
Novel In Vitro Models for Cell Differentiation and Drug Transport Studies of the Human Intestine

 ,
, 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Cell Culture
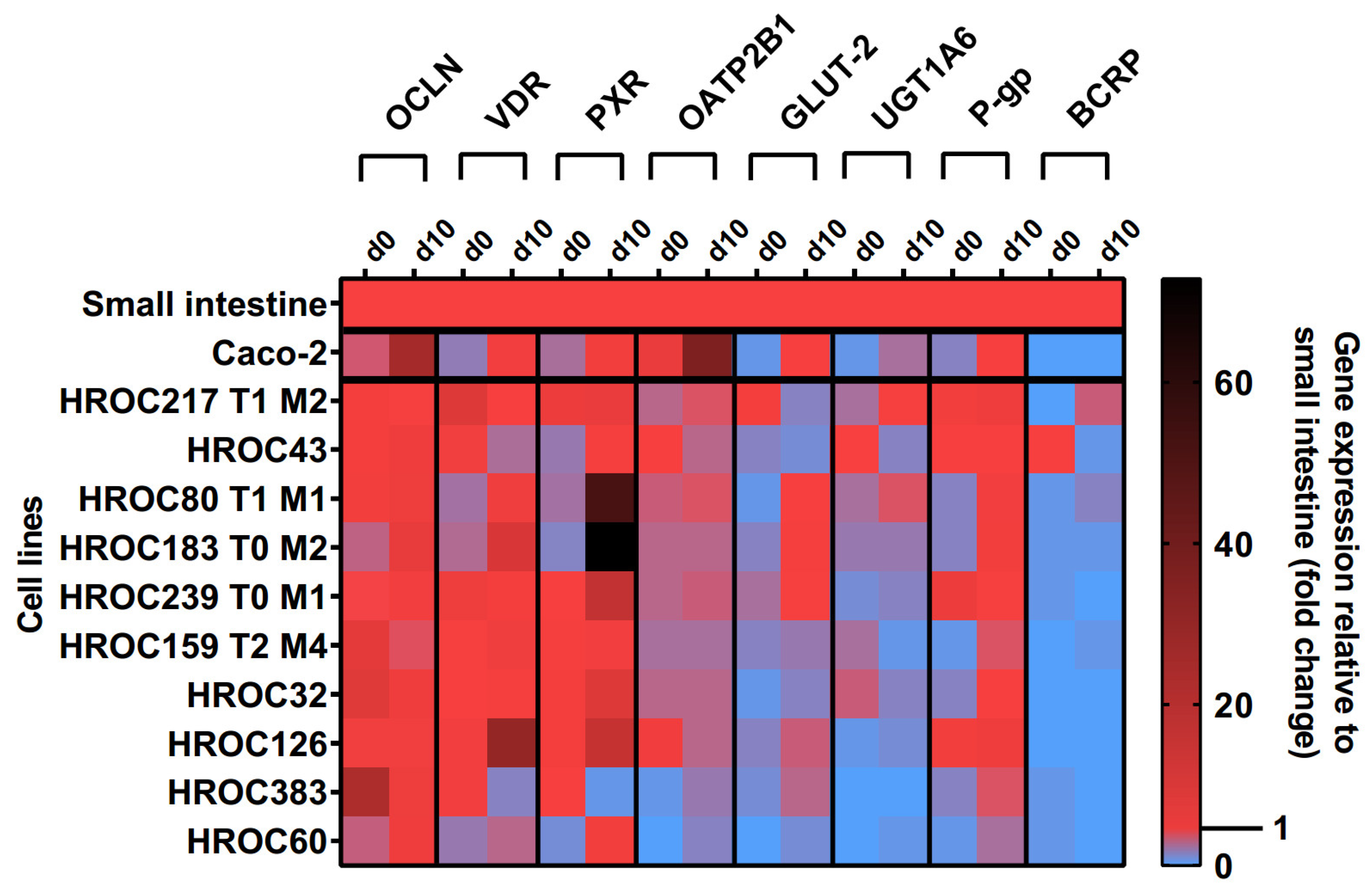
2.3. Gene Expression Analysis
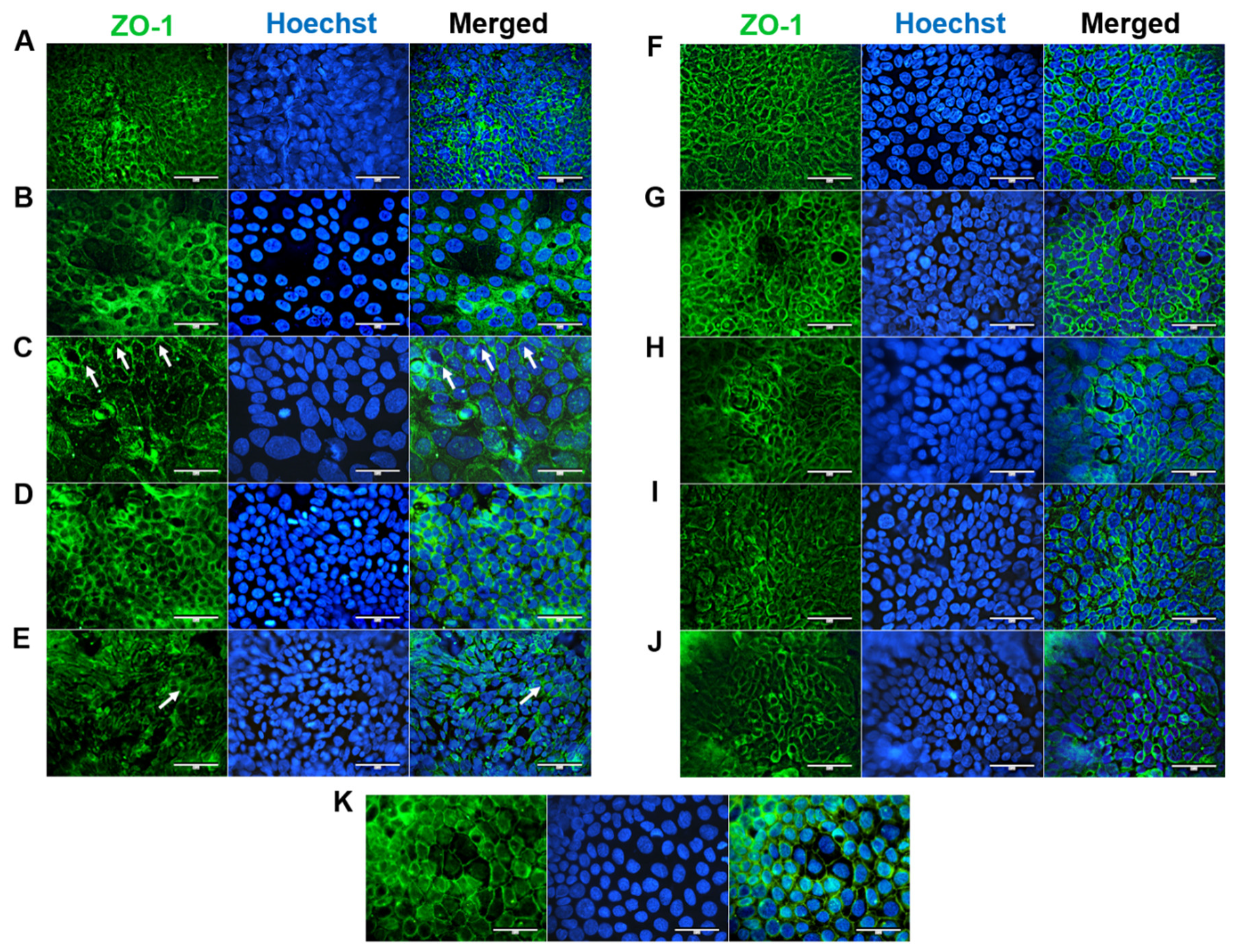
2.4. Immunocytochemistry
2.5. Protein Quantification by Liquid Chromatography–Mass Spectrometry/Mass Spectrometry (LC-MS/MS)
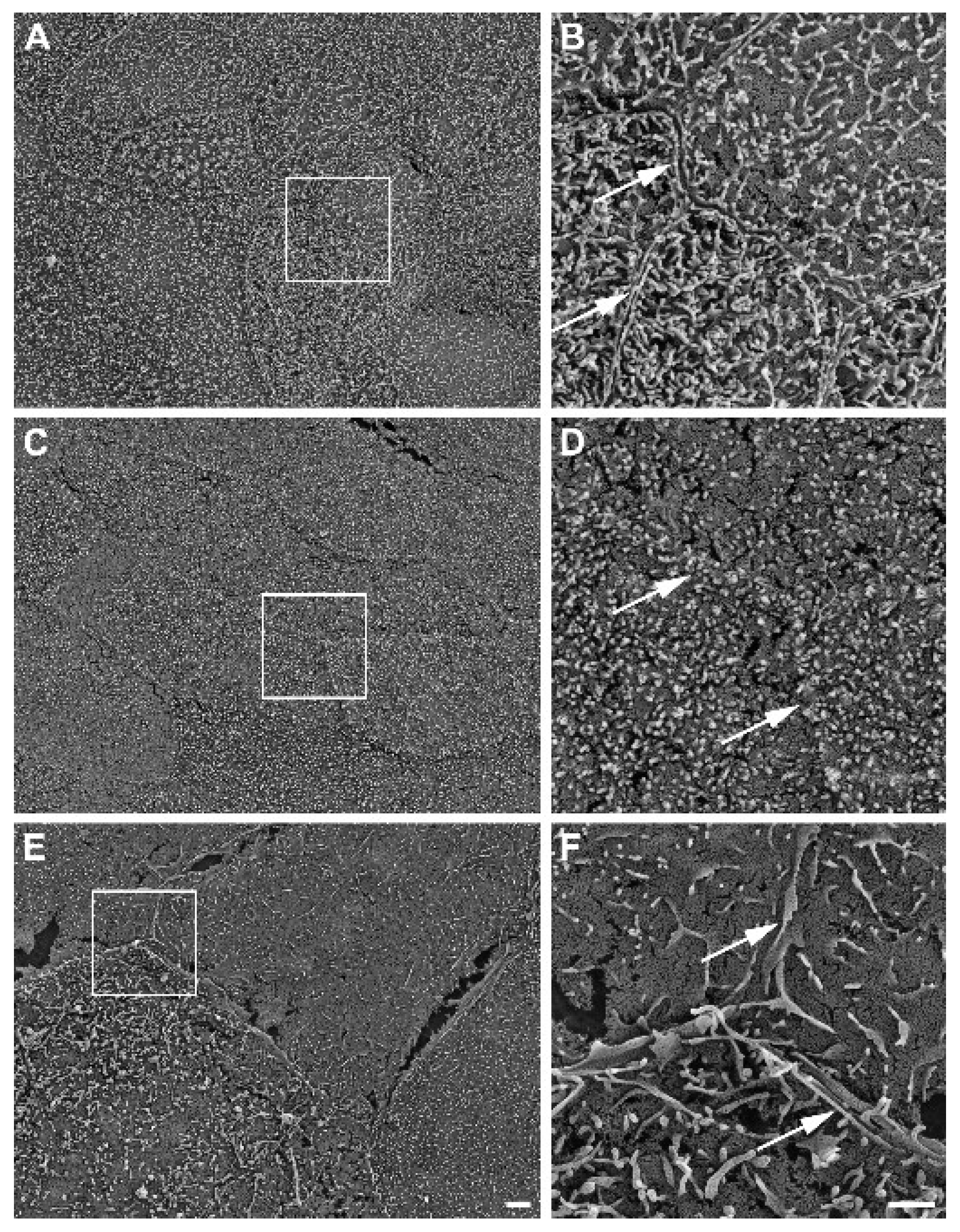
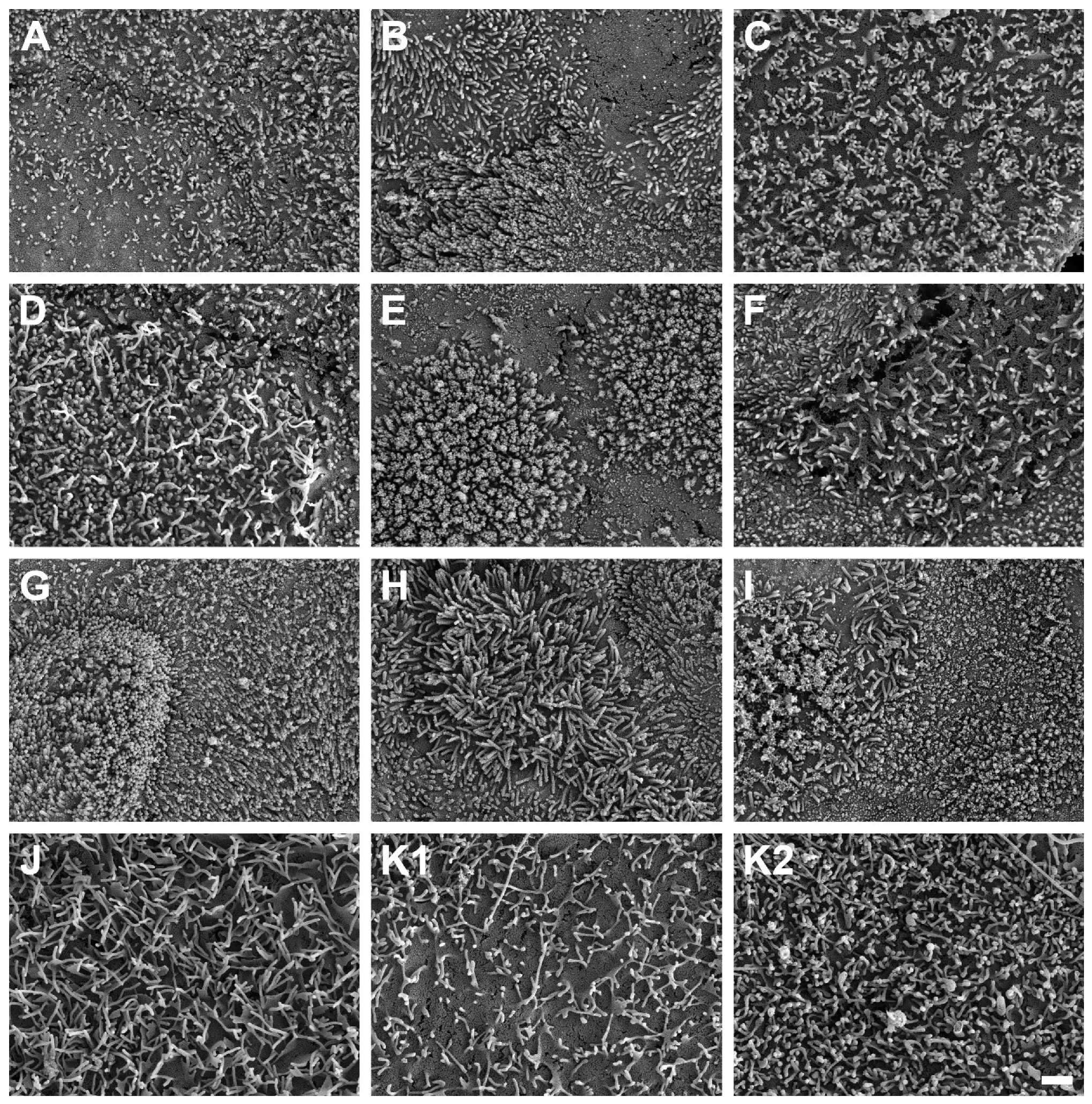
2.6. Scanning Electron Microscopy
2.7. Data Analysis
3. Results
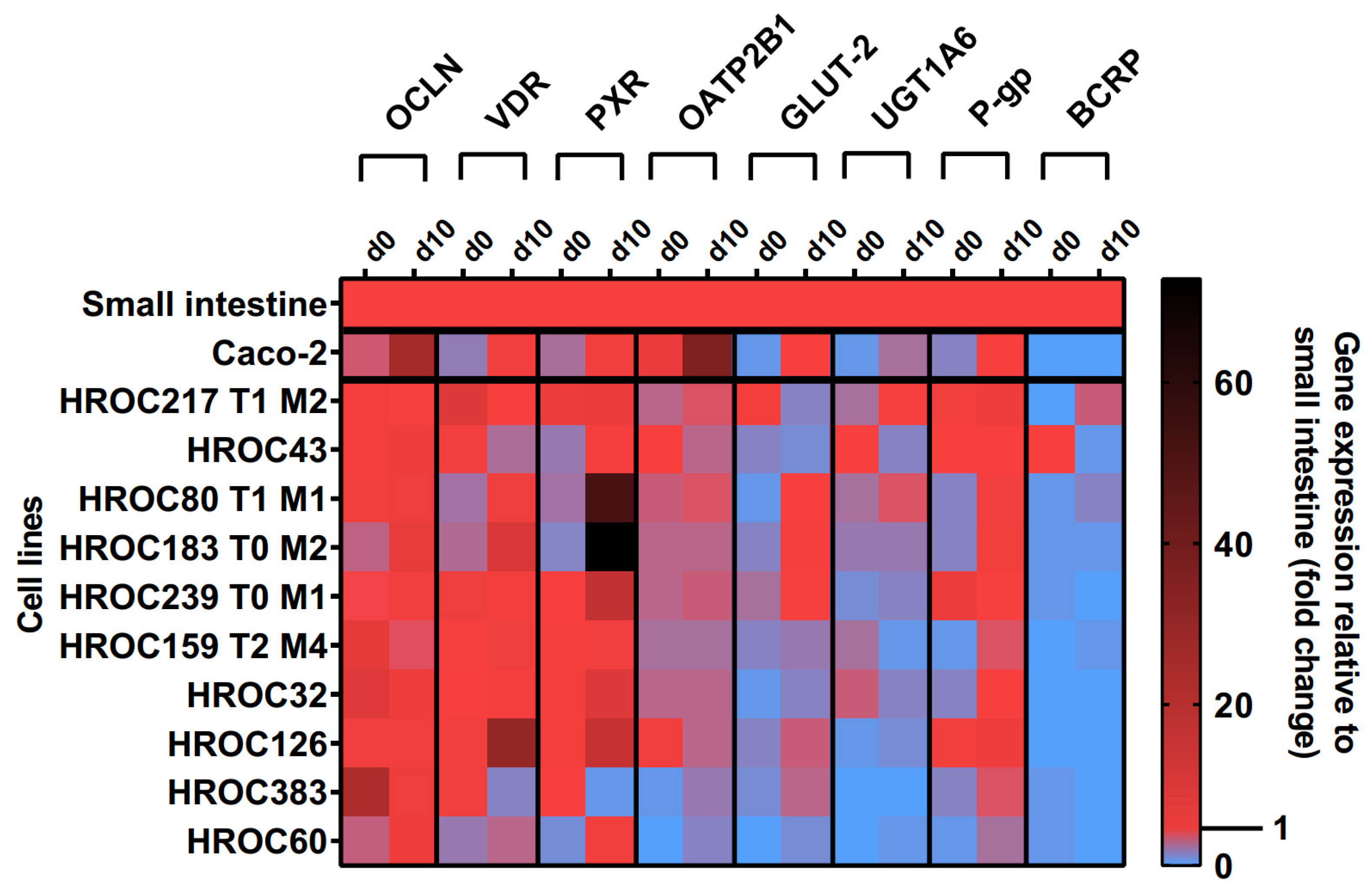
3.1. Culture Time-Dependent Gene Expression
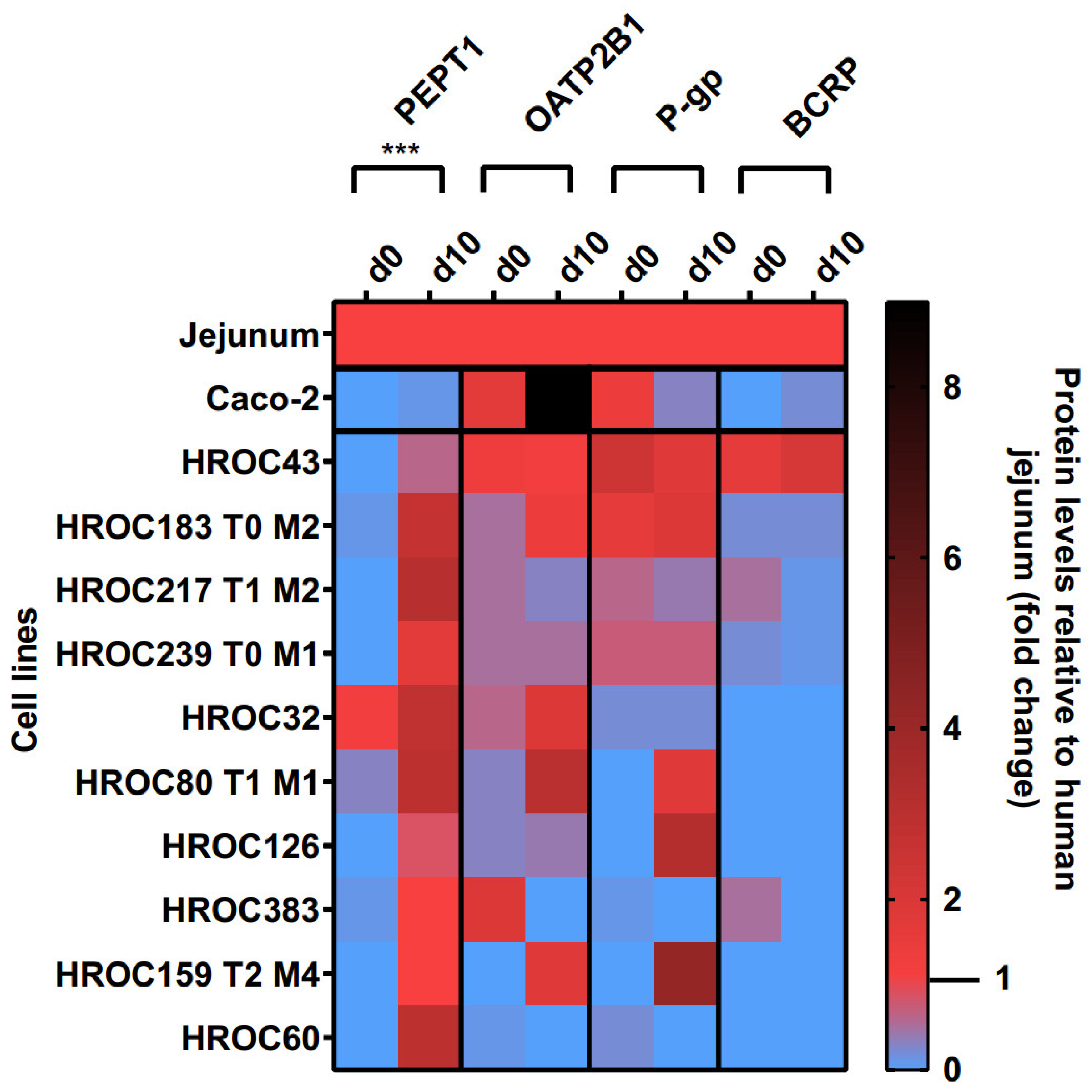
3.2. Culture Time-Dependent Protein Levels
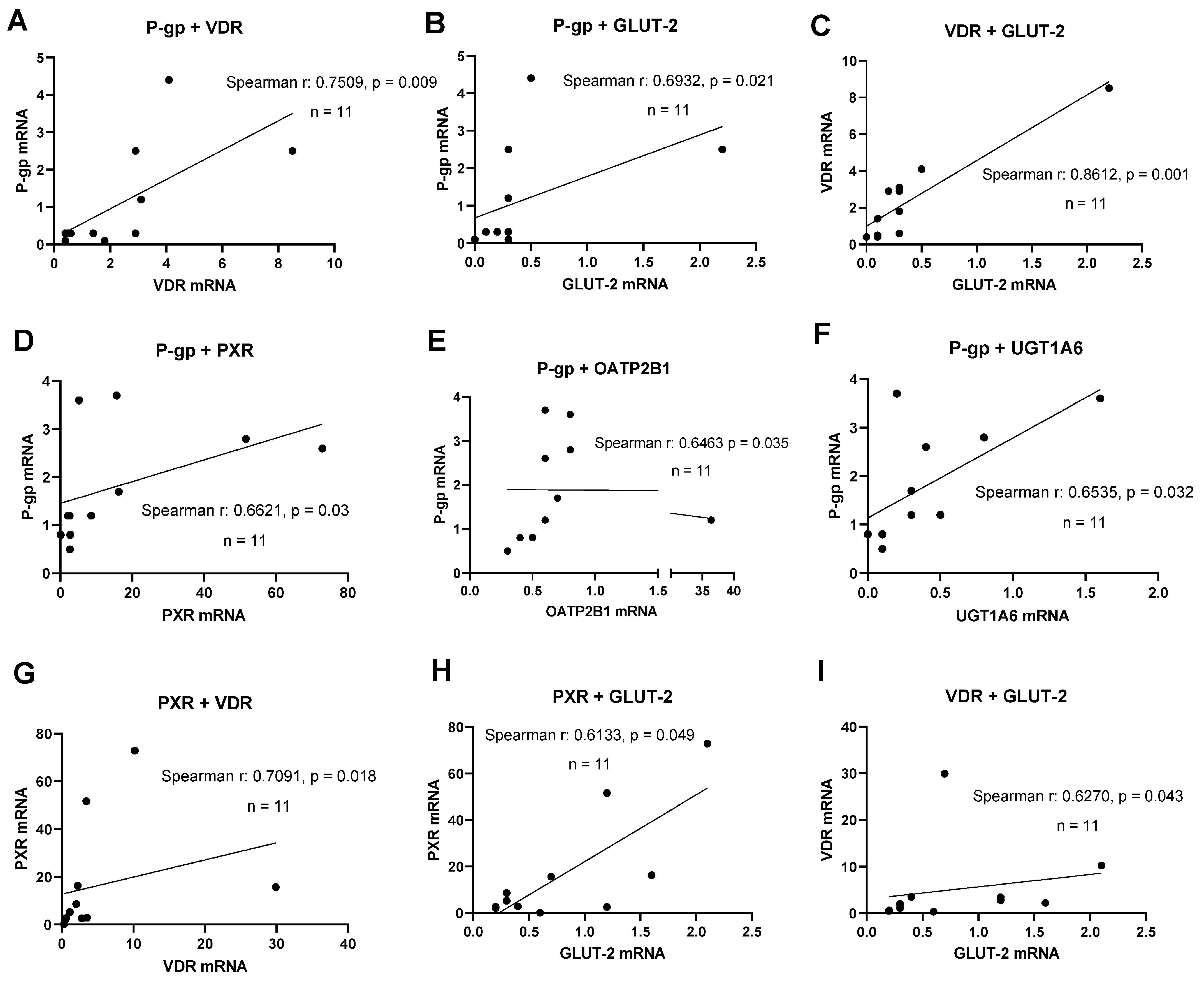
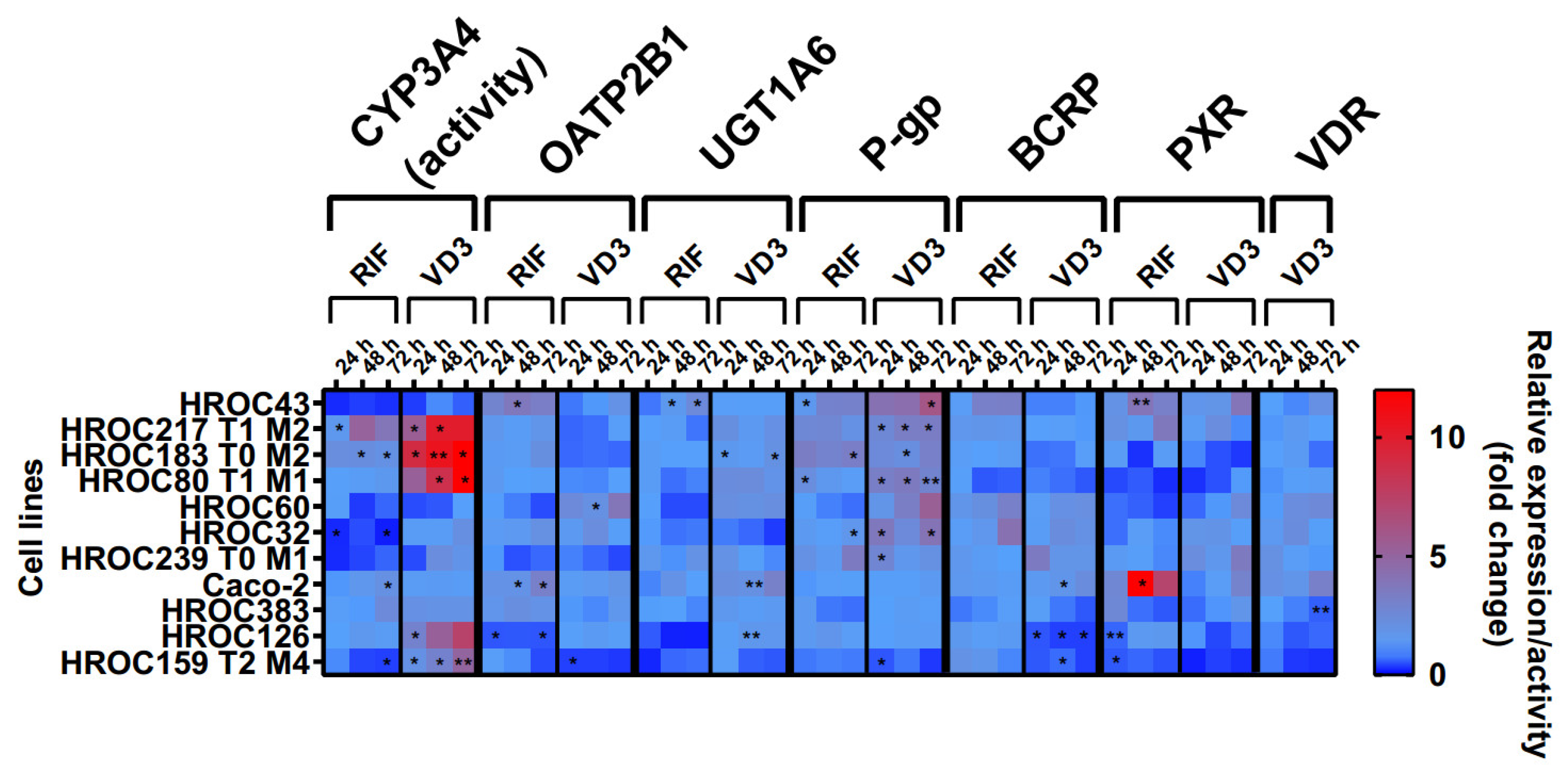
3.3. Regulation of Intestinal Drug Transporter Expression
3.4. Cell Morphology and Differentiation
3.5. Detection of Intestinal Brush Border
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ABC | ATP-binding cassette |
| ADME | administration distribution metabolism excretion |
| BCRP | breast cancer resistance protein |
| CHGA | chromogranin A |
| CYP3A4 | cytochrome P450 (CYP) 3A4 |
| GCs | goblet cells |
| DT | drug transporter |
| EECs | enteroendocrine cells |
| GLUT-2 | glucose transporter 2 |
| HROC | Hansestadt Rostock CRC |
| IECs | intestinal epithelial cells |
| LC-MS/MS | liquid chromatography–mass spectrometry/mass spectrometry |
| LYZ | lysozyme |
| MUC2 | mucin 2 |
| NRs | nuclear receptors |
| OATP2B1 | organic anion-transporting polypeptide 2B1 |
| PEPT1 | peptide transporter 1 |
| P-gp | permeability glycoprotein |
| PCs | Paneth cells |
| PXR | pregnane X receptor |
| RIF | rifampicin |
| SLC | solute carrier transporters |
| TJ | tight junction |
| VD3 | vitamin D3 |
| ZO-1 | zonula occludens-1 |
References
- Stewart, K.D.; Johnston, J.A.; Matza, L.S.; Curtis, S.E.; Havel, H.A.; Sweetana, S.A.; Gelhorn, H.L. Preference for pharmaceutical formulation and treatment process attributes. Patient Prefer. Adherence 2016, 10, 1385–1399. [Google Scholar] [CrossRef] [PubMed]
- Doherty, M.M.; Pang, K.S. First-pass effect: Significance of the intestine for absorption and metabolism. Drug Chem. Toxicol. 1997, 20, 329–344. [Google Scholar] [CrossRef] [PubMed]
- Berggren, S.; Gall, C.; Wollnitz, N.; Ekelund, M.; Karlbom, U.; Hoogstraate, J.; Schrenk, D.; Lennernäs, H. Gene and protein expression of P-glycoprotein, MRP1, MRP2, and CYP3A4 in the small and large human intestine. Mol. Pharm. 2007, 4, 252–257. [Google Scholar] [CrossRef] [PubMed]
