
Effective Treatment of Patients Experiencing Primary, Acute HIV Infection Decreases Exhausted/Activated CD4+ T Cells and CD8+ T Memory Stem Cells

 , , , , ,
, , , , ,  , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients
2.2. Blood Samples Collection, Processing, and Storage
2.3. Sorting of Naïve and Effector Memory CD4+ T Cells
2.4. DNA Extraction and Quantification of HIV-DNA
2.5. Immunophenotyping of T Lymphocytes Using Flow Cytometry
2.6. Immunophenotyping of B Lymphocytes Using Flow Cytometry
2.7. Statistical Analysis
2.7.1. Analysis of Clinical Data
2.7.2. High-Dimensional Data Analysis of Flow Cytometry Data
2.7.3. Principal Component Analysis and Correlation Plots
3. Results
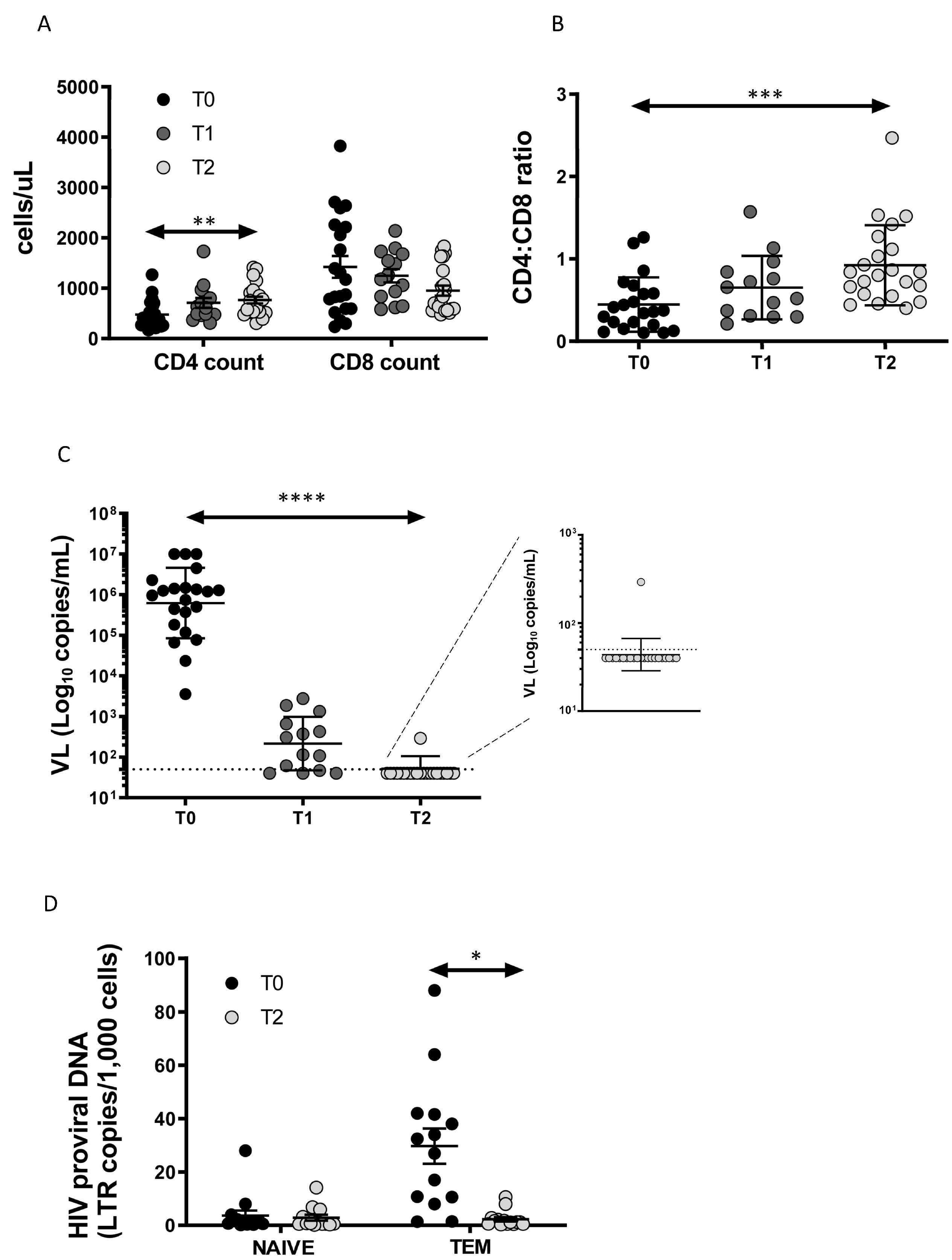
3.1. Enrolled Patients and Clinical Data
3.2. Total HIV-DNA Decrease after ART in TEM Cells
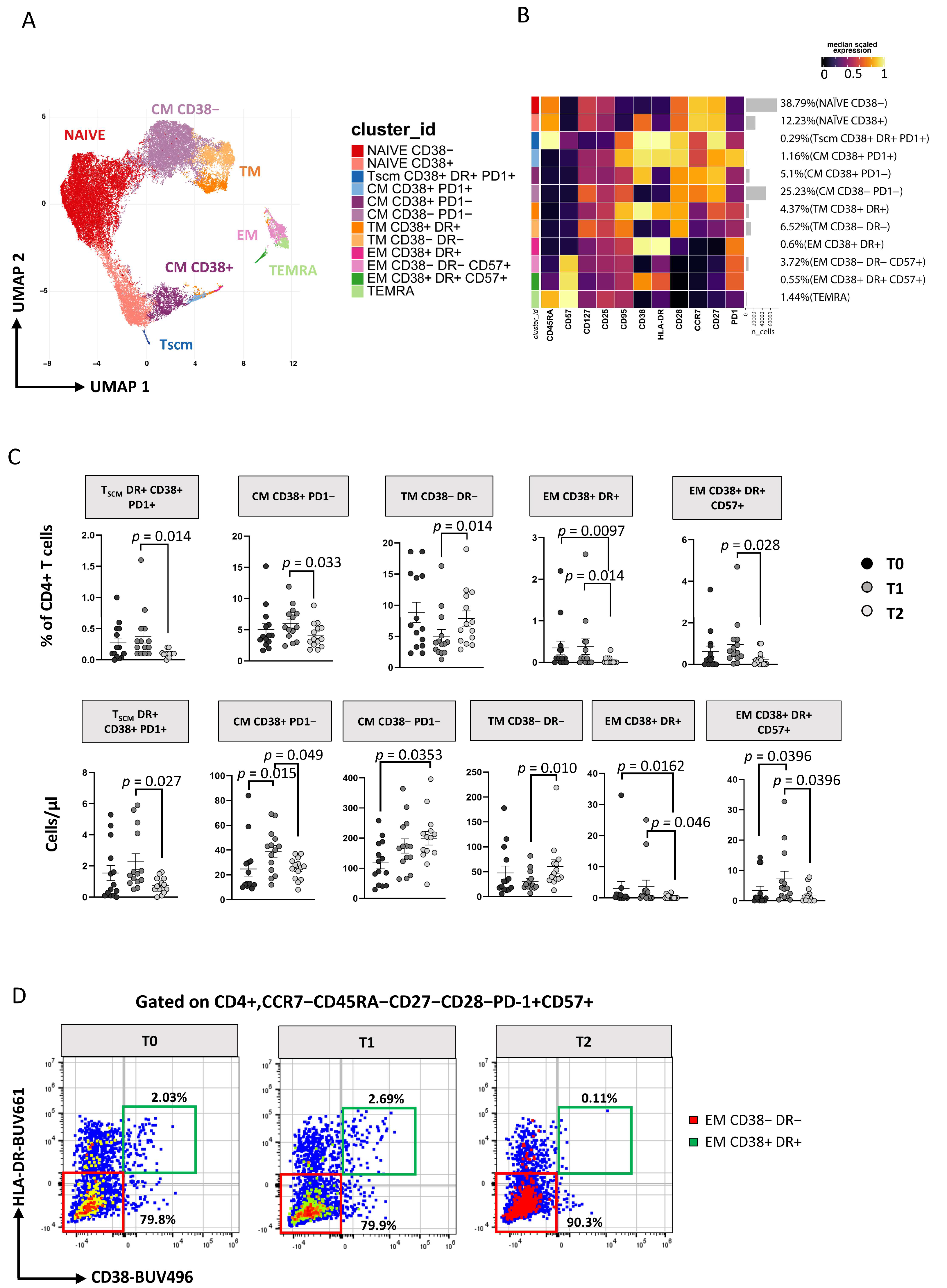
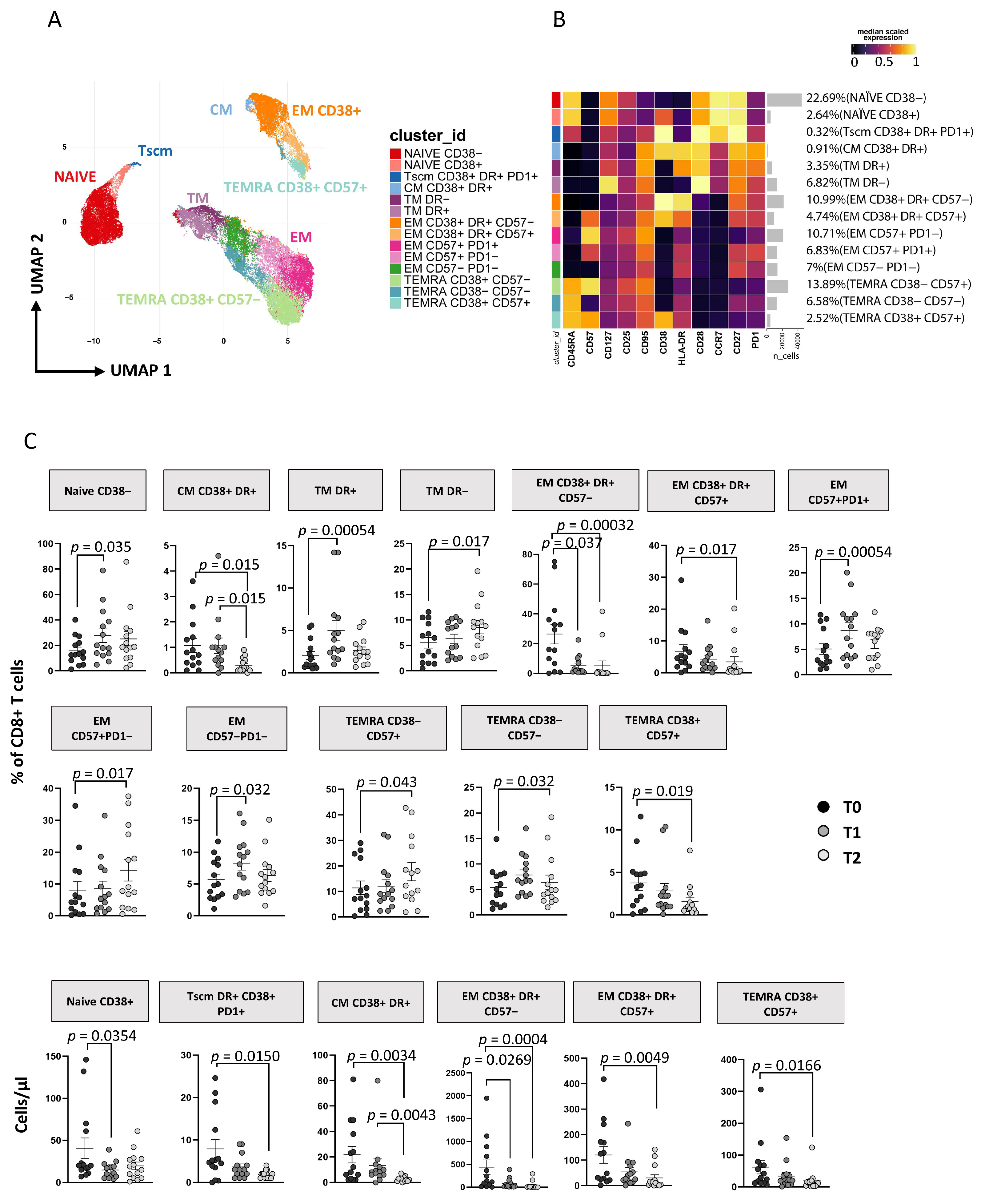
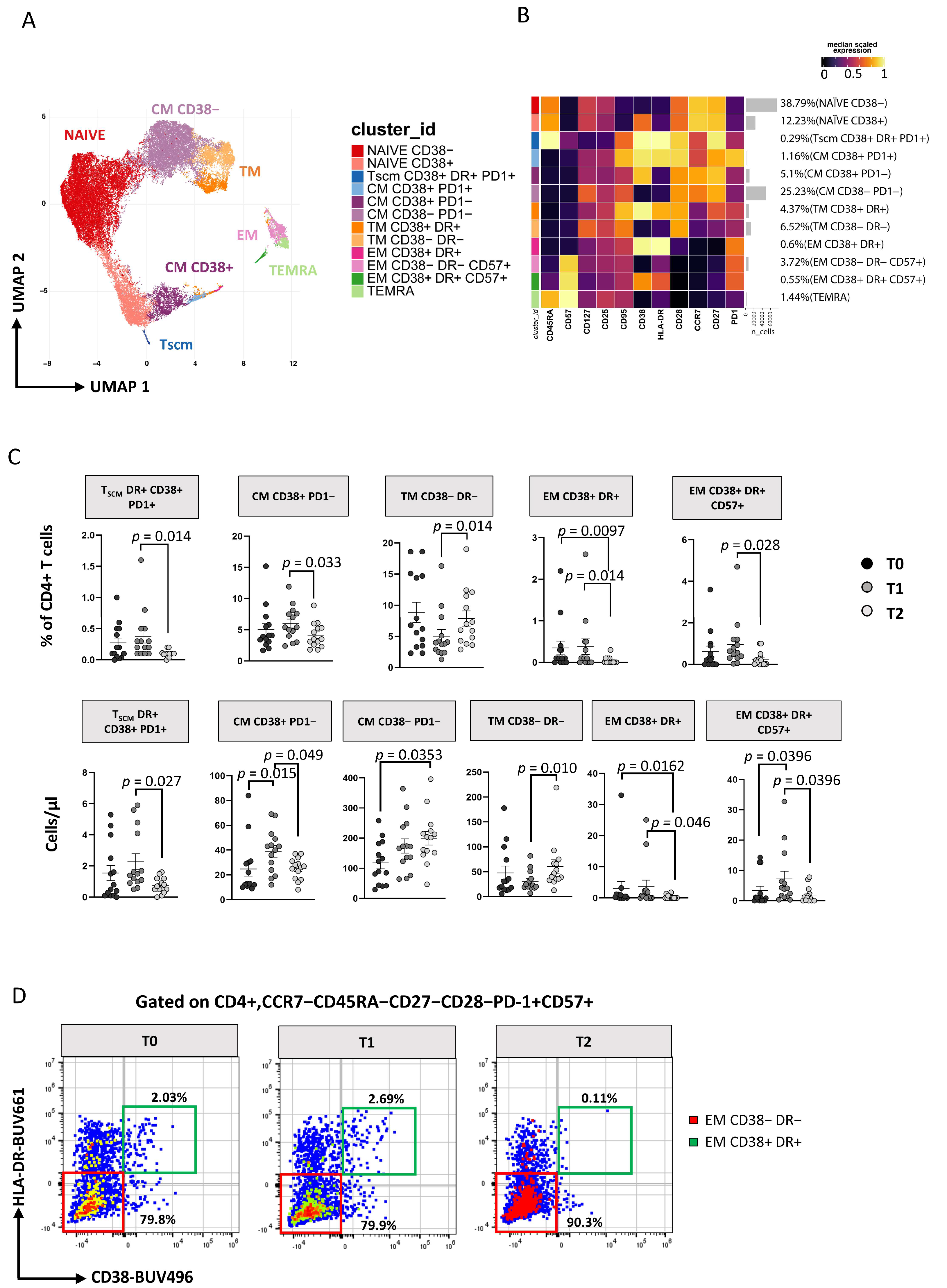
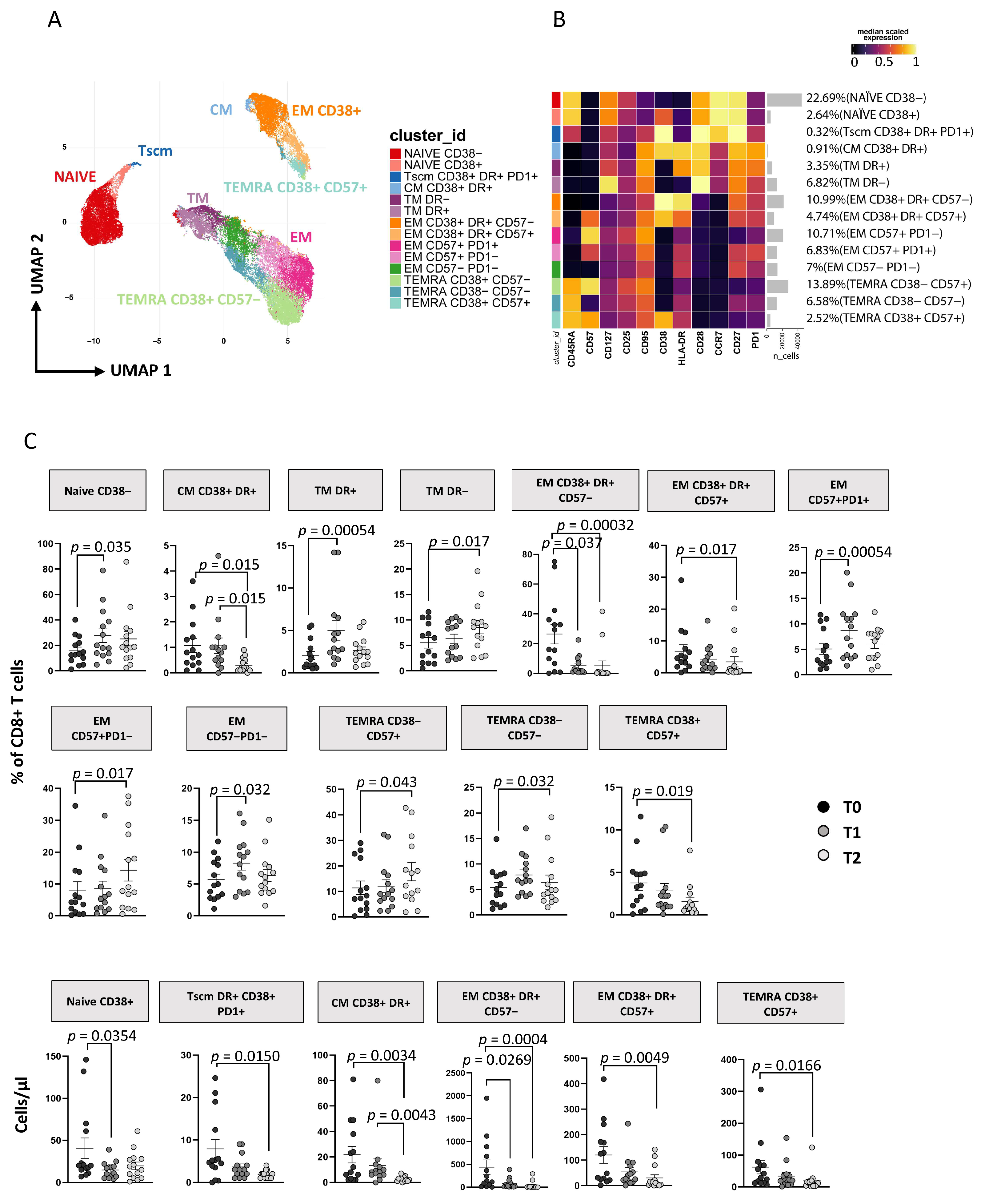
3.3. Immunophenotyping of T Lymphocytes
3.3.1. TCM and TSCM Cells Reduction within CD4 T Cell after 12 Months of ART
3.3.2. ART Is Associated with a Huge Reduction of CD8+ T Cell Activation and TSCM
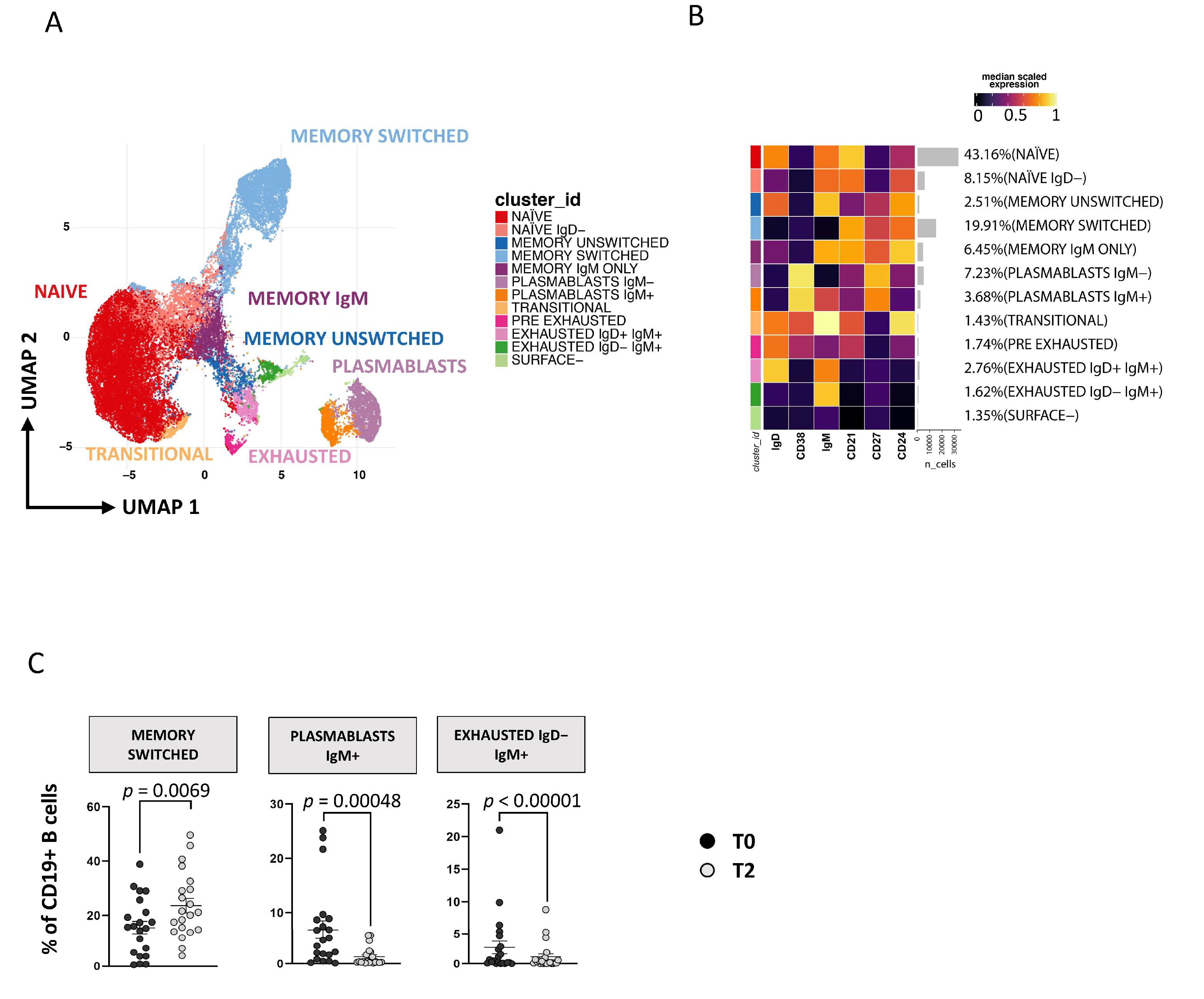
3.4. Immunophenotyping of B Lymphocytes
Plasmablasts and Exhausted B Cell Reduction Occurs following ART
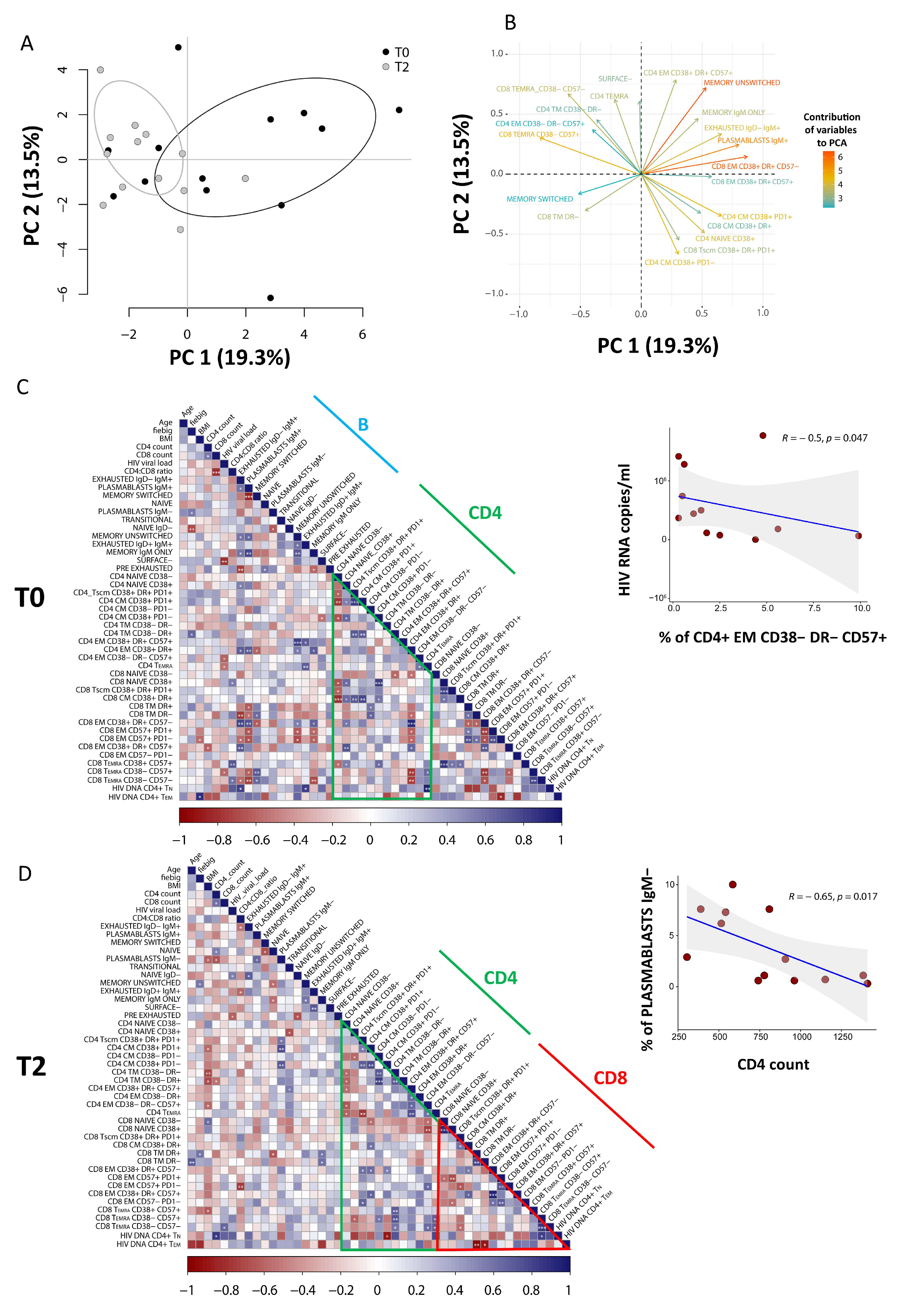
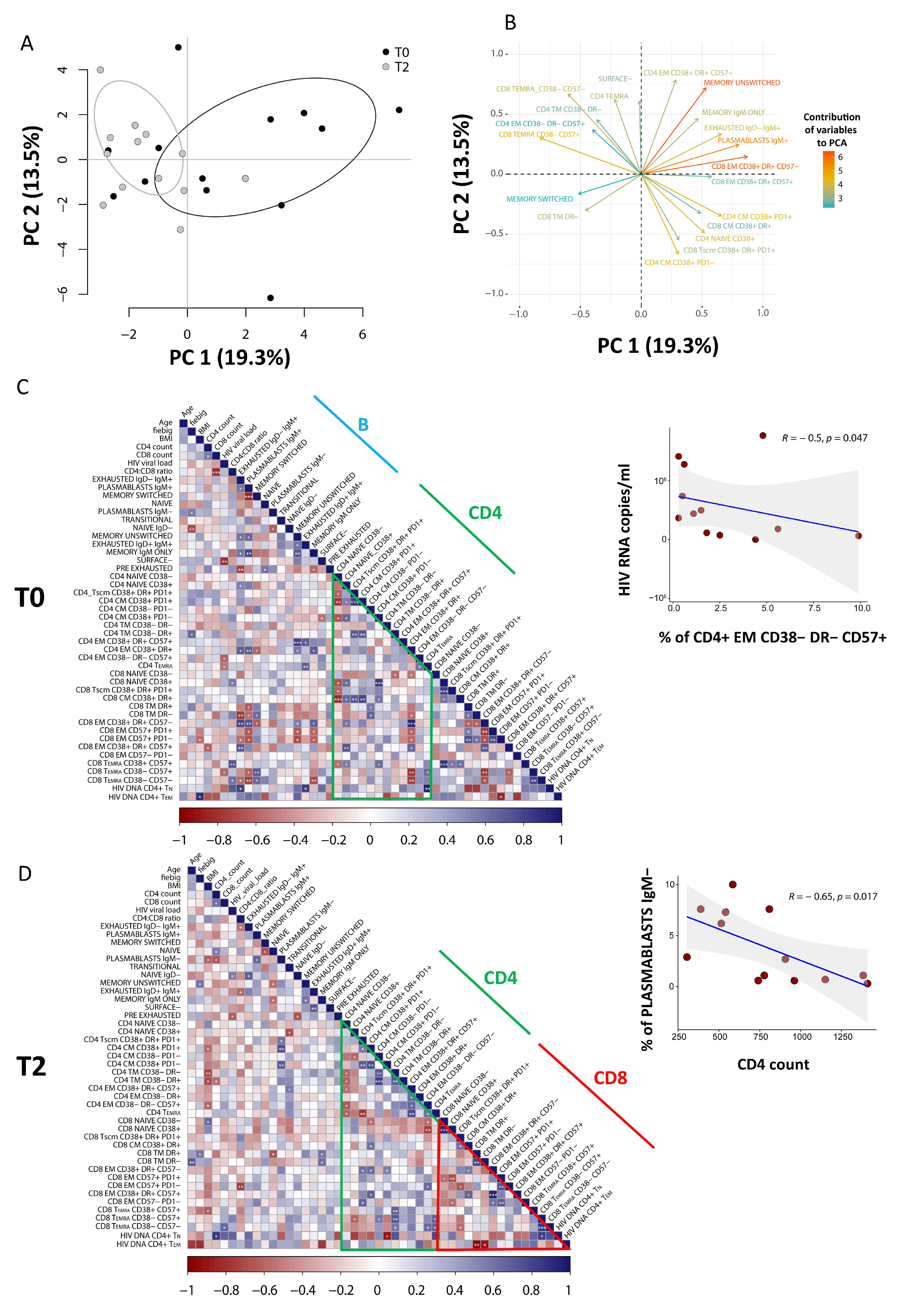
3.5. PCA of CD8 T Cells Identifies Critical Biomarkers Associated with Clinical Outcome
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cohen, M.S.; Shaw, G.M.; McMichael, A.J.; Haynes, B.F. Acute HIV-1 Infection. N. Engl. J. Med. 2011, 364, 1943–1954. [Google Scholar] [CrossRef] [Green Version]
- Strain, M.C.; Little, S.J.; Daar, E.S.; Havlir, D.V.; Gunthard, H.F.; Lam, R.Y.; Daly, O.A.; Nguyen, J.; Ignacio, C.C.; Spina, C.A.; et al. Effect of treatment, during primary infection, on establishment and clearance of cellular reservoirs of HIV-1. J. Infect. Dis. 2005, 191, 1410–1418. [Google Scholar] [CrossRef] [Green Version]
