
Selective Internal Radiotherapy Changes the Immune Profiles of Extracellular Vesicles and Their Immune Origin in Patients with Inoperable Cholangiocarcinoma

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics
2.2. Study Settings and Population
2.3. Technique of 90Y-Radioembolization
2.4. Data Acquisition and Blood Sampling
2.5. EV Isolation
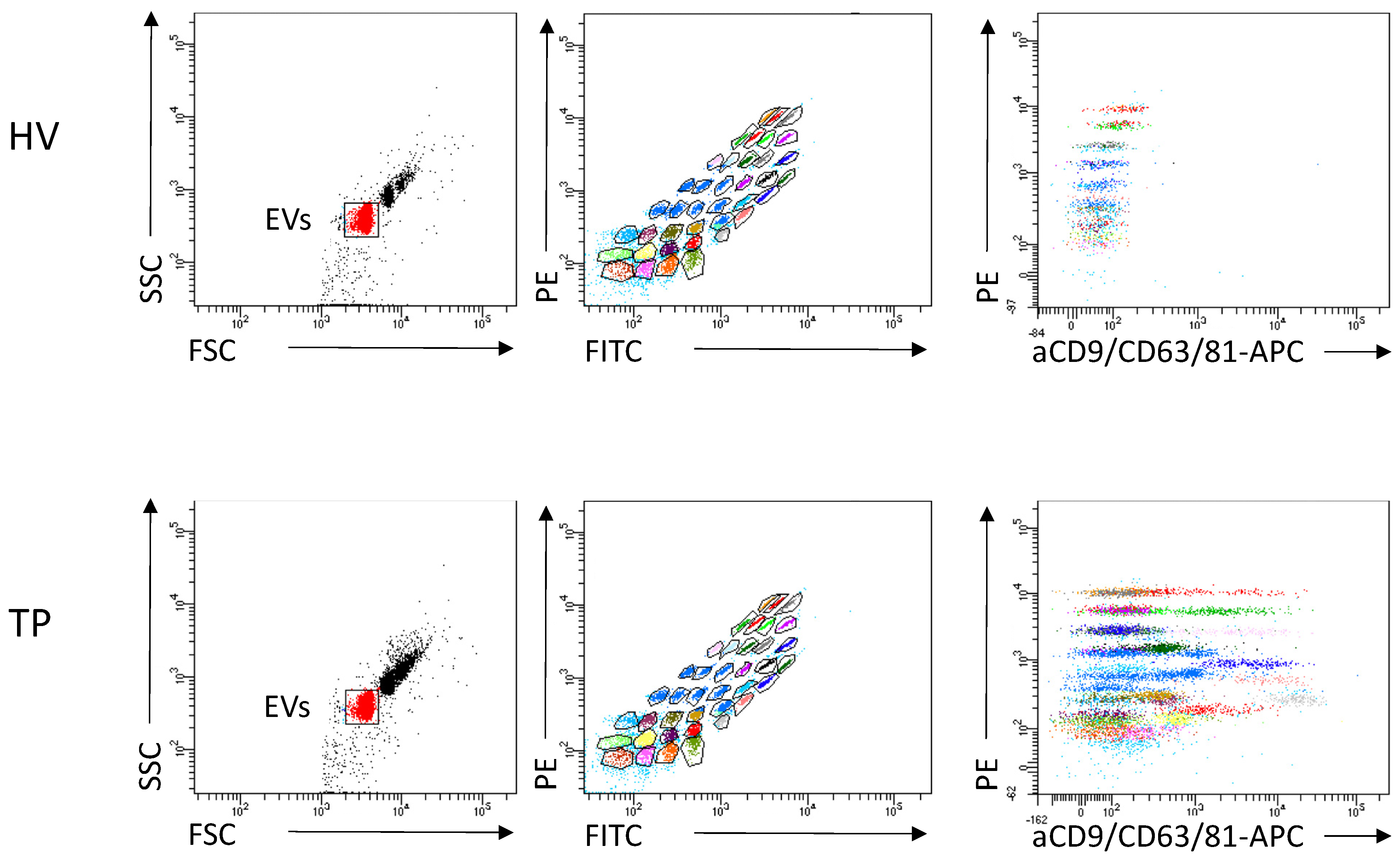
2.6. EV Measurement via FACS
2.7. Western Blotting
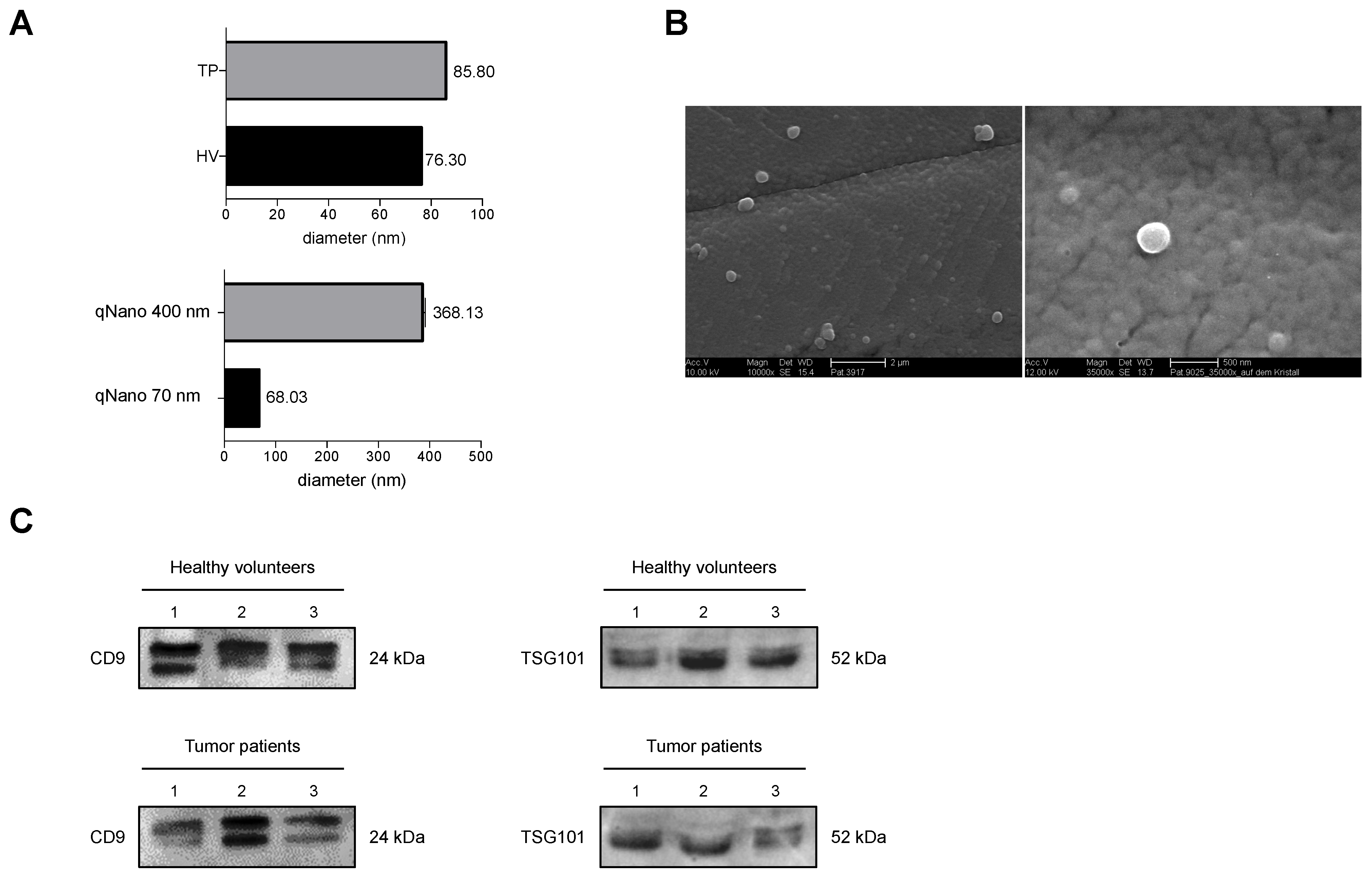
2.8. Dynamic Light Scattering
2.9. Scanning Electron Microscopy Analysis
2.10. Statistics
3. Results
3.1. Data Description
3.1.1. Patient Cohort
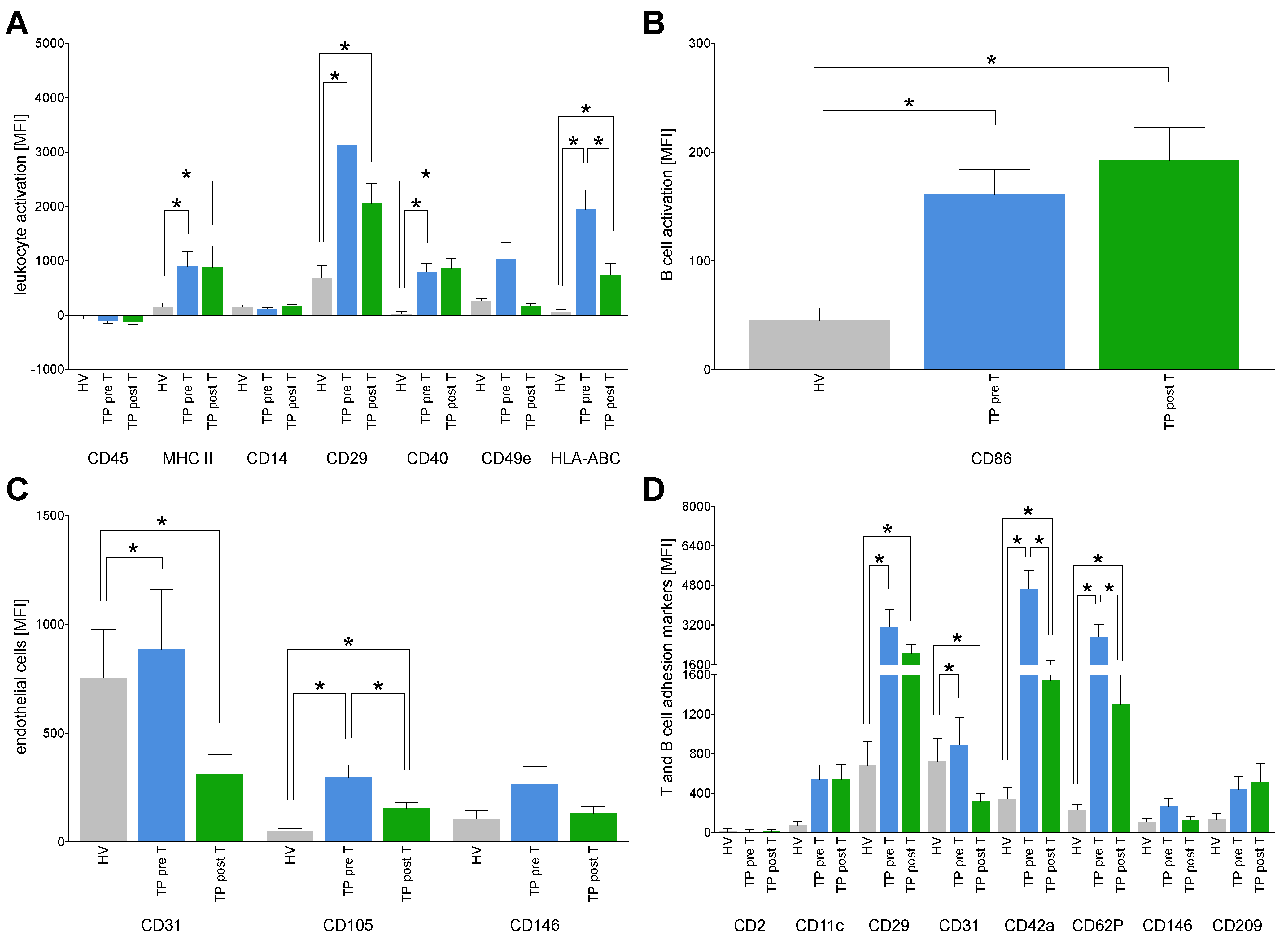
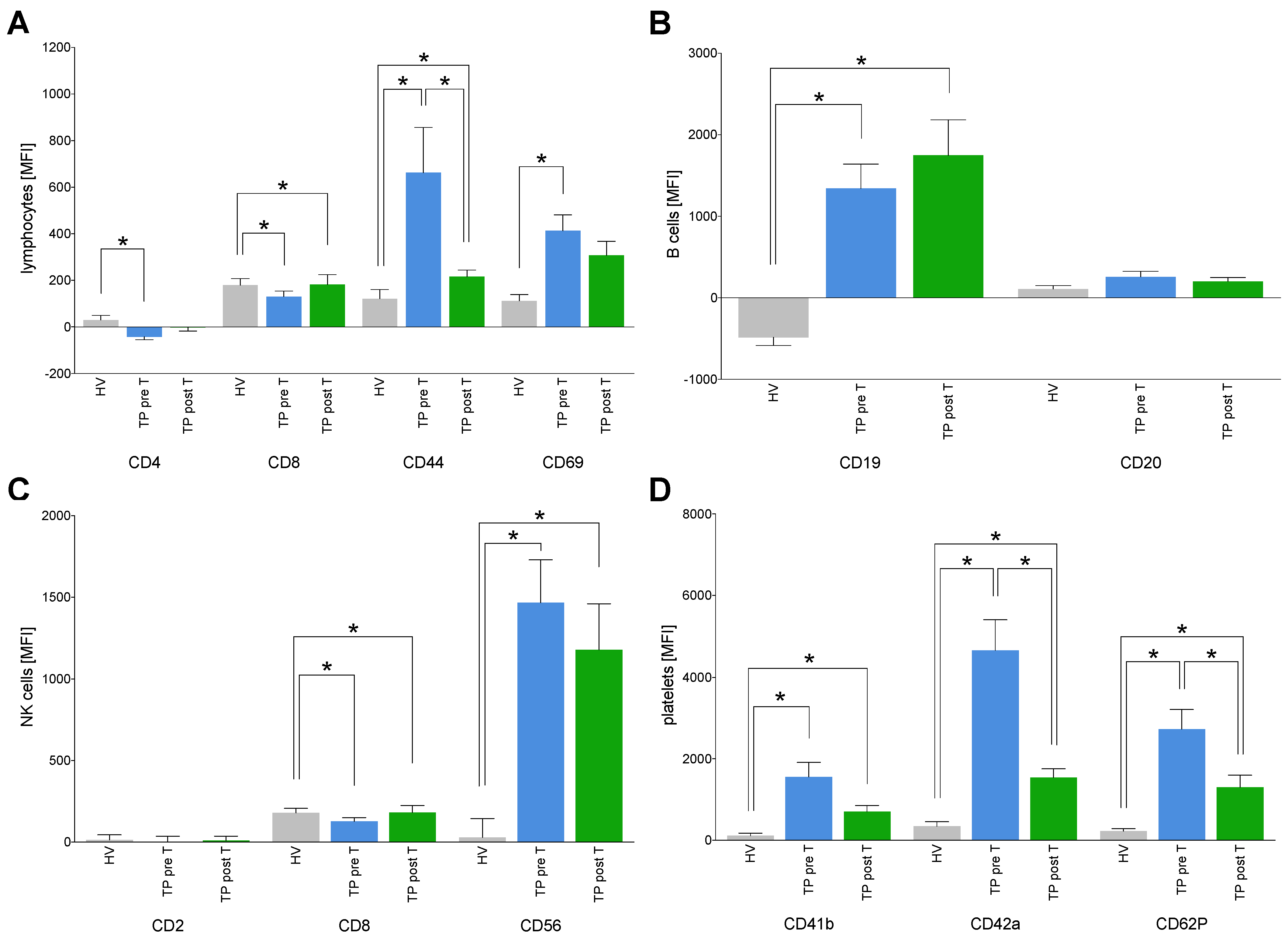
3.1.2. Immune Cell Status
3.1.3. Verification of Isolated EVs
3.1.4. Immune Origin of EVs
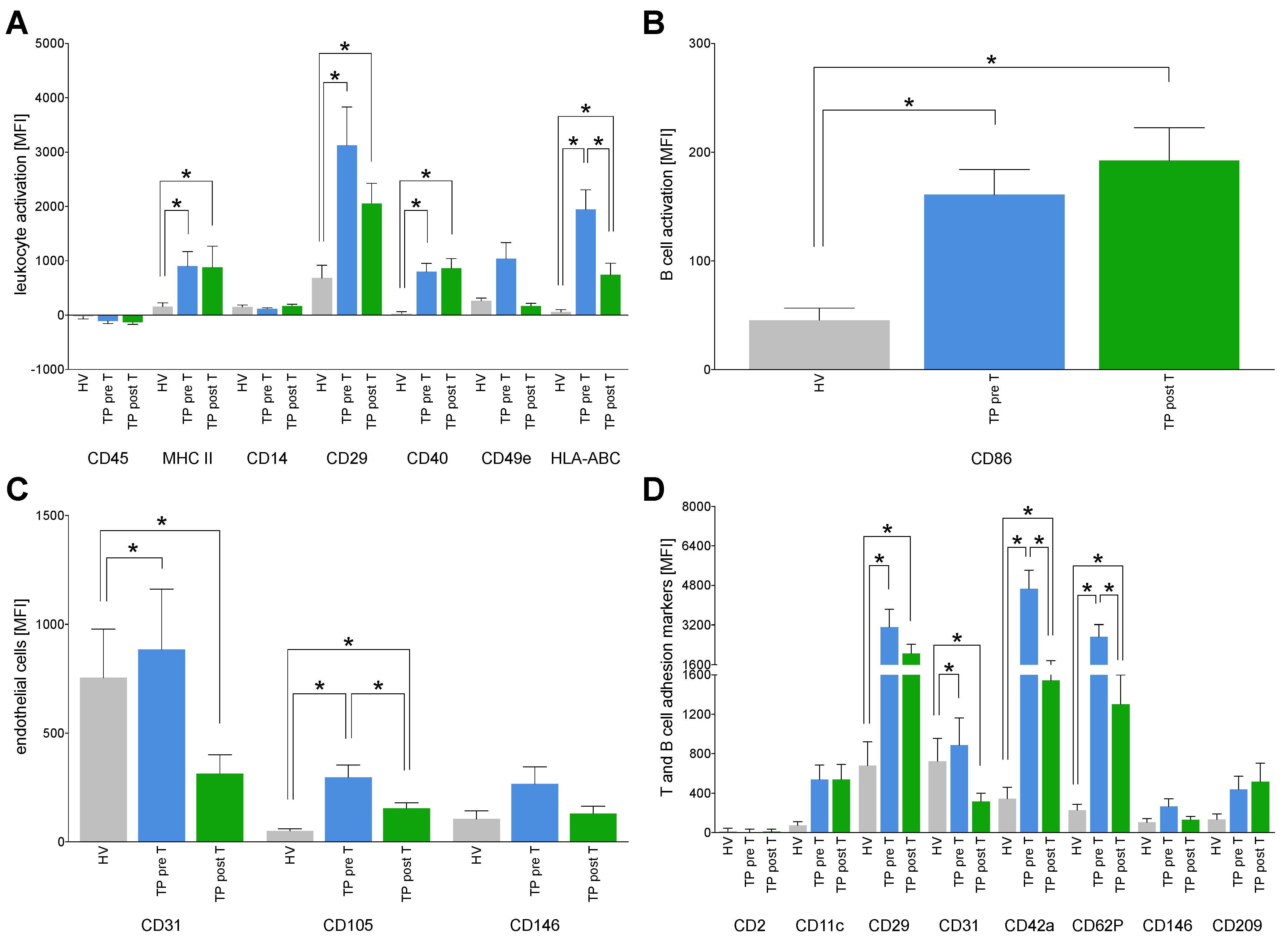
3.1.5. Immune Activation Markers on EVs
3.1.6. Expression of Tumor Markers on EVs
3.1.7. Correlation Analyses of the Expression Markers on EVs with the Administered Activity Dose as Well as Interval until Death
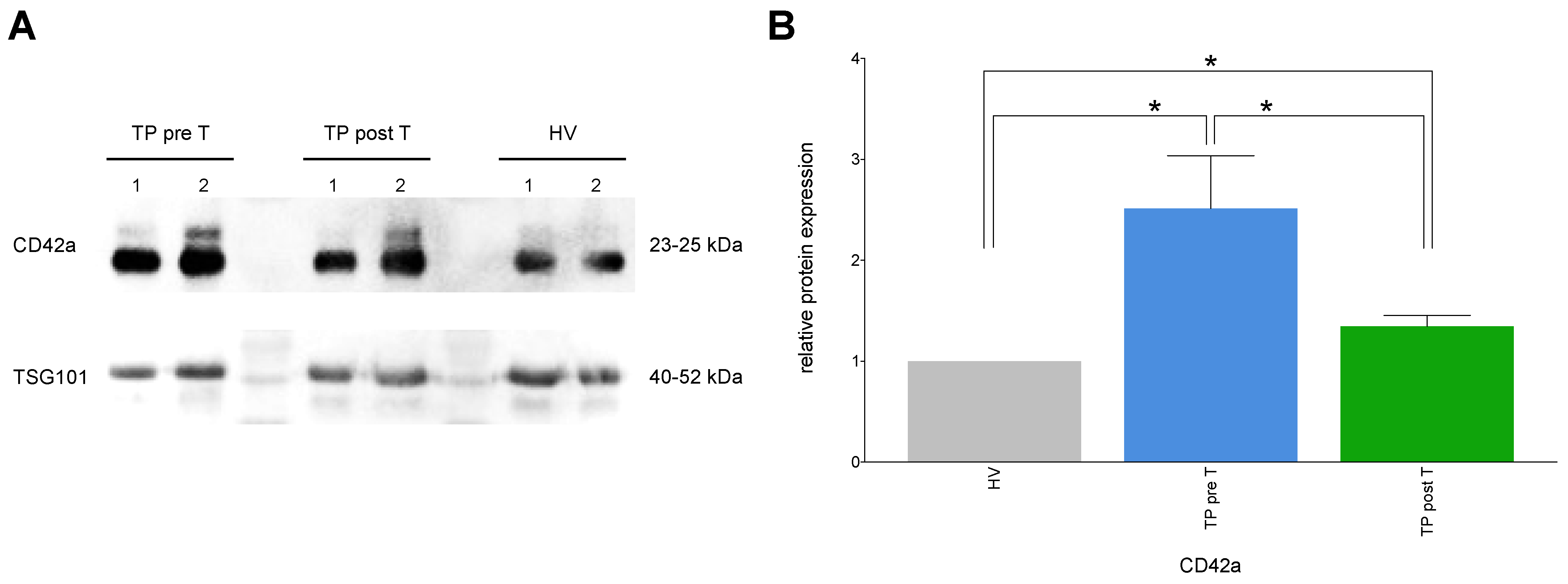
3.1.8. Exemplary Confirmation of Flow Cytometry Data by Western Blot
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Khan, S.A.; Tavolari, S.; Brandi, G. Cholangiocarcinoma: Epidemiology and risk factors. Liver Int. 2019, 39, 19–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banales, J.M.; Marin, J.J.G.; Lamarca, A.; Rodrigues, P.M.; Khan, S.A.; Roberts, L.R.; Cardinale, V.; Carpino, G.; Andersen, G.B.; Braconi, C.; et al. Cholangiocarcinoma 2020: The next horizon in mechanisms and management. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 557–588. [Google Scholar] [CrossRef]
- Macias, R.I.R.; Kornek, M.; Rodrigues, P.M.; Paiva, N.A.; Castro, R.E.; Urban, S.; Pereira, S.P.; Cadamuro, M.; Rupp, C.; Loosen, S.H.; et al. Diagnostic and prognostic biomarkers in cholangiocarcinoma. Liver Int. 2019, 39, 108–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banales, J.M.; Cardinale, V.; Carpino, G.; Marzioni, M.; Andersen, J.B.; Invernizzi, P.; Lind, G.E.; Folseraas, T.; Forbes, S.J.; Fouassier, L.; et al. Cholangiocarcinoma: Current knowledge and future perspectives consensus statement from the European Network for the Study of Cholangiocarcinoma (ENS-CCA). Nat. Rev. Gastroenterol. Hepatol. 2016, 13, 261–280. [Google Scholar] [CrossRef]
