

RIPOR2 Expression Decreased by HPV-16 E6 and E7 Oncoproteins: An Opportunity in the Search for Prognostic Biomarkers in Cervical Cancer


 , , , ,
, , , ,
Abstract
1. Introduction
2. Materials and Methods
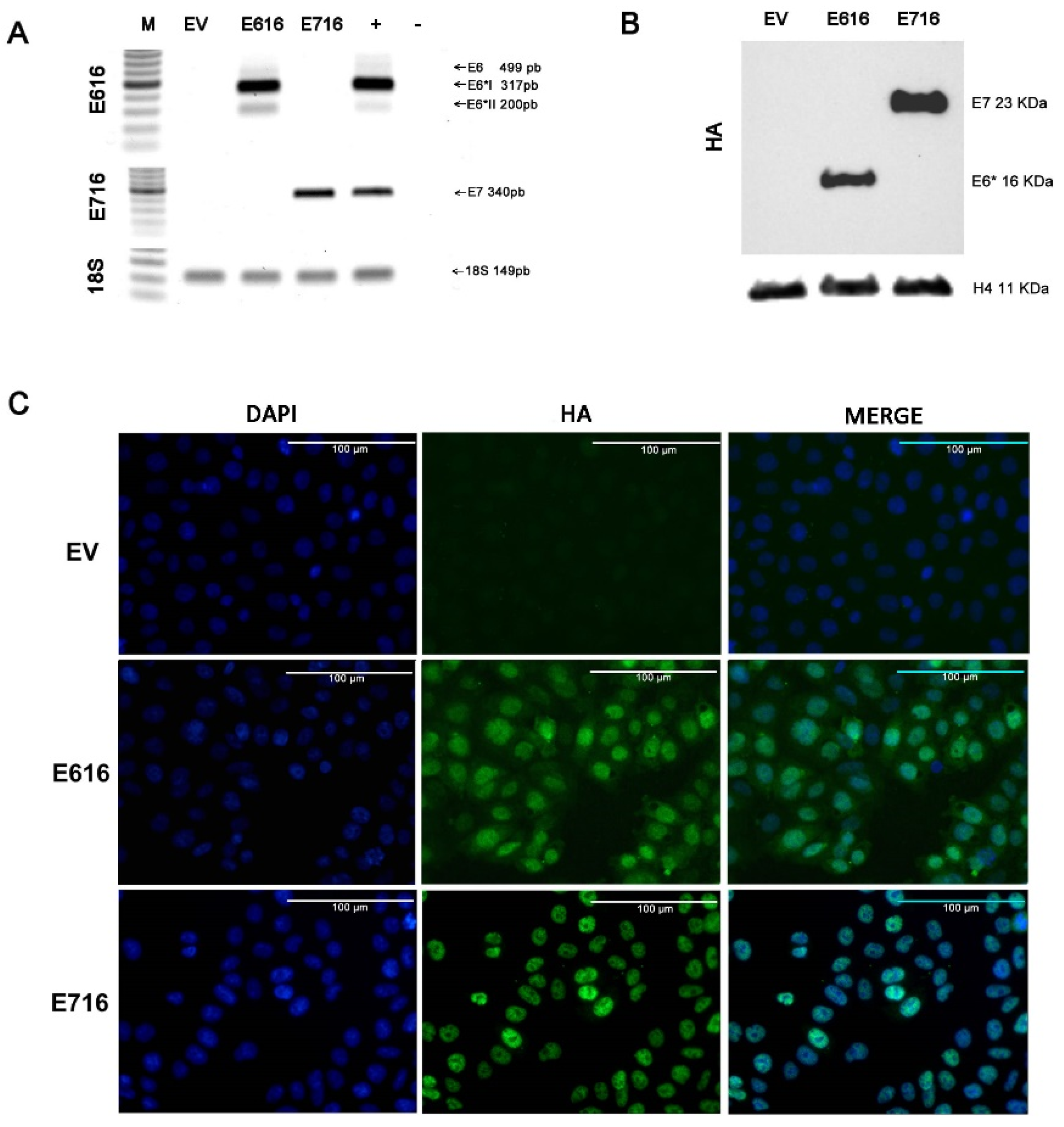
2.1. Cell Lines and Culture
2.2. Plasmids
2.3. Western Blotting
2.4. Immunofluorescence Staining
2.5. RNA Sequencing and Data Analysis
2.6. TCGA Analysis
