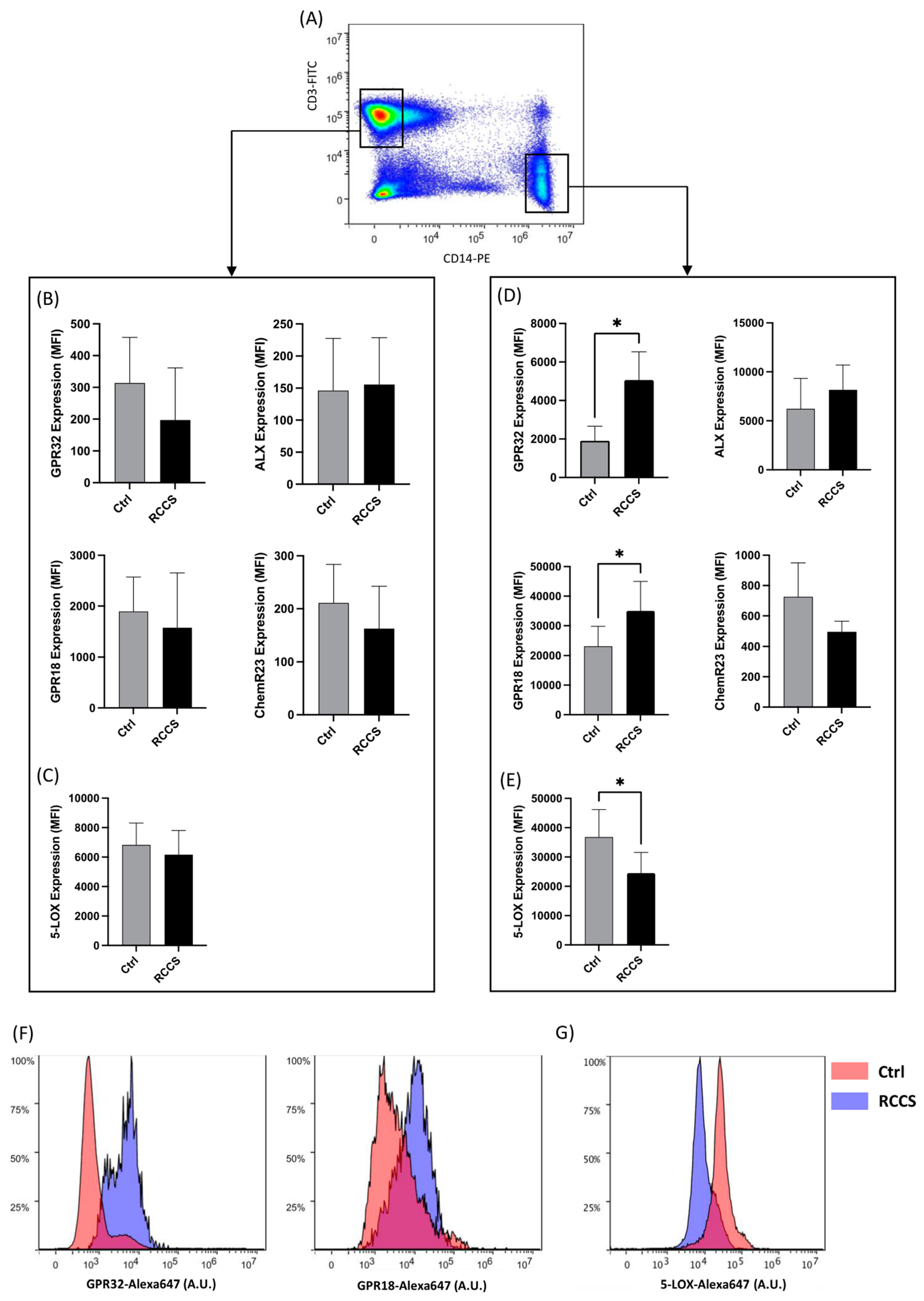
2.4. Flow Cytometry
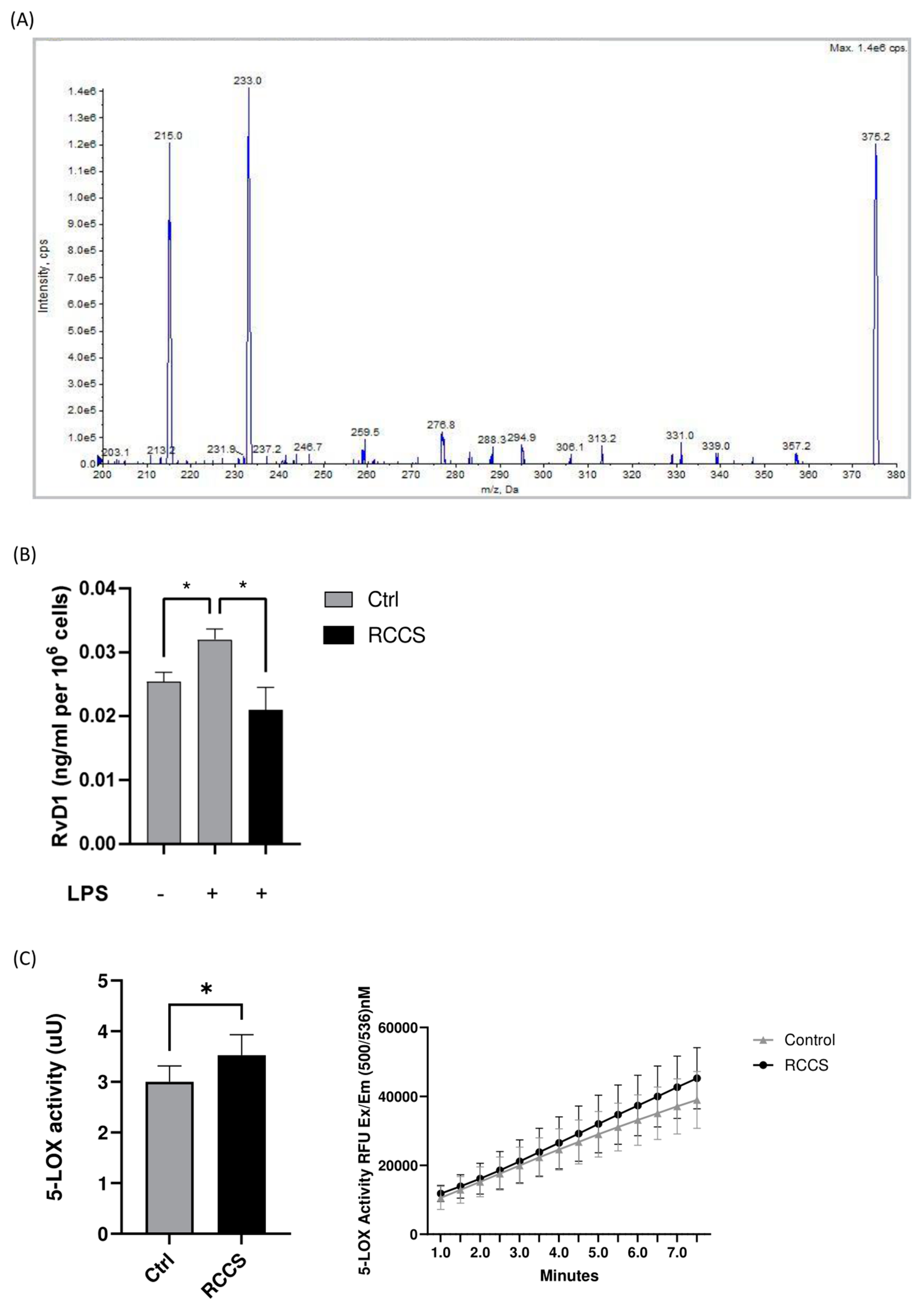
2.5. UPLC-MS/MS Analysis of SPMs
2.6. Statistical Analysis
3. Results
3.1. Simulated Microgravity Rearranges the Expression of Pro-Resolving Genes in PBMCs
3.2. Simulated Microgravity Changes Protein Expression of SPM Receptors and Enzymes in Monocytes
3.3. SPM Biosynthesis Changes in PBMCs Exposed to Simulated Microgravity
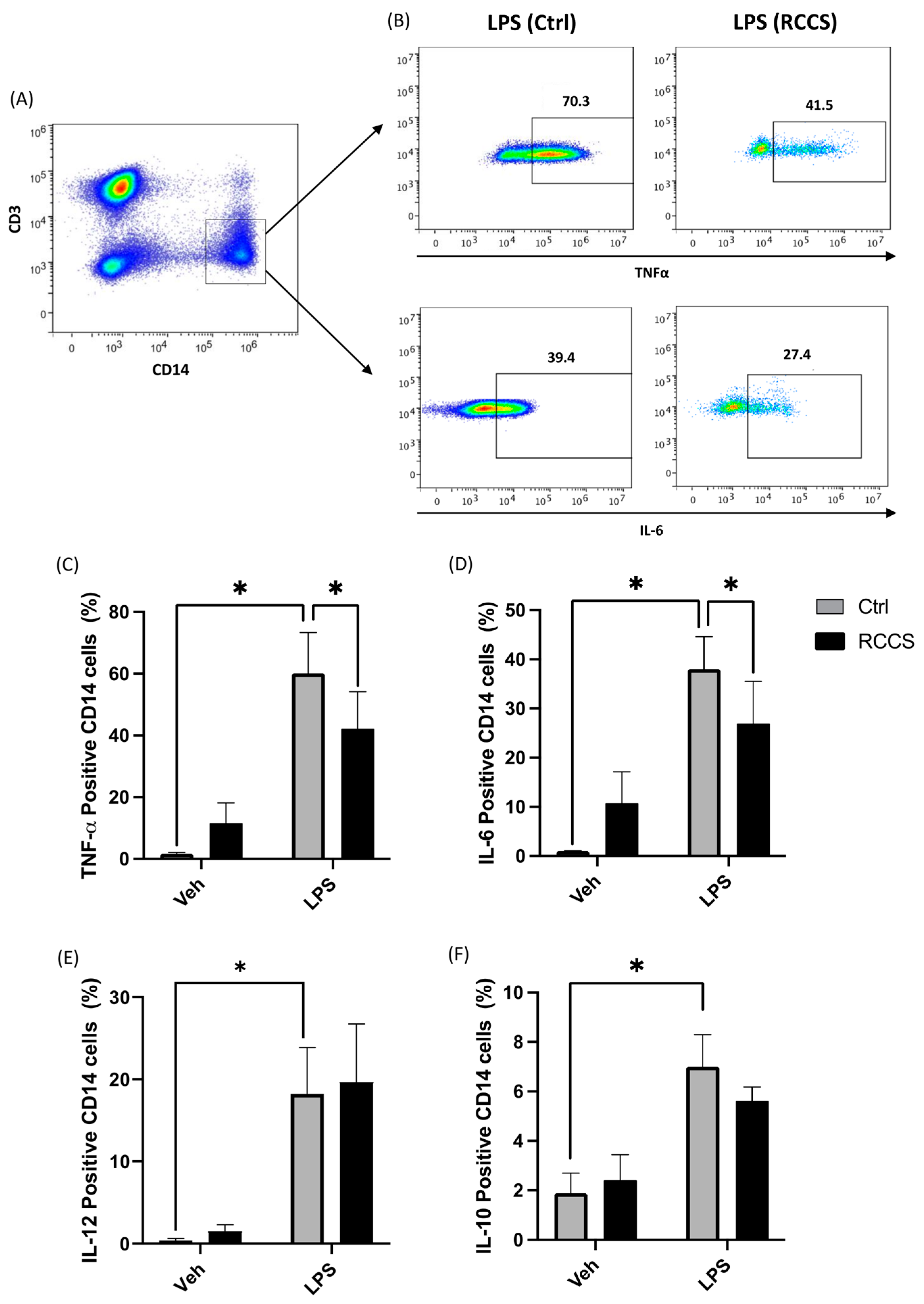
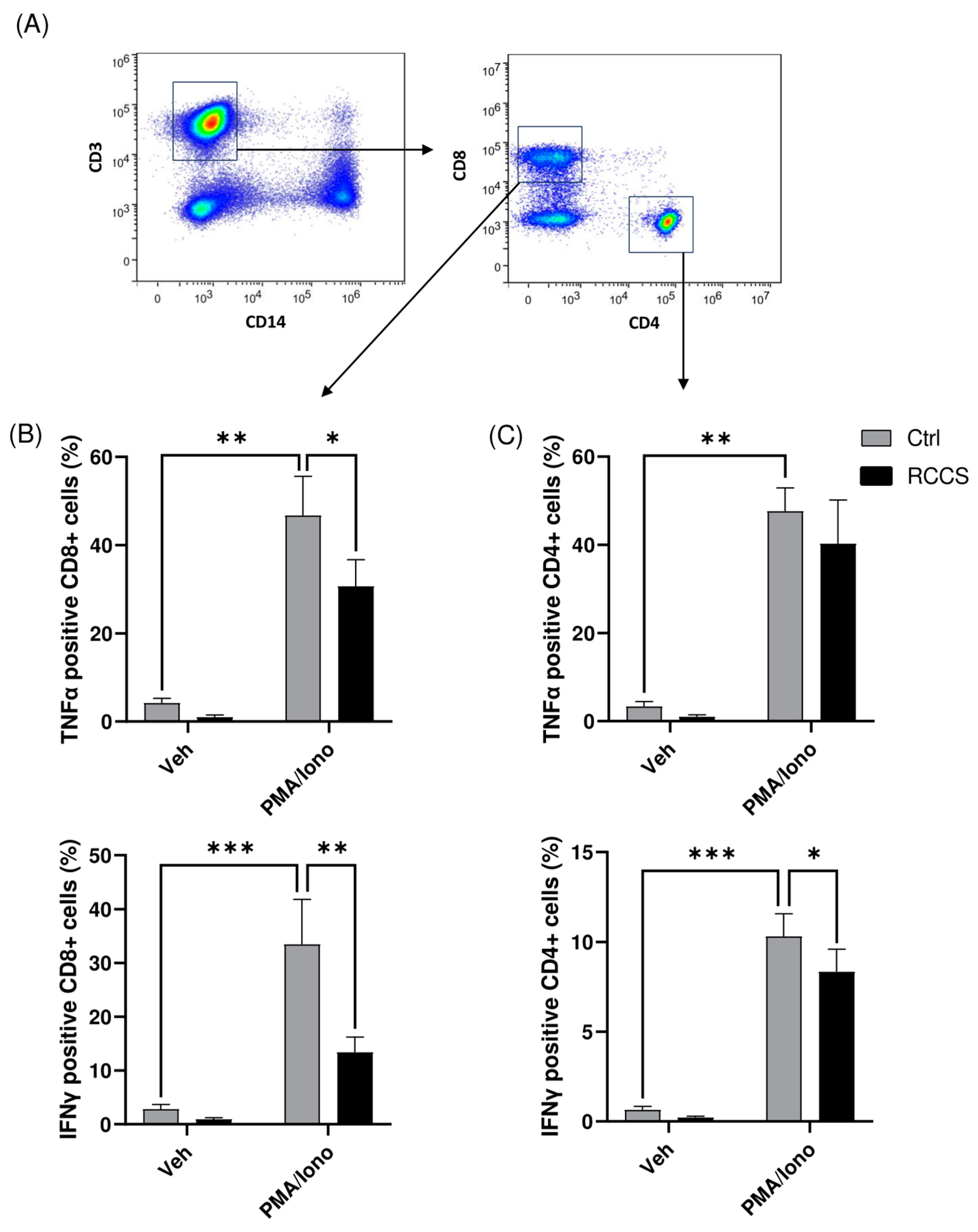
3.4. Simulated Microgravity Modifies Cytokine Production in Both Lymphocytes and Monocytes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hughes-Fulford, M. To Infinity … and beyond! Human Spaceflight and Life Science. FASEB J. 2011, 25, 2858–2864. [Google Scholar] [CrossRef] [PubMed]
- Kimzey, S.L.; Johnson, P.C.; Ritzman, S.E.; Mengel, C.E. Hematology and Immunology Studies: The Second Manned Skylab Mission. Aviat. Space Env. Med 1976, 47, 383–390. [Google Scholar]
- Man, J.; Graham, T.; Squires-Donelly, G.; Laslett, A.L. The Effects of Microgravity on Bone Structure and Function. npj Microgravity 2022, 8, 9. [Google Scholar] [CrossRef] [PubMed]
- Fava, M.; Leuti, A.; Maccarrone, M. Lipid Signalling in Human Immune Response and Bone Remodelling under Microgravity. Appl. Sci. 2020, 10, 4309. [Google Scholar] [CrossRef]