- Cholangiozelluläres Karzinom und Gallenblasenkarzinom. Z. Für Gastroenterol. 2004, 42, 397–402. [CrossRef] [PubMed]
- Lamarca, A.; Hubner, R.A.; David Ryder, W.; Valle, J.W. Second-line chemotherapy in advanced biliary cancer: A systematic review. Ann. Oncol. 2014, 25, 2328–2338. [Google Scholar] [CrossRef] [PubMed]
- Valle, J.; Wasan, H.; Palmer, D.H.; Cunningham, D.; Anthoney, A.; Maraveyas, A.; Madhusudan, S.; Iveson, T.; Hughes, S.; Pereira, S.P.; et al. Cisplatin plus Gemcitabine versus Gemcitabine for Biliary Tract Cancer. N. Engl. J. Med. 2010, 362, 1273–1281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boehm, L.M.; Jayakrishnan, T.T.; Miura, J.T.; Zacharias, A.J.; Johnston, F.M.; Turaga, K.K. Comparative effectiveness of hepatic artery based therapies for unresectable intrahepatic cholangiocarcinoma: Hepatic Artery Therapy for Unresectable, I.C.C. J. Surg. Oncol. 2015, 111, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Mouli, S.; Memon, K.; Baker, T.; Benson, A.B.; Mulcahy, M.F.; Gupta, R.; Ryu, R.K.; Salem, R.; Lewandowski, R.J. Yttrium-90 Radioembolization for Intrahepatic Cholangiocarcinoma: Safety, Response, and Survival Analysis. J. Vasc. Interv. Radiol. 2013, 24, 1227–1234. [Google Scholar] [CrossRef] [Green Version]
- Relja, B.; Pech, M. Transarterielle Radioembolisation: Immuneffekte und Immunonkologie. Radiologe 2020, 60, 693–703. [Google Scholar] [CrossRef]
- Powerski, M.; Drewes, R.; Omari, J.; Relja, B.; Surov, A.; Pech, M. Intra-hepatic Abscopal Effect Following Radioembolization of Hepatic Metastases. Cardiovasc. Intervent. Radiol. 2020, 43, 1641–1649. [Google Scholar] [CrossRef] [PubMed]
- Willms, E.; Cabañas, C.; Mäger, I.; Wood, M.J.A.; Vader, P. Extracellular Vesicle Heterogeneity: Subpopulations, Isolation Techniques, and Diverse Functions in Cancer Progression. Front. Immunol. 2018, 9, 738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Niel, G.; D’Angelo, G.; Raposo, G. Shedding light on the cell biology of extracellular vesicles. Nat. Rev. Mol. Cell. Biol. 2018, 19, 213–228. [Google Scholar] [CrossRef]
- Tkach, M.; Théry, C. Communication by Extracellular Vesicles: Where We Are and Where We Need to Go. Cell 2016, 164, 1226–1232. [Google Scholar] [CrossRef] [Green Version]