- Lea, T. The Impact of Food Bioactives on Health: In Vitro and Ex Vivo Models; Verhoeckx, K., Cotter, P., López-Expósito, I., Kleiveland, C., Lea, T., Mackie, A., Requena, T., Swiatecka, D., Wichers, H., Eds.; Springer: Cham, Switzerland, 2015. [Google Scholar]
- Brück, S.; Strohmeier, J.; Busch, D.; Drozdzik, M.; Oswald, S. Caco-2 cells—Expression, regulation and function of drug transporters compared with human jejunal tissue. Biopharm. Drug Dispos. 2017, 38, 115–126. [Google Scholar] [CrossRef]
- Sancho, E.; Batlle, E.; Clevers, H. Live and let die in the intestinal epithelium. Curr. Opin. Cell Biol. 2003, 15, 763–770. [Google Scholar] [CrossRef]
- Cheng, H.; Leblond, C.P. Origin, differentiation and renewal of the four main epithelial cell types in the mouse small intestine. V. Unitarian Theory of the origin of the four epithelial cell types. Am. J. Anat. 1974, 141, 537–561. [Google Scholar] [CrossRef]
- Potten, C.S. Stem cells in gastrointestinal epithelium: Numbers, characteristics and death. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 1998, 353, 821–830. [Google Scholar] [CrossRef]
- Ménochet, K.; Chanteux, H.; Henshall, J.; Nicolas, J.M.; Wright, S.; van Asperen, J.; Ungell, A.L. Intestinal Epithelium and Drug Transporters. In Oral Drug Delivery for Modified Release Formulations; Kostewicz, E.S., Vertzoni, M., Benson, H.A.E., Roberts, M.S., Eds.; Wiley: Hoboken, NJ, USA, 2022; pp. 39–64. [Google Scholar]
- Hosomi, A.; Nakanishi, T.; Fujita, T.; Tamai, I. Extra-renal elimination of uric acid via intestinal efflux transporter BCRP/ABCG2. PLoS ONE 2012, 7, e30456. [Google Scholar] [CrossRef]
- Urquhart, B.L.; Tirona, R.G.; Kim, R.B. Nuclear receptors and the regulation of drug-metabolizing enzymes and drug transporters: Implications for interindividual variability in response to drugs. J. Clin. Pharmacol. 2007, 47, 566–578. [Google Scholar] [CrossRef]
- Przybylla, R.; Mullins, C.S.; Krohn, M.; Oswald, S.; Linnebacher, M. Establishment and Characterization of Novel Human Intestinal In Vitro Models for Absorption and First-Pass Metabolism Studies. Int. J. Mol. Sci. 2022, 23, 9861. [Google Scholar] [CrossRef]
- Mullins, C.S.; Micheel, B.; Matschos, S.; Leuchter, M.; Bürtin, F.; Krohn, M.; Hühns, M.; Klar, E.; Prall, F.; Linnebacher, M. Integrated Biobanking and Tumor Model Establishment of Human Colorectal Carcinoma Provides Excellent Tools for Preclinical Research. Cancers 2019, 11, 1520. [Google Scholar] [CrossRef] [PubMed]