- Hellmuth, J.; Slike, B.M.; Sacdalan, C.; Best, J.; Kroon, E.; Phanuphak, N.; Fletcher, J.L.K.; Prueksakaew, P.; Jagodzinski, L.L.; Valcour, V.; et al. Very Early Initiation of Antiretroviral Therapy during Acute HIV Infection Is Associated with Normalized Levels of Immune Activation Markers in Cerebrospinal Fluid but Not in Plasma. J. Infect. Dis. 2019, 220, 1885–1891. [Google Scholar] [CrossRef] [Green Version]
- Ndhlovu, Z.M.; Kazer, S.W.; Nkosi, T.; Ogunshola, F.; Muema, D.M.; Anmole, G.; Swann, S.A.; Moodley, A.; Dong, K.; Reddy, T.; et al. Augmentation of HIV-specific T cell function by immediate treatment of hyperacute HIV-1 infection. Sci. Transl. Med. 2019, 11, eaau0528. [Google Scholar] [CrossRef] [Green Version]
- Schuetz, A.; Deleage, A.; Sereti, I.; Rerknimitr, R.; Phanuphak, N.; Phuang-Ngern, P.; Estes, J.D.; Sandler, N.G.; Sukhumvittaya, S.; Marovich, M.; et al. RV254/SEARCH 010 and RV304/SEARCH 013 Study Groups. Initiation of ART during early acute HIV infection preserves mucosal Th17 function and reverses HIV-related immune activation. PLoS Pathog. 2014, 10, e1004543. [Google Scholar] [CrossRef] [Green Version]
- Nasi, M.; Pinti, M.; Mussini, C.; Cossarizza, A. Persistent inflammation in HIV infection: Established concepts, new perspectives. Immunol. Lett. 2014, 161, 184–188. [Google Scholar] [CrossRef]
- Nasi, M.; Pinti, M.; De Biasi, S.; Gibellini, L.; Ferraro, D.; Mussini, C.; Cossarizza, A. Aging with HIV infection: A journey to the center of inflammAIDS, immunosenescence and neuroHIV. Immunol. Lett. 2014, 162, 329–333. [Google Scholar] [CrossRef] [PubMed]
- Nasi, M.; De Biasi, S.; Gibellini, L.; Bianchini, E.; Pecorini, S.; Bacca, V.; Guaraldi, G.; Mussini, C.; Pinti, M.; Cossarizza, A. Ageing and inflammation in patients with HIV infection. Clin. Exp. Immunol. 2017, 187, 44–52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fulop, T.; Herbein, G.; Cossarizza, A.; Witkowski, J.M.; Frost, E.; Dupuis, G.; Pawelec, G.; Larbi, A. Cellular Senescence, Immunosenescence and HIV. Interdiscip. Top. Gerontol. Geriatr. 2017, 42, 28–46. [Google Scholar]
