
Therapeutic Potential of Complementary and Alternative Medicines in Peripheral Nerve Regeneration: A Systematic Review

 ,
,  , ,
, , 
Abstract
1. Introduction
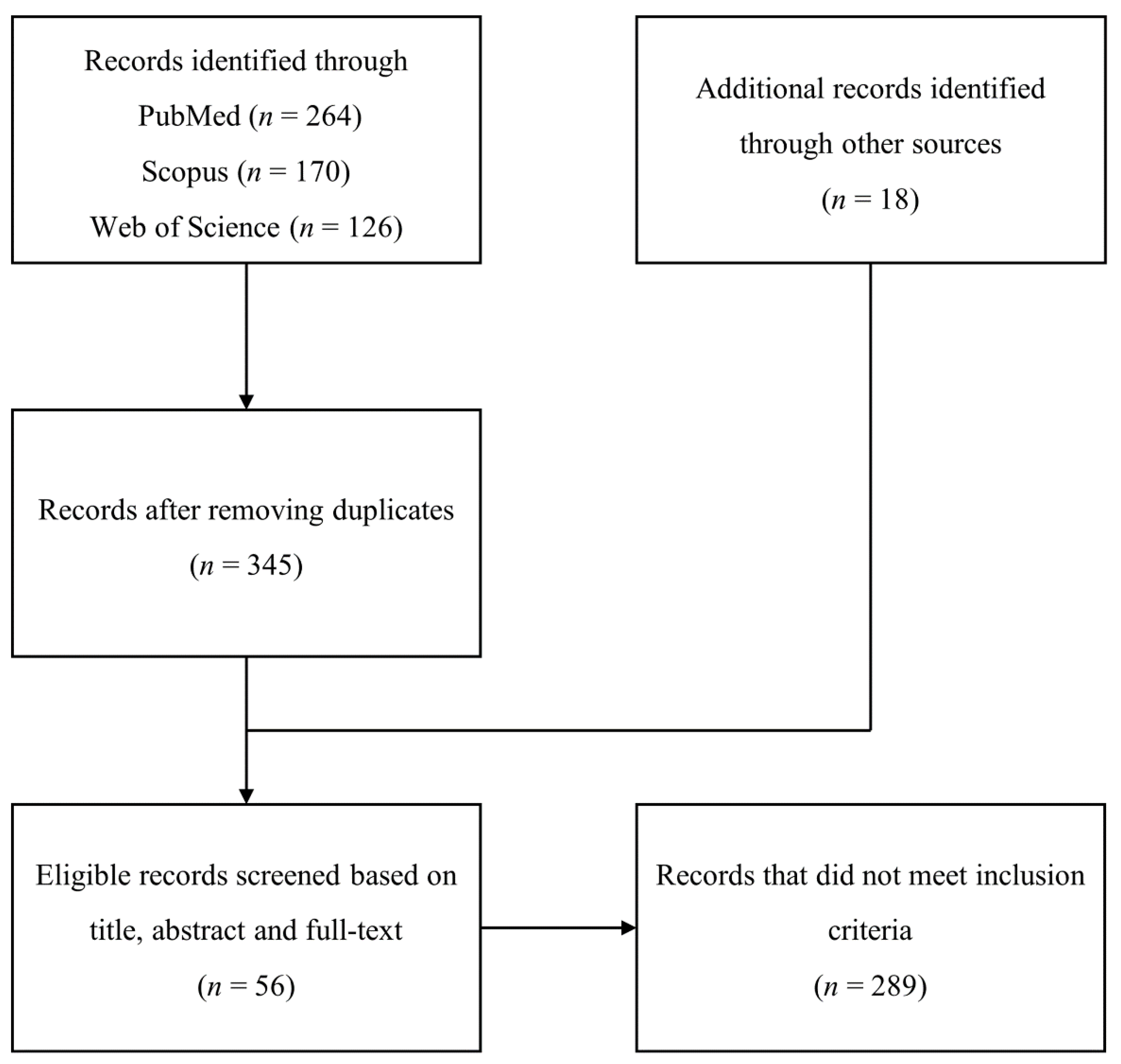
2. Materials and Methods
2.1. Search Strategy and Data Extraction
2.2. Eligibility Criteria
3. Results
4. Discussion
4.1. Current Therapeutic Approaches against Peripheral Nerve Injuries
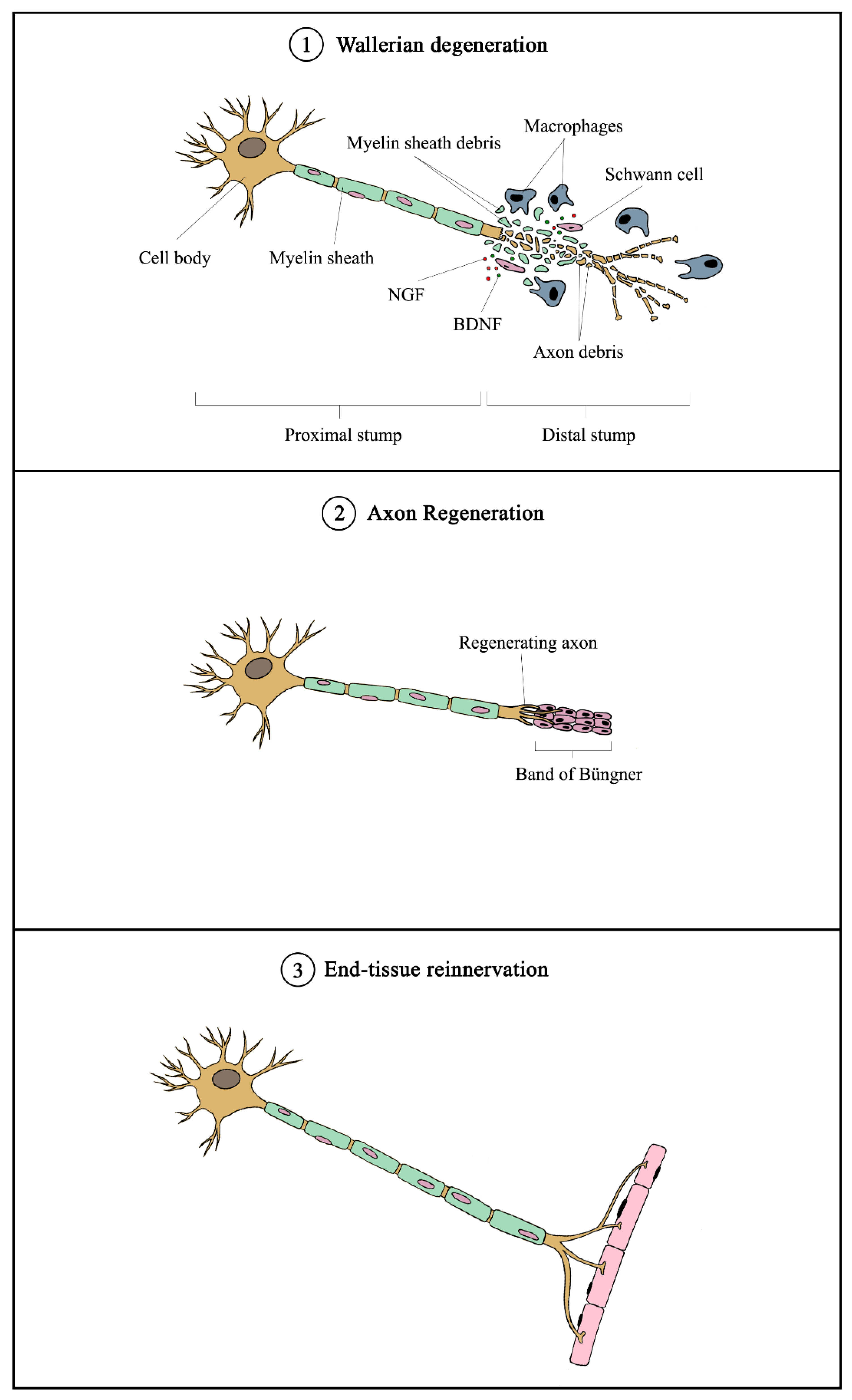
4.2. Mechanisms of Peripheral Nerve Injury and Regeneration
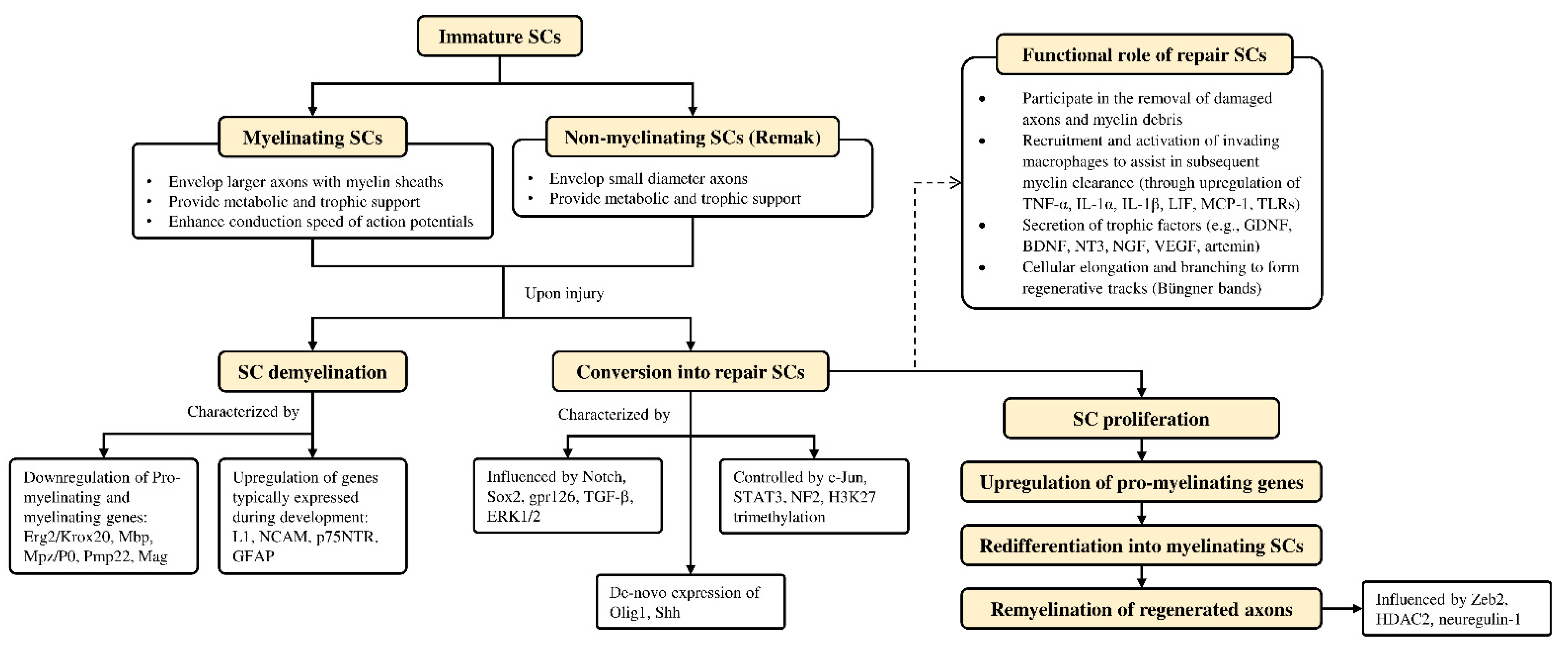
4.3. Role of Schwann Cells in Nerve Regeneration

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source | Molecule(s)/Ingredients | Experimental Model | Effective Concentration | Application Method | Biological Effect | Mechanism | Reference |
|---|---|---|---|---|---|---|---|
| PLANT | |||||||
| Achyranthes bidentata | Polypeptides | In vitro (SCs isolated from the sciatic nerves of 1-day old SD rats) | 0.1 µg/mL | Incubation | Promoted migration of SCs | Upregulation of NOX4/DUOX2-derived ROS production | [44] |
| Polypeptides | In vitro (DRG explants harvested from spinal and peripheral roots of postnatal day 1 SD rats) | 0.01, 0.1, 1 µg/mL (dose-dependent manner) | Incubation | Promoted neurite outgrowth from cultured DRG explants/neurons | Activation of ERK1/2 | [45] | |
| In vivo (Adult New Zealand rabbits) | 6.0 mg/kg | Intravenous injection | Enhanced nerve regeneration and functional restoration after crush injury to rabbit common peroneal nerve (increased CMAP, density, diameter and thickness of myelinated fibers, and number of motor neurons in anterior horn) | N/A | |||
| Polypeptides (Fraction K) | In vitro (DRG explants harvested from spinal and peripheral roots of postnatal day 1 SD rats) | 50, 250 ng/mL (dose-dependent manner) | Incubation | Promoted neurite outgrowth in DRG explant and neurons | Activation of ERK1/2 | [46] | |
| In vivo (ICR mice) | 10 mg/kg | Intravenous injection | Promoted peripheral nerve regeneration in mice after SNI (increased diameter and thickness of myelinated fibers, CSA of gastrocnemius muscle fibers, SFI, and CMAP) | N/A | |||
| Polypeptides | In vivo (SD rats) | 2 mg in 0.2 mL saline | Intraperitoneal injection | Promoted functional and histological recovery after rat sciatic nerve crush (increased SFI, CMAP, MNCV, myelin thickness, lamellae number, CSA of gastrocnemius muscle fibers) | Modulation of mRNA expression of GAP-43, neurotrophic factors (NGF, BDNF, CNTF), and neurotrophic factor receptors (TrkA, TrkB) | [47] | |