- Gröer, C.; Brück, S.; Lai, Y.; Paulick, A.; Busemann, A.; Heidecke, C.D.; Siegmund, W.; Oswald, S. LC-MS/MS-based quantification of clinically relevant intestinal uptake and efflux transporter proteins. J. Pharm. Biomed. Anal. 2013, 85, 253–261. [Google Scholar] [CrossRef] [PubMed]
- Wenzel, C.; Drozdzik, M.; Oswald, S. Mass spectrometry-based targeted proteomics method for the quantification of clinically relevant drug metabolizing enzymes in human specimens. J. Chromatogr. B 2021, 1180, 122891. [Google Scholar] [CrossRef] [PubMed]
- Wenzel, C.; Gödtke, L.; Reichstein, A.; Keiser, M.; Busch, D.; Drozdzik, M.; Oswald, S. Gene Expression and Protein Abundance of Nuclear Receptors in Human Intestine and Liver: A New Application for Mass Spectrometry-Based Targeted Proteomics. Molecules 2022, 27, 4629. [Google Scholar] [CrossRef] [PubMed]
- Howarth, A.G.; Hughes, M.R.; Stevenson, B.R. Detection of the tight junction-associated protein ZO-1 in astrocytes and other nonepithelial cell types. Am. J. Physiol. 1992, 262, C461–C469. [Google Scholar] [CrossRef]
- Speer, J.E.; Gunasekara, D.B.; Wang, Y.; Fallon, J.K.; Attayek, P.J.; Smith, P.C.; Sims, C.E.; Allbritton, N.L. Molecular transport through primary human small intestinal monolayers by culture on a collagen scaffold with a gradient of chemical cross-linking. J. Biol. Eng. 2019, 13, 36. [Google Scholar] [CrossRef]
- Herath, M.; Hosie, S.; Bornstein, J.C.; Franks, A.E.; Hill-Yardin, E.L. The Role of the Gastrointestinal Mucus System in Intestinal Homeostasis: Implications for Neurological Disorders. Front. Cell. Infect. Microbiol. 2020, 10, 248. [Google Scholar] [CrossRef]
- Lueschow, S.R.; McElroy, S.J. The Paneth Cell: The Curator and Defender of the Immature Small Intestine. Front. Immunol. 2020, 11, 587. [Google Scholar] [CrossRef]
- Rehfeld, J.F. A centenary of gastrointestinal endocrinology. Horm. Metab. Res. 2004, 36, 735–741. [Google Scholar] [CrossRef]
- Müller, J.; Keiser, M.; Drozdzik, M.; Oswald, S. Expression, regulation and function of intestinal drug transporters: An update. Biol. Chem. 2017, 398, 175–192. [Google Scholar] [CrossRef]
- Qiu, Y.; Chen, Y.; Zhang, G.G.Z.; Yu, L.X.; Mantri, R.V. Developing solid oral dosage forms. In Pharmaceutical Theory & Practice, 2nd ed.; Elsevier: Amsterdam, The Netherlands; Academic Press: Cambridge, MA, USA, 2017. [Google Scholar]
- Mueckler, M. Facilitative glucose transporters. Eur. J. Biochem. 1994, 219, 713–725. [Google Scholar] [CrossRef] [PubMed]