2.7. Real-Time Quantitative PCR
2.8. Cervical Samples
2.9. Statistical Analysis
3. Results
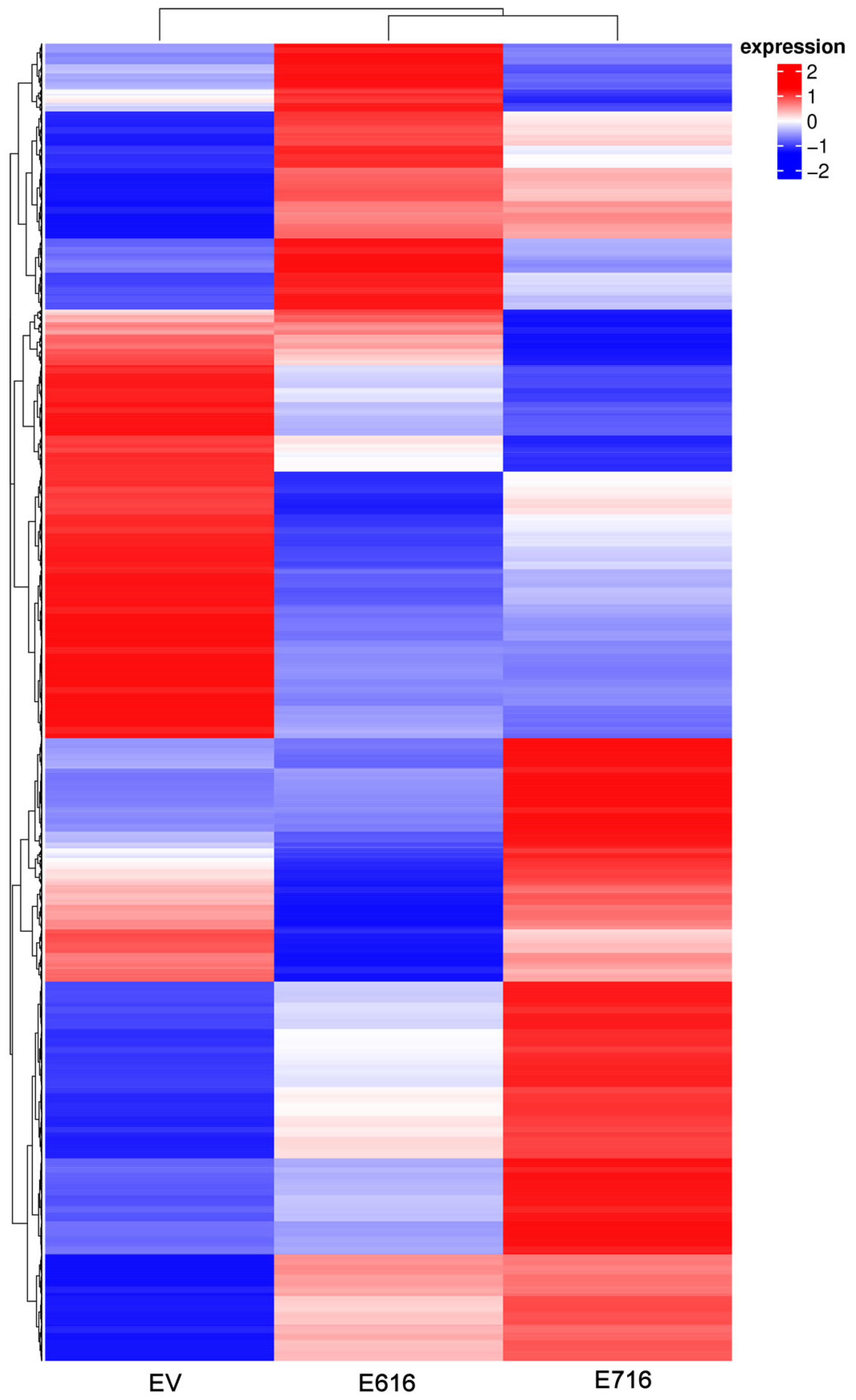
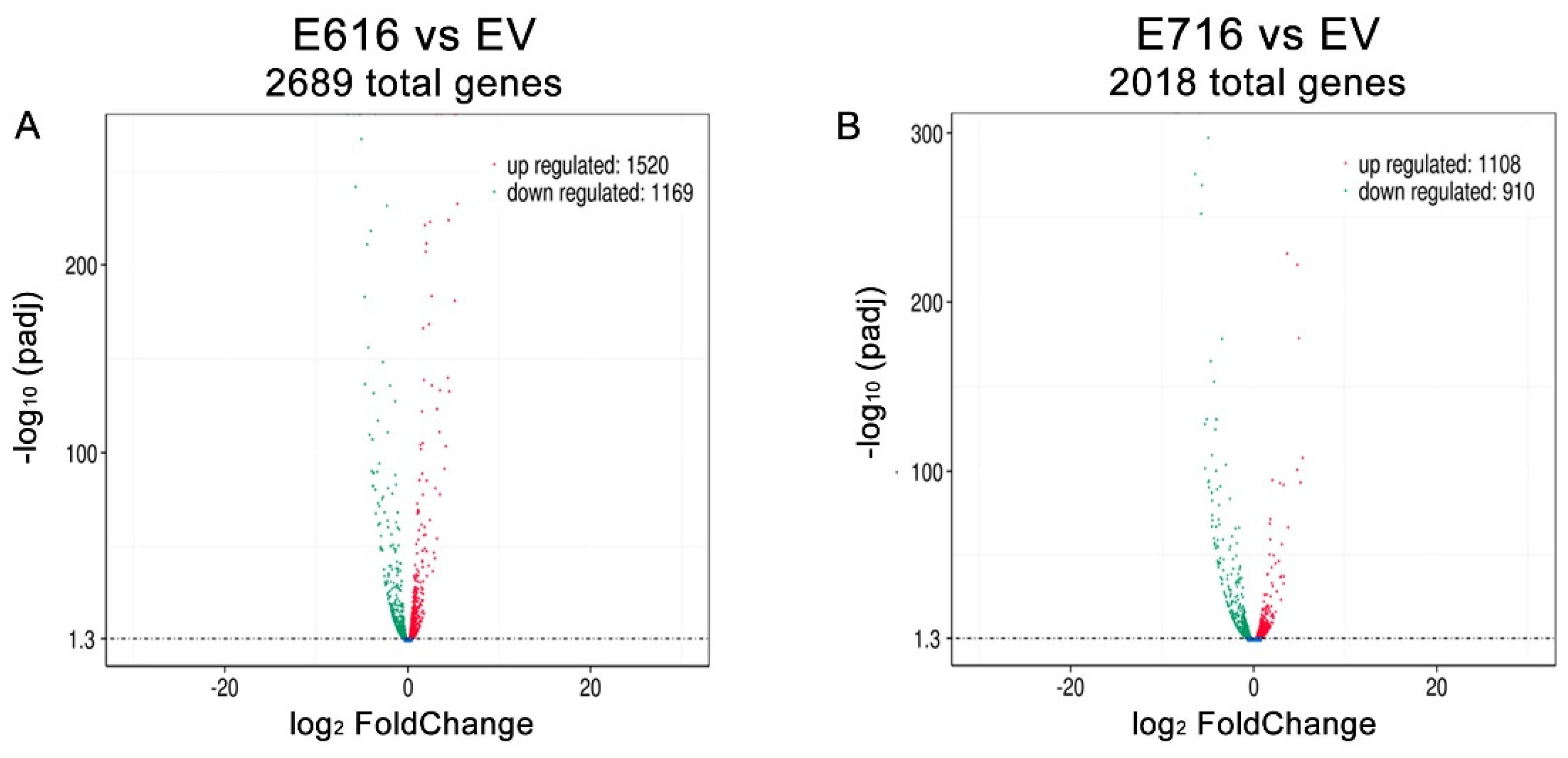
3.1. HPV-16 E6 and E7 Oncoproteins Differentially Modify Transcriptome of Cervical Cancer Cells
3.2. Cellular Processes and Signaling Pathways Modified by E6 and E7
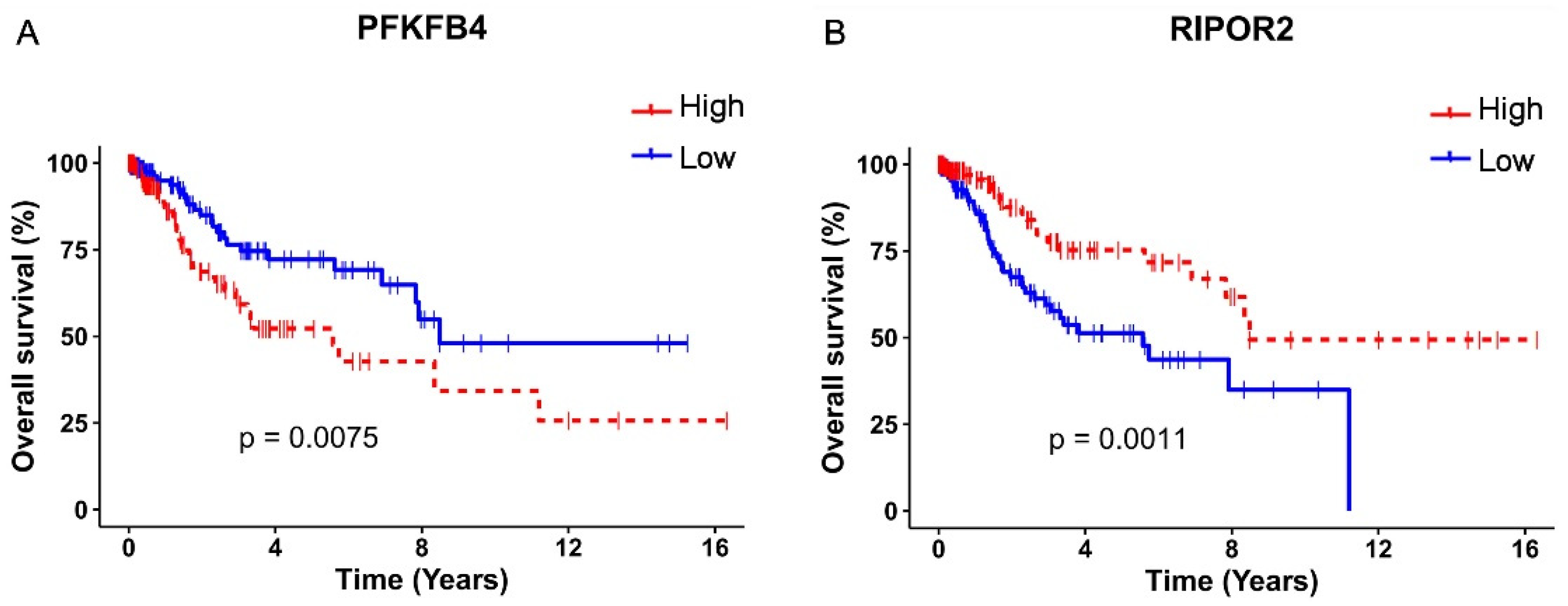