- Li, P.; Shi, J.; Zhang, P.; Wang, K.; Li, J.; Liu, H.; Zhou, Y.; Xu, X.; Hao, J.; Sun, X.; et al. Simulated Microgravity Disrupts Intestinal Homeostasis and Increases Colitis Susceptibility. FASEB J 2015, 29, 3263–3273. [Google Scholar] [CrossRef]
- Roy-O’Reilly, M.; Mulavara, A.; Williams, T. A Review of Alterations to the Brain during Spaceflight and the Potential Relevance to Crew in Long-Duration Space Exploration. npj Microgravity 2021, 7, 5. [Google Scholar] [CrossRef]
- Bradbury, P.; Wu, H.; Choi, J.U.; Rowan, A.E.; Zhang, H.; Poole, K.; Lauko, J.; Chou, J. Modeling the Impact of Microgravity at the Cellular Level: Implications for Human Disease. Front. Cell Dev. Biol. 2020, 8, 96. [Google Scholar] [CrossRef]
- Hauschild, S.; Tauber, S.; Lauber, B.; Thiel, C.S.; Layer, L.E.; Ullrich, O. T Cell Regulation in Microgravity—The Current Knowledge from in Vitro Experiments Conducted in Space, Parabolic Flights and Ground-Based Facilities. Acta Astronaut. 2014, 104, 365–377. [Google Scholar] [CrossRef]
- Ludtka, C.; Silberman, J.; Moore, E.; Allen, J.B. Macrophages in Microgravity: The Impact of Space on Immune Cells. npj Microgravity 2021, 7, 13. [Google Scholar] [CrossRef]
- Serhan, C.N. Discovery of Specialized Pro-Resolving Mediators Marks the Dawn of Resolution Physiology and Pharmacology. Mol. Asp. Med. 2017, 58, 1–11. [Google Scholar] [CrossRef]
- Leuti, A.; Fazio, D.; Fava, M.; Piccoli, A.; Oddi, S.; Maccarrone, M. Bioactive Lipids, Inflammation and Chronic Diseases. Adv. Drug Deliv. Rev. 2020, 159, 133–169. [Google Scholar] [CrossRef] [PubMed]