- Kleiveland, C.R. Co-cultivation of Caco-2 and HT-29MTX. In The Impact of Food Bioactives on Health: In Vitro and Ex Vivo Models; Verhoeckx, K., Cotter, P., López-Expósito, I., Kleiveland, C., Lea, T., Mackie, A., Requena, T., Swiatecka, D., Wichers, H., Eds.; Springer: Cham, Switzerland, 2015. [Google Scholar]
- Sommers, C.L.; Byers, S.W.; Thompson, E.W.; Torri, J.A.; Gelmann, E.P. Differentiation state and invasiveness of human breast cancer cell lines. Breast Cancer Res. Treat. 1994, 31, 325–335. [Google Scholar] [CrossRef] [PubMed]
- Sheehan, J.K.; Thornton, D.J.; Howard, M.; Carlstedt, I.; Corfield, A.P.; Paraskeva, C. Biosynthesis of the MUC2 mucin: Evidence for a slow assembly of fully glycosylated units. Biochem. J. 1996, 315, 1055–1060. [Google Scholar] [CrossRef]
- Blank, M.; Klussmann, E.; Krüger-Krasagakes, S.; Schmitt-Gräff, A.; Stolte, M.; Bornhoeft, G.; Stein, H.; Xing, P.X.; McKenzie, I.F.; Verstijnen, C.P.; et al. Expression of MUC2-mucin in colorectal adenomas and carcinomas of different histological types. Int. J. Cancer 1994, 59, 301–306. [Google Scholar] [CrossRef]
- Wang, H.; Jin, S.; Lu, H.; Mi, S.; Shao, W.; Zuo, X.; Yin, H.; Zeng, S.; Shimamoto, F.; Qi, G. Expression of survivin, MUC2 and MUC5 in colorectal cancer and their association with clinicopathological characteristics. Oncol. Lett. 2017, 14, 1011–1016. [Google Scholar] [CrossRef]
- Porter, E.M.; Bevins, C.L.; Ghosh, D.; Ganz, T. The multifaceted Paneth cell. Cell. Mol. Life Sci. CMLS 2002, 59, 156–170. [Google Scholar] [CrossRef]
- Luo, W.; Hofstetter, W.L.; Tan, D. Gastroesophageal junction Paneth cell carcinoma with extensive cystic and secretory features-case report and literature review. Diagn. Pathol. 2019, 14, 1. [Google Scholar] [CrossRef]
- Fodde, R.; Verhagen, M.; Joosten, R.; Schmitt, M.; Sacchetti, A.; Choi, J.; Välimäki, N.; Aaltonen, L.; Augenlicht, L. Paneth cells as the origin of intestinal cancer in the context of inflammation. Res. Sq. 2023, 2023, rs.3.rs-2458794. [Google Scholar] [CrossRef]
- Sasaki, N.; Sachs, N.; Wiebrands, K.; Ellenbroek, S.I.J.; Fumagalli, A.; Lyubimova, A.; Begthel, H.; van den Born, M.; van Es, J.H.; Karthaus, W.R.; et al. Reg4+ deep crypt secretory cells function as epithelial niche for Lgr5+ stem cells in colon. Proc. Natl. Acad. Sci. USA 2016, 113, E5399–E5407. [Google Scholar] [CrossRef]
- Atanga, R.; Singh, V.; In, J.G. Intestinal Enteroendocrine Cells: Present and Future Druggable Targets. Int. J. Mol. Sci. 2023, 24, 8836. [Google Scholar] [CrossRef]