| Polypeptides | In vivo (ICR mice) | 1, 4, 16 mg/kg (dose-independent manner) | Tail vein injection | Promoted functional and histological recovery after rat sciatic nerve crush (increased SFI, CMAP, MNCV, number, and diameter of myelinated fibers, axon diameter, myelin thickness, lamellae number, CSA of gastrocnemius muscle fibers) | N/A | [48] | |
| Aqueous extract | In vivo (Adult New Zealand rabbits) | 10, 20 mg/kg (dose-dependent manner) | Intravenous injection | Promoted peripheral nerve regeneration in the crushed common peroneal nerve in rabbits (increased CMAP, CSA of tibialis posterior muscle, number of regenerated myelinated nerve fibers, and motoneurons in anterior horn of the spinal cord) | N/A | [49] | |
| Alpinate Oxyphyllae Fructus (Alpinia oxyphylla Miq) | Protocatechuic acid | In vitro (RSC96 SCs) | 1 mM | Incubation | Promoted proliferation and survival of RSC96 SCs | Upregulation of IGF-1 and activation of PI3K/Akt signaling | [50] |
| Aqueous extract | In vitro (RSC96 SCs) | Proliferation: 20, 60, 200 µg/mL (dose-independent manner Migration: 20–200 µg/mL (dose-dependent manner | Incubation | Promoted proliferation and migration of RSC96 SCs | Upregulation of PAs (uPA, tPA) and MMP2/9 mediated through the activation of MAPK pathway (ERK1/2, JNK, p38) | [51] | |
| In vivo (SD rats) | 30, 60, 100, 150, 200 µg/mL (dose-independent manner) | Injection into a silicone rubber tube bridging a 15mm sciatic nerve defect | Promoted peripheral nerve regeneration in rats with SNI | ||||
| Astragalus membranaceus | Astragaloside IV | In vivo (BALB/c mice) | 2.5, 5, 10 mg/kg (dose-dependent manner) | Intraperitoneal injection | Promoted sciatic nerve regeneration and functional recovery in mice (increased number and diameter of myelinated nerve fibers, MNCV, CMAP) | Upregulation of GAP-43 expression | [52] |
| Astragaloside IV | In vivo (SD rats) | 50 µM | Injection into a silicone rubber tube bridging a 15mm sciatic nerve defect | Promoted peripheral nerve regeneration in rats with SNI (increased number of myelinated axons and CMAP) | N/A | [53] | |
| Extract | In vivo (SD rats) | 3 g/kg in 0.01 M of PBS | Intragastric gavage | Promoted peripheral nerve regeneration in rats with SNI (increased MNCV and latency, fluorogold labeling in the DRG, mean axonal density, percentage of CGRP area ratio, and macrophage density) | Modulation of local growth factors (FGF, NGF, PDGF, TGF-β) and immunoregulatory factors (IL-1, IFN-γ) | [54] | |
| Aqueous extract | In vitro (RSC96 SCs) | Proliferation: 12.5, 125, 250, 500 µg/mL (optimal at 12.5 µg/mL) Migration: 1.25, 12.5, 125, 250, 500 µg/mL (optimal at 1.25 µg/mL) | Incubation | Promoted proliferation and migration of RSC96 SCs | Proliferation: Increased cyclin protein A, D1, and E via ERK and p38 signaling pathways Migration: Activation of FGF-2 signaling, leading to upregulation of uPA and downregulation of PAI-1 | [55] | |