- Estes, J.D.; LeGrand, R.; Petrovas, C. Visualizing the Immune System: Providing Key Insights into HIV/SIV Infections. Front. Immunol. 2018, 9, 423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Tellez, T.; Huot, N.; Ploquin, M.J.; Rascle, P.; Jacquelin, B.; Müller-Trutwin, T. Non-human primates in HIV research: Achievements, limits and alternatives. Infect. Genet. Evol. 2016, 46, 324–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McMichael, A.J.; Borrow, P.; Tomaras, G.D.; Goonetilleke, N.; Haynes, B.F. The immune response during acute HIV-1 infection: Clues for vaccine development. Nat. Rev. Immunol. 2010, 10, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Nemes, E.; Bertoncelli, L.; Lugli, E.; Pinti, M.; Nasi, M.; Manzini, L.; Manzini, S.; Prati, F.; Borghi, V.; Cossarizza, A.; et al. Cytotoxic granule release dominates gag-specific CD4+ T-cell response in different phases of HIV infection. AIDS 2010, 24, 947–957. [Google Scholar] [CrossRef]
- Nemes, E.; Lugli, E.; Nasi, M.; Ferraresi, R.; Pinti, M.; Bugarini, R.; Borghi, V.; Prati, F.; Esposito, R.; Cossarizza, A.; et al. Immunophenotype of HIV+ patients during CD4 cell-monitored treatment interruption: Role of the IL-7/IL-7 receptor system. AIDS 2006, 20, 2021–2032. [Google Scholar] [CrossRef] [PubMed]
- Nemes, E.; Lugli, E.; Bertoncelli, L.; Nasi, M.; Pinti, M.; Manzini, S.; Prati, F.; Manzini, L.; Del Giovane, C.; D’Amico, R.; et al. CD4+ T-cell differentiation, regulatory T cells and gag-specific T lymphocytes are unaffected by CD4-guided treatment interruption and therapy resumption. AIDS 2011, 25, 1443–1453. [Google Scholar] [CrossRef]
- Zaunders, J.J.; Munier, M.L.; Kaufmann, D.E.; Ip, S.; Grey, P.; Smith, D.; Ramacciotti, T.; Quan, D.; Finlayson, R.; Kaldor, J.; et al. Early proliferation of CCR5+ CD38+++ antigen-specific CD4+ Th1 effector cells during primary HIV-1 infection. Blood 2005, 106, 1660–1667. [Google Scholar] [CrossRef] [Green Version]