- Kim, T.; Tao-Cheng, J.-H.; Eiden, L.E.; Loh, Y. Chromogranin A, an “On/Off” Switch Controlling Dense-Core Secretory Granule Biogenesis. Cell 2001, 106, 499–509. [Google Scholar] [CrossRef] [PubMed]
- Eissa, N.; Hussein, H.; Hendy, G.N.; Bernstein, C.N.; Ghia, J.-E. Chromogranin-A and its derived peptides and their pharmacological effects during intestinal inflammation. Biochem. Pharmacol. 2018, 152, 315–326. [Google Scholar] [CrossRef] [PubMed]
- Schulte, R.R.; Ho, R.H. Organic Anion Transporting Polypeptides: Emerging Roles in Cancer Pharmacology. Mol. Pharmacol. 2019, 95, 490–506. [Google Scholar] [CrossRef] [PubMed]
- Van Klinken, B.J.W.; Oussoren, E.; Weenink, J.J.; Strous, G.J.; Büller, H.A.; Dekker, J.; Einerhand, A.W. The human intestinal cell lines Caco-2 and LS174T as models to study cell-type specific mucin expression. Glycoconj. J. 1996, 13, 757–768. [Google Scholar] [CrossRef] [PubMed]
- Pshezhetsky, A.V.; Fedjaev, M.; Ashmarina, L.; Mazur, A.; Budman, L.; Sinnett, D.; Labuda, D.; Beaulieu, J.F.; Ménard, D.; Nifant’ev, I.; et al. Subcellular proteomics of cell differentiation: Quantitative analysis of the plasma membrane proteome of Caco-2 cells. Proteomics 2007, 7, 2201–2215. [Google Scholar] [CrossRef]
- Sinnecker, H.; Ramaker, K.; Frey, A. Coating with luminal gut-constituents alters adherence of nanoparticles to intestinal epithelial cells. Beilstein J. Nanotechnol. 2014, 5, 2308–2315. [Google Scholar] [CrossRef]
- Lopez-Escalera, S.; Wellejus, A. Evaluation of Caco-2 and human intestinal epithelial cells as in vitro models of colonic and small intestinal integrity. Biochem. Biophys. Rep. 2022, 31, 101314. [Google Scholar] [CrossRef]
- Hubatsch, I.; Ragnarsson, E.G.E.; Artursson, P. Determination of drug permeability and prediction of drug absorption in Caco-2 monolayers. Nat. Protoc. 2007, 2, 2111–2119. [Google Scholar] [CrossRef]
- Simon-Assmann, P.; Turck, N.; Sidhoum-Jenny, M.; Gradwohl, G.; Kedinger, M. In vitro models of intestinal epithelial cell differentiation. Cell Biol. Toxicol. 2007, 23, 241–256. [Google Scholar] [CrossRef]
- Ohura, K.; Nishiyama, H.; Saco, S.; Kurokawa, K.; Imai, T. Establishment and Characterization of a Novel Caco-2 Subclone with a Similar Low Expression Level of Human Carboxylesterase 1 to Human Small Intestine. Drug Metab. Dispos. Biol. Fate Chem. 2016, 44, 1890–1898. [Google Scholar] [CrossRef]