- Battista, N.; Meloni, M.A.; Bari, M.; Mastrangelo, N.; Galleri, G.; Rapino, C.; Dainese, E.; Agrò, A.F.; Pippia, P.; Maccarrone, M. 5-Lipoxygenase-Dependent Apoptosis of Human Lymphocytes in the International Space Station: Data from the ROALD Experiment. FASEB J. 2012, 26, 1791–1798. [Google Scholar] [CrossRef]
- Chiurchiù, V.; Leuti, A.; Cencioni, M.T.; Albanese, M.; De Bardi, M.; Bisogno, T.; Centonze, D.; Battistini, L.; Maccarrone, M. Modulation of Monocytes by Bioactive Lipid Anandamide in Multiple Sclerosis Involves Distinct Toll-like Receptors. Pharmacol. Res. 2016, 113, 313–319. [Google Scholar] [CrossRef]
- Fanti, F.; Oliva, E.; Tortolani, D.; Di Meo, C.; Fava, M.; Leuti, A.; Rapino, C.; Sergi, M.; Maccarrone, M.; Compagnone, D. μSPE Followed by HPLC-MS/MS for the Determination of Series D and E Resolvins in Biological Matrices. J. Pharm. Biomed. Anal. 2021, 203, 114181. [Google Scholar] [CrossRef] [PubMed]
- Chiang, N.; Serhan, C.N. Specialized Pro-Resolving Mediator Network: An Update on Production and Actions. Essays Biochem. 2020, 64, 443–462. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N.; Chiang, N.; Dalli, J. The Resolution Code of Acute Inflammation: Novel pro-Resolving Lipid Mediators in Resolution. Semin. Immunol. 2015, 27, 200–215. [Google Scholar] [CrossRef] [PubMed]