- Escola, J.-M.; Kleijmeer, M.J.; Stoorvogel, W.; Griffith, J.M.; Yoshie, O.; Geuze, H.J. Selective Enrichment of Tetraspan Proteins on the Internal Vesicles of Multivesicular Endosomes and on Exosomes Secreted by Human B-lymphocytes. J. Biol. Chem. 1998, 273, 20121–20127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.-K.; Kang, B.; Kim, O.Y.; Choi, D.; Lee, J.; Kim, S.R.; Go, G.; Yoon, Y.J.; Kim, J.H.; Jang, S.C.; et al. EVpedia: An integrated database of high-throughput data for systemic analyses of extracellular vesicles. J. Extracell. Vesicles 2013, 2, 20384. [Google Scholar] [CrossRef]
- Brahmer, A.; Neuberger, E.; Esch-Heisser, L.; Haller, N.; Jorgensen, M.M.; Baek, R.; Möbius, W.; Simon, P.; Krämer-Albers, E.-M. Platelets, endothelial cells and leu-kocytes contribute to the exercise-triggered release of extracellular vesicles into the circulation. J. Extracell. Vesicles 2019, 8, 1615820. [Google Scholar] [CrossRef] [PubMed]
- Naito, Y.; Yoshioka, Y.; Yamamoto, Y.; Ochiya, T. How cancer cells dictate their microenvironment: Present roles of ex-tracellular vesicles. Cell. Mol. Life Sci. 2017, 74, 697–713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Liu, Y.; Liu, H.; Tang, W.H. Exosomes: Biogenesis, biologic function and clinical potential. Cell Biosci. 2019, 9, 19. [Google Scholar] [CrossRef]
- Webber, J.; Steadman, R.; Mason, M.D.; Tabi, Z.; Clayton, A. Cancer Exosomes Trigger Fibroblast to Myofibroblast Differen-tiation. Cancer Res. 2010, 70, 9621–9630. [Google Scholar] [CrossRef] [Green Version]
- Webber, J.P.; Spary, L.K.; Sanders, A.J.; Chowdhury, R.; Jiang, W.G.; Steadman, R.; Wymant, J.; Jones, A.T.; Kynaston, H.; Mason, M.D.; et al. Differentiation of tumour-promoting stromal myofibroblasts by cancer exosomes. Oncogene 2015, 34, 290–302. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Zhang, H.; Chen, N.; Hao, J.; Jin, H.; Ma, X. Diagnostic value of various liquid biopsy methods for pancreatic cancer: A systematic review and meta-analysis. Medicine 2020, 99, e18581. [Google Scholar] [CrossRef]
- Guo, D.; Yuan, J.; Xie, A.; Lin, Z.; Li, X.; Chen, J. Diagnostic performance of circulating exosomes in human cancer: A meta-analysis. J. Clin. Lab. Anal. 2020, 3, e23341. Available online: https://onlinelibrary.wiley.com/doi/10.1002/jcla.23341 (accessed on 4 April 2022). [CrossRef] [PubMed] [Green Version]