- Johnson, S.; Eller, M.; Teigler, J.E.; Maloveste, S.M.; Schultz, B.T.; Soghoian, D.Z.; Lu, R.; Oster, A.F.; Chenine, A.L.; Alter, G.; et al. Cooperativity of HIV-specific cytolytic CD4 T cells and CD8 T cells in control of HIV viremia. J. Virol. 2015, 89, 7494. [Google Scholar] [CrossRef] [Green Version]
- Wherry, E.J. T cell exhaustion. Nat. Immunol. 2011, 12, 492–499. [Google Scholar] [CrossRef]
- Fromentin, R.; DaFonseca, S.; Costiniuk, C.T.; El-Far, M.; Procopio, F.A.; Hecht, F.M.; Hoh, R.; Deeks, S.G.; Hazuda, D.J.; Lewin, S.R.; et al. PD-1 blockade potentiates HIV latency reversal ex vivo in CD4+ T cells from ART-suppressed individuals. Nat. Commun. 2019, 10, 814. [Google Scholar] [CrossRef] [Green Version]
- Moir, S.; Ho, J.; Malaspina, A.; Wang, W.; DiPoto, A.C.; O’Shea, M.A.; Roby, G.; Kottilil, S.; Arthos, J.; Proschan, M.A.; et al. Evidence for HIV-associated B cell exhaustion in a dysfunctional memory B cell compartment in HIV-infected viremic individuals. J. Exp. Med. 2008, 205, 1797–1805. [Google Scholar]
- Moir, S.; Malaspina, A.; Ho, J.; Wang, W.; Dipoto, A.C.; O’Shea, M.A.; Roby, G.; Mican, J.M.; Kottilil, S.; Chun, T.W.; et al. Normalization of B cell counts and subpopulations after antiretroviral therapy in chronic HIV disease. J. Infect. Dis. 2008, 197, 572–579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agrati, C.; De Biasi, S.; Fidanza, L.; Gibellini, L.; Nasi, M.; Pinti, M.; Cossarizza, A. The importance of advanced cytometry in defining new immune cell types and functions relevant for the immunopathogenesis of HIV infection. AIDS 2020, 34, 2169–2185. [Google Scholar] [CrossRef] [PubMed]
- Fiebig, E.W.; Wright, D.J.; Rawal, B.D.; Garrett, P.E.; Schumacher, R.T.; Peddada, L.; Heldebrant, C.; Smith, R.; Conrad, A.; Kleinman, S.H. Dynamics of HIV viremia and antibody seroconversion in plasma donors: Implications for diagnosis and staging of primary HIV infection. AIDS 2003, 17, 1871–1879. [Google Scholar] [CrossRef]