- Kooij, G.; Troletti, C.D.; Leuti, A.; Norris, P.C.; Riley, I.; Albanese, M.; Ruggieri, S.; Libreros, S.; van der Pol, S.M.A.; van Het Hof, B.; et al. Specialized Pro-Resolving Lipid Mediators Are Differentially Altered in Peripheral Blood of Patients with Multiple Sclerosis and Attenuate Monocyte and Blood-Brain Barrier Dysfunction. Haematologica 2020, 105, 2056–2070. [Google Scholar] [CrossRef]
- Norris, P.C.; Serhan, C.N. Metabololipidomic Profiling of Functional Immunoresolvent Clusters and Eicosanoids in Mammalian Tissues. Biochem. Biophys. Res. Commun. 2018, 504, 553–561. [Google Scholar] [CrossRef]
- Maccarrone, M.; Battista, N.; Meloni, M.; Bari, M.; Galleri, G.; Pippia, P.; Cogoli, A.; Finazzi-Agrò, A. Creating Conditions Similar to Those That Occur during Exposure of Cells to Microgravity Induces Apoptosis in Human Lymphocytes by 5-Lipoxygenase-Mediated Mitochondrial Uncoupling and Cytochrome c Release. J. Leukoc. Biol. 2003, 73, 472–481. [Google Scholar] [CrossRef]
- Maccarrone, M.; Finazzi-Agrò, A. Microgravity Increases the Affinity of Lipoxygenases for Free Fatty Acids. FEBS Lett. 2001, 489, 283. [Google Scholar] [CrossRef]
- Werz, O.; Gerstmeier, J.; Libreros, S.; De la Rosa, X.; Werner, M.; Norris, P.C.; Chiang, N.; Serhan, C.N. Human Macrophages Differentially Produce Specific Resolvin or Leukotriene Signals That Depend on Bacterial Pathogenicity. Nat. Commun. 2018, 9, 59. [Google Scholar] [CrossRef]