- Antunes, F.; Andrade, F.; Araújo, F.; Ferreira, D.; Sarmento, B. Establishment of a triple co-culture in vitro cell models to study intestinal absorption of peptide drugs. Eur. J. Pharm. Biopharm. 2013, 83, 427–435. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Xu, C.; Chen, P.; Hu, J.; Hu, R.; Huang, M.; Bi, H. Development, validation, and application of a novel 7-day Caco-2 cell culture system. J. Pharmacol. Toxicol. Methods 2014, 70, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Chong, S.; Dando, S.A.; Morrison, R.A. Evaluation of Biocoat intestinal epithelium differentiation environment (3-day cultured Caco-2 cells) as an absorption screening model with improved productivity. Pharm. Res. 1997, 14, 1835–1837. [Google Scholar] [CrossRef] [PubMed]
- Sevin, E.; Dehouck, L.; Fabulas-da Costa, A.; Cecchelli, R.; Dehouck, M.P.; Lundquist, S.; Culot, M. Accelerated Caco-2 cell permeability model for drug discovery. J. Pharmacol. Toxicol. Methods 2013, 68, 334–339. [Google Scholar] [CrossRef] [PubMed]
- Miura, S.; Sato, K.; Kato-Negishi, M.; Teshima, T.; Takeuchi, S. Fluid shear triggers microvilli formation via mechanosensitive activation of TRPV6. Nat. Commun. 2015, 6, 8871. [Google Scholar] [CrossRef]
- Crawley, S.W.; Mooseker, M.S.; Tyska, M.J. Shaping the intestinal brush border. J. Cell Biol. 2014, 207, 441–451. [Google Scholar] [CrossRef]
- Glaeser, H.; Bailey, D.G.; Dresser, G.K.; Gregor, J.C.; Schwarz, U.I.; McGrath, J.S.; Jolicoeur, E.; Lee, W.; Leake, B.F.; Tirona, R.G.; et al. Intestinal Drug Transporter Expression and the Impact of Grapefruit Juice in Humans. Clin. Pharmacol. Ther. 2007, 81, 362–370. [Google Scholar] [CrossRef]
- Ranhotra, H.S.; Flannigan, K.L.; Brave, M.; Mukherjee, S.; Lukin, D.J.; Hirota, S.A.; Mani, S. Xenobiotic Receptor-Mediated Regulation of Intestinal Barrier Function and Innate Immunity. Nucl. Recept. Res. 2016, 3, 101199. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | 5′-3′ | Sequences | Product Size (bp) |
|---|---|---|---|
| ABCB1 (P-gp) | Forward Reverse | GCTGTCAAGGAAGCCAATGCCT TGCAATGGCGATCCTCTGCTTC | 120 |
| ABCG2 (BCRP) | Forward Reverse | GTTCTCAGCAGCTCTTCGGCTT TCCTCCAGACACACCACGGATA | 145 |
| GAPDH | Forward Reverse | GAAGGTGAAGGTCGGAGTC GAAGATGGTGATGGGATTTC | 226 |
| SLC2A2 (GLUT-2) | Forward Reverse | ATGTCAGTGGGACTTGTGCTGC AACTCAGCCACCATGAACCAGG | 131 |
| SLCO2B1 (OATP2B1) | Forward Reverse | TGGGCACAGAAAACACACCT CGGCTGCCAAAATAGCTCAC | 265 |
| OCLN | Forward Reverse | ATGGCAAAGTGAATGACAAGCGG CTGTAACGAGGCTGCCTGAAGT | 124 |
| NR1I2 (PXR) | Forward Reverse | GCTGTCCTACTGCTTGGAAGAC CTGCATCAGCACATACTCCTCC | 124 |