3.3. E616 and E716 Regulated Genes Involved in Overall Survival of Cervical Cancer Patients
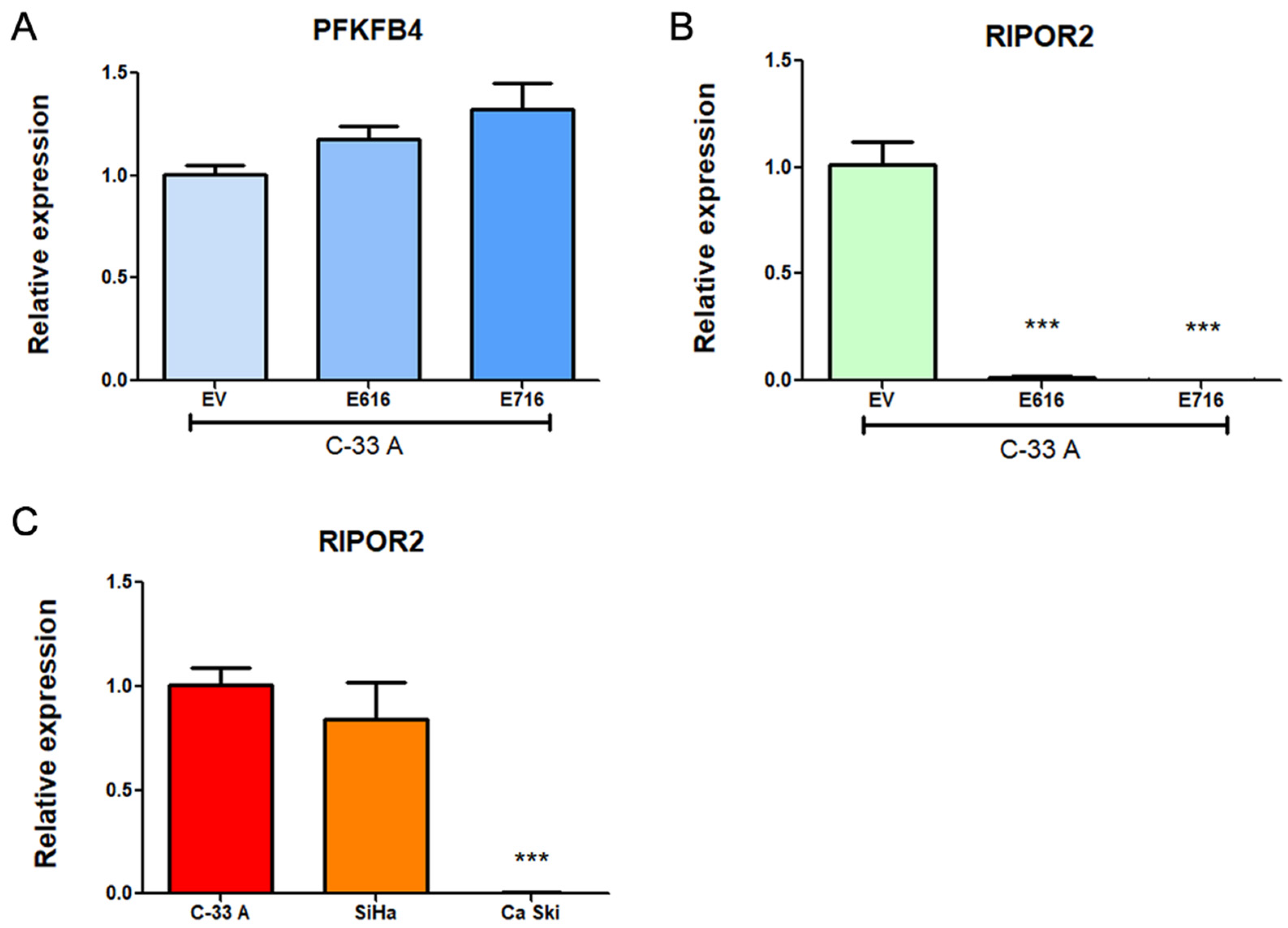
3.4. PFKFB4 and RIPOR2 Transcripts Are Affected by E6 and E7 in Cervical Cancer Cells
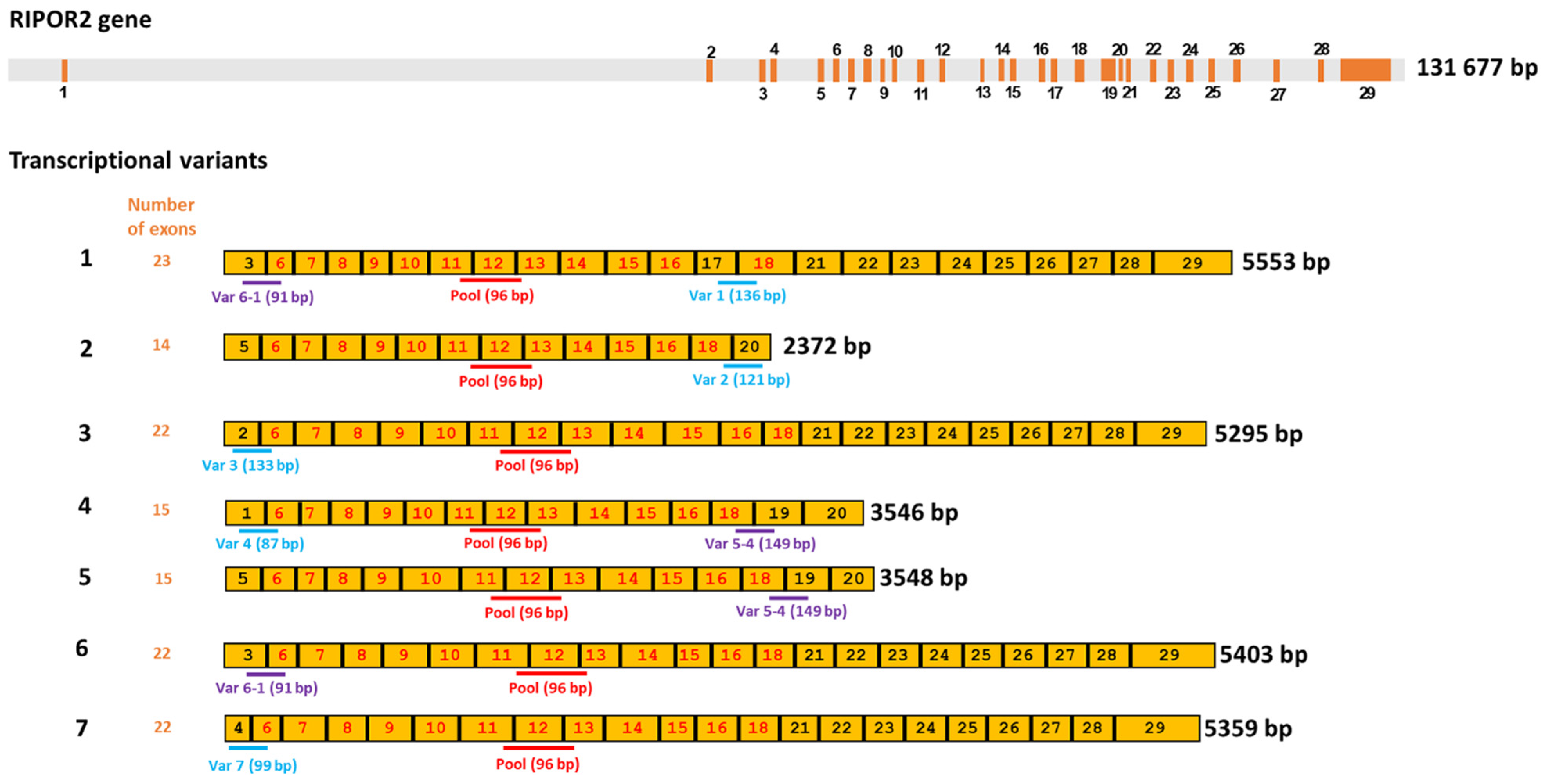
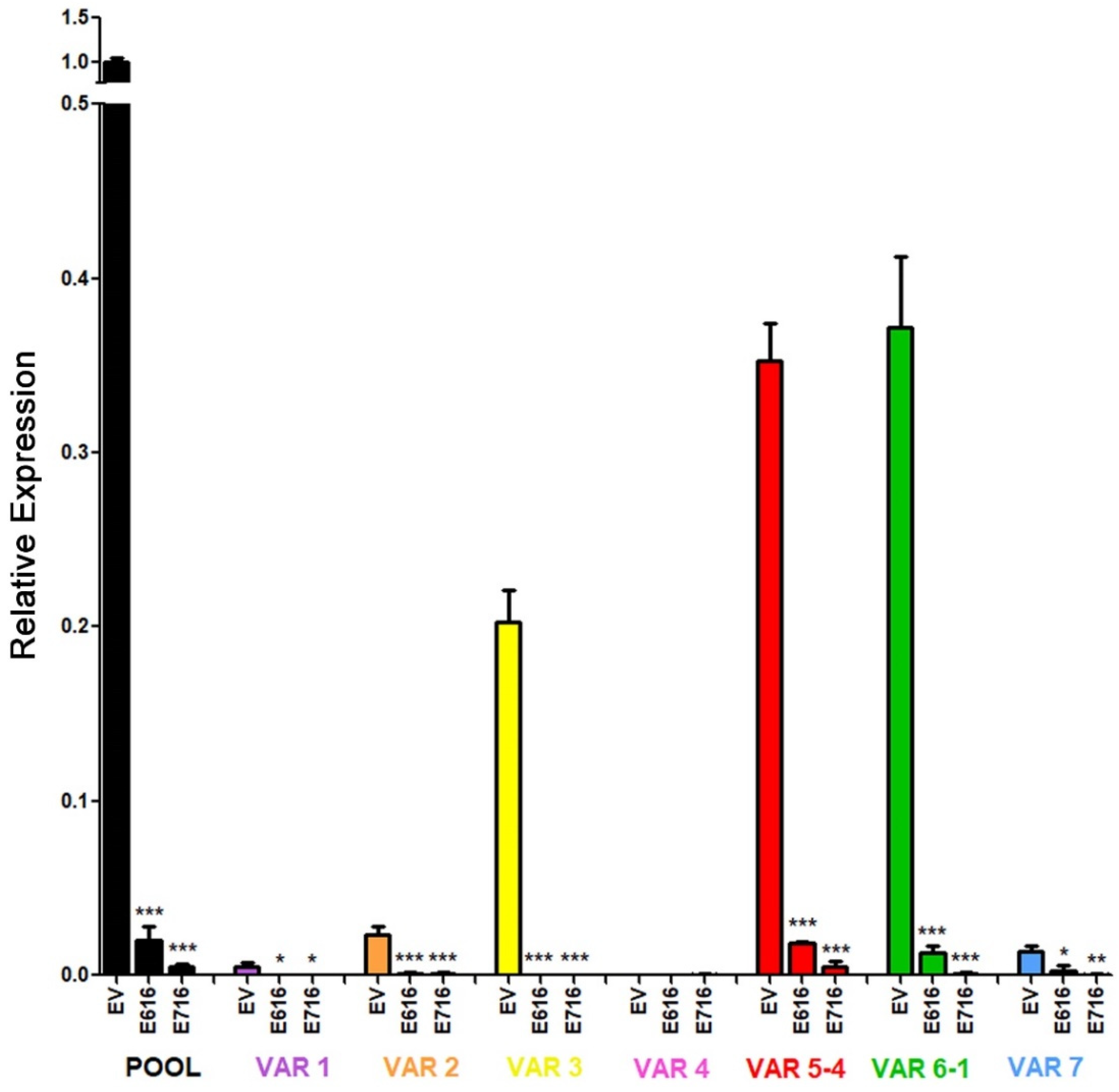
3.5. HPV-16 E6 and E7 Oncoproteins Decrease the Levels of Six Transcriptional Variants of RIPOR2 in C-33 A Cells
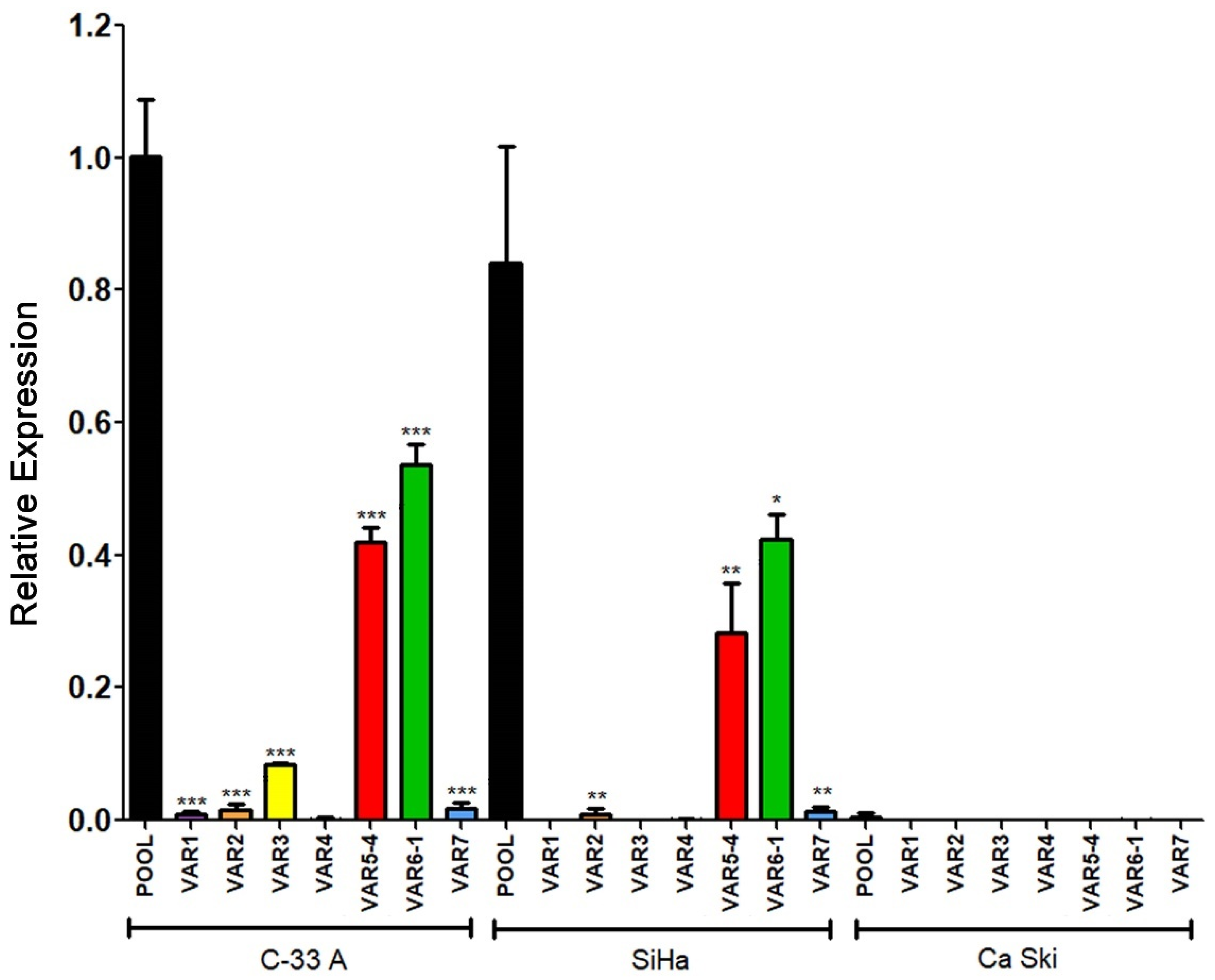
3.6. RIPOR2 Expression Is Downregulated in Premalignant Lesions and Lower Levels of RIPOR2 Are Associated with Worse Prognosis of Cervical Cancer
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ferlay, J.; Ervik, M.; Lam, F.; Colombet, M.; Mery, L.; Piñeros, M.; Znaor, A.; Soerjomataram, I.B.F. IARC: Cancer Today. Available online: https://gco.iarc.fr/today/home (accessed on 29 June 2022).