| Centella asiatica | Hydro-ethanolic extract | In vivo (SD rats) | 400 µg/mL | Nerve conduit developed using decellularized artery seeded with C. asiatca-neurodifferentiated mesenchymal stem cells bridging a 15mm sciatic nerve defect | Promoted nerve regeneration and functional restoration in rats with SNI (increased CMAP, latency, MNCV, confirmation of angiogenesis, increased MBP expression, and number of myelinated axons) | N/A | [56] |
| Citrus medica var. sarcodactylis | Aqueous extract | In vitro (RSC96 SCs) | 0.85, 1.7, 2.55, 3.4, 4.25 µg/mL (dose-dependent manner) | Incubation | Promoted proliferation and migration of RSC96 SCs | Proliferation: Upregulation of cyclin A and B1 Migration: Activation of FGF-2 signaling, leading to the upregulation of uPA and MMP-9 | [57] |
| Codonopsis pilosula | Aqueous extract | In vitro (RSC96 SCs) | 20, 40, 60, 80, 100 µg/mL (dose-independent manner) | Incubation | Promoted proliferation and migration of RSC96 SCs | Proliferation: Enhanced IGF-I signaling pathway, cell cycle controlling protein expressions (cyclin A, D1, E) and MAPK pathway (ERK, p38) Migration: Stimulated FGF-2-uPA-MMP9 migration pathway | [58] |
| Crocus sativus | Crocin | In vivo (Wistar rats) | 20, 80 mg/kg | Intraperitoneal injection | Promoted functional recovery in rats with SNI (Increased SFI, reduced plasma MDA levels, alleviated histological changes due to a crushing injury) | N/A | [59] |
| Curcuma longa | Alcoholic extract | In vivo (Wistar rats) | 100 mg/kg (3, 6, or 9 times across 28 days) | Intraperitoneal injection | Protected against peripheral nerve degeneration in rats with SNI (Increased number of intact neurons in the right ventral horn of spinal cord region) | N/A | [60] |
| Curcumin | In vivo (SD rats) | 100 mg/kg (dissolved in olive oil) | Oral gavage | Promoted peripheral nerve regeneration in rats with SNI (increased mean cell volume, total volume and surface of DRG cells, total number, diameter, and area of myelinated nerve fibers) | N/A | [61] | |
| Curcumin | In vivo (SD rats) | 100 mg/kg (dissolved in olive oil) | Oral gavage | Promoted functional recovery (improved SFI) and protective effect on DRG (increased volume and number of A- and B- cells, number of satellite cells) in rats with SNI | N/A | [62] | |
| Curcumin | In vivo (SD rats) | 50, 100, 300 mg/kg | Intraperitoneal injection | Promoted peripheral nerve regeneration in rats with SNI (increased number of motoneurons, number and diameter of myelinated axons, SFI, MNCV, amplitude of CMAP, muscle fiber area and reduced latency of CMAP, mechanical withdrawal threshold, thermal withdrawal latency) | N/A | [63] | |