- Lv, H.; Yang, H.; Jiang, C.; Shi, J.; Chen, R.; Huang, Q.; Shao, D. Microgravity and Immune Cells. J. R. Soc. Interface 2023, 20, 20220869. [Google Scholar] [CrossRef] [PubMed]
- Kaur, I.; Simons, E.R.; Kapadia, A.S.; Ott, C.M.; Pierson, D.L. Effect of Spaceflight on Ability of Monocytes To Respond to Endotoxins of Gram-Negative Bacteria. Clin. Vaccine Immunol. 2008, 15, 1523–1528. [Google Scholar] [CrossRef] [PubMed]
- Crucian, B.; Stowe, R.; Mehta, S.; Uchakin, P.; Quiriarte, H.; Pierson, D.; Sams, C. Immune System Dysregulation Occurs During Short Duration Spaceflight On Board the Space Shuttle. J. Clin. Immunol. 2013, 33, 456–465. [Google Scholar] [CrossRef] [PubMed]
- Crucian, B.; Stowe, R.; Quiriarte, H.; Pierson, D.; Sams, C. Monocyte Phenotype and Cytokine Production Profiles Are Dysregulated by Short-Duration Spaceflight. Aviat. Space Environ. Med. 2011, 82, 857–862. [Google Scholar] [CrossRef] [PubMed]
- Kaur, I.; Simons, E.R.; Castro, V.A.; Ott, C.M.; Pierson, D.L. Changes in Monocyte Functions of Astronauts. Brain Behav. Immun. 2005, 19, 547–554. [Google Scholar] [CrossRef]
- Gridley, D.S.; Slater, J.M.; Luo-Owen, X.; Rizvi, A.; Chapes, S.K.; Stodieck, L.S.; Ferguson, V.L.; Pecaut, M.J. Spaceflight Effects on T Lymphocyte Distribution, Function and Gene Expression. J. Appl. Physiol. 2009, 106, 194–202. [Google Scholar] [CrossRef]