- Li, N.; Franceschi, S.; Howell-Jones, R.; Snijders, P.J.F.; Clifford, G.M. Human Papillomavirus Type Distribution in 30,848 Invasive Cervical Cancers Worldwide: Variation by Geographical Region, Histological Type and Year of Publication. Int. J. cancer 2011, 128, 927–935. [Google Scholar] [CrossRef]
- Alfaro, A.; Juárez-Torres, E.; Medina-Martínez, I.; Mateos-Guerrero, N.; Bautista-Huerta, M.; Román-Bassaure, E.; Villegas-Sepúlveda, N.; Berumen, J. Different Association of Human Papillomavirus 16 Variants with Early and Late Presentation of Cervical Cancer. PLoS ONE 2016, 11, e0169315. [Google Scholar] [CrossRef]
- Pal, A.; Kundu, R. Human Papillomavirus E6 and E7: The Cervical Cancer Hallmarks and Targets for Therapy. Front. Microbiol. 2020, 10, 3116. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Zapien, D.; Ruiz, F.X.; Poirson, J.; Mitschler, A.; Ramirez, J.; Forster, A.; Cousido-Siah, A.; Masson, M.; Pol, S.V.; Podjarny, A.; et al. Structure of the E6/E6AP/P53 Complex Required for HPV-Mediated Degradation of P53. Nature 2016, 529, 541–545. [Google Scholar] [CrossRef] [PubMed]
- Scheffner, M.; Huibregtse, J.M.; Vierstra, R.D.; Howley, P.M. The HPV-16 E6 and E6-AP Complex Functions as a Ubiquitin-Protein Ligase in the Ubiquitination of P53. Cell 1993, 75, 495–505. [Google Scholar] [CrossRef] [PubMed]
- Huh, K.; Zhou, X.; Hayakawa, H.; Cho, J.-Y.; Libermann, T.A.; Jin, J.; Wade Harper, J.; Munger, K. Human Papillomavirus Type 16 E7 Oncoprotein Associates with the Cullin 2 Ubiquitin Ligase Complex, Which Contributes to Degradation of the Retinoblastoma Tumor Suppressor. J. Virol. 2007, 81, 9737–9747. [Google Scholar] [CrossRef]
- Singh, G.K.; Azuine, R.E.; Siahpush, M. Global Inequalities in Cervical Cancer Incidence and Mortality Are Linked to Deprivation, Low Socioeconomic Status, and Human Development. Int. J. MCH AIDS 2012, 1, 17–30. [Google Scholar] [CrossRef] [PubMed]
- Isla-Ortiz, D.; Palomares-Castillo, E.; Mille-Loera, J.E.; Ramírez-Calderón, N.; Mohar-Betancourt, A.; Meneses-García, A.A.; Reynoso-Noverón, N. Cervical Cancer in Young Women: Do They Have a Worse Prognosis? A Retrospective Cohort Analysis in a Population of Mexico. Oncologist 2020, 25, e1363–e1371. [Google Scholar] [CrossRef]
- Torreglosa-Hernández, S.; Grisales-Romero, H.; Morales-Carmona, E.; Hernández-Ávila, J.E.; Huerta-Gutiérrez, R.; Barquet-Muñoz, S.A.; Palacio-Mejía, L.S. Supervivencia y Factores Asociados En Pacientes Con Cáncer Cervicouterino Atendidas Por El Seguro Popular En México. Salud Publica Mex. 2022, 64, 76–86. [Google Scholar] [CrossRef]
- Piri, R.; Ghaffari, A.; Gholami, N.; Azami-Aghdash, S.; PourAli-Akbar, Y.; Saleh, P.; Naghavi-Behzad, M. Ki-67/MIB-1 as a Prognostic Marker in Cervical Cancer—A Systematic Review with Meta-Analysis. Asian Pac. J. Cancer Prev. 2015, 16, 6997–7002. [Google Scholar] [CrossRef]
- Zhu, K.; Deng, C.; Du, P.; Liu, T.; Piao, J.; Piao, Y.; Yang, M.; Chen, L. G6PC Indicated Poor Prognosis in Cervical Cancer and Promoted Cervical Carcinogenesis in Vitro and in Vivo. Reprod. Biol. Endocrinol. 2022, 20, 50. [Google Scholar] [CrossRef]
- Dong, Z.; Chang, X.; Xie, L.; Wang, Y.; Hou, Y. Increased Expression of SRPK1 (Serine/Arginine-Rich Protein-Specific Kinase 1) Is Associated with Progression and Unfavorable Prognosis in Cervical Squamous Cell Carcinoma. Bioengineered 2022, 13, 6100–6112. [Google Scholar] [CrossRef] [PubMed]
- Beyer, S.; Wehrmann, M.; Meister, S.; Kolben, T.M.; Trillsch, F.; Burges, A.; Czogalla, B.; Schmoeckel, E.; Mahner, S.; Jeschke, U.; et al. Galectin-8 and -9 as Prognostic Factors for Cervical Cancer. Arch. Gynecol. Obstet. 2022, 306, 105. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.; Ma, R.; Hu, T.; Xiao, G.G.; Wu, C. Bioinformatics Analysis Highlights Five Differentially Expressed Genes as Prognostic Biomarkers of Cervical Cancer and Novel Option for Anticancer Treatment. Front. Cell. Infect. Microbiol. 2022, 12, 926348. [Google Scholar] [CrossRef]
- Campos-Parra, A.D.; Pérez-Quintanilla, M.; Martínez-Gutierrez, A.D.; Pérez-Montiel, D.; Coronel-Martínez, J.; Millan-Catalan, O.; De León, D.C.; Pérez-Plasencia, C. Molecular Differences between Squamous Cell Carcinoma and Adenocarcinoma Cervical Cancer Subtypes: Potential Prognostic Biomarkers. Curr. Oncol. 2022, 29, 4689–4702. [Google Scholar] [CrossRef] [PubMed]
- Paik, E.S.; Chang, C.S.; Chae, Y.L.; Oh, S.Y.; Byeon, S.J.; Kim, C.J.; Lee, Y.Y.; Kim, T.J.; Lee, J.W.; Kim, B.G.; et al. Prognostic Relevance of BRCA1 Expression in Survival of Patients With Cervical Cancer. Front. Oncol. 2021, 11, 770103. [Google Scholar] [CrossRef] [PubMed]