- Cossarizza, A.; Chang, H.D.; Radbruch, A.; Abrignani, S.; Addo, R.; Akdis, M.; Andra, I.; Andreata, F.; Annunziato, F.; Arranz, E.; et al. Guidelines for the use of flow cytometry and cell sorting in immunological studies (third edition). Eur. J. Immunol. 2021, 51, 2708–3145. [Google Scholar] [CrossRef]
- Lamoreaux, L.; Roederer, M.; Koup, R. Intracellular cytokine optimization and standard operating procedure. Nat. Protoc. 2006, 1, 1507–1516. [Google Scholar] [CrossRef]
- Gibellini, L.; Pecorini, S.; De Biasi, S.; Bianchini, E.; Digaetano, M.; Pinti, M.; Carnevale, G.; Borghi, V.; Guaraldi, G.; Mussini, C.; et al. HIV-DNA content in different CD4+ T-cell subsets correlates with CD4+ cell: CD8+ cell ratio or length of efficient treatment. AIDS 2017, 31, 1387–1392. [Google Scholar] [CrossRef] [PubMed]
- Crowell, H.Z.V.; Chevrier, S.; Robinson, M.; Crowell, H.; Zanotelli, V.; Chevrier, S.; Robinson, M. CATALYST: Cytometry dATa anALYSis Tools. R package version 1.14.0. 2020. Available online: https://github.com/HelenaLC/CATALYST (accessed on 22 June 2022).
- Weber, L.M.; Nowicka, M.; Soneson, C.; Robinson, M.D. diffcyt: Differential discovery in high-dimensional cytometry via high-resolution clustering. Commun. Biol. 2019, 2, 183. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Fu, Y.; Zhang, Z.; Tang, T.; Liu, J.; Ding, H.; Han, X.; Hu, J.; Chu, Z.; Shang, H.; et al. The investigation of CD4+ T-cell functions in primary HIV infection with antiretroviral therapy. Medicine 2017, 96, e7430. [Google Scholar] [CrossRef]
- Cheret, A.; Bacchus-Souffan, C.; Avettand-Fenoël, V.; Mélard, A.; Nembot, G.; Blanc, C.; Samri, A.; Saez-Cirion, A.; Hocqueloux, L.; Lascoux-Combe, C.; et al. Combined ART started during acute HIV infection protects central memory CD4+ T cells and can induce remission. J. Antimicrob. Chemother. 2015, 70, 2108. [Google Scholar] [CrossRef] [Green Version]