| Curcumin | In vitro (SCs isolated from S100β-DsRed transgenic mice) | 0.04-1 µM (0.1 µM having the highest proliferative effect) | Incubation | Promoted proliferation and migration of SCs | Proliferation: Modulated by ERK and p38 kinase pathways | [64] | |
| Curcuma longa (curcumin); from honeybees (propolis) | Curcumin, propolis | In vivo (Wistar rats) | Curcumin (100 mg/kg) Propolis (200 mg/kg) | Administration through a nasogastric tube | Promoted functional recovery in rats with SNI (Increased SFI and amplitude of CMAP, reduced latency time) | N/A | [65] |
| Dioscoreae rhizoma | Aqueous extract | In vivo (SD rats) | 10 mg/mL | Applied directly into the crush site | Promoted peripheral nerve regeneration in rats with SNI (increased number of DRG sensory neurons and motor neurons in the spinal cord) | Increasing protein levels of GAP-43 and Cdc2 | [66] |
| Epimedium | Icariin | In vivo (SD rats) | 20 mg in 5 mL | Injection into a poly(lactic-co-glycolic acid) biological conduit sleeve bridging a 5mm sciatic nerve defect | Promoted peripheral nerve regeneration in rats with SNI (increased sciatic nerve conduction velocity and number of myelinated fibers) | N/A | [67] |
| Epimedium extract, icariin | In vivo (SD rats) | 4.873 mg/mL | Intragastric administration | Promoted peripheral nerve regeneration in rats with SNI (Increased SFI, nerve regeneration based on nerve pinch test, MNCV, muscle wet weight) | N/A | [68] | |
| Gardenia jasminoides Ellis | Genipin | In vivo (SD rats) | 3% aqueous gelatin solution fixed with 3% genipin | Injection into a silicone rubber tube bridging a 10mm sciatic nerve defect | Promoted peripheral nerve regeneration in rats with SNI | N/A | [69] |
| Gastrodia elata Blume | Gastrodin | In vitro (RSC96 SCs) | 50, 100, 200 µM (dose-dependent manner) | Incubation | Promoted proliferation of RSC96 SCs in a dose- and time-dependent manner | Inhibition of ERK1/2 phosphorylation and activation of Akt phosphorylation | [70] |
| Ginkgo biloba | Ginkgo biloba extract (EGb 761) | In vivo (SD rats) | 50 mg/kg | Intraperitoneal injection paired with an 18mm acellular nerve allograft bridging a 15mm sciatic nerve defect | Promoted peripheral nerve regeneration in rats with SNI (increased density of regenerated axons, muscle mass, axon number and diameter, expression of CD34 and NF200) | Increasing expression of angiogenesis-related genes (Vegf, Sox18, Prom1, IL-6) | [71] |
| Ginkgo biloba extract (EGb 761) | In vitro (SCs isolated from spinal nerves of 1-day old SD rats) | 1, 10, 20, 50, 100 µg/mL (dose-dependent manner) | Incubation | Promoted cell attachment and survival of SCs | N/A | [72] | |