- Patel, K.A.; Patel, B.M.; Thobias, A.R.; Gokani, R.A.; Chhikara, A.B.; Desai, A.D.; Patel, P.S. Overexpression of VEGF165 Is Associated with Poor Prognosis of Cervical Cancer. J. Obstet. Gynaecol. Res. 2020, 46, 2397–2406. [Google Scholar] [CrossRef]
- Nahand, J.S.; Taghizadeh-boroujeni, S.; Karimzadeh, M.; Borran, S.; Pourhanifeh, M.H.; Moghoofei, M.; Bokharaei-Salim, F.; Karampoor, S.; Jafari, A.; Asemi, Z.; et al. MicroRNAs: New Prognostic, Diagnostic, and Therapeutic Biomarkers in Cervical Cancer. J. Cell. Physiol. 2019, 234, 17064–17099. [Google Scholar] [CrossRef]
- Zhang, G.; Zhang, R.; Bai, P.; Li, S.; Zuo, J.; Zhang, Y.; Liu, M.; Wu, L. Down-Regulated Expression of MiR-99a Is Associated with Lymph Node Metastasis and Predicts Poor Outcome in Stage IB Cervical Squamous Cell Carcinoma: A Case-Control Study. Ann. Transl. Med. 2022, 10, 663. [Google Scholar] [CrossRef]
- Chang, A.; Shi, Y.; Wang, P.; Ren, J. LINC00963 May Be Associated with a Poor Prognosis in Patients with Cervical Cancer. Med. Sci. Monit. 2022, 28, e935070. [Google Scholar] [CrossRef]
- Coquillard, G.; Palao, B.; Patterson, B.K. Quantification of Intracellular HPV E6/E7 MRNA Expression Increases the Specificity and Positive Predictive Value of Cervical Cancer Screening Compared to HPV DNA. Gynecol. Oncol. 2011, 120, 89–93. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.M.; Warke, H.; Chaudhari, H.; Mavani, P.; Katke, R.D.; Kerkar, S.C.; Mania-Pramanik, J. Human Papillomavirus E6/E7 Oncogene Transcripts as Biomarkers for the Early Detection of Cervical Cancer. J. Med. Virol. 2022, 94, 3368–3375. [Google Scholar] [CrossRef]
- Ho, C.M.; Lee, B.H.; Chang, S.F.; Chien, T.Y.; Huang, S.H.; Yan, C.C.; Cheng, W.F. Type-Specific Human Papillomavirus Oncogene Messenger RNA Levels Correlate with the Severity of Cervical Neoplasia. Int. J. cancer 2010, 127, 622–632. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, F.J.; Inkman, M.; Rashmi, R.; Muhammad, N.; Gabriel, N.; Miller, C.A.; McLellan, M.D.; Goldstein, M.; Markovina, S.; Grigsby, P.W.; et al. HPV Transcript Expression Affects Cervical Cancer Response to Chemoradiation. JCI insight 2021, 6, e138734. [Google Scholar] [CrossRef]
- Rose, B.R.; Thompson, C.H.; Jiang, X.M.; Tattersall, M.H.N.; Elliott, P.M.; Dalrymple, C.; Cossart, Y.E. Detection of Human Papillomavirus Type 16 E6/E7 Transcripts in Histologically Cancer-Free Pelvic Lymph Nodes of Patients with Cervical Carcinoma. Gynecol. Oncol. 1994, 52, 212–217. [Google Scholar] [CrossRef] [PubMed]
- Dürst, M.; Hoyer, H.; Altgassen, C.; Greinke, C.; Häfner, N.; Fishta, A.; Gajda, M.; Mahnert, U.; Hillemanns, P.; Dimpfl, T.; et al. Prognostic Value of HPV-mRNA in Sentinel Lymph Nodes of Cervical Cancer Patients with pN0-Status. Oncotarget 2015, 6, 23015–23025. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Gene Ontology Resource. Available online: http://geneontology.org/ (accessed on 5 October 2022).
- KEGG: Kyoto Encyclopedia of Genes and Genomes. Available online: https://www.genome.jp/kegg/ (accessed on 5 October 2022).
- Home—Reactome Pathway Database. Available online: https://reactome.org/ (accessed on 5 October 2022).
- TCGAbiolinks: An R/Bioconductor Package for Integrative Analysis of TCGA Data|Nucleic Acids Research|Oxford Academic. Available online: https://academic.oup.com/nar/article/44/8/e71/2465925 (accessed on 5 October 2022).
- Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2|Genome Biology|Full Text. Available online: https://genomebiology.biomedcentral.com/articles/10.1186/s13059-014-0550-8 (accessed on 5 October 2022).
- Yu, L.; Zheng, Z.M. Human Papillomavirus Type 16 Circular RNA Is Barely Detectable for the Claimed Biological Activity. MBio 2022, 13, e0359421. [Google Scholar] [CrossRef] [PubMed]
- Paget-Bailly, P.; Meznad, K.; Bruyère, D.; Perrard, J.; Herfs, M.; Jung, A.C.; Mougin, C.; Prétet, J.L.; Baguet, A. Comparative RNA Sequencing Reveals That HPV16 E6 Abrogates the Effect of E6*I on ROS Metabolism. Sci. Rep. 2019, 9, 5938. [Google Scholar] [CrossRef] [PubMed]
- Mincheva, A.; Gissmann, L.; Hausen, H.Z. Chromosomal integration sites of human papillomavirus DNA in three cervical cancer cell lines mapped by in situ hybridization. Med. Microbiol. Immunol. 1987, 176, 245–256. [Google Scholar]
- NCBI. RIPOR2 RHO Family Interacting Cell Polarization Regulator 2 [Homo Sapiens (Human)]-Gene-NCBI. Available online: https://www.ncbi.nlm.nih.gov/gene/9750 (accessed on 5 October 2022).