- Baqai, F.P.; Gridley, D.S.; Slater, J.M.; Luo-Owen, X.; Stodieck, L.S.; Ferguson, V.; Chapes, S.K.; Pecaut, M.J. Effects of Spaceflight on Innate Immune Function and Antioxidant Gene Expression. J. Appl. Physiol. 2009, 106, 1935–1942. [Google Scholar] [CrossRef]
- Akiyama, T.; Horie, K.; Hinoi, E.; Hiraiwa, M.; Kato, A.; Maekawa, Y.; Takahashi, A.; Furukawa, S. How Does Spaceflight Affect the Acquired Immune System? npj Microgravity 2020, 6, 14. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Ab-Conjugated Fluorophore | Manufacturer | Dilution |
|---|---|---|---|
| CD3 | FITC | Miltenyi Biotec, Bergisch Gladbach, Germany | 1:80 |
| CD4 | APC-Vio770 | Miltenyi Biotec, Bergisch Gladbach, Germany | 1:50 |
| CD8 | VioGreen | Miltenyi Biotec, Bergisch Gladbach, Germany | 1:50 |
| CD14 | APC | Miltenyi Biotec, Bergisch Gladbach, Germany | 1:50 |
| GPR-18 | Non-conjugated | Novus Bio, Minneapolis, MI, USA | 1:200 |
| GPR-32 | Non-conjugated | Genetex, Irvine, CA, USA | 1:200 |
| 5-LOX | Non-conjugated | Abcam, Cambridge, UK | 1:100 |
| ALX | APC | Miltenyi Biotec, Bergisch Gladbach, Germany | 1:50 |
| ChemR23 | APC | Miltenyi Biotec, Bergisch Gladbach, Germany | 1:50 |
| TNF-α | PE | Miltenyi Biotec, Bergisch Gladbach, Germany | 1:80 |
| IFN- γ | Vioblue | Miltenyi Biotec, Bergisch Gladbach, Germany | 1:50 |
| IL-6 | PE CF594 | Biolegend, San Diego, CA, USA | 1:400 |
| IL-10 | PE | BD Biosciences, San Jose, CA, USA | 1:50 |