- Xiang, H.; Li, F.; Luo, J.; Long, W.; Hong, L.; Hu, Y. A meta-analysis on the relationship of exosomes and the prognosis of lung cancer. Medicine 2021, 100, e25332. [Google Scholar] [CrossRef] [PubMed]
- Nanou, A.; Mol, L.; Coumans, F.A.W.; Koopman, M.; Punt, C.J.A.; Terstappen, L.W.M.M. Endothelium-Derived Extracellular Vesicles Associate with Poor Prognosis in Metastatic Colorectal Cancer. Cells 2020, 9, 2688. [Google Scholar] [CrossRef]
- Laschos, K.; Lampropoulou, D.I.; Aravantinos, G.; Piperis, M.; Filippou, D.; Theodoropoulos, G.; Gazouli, M. Exosomal noncoding RNAs in cholangiocarcinoma: Laboratory noise or hope? World J. Gastrointest. Surg. 2020, 12, 407–424. [Google Scholar] [CrossRef]
- Salem, R.; Thurston, K.G. Radioembolization with 90Yttrium Microspheres: A State-of-the-Art Brachytherapy Treatment for Primary and Secondary Liver Malignancies. J. Vasc. Interv. Radiol. 2006, 17, 1251–1278. [Google Scholar] [CrossRef] [Green Version]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Gardiner, C.; Shaw, M.; Hole, P.; Smith, J.; Tannetta, D.; Redman, C.W.; Sargent, I.L. Measurement of refractive index by nanoparticle tracking analysis reveals heterogeneity in extracellular vesicles. J. Extracell. Vesicles 2014, 3, 25361. [Google Scholar] [CrossRef]
- Harmati, M.; Gyukity-Sebestyen, E.; Dobra, G.; Janovak, L.; Dekany, I.; Saydam, O.; Hunyadi-Gulyas, E.; Nagy, I.; Farkas, A.; Pankotai, T.; et al. Small extracellular vesicles convey the stress-induced adaptive responses of melanoma cells. Sci. Rep. 2019, 9, 15329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malenica, M.; Vukomanović, M.; Kurtjak, M.; Masciotti, V.; Dal Zilio, S.; Greco, S.; Lazzarino, M.; Krušić, V.; Perčić, M.; Badovinac, I.J.; et al. Perspectives of Microscopy Methods for Morphology Characterisation of Extracellular Vesicles from Human Biofluids. Biomedicines 2021, 9, 603. [Google Scholar] [CrossRef]
- CLUSTER OF DIFFERENTIATION (CD) ANTIGENS. In Immunology Guidebook [Internet]; Elsevier: Amsterdam, The Netherlands, 2004; pp. 47–124. Available online: https://linkinghub.elsevier.com/retrieve/pii/B9780121983826500273 (accessed on 4 April 2022).