- Dakour, J.; Li, H.; Morrish, D.W. PL48: A Novel Gene Associated with Cytotrophoblast and Lineage-Specific HL-60 Cell Differentiation. Gene 1997, 185, 153–157. [Google Scholar] [CrossRef]
- Yoon, S.; Molloy, M.J.; Wu, M.P.; Cowan, D.B.; Gussoni, E. C6ORF32 Is Upregulated during Muscle Cell Differentiation and Induces the Formation of Cellular Filopodia. Dev. Biol. 2007, 301, 70–81. [Google Scholar] [CrossRef]
- Ling, K.; Yang, L.; Yang, N.; Chen, M.; Wang, Y.; Liang, S.; Li, Y.; Jiang, L.; Yan, P.; Liang, Z. Gene Targeting of HPV18 E6 and E7 Synchronously by Nonviral Transfection of CRISPR/Cas9 System in Cervical Cancer. Hum. Gene Ther. 2020, 31, 297–308. [Google Scholar] [CrossRef]
- Zhen, S.; Liu, Y.; Lu, J.; Tuo, X.; Yang, X.; Chen, H.; Chen, W.; Li, X. Human Papillomavirus Oncogene Manipulation Using Clustered Regularly Interspersed Short Palindromic Repeats/Cas9 Delivered by PH-Sensitive Cationic Liposomes. Hum. Gene Ther. 2020, 31, 309–324. [Google Scholar] [CrossRef] [PubMed]
- Xiong, J.; Li, G.; Mei, X.; Ding, J.; Shen, H.; Zhu, D.; Wang, H. Co-Delivery of P53 Restored and E7 Targeted Nucleic Acids by Poly (Beta-Amino Ester) Complex Nanoparticles for the Treatment of HPV Related Cervical Lesions. Front. Pharmacol. 2022, 13, 826771. [Google Scholar] [CrossRef]
- Volkova, L.V.; Pashov, A.I.; Omelchuk, N.N. Cervical Carcinoma: Oncobiology and Biomarkers. Int. J. Mol. Sci. 2021, 22, 12571. [Google Scholar] [CrossRef] [PubMed]
- Dong, A.; Xu, B.; Wang, Z.; Miao, X. Survival-related DLEU1 Is Associated with HPV Infection Status and Serves as a Biomarker in HPV-infected Cervical Cancer. Mol. Med. Rep. 2022, 25, 77. [Google Scholar] [CrossRef] [PubMed]
- Yalcin, A.; Telang, S.; Clem, B.; Chesney, J. Regulation of Glucose Metabolism by 6-Phosphofructo-2-Kinase/Fructose-2,6-Bisphosphatases in Cancer. Exp. Mol. Pathol. 2009, 86, 174–179. [Google Scholar] [CrossRef]
- Cai, Y.C.; Yang, H.; Shan, H.B.; Su, H.F.; Jiang, W.Q.; Shi, Y.X. PFKFB4 Overexpression Facilitates Proliferation by Promoting the G1/S Transition and Is Associated with a Poor Prognosis in Triple-Negative Breast Cancer. Dis. Markers 2021, 2021, 8824589. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Wu, X.; Li, Y.; Cao, X.; Zhang, C.; Gao, Y. PFKFB4 as a Promising Biomarker to Predict a Poor Prognosis in Patients with Gastric Cancer. Oncol. Lett. 2021, 21, 296. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Fan, Y.; Qiu, B.; Lou, M.; Liu, X.; Yuan, K.; Tong, J. Effect of PFKFB4 on the Prognosis and Immune Regulation of NSCLC and Its Mechanism. Int. J. Gen. Med. 2022, 15, 6341–6353. [Google Scholar] [CrossRef]
- Trojan, S.E.; Piwowar, M.; Ostrowska, B.; Laidler, P.; Kocemba-Pilarczyk, K.A. Analysis of Malignant Melanoma Cell Lines Exposed to Hypoxia Reveals the Importance of PFKFB4 Overexpression for Disease Progression. Anticancer Res. 2018, 38, 6745–6752. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.; Chen, S.; You, Z.; Xie, C.; Huang, S.; Hu, X. PFKFB4 Negatively Regulated the Expression of Histone Acetyltransferase GCN5 to Mediate the Tumorigenesis of Thyroid Cancer. Dev. Growth Differ. 2020, 62, 129–138. [Google Scholar] [CrossRef]
- Zhang, H.; Lu, C.; Fang, M.; Yan, W.; Chen, M.; Ji, Y.; He, S.; Liu, T.; Chen, T.; Xiao, J. HIF-1α Activates Hypoxia-Induced PFKFB4 Expression in Human Bladder Cancer Cells. Biochem. Biophys. Res. Commun. 2016, 476, 146–152. [Google Scholar] [CrossRef]
- Lv, Z.; Ding, Y.; Cao, W.; Wang, S.; Gao, K. Role of RHO Family Interacting Cell Polarization Regulators (RIPORs) in Health and Disease: Recent Advances and Prospects. Int. J. Biol. Sci. 2022, 18, 800–808. [Google Scholar] [CrossRef]
- Zhang, K.; Waxman, D.J. PC3 Prostate Tumor-Initiating Cells with Molecular Profile FAM65Bhigh/MFI2low/LEF1low Increase Tumor Angiogenesis. Mol. Cancer 2010, 9, 319. [Google Scholar] [CrossRef]
- Mei, J.; Xing, Y.; Lv, J.; Gu, D.; Pan, J.; Zhang, Y.; Liu, J. Construction of an Immune-Related Gene Signature for Prediction of Prognosis in Patients with Cervical Cancer. Int. Immunopharmacol. 2020, 88, 106882. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Zou, C.; Gao, Y.; Shen, J.; Liu, T.; He, Q.; Li, S.; Xu, S. Comprehensive Analyses Identify RIPOR2 as a Genomic Instability-Associated Immune Prognostic Biomarker in Cervical Cancer. Front. Immunol. 2022, 13, 930488. [Google Scholar] [CrossRef] [PubMed]
- Rougerie, P.; Largeteau, Q.; Megrelis, L.; Carrette, F.; Lejeune, T.; Toffali, L.; Rossi, B.; Zeghouf, M.; Cherfils, J.; Constantin, G.; et al. Fam65b Is a New Transcriptional Target of FOXO1 That Regulates RhoA Signaling for T Lymphocyte Migration. J. Immunol. 2013, 190, 748–755. [Google Scholar] [CrossRef]
- Zhang, B.; Gui, L.S.; Zhao, X.L.; Zhu, L.L.; Li, Q.W. FOXO1 Is a Tumor Suppressor in Cervical Cancer. Genet. Mol. Res. 2015, 14, 6605–6616. [Google Scholar] [CrossRef] [PubMed]
- Chay, D.B.; Han, G.H.; Nam, S.; Cho, H.; Chung, J.Y.; Hewitt, S.M. Forkhead Box Protein O1 (FOXO1) and Paired Box Gene 3 (PAX3) Overexpression Is Associated with Poor Prognosis in Patients with Cervical Cancer. Int. J. Clin. Oncol. 2019, 24, 1429–1439. [Google Scholar] [CrossRef] [PubMed]
- Javadi, H.; Lotfi, A.S.; Hosseinkhani, S.; Mehrani, H.; Amani, J.; Soheili, Z.S.; Hojati, Z.; Kamali, M. The Combinational Effect of E6/E7 SiRNA and Anti-MiR-182 on Apoptosis Induction in HPV16-Positive Cervical Cells. Artif. Cells Nanomed. Biotechnol. 2018, 46, 727–736. [Google Scholar] [CrossRef]
- Tanaka, K.; Matsumoto, Y.; Ishikawa, H.; Fukumitsu, N.; Numajiri, H.; Murofushi, K.; Oshiro, Y.; Okumura, T.; Satoh, T.; Sakurai, H. Impact of RhoA Overexpression on Clinical Outcomes in Cervical Squamous Cell Carcinoma Treated with Concurrent Chemoradiotherapy. J. Radiat. Res. 2020, 61, 221–230. [Google Scholar] [CrossRef]
- Hampson, L.; Li, C.; Oliver, A.W.; Kitchener, H.C.; Hampson, I.N. The PDZ Protein Tip-1 Is a Gain of Function Target of the HPV16 E6 Oncoprotein. Int. J. Oncol. 2004, 25, 1249–1256. [Google Scholar] [PubMed]
- Charette, S.T.; McCance, D.J. The E7 Protein from Human Papillomavirus Type 16 Enhances Keratinocyte Migration in an Akt-Dependent Manner. Oncogene 2007, 26, 7386–7390. [Google Scholar] [CrossRef] [PubMed]
- Morrish, D.W.; Linetsky, E.; Bhardwaj, D.; Li, H.; Dakour, J.; Marsh, R.G.; Paterson, M.C.; Godbout, R. Identification by Subtractive Hybridization of a Spectrum of Novel and Unexpected Genes Associated with in Vitro Differentiation of Human Cytotrophoblast Cells. Placenta 1996, 17, 431–441. [Google Scholar] [CrossRef]
- Sen, P.; Ganguly, P.; Ganguly, N. Modulation of DNA Methylation by Human Papillomavirus E6 and E7 Oncoproteins in Cervical Cancer. Oncol. Lett. 2018, 15, 11–22. [Google Scholar] [CrossRef] [PubMed]
- EPD The Eukaryotic Promoter Database. Available online: https://epd.epfl.ch/search_EPDnew.php?query=ripor2&db=human (accessed on 30 September 2022).