| UGT1A6 | Forward Reverse | GCAAAGCGCATGGAGACTAAGG GGTCCTTGTGAAGGCTGGAGAG | 148 |
| NR1I1 (VDR) | Forward Reverse | CGCATCATTGCCATACTGCTGG CCACCATCATTCACACGAACTGG | 101 |
| Antibody | Host | Company | Dilution | Catalogue Number |
|---|---|---|---|---|
| anti chromogranin A | mouse | Santa Cruz | 1:50 | sc-393941 |
| anti lysozyme C | mouse | Santa Cruz | 1:50 | sc-518012 |
| anti mucin 2 | mouse | Santa Cruz | 1:50 | sc-515032 |
| anti ZO-1 | rat | Santa Cruz | 1:50 | sc-33725 |
| Cell Line | Differentiation Grade | Advantages | Limitations | Applications |
|---|---|---|---|---|
| HROC32 | Presence of GCs, PCs, and EECs; villi formation | Protein abundance of PEPT1, OATP2B1, and P-gp; high basal CYP3A4 activity; high efflux effects | Lack of BCRP protein and mRNA, lower levels of GLUT-2 mRNA compared with small intestine, colonic origin | VDR-mediated first-pass metabolism, cell differentiation, cell interaction, HTS |
| HROC43 | Well-developed TJs; closely resembles in vivo jejunal properties; presence of GCs, PCs, and EECs; villi formation | High basal CYP3A4 activity; protein abundance of PEPT1, OATP2B1, P-gp, and BCRP; PXR-/VDR-mediated regulation of CYP3A4 activity + DT mRNA | Lower levels of GLUT-2 mRNA compared with small intestine, colonic origin | First-pass metabolism, permeability, host–microbe interaction, tissue regeneration, HTS |
| HROC60 | Presence of GCs and EECs, villi formation, LYZ expressed | Well-developed TJs, protein abundance of PEPT1, high TEER, stable long-term culture, high VDR-mediated regulation of DTs | Limited degree of differentiation; poor abundance of CYP3A4 and DTs, limited reflection of the in vivo situation; colonic origin | Gut-on-a-chip, tissue regeneration, HTS |
| HROC80 T1 M1 | Presence of GCs, villi formation | Well-developed TJs, PXR-/VDR-mediated regulation of CYP3A4 + DTs | Lack of BCRP protein, limited degree of differentiation (lack of EECs and LYZ), poor PXR-mediated drug response, colonic origin | First-pass metabolism, HTS |
| HROC126 | EECs present, villi formation | VDR-mediated induction of CYP3A4 activity | Limited degree of differentiation (lack of GCs and LYZ), rectal cancer origin, poor PXR-mediated regulation of DTs, VDR overexpressed | VDR-mediated first-pass metabolism, rectal drug administration, HTS |
| HROC159 T2 M4 | GCs present, LYZ expressed, villi formation | Well-developed TJs, high basal CYP3A4 activity, appropriate VDR-mediated regulation of CYP3A4 activity | Poor gene expression and protein levels of multiple DTs, colonic origin | First-pass metabolism, tissue regeneration, HTS |