- Moir, S.; Buckner, C.M.; Ho, J.; Wang, W.; Chen, J.; Waldner, A.J.; Posada, G.J.; Kardava, L.; O’Shea, M.A.; Kottilil, S.; et al. B cells in early and chronic HIV infection: Evidence for preservation of immune function associated with early initiation of antiretroviral therapy. Blood 2010, 116, 5571–5579. [Google Scholar]
- Martin, G.E.; Pace, M.; Shearer, F.M.; Zilber, E.; Hurst, J.; Meyerowitz, J.; Thornhill, J.P.; Lwanga, J.; Brown, H.; Robinson, N.; et al. Levels of Human Immunodeficiency Virus DNA Are Determined before ART Initiation and Linked to CD8 T-Cell Activation and Memory Expansion. J. Infect. Dis. 2020, 221, 1135–1145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juno, J.A.; Van Bockel, D.; Kent, S.J.; Kelleher, A.D.; Zaunders, J.J.; Munier, C.M.L. Cytotoxic CD4 T cells–friends or foe during viral infection? Front. Immunol. 2017, 8, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pusnik, J.; Eller, M.A.; Tassaneetrithep, B.; Schultz, B.T.; Eller, L.A.; Nitayaphan, S.; Kosgei, J.; Maganga, L.; Kibuuka, H.; Alter, G.; et al. Expansion of Stem Cell-Like CD4(+) Memory T Cells during Acute HIV-1 Infection Is Linked to Rapid Disease Progression. J. Virol. 2019, 93, e00377-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabler, C.O.; Lucera, M.B.; Haqqani, A.A.; McDonald, D.J.; Migueles, S.A.; Connors, M.; Tilton, J.C. CD4+ memory stem cells are infected by HIV-1 in a manner regulated in part by SAMHD1 expression. J. Virol. 2014, 88, 4976–4986. [Google Scholar] [CrossRef] [Green Version]
- Buzon, M.J.; Sun, H.; Li, C.; Shaw, A.; Seiss, K.; Ouyang, Z.; Martin-Gayo, E.; Leng, J.; Henrich, T.J.; Li, J.Z.; et al. HIV-1 persistence in CD4+ T cells with stem cell-like properties. Nat. Med. 2014, 20, 139–142. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.A.; Sinclair, E.; Jain, V.; Huang, Y.; Epling, L.; Van Natta, M.; Meinert, C.L.; Martin, J.N.; McCune, J.M.; Deeks, S.G.; et al. Low proportions of CD28− CD8+ T cells expressing CD57 can be reversed by early ART initiation and predict mortality in treated HIV infection. J. Infect. Dis. 2014, 210, 374. [Google Scholar] [CrossRef]