- FANTOM. Available online: https://fantom.gsc.riken.jp/ (accessed on 5 October 2022).
- Vanhove, K.; Graulus, G.J.; Mesotten, L.; Thomeer, M.; Derveaux, E.; Noben, J.P.; Guedens, W.; Adriaensens, P. The Metabolic Landscape of Lung Cancer: New Insights in a Disturbed Glucose Metabolism. Front. Oncol. 2019, 9, 1215. [Google Scholar] [CrossRef] [PubMed]
- Hao, P.; Yu, J.; Ward, R.; Liu, Y.; Hao, Q.; An, S.; Xu, T. Eukaryotic Translation Initiation Factors as Promising Targets in Cancer Therapy. Cell Commun. Signal. 2020, 18, 175. [Google Scholar] [CrossRef]
- Zhou, W.J.; Geng, Z.H.; Chi, S.; Zhang, W.; Niu, X.F.; Lan, S.J.; Ma, L.; Yang, X.; Wang, L.J.; Ding, Y.Q.; et al. Slit-Robo Signaling Induces Malignant Transformation through Hakai-Mediated E-Cadherin Degradation during Colorectal Epithelial Cell Carcinogenesis. Cell Res. 2011, 21, 609–626. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Univariate Analysis | Multivariate Analysis | ||||
|---|---|---|---|---|---|
| Overall Survival | HR (95% CI) | p-Value | HR (95% CI) | p-Value | |
| SLC4A11 | High vs. low expression | 2 (1.2–3.5) | 0.0081 | 1.42 (0.79–2.55) | 0.228 |
| NUP188 | 2 (1.2–3.4) | 0.0097 | 1.10 (0.54–2.23) | 0.773 | |
| CREM | 2 (1.2–3.3) | 0.013 | 0.80 (0.40–1.62) | 0.55 | |
| AP1B1 | 1.9 (1.1–3.1) | 0.016 | 0.99 (0.52–1.88) | 0.99 | |
| RIPOR2 | 2.4 (1.4–4.1) | 0.0016 | 1.80 (1.00–3.25) | 0.048 | |
| PFKFB4 | 0.5 (0.3–0.84) | 0.0085 | 0.50 (0.27–0.93) | 0.029 | |
| CC2D1A | 1.9 (1.1–3.2) | 0.015 | 1.14 (0.56–2.30) | 0.704 | |
| BICDL1 | 1.9 (1.1–3.2) | 0.015 | 1.16 (0.62–2.15) | 0.629 | |
| RHOT2 | 2 (1.2–3.4) | 0.0073 | 1.44 (0.74–2.79) | 0.278 | |
| NBEAL2 | 1.9 (1.1–3.2) | 0.016 | 1.27 (0.69–2.33) | 0.436 | |
| CPNE7 | 2.2 (1.3–3.7) | 0.0033 | 1.55 (0.83–2.90) | 0.165 | |
| FARSA | 1.9 (1.1–3.2) | 0.013 | 1.15 (0.55–2.40) | 0.692 | |
| SHTN1 | 2.2 (1.3–3.7) | 0.0033 | 1.46 (0.73–2.91) | 0.281 | |
| Clinical Stage | 1.5 (1.2–1.9) | 0.0003 | |||
| Transcript | Length (nt) | Transcript Type | Protein Isoform | Length (aa) |
|---|---|---|---|---|
| 1 | 5553 | protein coding | 1 | 1068 |
| 2 | 2372 | protein coding | 2 | 591 |
| 3 | 5295 | protein coding | 3 | 1047 |
| 4 | 3546 | protein coding | 4 | 647 |
| 5 | 3548 | protein coding | 5 | 613 |
| 6 | 5403 | protein coding | 6 | 1018 |
| 7 | 5359 | protein coding | 6 | 1018 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Olmedo-Nieva, L.; Muñoz-Bello, J.O.; Martínez-Ramírez, I.; Martínez-Gutiérrez, A.D.; Ortiz-Pedraza, Y.; González-Espinosa, C.; Madrid-Marina, V.; Torres-Poveda, K.; Bahena-Roman, M.; Lizano, M. RIPOR2 Expression Decreased by HPV-16 E6 and E7 Oncoproteins: An Opportunity in the Search for Prognostic Biomarkers in Cervical Cancer. Cells 2022, 11, 3942. https://doi.org/10.3390/cells11233942
Olmedo-Nieva L, Muñoz-Bello JO, Martínez-Ramírez I, Martínez-Gutiérrez AD, Ortiz-Pedraza Y, González-Espinosa C, Madrid-Marina V, Torres-Poveda K, Bahena-Roman M, Lizano M. RIPOR2 Expression Decreased by HPV-16 E6 and E7 Oncoproteins: An Opportunity in the Search for Prognostic Biomarkers in Cervical Cancer. Cells. 2022; 11(23):3942. https://doi.org/10.3390/cells11233942
Chicago/Turabian StyleOlmedo-Nieva, Leslie, J. Omar Muñoz-Bello, Imelda Martínez-Ramírez, Antonio Daniel Martínez-Gutiérrez, Yunuen Ortiz-Pedraza, Claudia González-Espinosa, Vicente Madrid-Marina, Kirvis Torres-Poveda, Margarita Bahena-Roman, and Marcela Lizano. 2022. "RIPOR2 Expression Decreased by HPV-16 E6 and E7 Oncoproteins: An Opportunity in the Search for Prognostic Biomarkers in Cervical Cancer" Cells 11, no. 23: 3942. https://doi.org/10.3390/cells11233942
APA StyleOlmedo-Nieva, L., Muñoz-Bello, J. O., Martínez-Ramírez, I., Martínez-Gutiérrez, A. D., Ortiz-Pedraza, Y., González-Espinosa, C., Madrid-Marina, V., Torres-Poveda, K., Bahena-Roman, M., & Lizano, M. (2022). RIPOR2 Expression Decreased by HPV-16 E6 and E7 Oncoproteins: An Opportunity in the Search for Prognostic Biomarkers in Cervical Cancer. Cells, 11(23), 3942. https://doi.org/10.3390/cells11233942