- Koliha, N.; Wiencek, Y.; Heider, U.; Jüngst, C.; Kladt, N.; Krauthäuser, S.; Johnston, I.C.D.; Bosio, A.; Schauss, A.; Wild, S. A novel multiplex bead-based platform highlights the diversity of extracellular vesicles. J. Extracell. Vesicles 2016, 5, 29975. [Google Scholar] [CrossRef] [PubMed]
- Panasiuk, A.; Zak, J.; Panasiuk, B.; Prokopowicz, D. Increase in expression of monocytic tissue factor (CD142) with monocytes and blood platelet activation in liver cirrhosis. Blood Coagul. Fibrinolysis 2007, 18, 739–744. [Google Scholar] [CrossRef] [PubMed]
- Uchida, N.; Buck, D.W.; He, D.; Reitsma, M.J.; Masek, M.; Phan, T.V.; Tsukamoto, A.S.; Gage, F.H.; Weissman, I.L. Direct isolation of human central nervous system stem cells. Proc. Natl. Acad. Sci. USA 2000, 97, 14720–14725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castelli, G.; Pelosi, E.; Testa, U. Liver Cancer: Molecular Characterization, Clonal Evolution and Cancer Stem Cells. Cancers 2017, 9, 127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weber, S. Intrahepatic Cholangiocarcinoma: Resectability, recurrence pattern, and outcomes. J. Am. Coll. Surg. 2001, 193, 384–391. [Google Scholar] [CrossRef]
- Endo, I.; Shimada, H.; Takimoto, A.; Fujii, Y.; Miura, Y.; Sugita, M.; Morioka, D.; Masunari, H.; Tanaka, K.; Sekido, H.; et al. Microscopic Liver Metastasis: Prognostic Factor for Patients with pT2 Gallbladder Carcinoma. World J. Surg. 2004, 28, 692–696. Available online: http://link.springer.com/10.1007/s00268-004-7289-4 (accessed on 4 April 2022). [CrossRef]
- Lapitz, A.; Arbelaiz, A.; Olaizola, P.; Aranburu, A.; Bujanda, L.; Perugorria, M.J.; Banales, J.M. Extracellular Vesicles in Hepatobiliary Malignancies. Front. Immunol. 2018, 9, 2270. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Peng, X.; Yang, S.; Li, X.; Huang, M.; Wei, S.; Zhang, S.; He, G.; Zheng, H.; Fan, Q.; et al. Extracellular vesicle PD-L1 in reshaping tumor immune microen-vironment: Biological function and potential therapy strategies. Cell Commun. Signal. 2022, 20, 14. [Google Scholar] [CrossRef]
- Seibold, T.; Schönfelder, J.; Weeber, F.; Lechel, A.; Armacki, M.; Waldenmaier, M.; Wille, S.; Palmer, A.; Halbgebauer, R.; Karasu, E.; et al. Small Extracellular Vesicles Propagate the Inflammatory Response After Trauma. Adv. Sci. 2021, 8, 2102381. [Google Scholar] [CrossRef]
- Shipkova, M.; Wieland, E. Surface markers of lymphocyte activation and markers of cell proliferation. Clin. Chim. Acta 2012, 413, 1338–1349. [Google Scholar] [CrossRef]
- Gerwing, M.; Kocman, V.; Stölting, M.; Helfen, A.; Masthoff, M.; Roth, J.; Barczyk-Kahlert, K.; Greune, L.; Schmidt, M.A.; Heindel, W.; et al. Tracking of Tumor Cell–Derived Extracellular Vesicles In Vivo Reveals a Specific Distribution Pattern with Consecutive Biological Effects on Target Sites of Metastasis. Mol. Imaging Biol. 2020, 22, 1501–1510. [Google Scholar] [CrossRef]
- Oba, R.; Isomura, M.; Igarashi, A.; Nagata, K. Circulating CD3 + HLA-DR + Extracellular Vesicles as a Marker for Th1/Tc1-Type Immune Responses. J. Immunol. Res. 2019, 2019, 6720819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oksvold, M.P.; Kullmann, A.; Forfang, L.; Kierulf, B.; Li, M.; Brech, A.; Vlassov, A.V.; Smeland, E.B.; Neurauter, A.; Pedersen, K.W. Expression of B-Cell Surface Antigens in Sub-populations of Exosomes Released From B-Cell Lymphoma Cells. Clin. Ther. 2014, 36, 847–862.e1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kin, N.W.; Sanders, V.M. CD86 Stimulation on a B Cell Activates the Phosphatidylinositol 3-Kinase/Akt and Phospho-lipase Cγ2/Protein Kinase Cαβ Signaling Pathways. J. Immunol. 2006, 176, 6727–6735. [Google Scholar] [CrossRef] [Green Version]
- Wolk, K.; Höflich, C.; Zuckermann-Becker, H.; Döcke, W.-D.; Volk, H.-D.; Sabat, R. Reduced monocyte CD86 expression in postinflammatory immunodeficiency. Crit. Care Med. 2007, 35, 458–467. [Google Scholar] [CrossRef] [PubMed]
- Tangye, S.G.; Phillips, J.H.; Lanier, L.L. The CD2-subset of the Ig superfamily of cell surface molecules: Receptor–ligand pairs expressed by NK cells and other immune cells. Semin. Immunol. 2000, 12, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Ji, C.G.; Zhang, J.Z.H. Glycosylation Modulates Human CD2-CD58 Adhesion via Conformational Adjustment. J. Phys. Chem. B 2015, 119, 6493–6501. [Google Scholar] [CrossRef]
- Gunesch, J.T.; Dixon, A.L.; Ebrahim, T.A.; Berrien-Elliott, M.M.; Tatineni, S.; Kumar, T.; Hegewisch-Solloa, E.; Fehniger, T.A.; Mace, E.M. CD56 regulates human NK cell cytotoxicity through Pyk2. Elife 2020, 9, e57346. [Google Scholar] [CrossRef]
- Panwong, S.; Wathikthinnakon, M.; Kaewkod, T.; Sawasdee, N.; Tragoolpua, Y.; Yenchitsomanus, P.; Panya, A. Cordycepin Sen-sitizes Cholangiocarcinoma Cells to Be Killed by Natural Killer-92 (NK-92) Cells. Molecules 2021, 26, 5973. [Google Scholar] [CrossRef]
- An, N.; Wang, H.; Jia, W.; Jing, W.; Liu, C.; Zhu, H.; Yu, J. The prognostic role of circulating CD8+ T cell proliferation in patients with untreated extensive stage small cell lung cancer. J. Transl. Med. 2019, 17, 402. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.Y.; Wang, R.-F. Regulatory T cells and cancer. Curr. Opin. Immunol. 2007, 19, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Kaech, S.M.; Cui, W. Transcriptional control of effector and memory CD8+ T cell differentiation. Nat. Rev. Immunol. 2012, 12, 749–761. [Google Scholar] [CrossRef] [Green Version]