| In vivo (SD rats) | 10, 50 µg/mL | Injection into poly(DL-lactic acid-co-glycolic acid) conduit seeded with Schwann cells bridging a 12mm sciatic nerve defect | Promoted histological and functional recovery in rats with SNI (increased number and area of myelinated axons, increased CMAP) | ||||
| Ginseng | Ginsenoside Rg1 | In vitro (RSC96 SCs) | Ginseng: 100, 200, 300, 400, 500 µg/mL Ginsenoside: 5, 10, 15, 20, 25 µg/mL) (Dose-dependent manner for both) | Incubation | Promoted proliferation and migration of RSC96 SCs | Proliferation: Enhancing protein expression of IGF-I pathway regulators (IGF-IR, PI3K, p-Akt, p-Bad, Bcl-2), cell cycle controlling proteins (cyclin D1, E, A), and MAPK signaling pathway (ERK, JNK, p38) Migration: Stimulating the FGF-2-uPA-MMP9 migrating pathway | [73] |
| Ginsenoside Rg1 | In vivo(SD rats) | 1.5 mg/kg | Intraperitoneal injection | Promoted peripheral nerve regeneration in rats with SNI (increased number of motoneurons, number, and diameter of myelinated axons, SFI, MNCV, improved CMAP latency and amplitude, the increased average percentage of muscle fiber) | N/A | [74] | |
| Ginsenoside Re | In vitro (SCs isolated from sciatic nerves of 3-day old SD rats) | 0.5 mg/mL | Incubation | Promoted proliferation and migration of SCs | Phosphorylation of ERK1/2 and JNK 1/2 | [75] | |
| In vivo (SD rats) | 2.0 mg/kg | Intraperitoneal injection | Promoted peripheral nerve regeneration in rats with SNI (increased SFI, TSI, PCNA expression level, improved pathological changes due to crushing injury, GAP43, and S-100 expression) | ||||
| Green tea | (-)-Epigallocatechin-3-gallate (EGCG) | In vivo (Wistar rats) | 50 mg/kg | Intraperitoneal injection | Promoted functional recovery (improved outcomes of foot position, toe spreading, extensor postural thrust, hopping reflex, von Frey hair, Randall–Sellito, hotplate, and tail-flick tests), improved morphological recovery in skeletal muscle tissues muscles, and protection towards muscle fibers in rats with SNI | Protection of muscle fibers from cellular death through activation of an anti-apoptotic signaling pathway (modulation of Bax, Bcl-2, and p53 expression) | [76] |
| (-)-Epigallocatechin-3-gallate (EGCG) | In vivo (Wistar rats) | 50 mg/kg | Intraperitoneal injection | Promoted peripheral nerve regeneration in rats with SNI (improved nerve morphology and functional recovery assessed by foot position, extensor postural thrust test, and withdrawal reflex threshold) | Reversal of Bax, Bcl-2, and survivin mRNA expression induced by sciatic nerve injury | [77] | |