| HROC183 T0 M2 | Closely resembles in vivo jejunal properties, presence of GCs and EECs, LYZ expressed, villi formation | Well-developed TJs, VDR-mediated induction of CYP3A4 activity, appropriate PXR-/VDR-mediated regulation of CYP3A4 activity and P-gp mRNA | Low basal CYP3A4 activity, colonic origin | PXR-mediated first-pass metabolism, HTS |
| HROC217 T1 M2 | Closely resembles in vivo jejunal properties, presence of GCs and EECs, LYZ expressed, villi formation | Well-developed TJs; protein abundance of PEPT1, OATP2B1, P-gp, and BCRP; appropriate PXR-/VDR-mediated regulation of CYP3A4 activity and P-gp mRNA | Lower protein levels of OATP2B1, P-gp, and BCRP compared with jejunum; colonic origin | PXR-/VDR-mediated first-pass metabolism, HTS |
| HROC239 T0 M1 | Closely resembles in vivo jejunal properties, presence of EECs, villi formation | Protein abundance of PEPT1, OATP2B1, P-gp, and BCRP | Rectal cancer origin; PXR overexpressed; low UGT1A6 mRNA expression; lack of GCs and PCs; colonic origin; lower protein levels of OATP2B1, P-gp and BCRP compared with jejunum | Rectal drug administration, HTS |
| HROC383 | Presence of GCs and villi formation | Protein abundance of PEPT1 and OATP2B1 | Limited degree of differentiation, poor PXR- and VDR-mediated drug response, poor abundance of OATP2B1, UGT1A6 not expressed, colonic origin | HTS |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Przybylla, R.; Krohn, M.; Sellin, M.-L.; Frank, M.; Oswald, S.; Linnebacher, M. Novel In Vitro Models for Cell Differentiation and Drug Transport Studies of the Human Intestine. Cells 2023, 12, 2371. https://doi.org/10.3390/cells12192371
Przybylla R, Krohn M, Sellin M-L, Frank M, Oswald S, Linnebacher M. Novel In Vitro Models for Cell Differentiation and Drug Transport Studies of the Human Intestine. Cells. 2023; 12(19):2371. https://doi.org/10.3390/cells12192371
Chicago/Turabian StylePrzybylla, Randy, Mathias Krohn, Marie-Luise Sellin, Marcus Frank, Stefan Oswald, and Michael Linnebacher. 2023. "Novel In Vitro Models for Cell Differentiation and Drug Transport Studies of the Human Intestine" Cells 12, no. 19: 2371. https://doi.org/10.3390/cells12192371
APA StylePrzybylla, R., Krohn, M., Sellin, M.-L., Frank, M., Oswald, S., & Linnebacher, M. (2023). Novel In Vitro Models for Cell Differentiation and Drug Transport Studies of the Human Intestine. Cells, 12(19), 2371. https://doi.org/10.3390/cells12192371