- Kasper, H.-U.; Drebber, U.; Stippel, D.L.; Dienes, H.P.; Gillessen, A. Liver tumor infiltrating lymphocytes: Comparison of hepatocellular and cholangiolar carcinoma. World J. Gastroenterol. 2009, 15, 5053. [Google Scholar] [CrossRef]
- Vigano, L.; Soldani, C.; Franceschini, B.; Cimino, M.; Lleo, A.; Donadon, M.; Roncalli, M.; Aghemo, A.; Di Tommaso, L.; Torzilli, G. Tumor-Infiltrating Lymphocytes and Mac-rophages in Intrahepatic Cholangiocellular Carcinoma. Impact on Prognosis after Complete Surgery. J. Gastrointest. Surg. 2019, 23, 2216–2224. [Google Scholar] [CrossRef]
- Grassberger, C.; Hong, T.S.; Hato, T.; Yeap, B.Y.; Wo, J.Y.; Tracy, M.; Bortfeld, T.; Wolfgang, J.A.; Eyler, C.A.; Goyal, L.; et al. Differential Association Between Circulating Lym-phocyte Populations With Outcome After Radiation Therapy in Subtypes of Liver Cancer. Int. J. Radiat. Oncol. 2018, 101, 1222–1225. [Google Scholar] [CrossRef] [PubMed]
- Subimerb, C.; Pinlaor, S.; Lulitanond, V.; Khuntikeo, N.; Okada, S.; McGrath, M.S.; Wongkham, S. Circulating CD14+CD16+ monocyte levels predict tissue invasive character of cholangiocarcinoma. Clin. Exp. Immunol. 2010, 161, 471–479. [Google Scholar] [CrossRef]
- Chang, S.-H.; Kim, H.J.; Park, C.-G. Allogeneic ADSCs Induce the Production of Alloreactive Memory-CD8 T Cells through HLA-ABC Antigens. Cells 2020, 9, 1246. [Google Scholar] [CrossRef] [PubMed]
- Laman, J.D.; Claassen, E.; Noelle, R.J. Functions of CD40 and Its Ligand, gp39 (CD40L). Crit. Rev. Immunol. 2017, 37, 371–420. [Google Scholar] [CrossRef]
- Marino, J.; Babiker-Mohamed, M.H.; Crosby-Bertorini, P.; Paster, J.T.; LeGuern, C.; Germana, S.; Abdi, R.; Uehara, M.; Kim, J.I.; Markmann, J.F.; et al. Donor exosomes rather than passenger leukocytes initiate alloreactive T cell responses after transplantation. Sci. Immunol. 2016, 1, aaf8759. [Google Scholar] [CrossRef] [Green Version]
- Pawelec, G. Tumour escape from the immune response. Cancer Immunol. Immunother. 2004, 53, 843. Available online: http://link.springer.com/10.1007/s00262-004-0531-y (accessed on 4 April 2022). [CrossRef]
- Zhang, Y.; Luo, J.; Dong, X.; Yang, F.; Zhang, M.; Zhao, J.; Wang, Q.; Zhou, F.; Sun, J.; Yang, X. Establishment and Characterization of Two Novel Cholan-giocarcinoma Cell Lines. Ann. Surg. Oncol. 2019, 26, 4134–4147. [Google Scholar] [CrossRef]
- Geng, S.; Guo, Y.; Wang, Q.; Li, L.; Wang, J. Cancer stem-like cells enriched with CD29 and CD44 markers exhibit molecular characteristics with epithelial–mesenchymal transition in squamous cell carcinoma. Arch. Dermatol. Res. 2013, 305, 35–47. [Google Scholar] [CrossRef] [PubMed]
- Lan, J.; Huang, B.; Liu, R.; Ju, X.; Zhou, Y.; Jiang, J.; Liang, W.; Shen, Y.; Li, F.; Pang, L. Expression of cancer stem cell markers and their correlation with pathogenesis in vascular tumors. Int. J. Clin. Exp. Pathol. 2015, 8, 12621–12633. [Google Scholar]
- Middleton, J.; Americh, L.; Gayon, R.; Julien, D.; Mansat, M.; Mansat, P.; Anract, P.; Cantagrel, A.; Cattan, P.; Reimund, J.M.; et al. A comparative study of endothelial cell markers expressed in chronically inflamed human tissues: MECA-79, Duffy antigen receptor for chemokines, von Willebrand factor, CD31, CD34, CD105 and CD146. J. Pathol. 2005, 206, 260–268. [Google Scholar] [CrossRef] [PubMed]
- Brocco, D.; Lanuti, P.; Simeone, P.; Bologna, G.; Pieragostino, D.; Cufaro, M.C.; Graziano, V.; Peri, M.; Di Marino, P.; De Tursi, M.; et al. Circulating Cancer Stem Cell-Derived Extracellular Vesicles as a Novel Biomarker for Clinical Outcome Evaluation. J. Oncol. 2019, 2019, 5879616. [Google Scholar] [CrossRef] [PubMed]
- Ghoroghi, S.; Mary, B.; Larnicol, A.; Asokan, N.; Klein, A.; Osmani, N.; Busnelli, I.; Delalande, F.; Paul, N.; Halary, S.; et al. Ral GTPases promote breast cancer metastasis by controlling biogenesis and organ targeting of exosomes. Elife 2021, 10, e61539. [Google Scholar] [CrossRef] [PubMed]
- Douglas, S.R.; Yeung, K.T.; Yang, J.; Blair, S.L.; Cohen, O.; Eliceiri, B.P. Identification of CD105+ Extracellular Vesicles as a Candidate Biomarker for Metastatic Breast Cancer. J. Surg. Res. 2021, 268, 168–173. [Google Scholar] [CrossRef] [PubMed]
- Burrello, J.; Bolis, S.; Balbi, C.; Burrello, A.; Provasi, E.; Caporali, E.; Gauthier, L.G.; Peirone, A.; D’Ascenzo, F.; Monticone, S.; et al. An extracellular vesicle epitope profile is associated with acute myocardial infarction. J. Cell. Mol. Med. 2020, 24, 9945–9957. [Google Scholar] [CrossRef]
- Cadamuro, M.; Nardo, G.; Indraccolo, S.; Dall’Olmo, L.; Sambado, L.; Moserle, L.; Franceschet, I.; Colledan, M.; Massani, M.; Stecca, T.; et al. Platelet-derived growth factor-D and Rho GTPases regulate recruitment of cancer-associated fibroblasts in cholangiocarcinoma. Hepatology 2013, 58, 1042–1053. [Google Scholar] [CrossRef]
- Pan, S.; Hu, Y.; Hu, M.; Jian, H.; Chen, M.; Gan, L.; Zheng, P.; He, Y.; Wang, J. Platelet-derived PDGF promotes the invasion and metastasis of cholangiocarcinoma by upregulating MMP2/MMP9 expression and inducing EMT via the p38/MAPK signalling pathway. Am. J. Transl. Res. 2020, 12, 3577–3595. [Google Scholar]
- Keeratichamroen, S.; Leelawat, K.; Thongtawee, T.; Narong, S.; Aegem, U.; Tujinda, S.; Praditphol, N.; Tohtong, R. Expression of CD24 in cholan-giocarcinoma cells is associated with disease progression and reduced patient survival. Int. J. Oncol. 2011, 39, 873–881. [Google Scholar] [PubMed]