| Can be found in a wide variety of plants | Syringic acid | In vitro (RSC96 SCs) | 600 µM | Incubation | Promoted proliferation and migration of RSC96 SCs | Downregulation of miR-451-5p | [78] |
| Can be found in a wide variety of plants | Ursolic acid | In vivo (BALB/c mice) | 2.5, 5, 10 mg/kg (dose-dependent manner) | Intraperitoneal injection | Promoted peripheral nerve regeneration in rats with SNI (increased number and diameter of myelinated nerve fibers and soleus muscle mass) | Increasing S100 protein expression levels | [79] |
| Lycium barbarum | Polysaccharide | In vitro (1) PC12 cells (2) Rat SCs (3) DRG neurons isolated from the embryo of 14-day pregnant rat | 10, 30, 50 mg/mL (optimal at 30 mg/mL) | Incorporated into core-shell structured nanofibrous scaffolds by coaxial electrospinning | (1) Promoted proliferation and neuronal differentiation of PC12 cells (2) Promoted proliferation and myelination of SCs (3) Promoted neurite outgrowth of DRG neurons | N/A | [80] |
| Can be found in a wide variety of plants | Quercetin | In vitro (RSC96 SCs) | 0.1, 1, 10 µg/mL | Incubation | Promoted proliferation of RSC96 SCs | N/A | [81] |
| In vivo (SD rats) | 0.1, 1, 10 µg/mL | Injection into a silicone rubber tube bridging a 15mm sciatic nerve defect | Promoted peripheral nerve regeneration in rats with SNI (increased count and density of myelinated axons, and resulted in larger area and amplitude of CMAP) | ||||
| Morus sp. | Cortex Mori Radicis (aqueous extract) | In vivo (SD rats) | 100 mg/kg | Gastrointestinal administration | Reduced blood glucose levels, improved nerve functions (thermal latency and mechanical threshold), reversed the loss of Nissl bodies and induced neurite outgrowth in DRG neurons, and restored the response of growth cones to NGF in diabetic rats | Neurite outgrowth: Increased expression of TRPC1, reduced Ca2+ influx, and activation of PI3K/Akt signaling | [82] |
| Pueraria lobata | Puerarin | In vitro (RSC96 SCs) | 1, 10, 100 µM (dose-independent manner) | Incubation | Promoted growth of SCs | N/A | [83] |
| In vivo (SD rats) | 1, 10, 100 µM (dose-independent manner) | Injection into a silicone rubber tube bridging a 15mm sciatic nerve defect | Promoted peripheral nerve regeneration in rats with SNI (increased density of myelinated axons, CMAP, and MNCV) | ||||
| Serum metabolites (obtained from rats fed with Pueraria lobata extract) | In vitro (PC12 cells) | 0.01, 0.1, 1 unit | Incubation | Enhanced NGF-mediated neurite outgrowth and expression of synapsin I in PC12 cells | N/A | [84] | |
| In vivo (SD rats) | 0.01, 0.1, 1 unit | Injection into silicone rubber chamber bridging a 10mm sciatic nerve defect | Promoted peripheral nerve regeneration in rats with SNI (increased mean values of myelinated axon number, endoneurial area, and total nerve area) | ||||