| IL-12 | BV421 | Biolegend, San Diego, CA, USA | 1:400 |
| IL-17 | PE-Vio770 | Miltenyi Biotec, Bergisch Gladbach, Germany | 1:50 |
| Goat anti-Rabbit | Alexa Fluor 647 | Southern Biotech, Birmingham, AL, USA | 1:200 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leuti, A.; Fava, M.; Pellegrini, N.; Forte, G.; Fanti, F.; Della Valle, F.; De Dominicis, N.; Sergi, M.; Maccarrone, M. Simulated Microgravity Affects Pro-Resolving Properties of Primary Human Monocytes. Cells 2024, 13, 100. https://doi.org/10.3390/cells13010100
Leuti A, Fava M, Pellegrini N, Forte G, Fanti F, Della Valle F, De Dominicis N, Sergi M, Maccarrone M. Simulated Microgravity Affects Pro-Resolving Properties of Primary Human Monocytes. Cells. 2024; 13(1):100. https://doi.org/10.3390/cells13010100
Chicago/Turabian StyleLeuti, Alessandro, Marina Fava, Niccolò Pellegrini, Giulia Forte, Federico Fanti, Francesco Della Valle, Noemi De Dominicis, Manuel Sergi, and Mauro Maccarrone. 2024. "Simulated Microgravity Affects Pro-Resolving Properties of Primary Human Monocytes" Cells 13, no. 1: 100. https://doi.org/10.3390/cells13010100
APA StyleLeuti, A., Fava, M., Pellegrini, N., Forte, G., Fanti, F., Della Valle, F., De Dominicis, N., Sergi, M., & Maccarrone, M. (2024). Simulated Microgravity Affects Pro-Resolving Properties of Primary Human Monocytes. Cells, 13(1), 100. https://doi.org/10.3390/cells13010100