- Agrawal, S.; Kuvshinoff, B.W.; Khoury, T.; Yu, J.; Javle, M.M.; LeVea, C.; Groth, J.; Coignet, L.J.; Gibbs, J.F. CD24 Expression is an Independent Prognostic Marker in Cholangiocarcinoma. J. Gastrointest. Surg. 2007, 11, 445–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, M.-C.; Hsu, C.; Kao, H.-L.; Jeng, Y.-M. CD24 expression is a prognostic factor in intrahepatic cholangiocarcinoma. Cancer Lett. 2006, 235, 34–39. [Google Scholar] [CrossRef]
- Wattanawongdon, W.; Simawaranon Bartpho, T.; Tongtawee, T. Expression of CD44 and MDM2 in cholangiocarcinoma is correlated with poor clinicopathologic characteristics. Int. J. Clin. Exp. Pathol. 2019, 12, 3961–3967. [Google Scholar] [PubMed]
- Thanee, M.; Dokduang, H.; Kittirat, Y.; Phetcharaburanin, J.; Klanrit, P.; Titapun, A.; Namwat, N.; Khuntikeo, N.; Wangwiwatsin, A.; Saya, H.; et al. CD44 modulates metabolic pathways and altered ROS-mediated Akt signal promoting cholangiocarcinoma progression. Pizzo SV, Herausgeber. PLoS ONE 2021, 16, e0245871. [Google Scholar] [CrossRef] [PubMed]
- Haga, H.; Yan, I.K.; Takahashi, K.; Wood, J.; Zubair, A.; Patel, T. Tumour cell–derived extracellular vesicles interact with mesenchymal stem cells to modulate the microenvironment and enhance cholangiocarcinoma growth. J. Extracell. Vesicles 2015, 4, 24900. [Google Scholar] [CrossRef]
- Yoshikawa, S.; Zen, Y.; Fujii, T.; Sato, Y.; Ohta, T.; Aoyagi, Y.; Nakanuma, Y. Characterization of CD133 + parenchymal cells in the liver: Histology and culture. World J. Gastroenterol. 2009, 15, 4896. [Google Scholar] [CrossRef]
- Cai, X.; Li, J.; Yuan, X.; Xiao, J.; Dooley, S.; Wan, X.; Weng, H.; Lu, L. CD133 expression in cancer cells predicts poor prognosis of non-mucin producing intrahepatic cholangiocarcinoma. J. Transl. Med. 2018, 16, 50. [Google Scholar] [CrossRef]
- Shimada, M.; Sugimoto, K.; Iwahashi, S.; Utsunomiya, T.; Morine, Y.; Imura, S.; Ikemoto, T. CD133 expression is a potential prognostic indicator in intrahepatic cholangiocarcinoma. J. Gastroenterol. 2010, 45, 896–902. [Google Scholar] [CrossRef]
- Tseng, J.-C.; Chang, L.-C.; Jiang, B.-Y.; Liu, Y.-C.; Chen, H.-J.; Yu, C.-T.; Hua, C.C. Elevated circulating levels of tissue factor-positive microvesicles are associated with distant metastasis in lung cancer. J. Cancer Res. Clin. Oncol. 2014, 140, 61–67. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | n = 47 |
|---|---|
| Age [median (range)] | 71.00 (60.50–78.50) |
| Gender (female, n) | 20 |
| Total liver volume (cm3) [median (range)] | 1888 (1434–2629) |
| Tumor volume (cm3) [median (range)] | 379.50 (117.40–619.60) |
| Tumor fraction (%) [median (range)] | 21.19 (9.09–26.62) |
| Administered activity dose (MBq) [median (range)] | 1055.00 (871.30–1461.00) |
| Creatinine (umol/L) [median (range)] | 69.00 (54.50–86.00) |
| Urea (mmol/L) [median (range)] | 5.00 (3.90–6.50) |
| Uric acid (umol/L) [median (range)] | 294.50 (239.30–335.80) |
| Bilirubin (umol/L) [median (range)] | 6.85 (5.17–11.78) |
| Albumin (g/L) [median (range)] | 39.90 (37.90–43.30) |
| Alanine aminotransferase (umol/s·L) [median (range)] | 0.32 (0.25–0.59) |
| Aspartate aminotransferase (umol/s·L) [median (range)] | 0.59 (0.45–0.79) |
| Alkaline phosphatase (umol/s·L) [median (range)] | 2.60 (1.75–4.24) |
| Gamma-glutamyl transferase (umol/s·L) [median (range)] | 2.04 (1.89–5.79) |
| Quick value (%) [median (range)] | 91.50 (81.75–96.75) |
| INR [median (range)] | 1.045 (1.020–1.115) |
| PTT (sec.) [median (range)] | 27.30 (26.30–29.98) |
| Thrombin time (sec.) [median (range)] | 16.65 (15.70–17.43) |
| C-reactive protein (mg/L) [median (range)] | 8.75 (5.75–28.35) |
| Variables | Pre T | Post T | p Value |
|---|---|---|---|
| Leukocytes (Gpt/L) [median (range)] | 7.34 (6.13–9.85) | 9.62 (5.85–12.10) | 0.2305 |
| Erythrocytes (Tpt/L) [median (range)] | 4.06 (3.76–4.40) | 3.89 (3.61–4.31) | 0.0001 |
| red cell distribution width (%) [median (range)] | 14.45 (13.18–15.33) | 14.30 (13.45–15.85) | 0.5723 |
| Thrombocytes (Gpt/L) [median (range)] | 217.00 (150.30–278.00) | 201.00 (130.50–246.50) | <0.0001 |
| Neutrophil granulocytes (%) [median (range)] | 74.70 (66.55–80.00) | 76.50 (66.93–81.75) | 0.3750 |
| Neutrophil granulocytes (Gpt/L) [median (range)] | 5.42 (4.53–7.68) | 6.58 (4.15–9.88) | 0.7422 |
| Immature granulocytes (%) [median (range)] | 0.00 (0.00–0.01) | 0.01 (0.00–0.23) | 0.5000 |
| Immature granulocytes (Gpt/L) [median (range)] | 0.04 (0.02–0.06) | 0.11 (0.04–0.14) | 0.6875 |
| Eosinophil granulocytes (%) [median (range)] | 1.00 (0.53–2.00) | 1.00 (0.00–1.80) | 0.5000 |
| Eosinophil granulocytes (Gpt/L) [median (range)] | 0.07 (0.04–0.16) | 0.04 (0.01–0.07) | 0.5625 |
| Basophil granulocytes (%) [median (range)] | 0.65 (0.00–1.00) | 0.30 (0.00–1.00) | 0.7500 |
| Basophil granulocytes (Gpt/L) [median (range)] | 0.04 (0.02–0.06) | 0.03 (0.02–0.04) | 0.1250 |
| Lymphocytes (%) [median (range)] | 16.40 (10.25–20.30) | 12.00 (9.00–19.80) | 0.1563 |
| Lymphocytes (Gpt/L) [median (range)] | 1.19 (0.75–1.75) | 1.18 (0.67–1.64) | 0.1953 |
| Monocytes (%) [median (range)] | 9.00 (6.70–11.00) | 8.00 (7.60–8.50) | 0.8750 |
| Monocytes (Gpt/L) [median (range)] | 0.67 (0.43–0.92) | 0.68 (0.45–1.00) | 0.2500 |
| Parameter | Administered Activity Dose (MBq) | Interval until Death (Month) | ||||
|---|---|---|---|---|---|---|