| Radix Hedysari | Aqueous extract | In vivo (SD rats) | 1 g/mL | Oral gavage paired with biodegradable chitin conduit bridging a 2mm sciatic nerve defect | Promoted peripheral nerve regeneration in rats with SNI (increased MNCV, fiber and axon diameter, g-ratio) | N/A | [85] |
| Polysaccharides | In vivo (SD rats) | 0.25 g/mL | Oral gavage | Promoted peripheral nerve regeneration in rats with sciatic nerve defect (increased SFI, TFI, PFI values, MNCV, and number of regenerated myelinated nerve fibers) | N/A | [86] | |
| Rhodiola rosea L. | Salidroside | In vivo (SD rats) | 5, 10 mg/kg | Intraperitoneal injection | Promoted peripheral nerve regeneration in rats with SNI (increased number and diameter of myelinated axons, number of motoneurons, SFI, amplitude of CMAP, MNCV) | N/A | [87] |
| Scutellaria baicalensis Georgi | Baicalin | In vitro (RSC96 SCs) | 5, 10, 20 µM (dose-dependent manner) | Incubation | Promoted proliferation of RSC96 SCs | Modulation of neurotrophic factors (GDNF, BDNF, CNTF) and S100β | [88] |
| Trigonella foenum-graecum (fenugreek) | IND01 (Fenugreek seed extract) | In vivo (Wistar rats) | 50, 100, 200 mg/kg | Oral administration | Promoted peripheral nerve regeneration in rats with: (1) partial sciatic nerve ligation (ameliorated thermal hyperalgesia, improved motor function test scores) (2) SNI (ameliorated thermal hyperalgesia, improved motor function test scores, increased MNCV) | N/A | [89] |
| Tripterygium wilfordii Hook. F. | Triptolide | In vivo (SD rats) | 100 µg/kg | Intraperitoneal injection | Promoted peripheral nerve regeneration in rats with SNI (increased number of motoneurons, number of myelinated axons, diameter of nerve fibers, SFI, CMAP amplitude, MNCV, muscle fiber area) | Reduction of TNF-α, IL-β, and IL-6 expression | [90] |
| MUSHROOM | |||||||
| Amanita muscaria | Muscimol | In vivo (SD rats) | 400 μg/mL | Applied directly to the right L5 DRG | Promoted peripheral nerve regeneration in rats with SNI (prevented the development of thermal and mechanical hypersensitivity and mechanical allodynia, improved basal membrane integrity, and increased nerve fibers) | Normalization of PMP22 protein expression level by GABAergic modulation in the ipsilateral DRG | [91] |
| Hericium erinaceus | Aqueous extract | In vivo (SD rats) | 10 mL/kg | Oral administration | Promoted peripheral nerve regeneration in rats following peroneal nerve crush | Activation of signaling pathways (Akt, MAPK, c-Jun, c-Fos) and protein synthesis | [92] |