- Galletti, G.; De Simone, G.; Mazza, E.M.C.; Puccio, S.; Mezzanotte, C.; Bi, T.M.; Davidov, A.N.; Metsger, M.; Scamardella, E.; Alvisi, G.; et al. Two subsets of stem-like CD8(+) memory T cell progenitors with distinct fate commitments in humans. Nat. Immunol. 2020, 21, 1552–1562. [Google Scholar] [CrossRef]
- Ribeiro, S.P.; Milush, J.M.; Cunha-Neto, E.; Kallas, E.G.; Kalil, J.; Somsouk, M.; Hunt, P.W.; Deeks, S.G.; Nixon, D.F.; SenGupta, D. The CD8(+) memory stem T cell (T(SCM)) subset is associated with improved prognosis in chronic HIV-1 infection. J. Virol. 2014, 88, 13836–13844. [Google Scholar] [CrossRef] [Green Version]
- Bordoni, V.; Brando, B.; Piselli, P.; Forini, O.; Perna, F.E.; Atripaldi, U.; Carputo, S.; Graziano, F.; Trento, E.; D’Agosto, G.; et al. Naive/Effector CD4 T cell ratio as a useful predictive marker of immune reconstitution in late presenter HIV patients: A multicenter study. PLoS ONE 2019, 14, e0225415. [Google Scholar] [CrossRef]
- Malaspina, A.; Moir, S.; Kottilil, S.; Hallahan, C.W.; Ehler, L.A.; Liu, S.; Planta, M.A.; Chun, T.W.; Fauci, A.S. Deleterious effect of HIV-1 plasma viremia on B cell costimulatory function. J. Immunol. 2003, 170, 5965–5972. [Google Scholar] [CrossRef] [Green Version]
- Pensieroso, S.; Galli, L.; Nozza, S.; Ruffin, N.; Castagna, A.; Tambussi, G.; Heideman, B.; Miscigna, D.; Riva, A.; Malnati, M.; et al. B-cell subset alterations and correlated factors in HIV-1 infection. AIDS 2013, 27, 1209–1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanko, R.F.; Soares, A.P.; Muller, T.L.; Garrett, N.J.; Samsunder, N.; Abdool Karim, Q.; Riou, C.; Burgers, W.A. Effect of Antiretroviral Therapy on the Memory and Activation Profiles of B Cells in HIV-Infected African Women. J. Immunol. 2017, 198, 1220–1228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Wang, J.; Kumar, V.; Karnell, J.L.; Naiman, B.; Gross, P.S.; Rahman, S.; Zerrouki, K.; Hanna, R.; Morehouse, C.; et al. IL-21 drives expansion and plasma cell differentiation of autoreactive CD11c(hi)T-bet(+) B cells in SLE. Nat. Commun. 2018, 9, 1758. [Google Scholar] [CrossRef]