| Immune System | EVs | Time Point | r | p Value | r | p Value |
| Lymphocytes | CD4 | pre T | 0.0446 | 0.8252 | 0.4290 | 0.1879 |
| post T | −0.0933 | 0.6956 | 0.1317 | 0.7168 | ||
| CD8 | pre T | −0.1500 | 0.4125 | 0.0100 | 0.9719 | |
| post T | −0.0498 | 0.8259 | 0.3174 | 0.3148 | ||
| CD44 | pre T | −0.0357 | 0.8488 | −0.0942 | 0.7384 | |
| post T | 0.2063 | 0.3571 | −0.1360 | 0.6735 | ||
| CD69 | pre T | 0.0462 | 0.8051 | −0.1122 | 0.6905 | |
| post T | 0.0271 | 0.9047 | 0.1284 | 0.6909 | ||
| B cells | CD19 | pre T | 0.3776 | 0.0331 | 0.0978 | 0.7287 |
| post T | −0.1689 | 0.4523 | 0.0653 | 0.8487 | ||
| CD20 | pre T | 0.3790 | 0.0355 | −0.2679 | 0.3158 | |
| post T | 0.0409 | 0.8602 | 0.1640 | 0.6105 | ||
| NK cells | CD2 | pre T | 0.0205 | 0.9174 | −0.2235 | 0.4424 |
| post T | −0.2465 | 0.2568 | 0.1705 | 0.6162 | ||
| CD8 | pre T | −0.1500 | 0.4125 | 0.0100 | 0.9719 | |
| post T | −0.0498 | 0.8259 | 0.3174 | 0.3148 | ||
| CD56 | pre T | −0.1662 | 0.3148 | 0.2597 | 0.3699 | |
| post T | −0.1617 | 0.4958 | 0.3016 | 0.3971 | ||
| Platelets | CD41b | pre T | −0.2308 | 0.2117 | 0.4734 | 0.0873 |
| post T | 0.0858 | 0.7192 | 0.0700 | 0.8381 | ||
| CD42a | pre T | −0.1941 | 0.9058 | −0.0060 | 0.9825 | |
| post T | −0.0518 | 0.8333 | −0.0183 | 0.9600 | ||
| CD62P | pre T | −0.0993 | 0.6223 | 0.1600 | 0.5848 | |
| post T | −0.2948 | 0.2070 | −0.1213 | 0.7225 | ||
| Leukocyte activation | CD45 | pre T | −0.2652 | 0.1358 | 0.0849 | 0.7546 |
| post T | −0.4881 | 0.0181 | 0.1177 | 0.7157 | ||
| MHC II | pre T | −0.4718 | 0.0064 | 0.5483 | 0.0227 | |
| post T | −0.3708 | 0.1075 | −0.0340 | 0.9258 | ||
| CD14 | pre T | 0.2318 | 0.2263 | 0.0673 | 0.8270 | |
| post T | −0.0627 | 0.7817 | 0.1498 | 0.6422 | ||
| CD29 | pre T | 0.0181 | 0.0477 | 0.1152 | 0.6950 | |
| post T | −0.4287 | 0.0525 | 0.1446 | 0.6715 | ||
| CD40 | pre T | −0.3594 | 0.0400 | 0.3061 | 0.2490 | |
| post T | −0.2248 | 0.3145 | 0.1336 | 0.6952 | ||
| CD49e | pre T | −0.3289 | 0.0616 | 0.5506 | 0.0271 | |
| post T | 0.0628 | 0.7760 | 0.1819 | 0.5716 | ||
| HLA-ABC | pre T | −0.1789 | 0.3355 | 0.0377 | 0.8983 | |
| post T | −0.1430 | 0.5591 | −0.0322 | 0.9252 | ||
| B cell activation | CD86 | pre T | −0.0369 | 0.8463 | 0.5185 | 0.0575 |
| post T | 0.2559 | 0.2504 | 0.0321 | 0.9211 | ||
| Endothelial cells | CD31 | pre T | −0.1417 | 0.1342 | 0.2048 | 0.5021 |
| post T | 0.0448 | 0.8470 | −0.2567 | 0.4205 | ||
| CD105 | pre T | 0.2721 | 0.1533 | 0.1248 | 0.6845 | |
| post T | −0.0624 | 0.7937 | −0.0886 | 0.7956 | ||
| CD146 | pre T | 0.1045 | 0.5759 | 0.0453 | 0.6845 | |
| post T | −0.2027 | 0.3781 | 0.3811 | 0.2216 | ||
| T and B cell adhesion markers | CD2 | pre T | 0.020 | 0.9174 | −0.2235 | 0.4424 |
| post T | −0.2465 | 0.2568 | 0.1705 | 0.6162 | ||
| CD11c | pre T | −0.2713 | 0.1267 | −0.0729 | 0.7884 | |
| post T | −0.4952 | 0.0163 | 0.3815 | 0.2210 | ||
| CD29 | pre T | 0.0181 | 0.0477 | 0.1152 | 0.6950 | |
| post T | −0.4287 | 0.0525 | 0.1446 | 0.6715 | ||
| CD31 | pre T | −0.1417 | 0.5401 | 0.2048 | 0.5021 | |
| post T | 0.0448 | 0.8470 | −0.2567 | 0.4205 | ||
| CD42a | pre T | −0.1941 | 0.2791 | −0.0060 | 0.9825 | |
| post T | −0.0518 | 0.8333 | −0.0183 | 0.9600 | ||
| CD146 | pre T | 0.1045 | 0.5759 | 0.0453 | 0.8727 | |
| post T | −0.2027 | 0.3781 | 0.3811 | 0.2216 | ||
| CD209 | pre T | 0.4003 | 0.0232 | −0.4524 | 0.0785 | |
| post T | −0.0554 | 0.8067 | −0.0392 | 0.9037 | ||
| Tissue Factor | CD142 | pre T | 0.3579 | 0.0443 | −0.2336 | 0.3838 |
| post T | −0.0209 | 0.9265 | 0.2318 | 0.4685 | ||
| “Don’t eat me signal” | CD24 | pre T | −0.0095 | 0.9596 | −0.2595 | 0.3502 |
| post T | −0.1191 | 0.5883 | 0.1287 | 0.7060 | ||
| CD44 | pre T | −0.0357 | 0.8488 | −0.0942 | 0.7384 | |
| post T | 0.2063 | 0.3571 | −0.1360 | 0.6735 | ||
| Stem cell/progenitor marker | CD133 | pre T | 0.1882 | 0.3106 | 0.2249 | 0.4204 |
| post T | 0.0527 | 0.8206 | 0.3540 | 0.2854 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haag, F.; Manikkam, A.; Kraft, D.; Bär, C.; Wilke, V.; Nowak, A.J.; Bertrand, J.; Omari, J.; Pech, M.; Gylstorff, S.; et al. Selective Internal Radiotherapy Changes the Immune Profiles of Extracellular Vesicles and Their Immune Origin in Patients with Inoperable Cholangiocarcinoma. Cells 2022, 11, 2309. https://doi.org/10.3390/cells11152309
Haag F, Manikkam A, Kraft D, Bär C, Wilke V, Nowak AJ, Bertrand J, Omari J, Pech M, Gylstorff S, et al. Selective Internal Radiotherapy Changes the Immune Profiles of Extracellular Vesicles and Their Immune Origin in Patients with Inoperable Cholangiocarcinoma. Cells. 2022; 11(15):2309. https://doi.org/10.3390/cells11152309
Chicago/Turabian StyleHaag, Florian, Anjana Manikkam, Daniel Kraft, Caroline Bär, Vanessa Wilke, Aleksander J. Nowak, Jessica Bertrand, Jazan Omari, Maciej Pech, Severin Gylstorff, and et al. 2022. "Selective Internal Radiotherapy Changes the Immune Profiles of Extracellular Vesicles and Their Immune Origin in Patients with Inoperable Cholangiocarcinoma" Cells 11, no. 15: 2309. https://doi.org/10.3390/cells11152309
APA StyleHaag, F., Manikkam, A., Kraft, D., Bär, C., Wilke, V., Nowak, A. J., Bertrand, J., Omari, J., Pech, M., Gylstorff, S., & Relja, B. (2022). Selective Internal Radiotherapy Changes the Immune Profiles of Extracellular Vesicles and Their Immune Origin in Patients with Inoperable Cholangiocarcinoma. Cells, 11(15), 2309. https://doi.org/10.3390/cells11152309