- Fink, K. Origin and Function of Circulating Plasmablasts during Acute Viral Infections. Front. Immunol. 2012, 3, 78. [Google Scholar] [CrossRef] [PubMed] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lo Tartaro, D.; Camiro-Zúñiga, A.; Nasi, M.; De Biasi, S.; Najera-Avila, M.A.; Jaramillo-Jante, M.D.R.; Gibellini, L.; Pinti, M.; Neroni, A.; Mussini, C.; et al. Effective Treatment of Patients Experiencing Primary, Acute HIV Infection Decreases Exhausted/Activated CD4+ T Cells and CD8+ T Memory Stem Cells. Cells 2022, 11, 2307. https://doi.org/10.3390/cells11152307
Lo Tartaro D, Camiro-Zúñiga A, Nasi M, De Biasi S, Najera-Avila MA, Jaramillo-Jante MDR, Gibellini L, Pinti M, Neroni A, Mussini C, et al. Effective Treatment of Patients Experiencing Primary, Acute HIV Infection Decreases Exhausted/Activated CD4+ T Cells and CD8+ T Memory Stem Cells. Cells. 2022; 11(15):2307. https://doi.org/10.3390/cells11152307
Chicago/Turabian StyleLo Tartaro, Domenico, Antonio Camiro-Zúñiga, Milena Nasi, Sara De Biasi, Marco A. Najera-Avila, Maria Del Rocio Jaramillo-Jante, Lara Gibellini, Marcello Pinti, Anita Neroni, Cristina Mussini, and et al. 2022. "Effective Treatment of Patients Experiencing Primary, Acute HIV Infection Decreases Exhausted/Activated CD4+ T Cells and CD8+ T Memory Stem Cells" Cells 11, no. 15: 2307. https://doi.org/10.3390/cells11152307
APA StyleLo Tartaro, D., Camiro-Zúñiga, A., Nasi, M., De Biasi, S., Najera-Avila, M. A., Jaramillo-Jante, M. D. R., Gibellini, L., Pinti, M., Neroni, A., Mussini, C., Soto-Ramírez, L. E., Calva, J. J., Belaunzarán-Zamudio, F., Crabtree-Ramirez, B., Hernández-Leon, C., Mosqueda-Gómez, J. L., Navarro-Álvarez, S., Perez-Patrigeon, S., & Cossarizza, A. (2022). Effective Treatment of Patients Experiencing Primary, Acute HIV Infection Decreases Exhausted/Activated CD4+ T Cells and CD8+ T Memory Stem Cells. Cells, 11(15), 2307. https://doi.org/10.3390/cells11152307