| Polysaccharide | In vivo (SD rats) | 30 mg/mL/kg | Oral administration | Promoted sensory functional recovery following peroneal nerve crush in rats (reduced withdrawal reflex latency) | Activation of Akt and p38 MAPK signaling and increased expression of RECA-1 | [93] | |
| Aqueous extract | In vivo (SD rats) | 10, 20 mL/kg | Oral administration | Promoted peripheral nerve regeneration in rats following peroneal nerve crush (increased PFI, improved axon morphology, and development of neuromuscular junction) | N/A | [94] | |
| Lignosus rhinocerotis | Aqueous extract | In vivo (SD rats) | 500, 1000 mg/kg | Oral administration | Promoted motor and sensory functional recovery in rats with SNI (improved WRL and toe-spreading reflex) | N/A | [95] |
| DECOCTION | |||||||
| Bogijetong | (1) Bogijietong decoction (18 ingredients) (2) A reconstituted formulation of BGJTD (BeD) with 4 ingredients (3) Angelica gigas (an ingredient in BeD) | In vitro (Primary neurons isolated from DRG at lumbar levels 4 and 5 in adult rats) | 400 mg/kg | Incubation | Promoted neurite outgrowth of DRG neurons | (1) BGJTD and BeD: Downregulation of TNF-α and p38, upregulation of p-ERK1/2; (2) Angelica gigas: Regulation of ERK1/2 activity and TNF-α production | [96] |
| In vivo (SD rats and BALB/c mice) | 400 mg/kg | Oral administration | Reduced latency time in rats | ||||
| Buyang Huanwu | Buyang Huanwu decoction (16 ingredients: Modified formulation) | In vivo (SD rats) | 1800 mg/kg | Oral administration paired with silicone rubber tube bridging a 10mm sciatic nerve defect | Promoted peripheral nerve regeneration in rats with sciatic nerve defect (increased nerve formation, myelinated axons, and endoneurial area) | N/A | [97] |
| Jiaweibugan | Jiaweibugan decoction (9 ingredients) | In vivo (Wistar rats) | 28.6 g/kg | Intragastric administration | Protective effect on peripheral nerve injury by playing an anti-oxidative role in a diabetic rat model (increased MNCV and serum levels of glutathione, decreased serum levels of MDA) | Downregulation of NF-kB p65 and p38 MAPK mRNA expression | [98] |
| Qian-Zheng-San | Qian-Zheng-San (3 ingredients: Typhonii rhizoma, Bombyx batryticatus, Scorpio) | In vivo (SD rats) | 1.75 g/mL | Oral gavage | Promoted peripheral nerve regeneration in rats with sciatic nerve defect (Increased SFI, MNCV, muscle wet weight, number of regenerated axons, axon diameter, nerve fiber diameter, myelin thickness, number of motor neurons in the lumbar spinal cord anterior horn) | N/A | [99] |

4.4. Experimental Strategies and Neuroprotective Effects of Complementary and Alternative Medicines (CAMs) against Peripheral Nerve Injury
4.4.1. CAMs with Neuroregenerative Potential
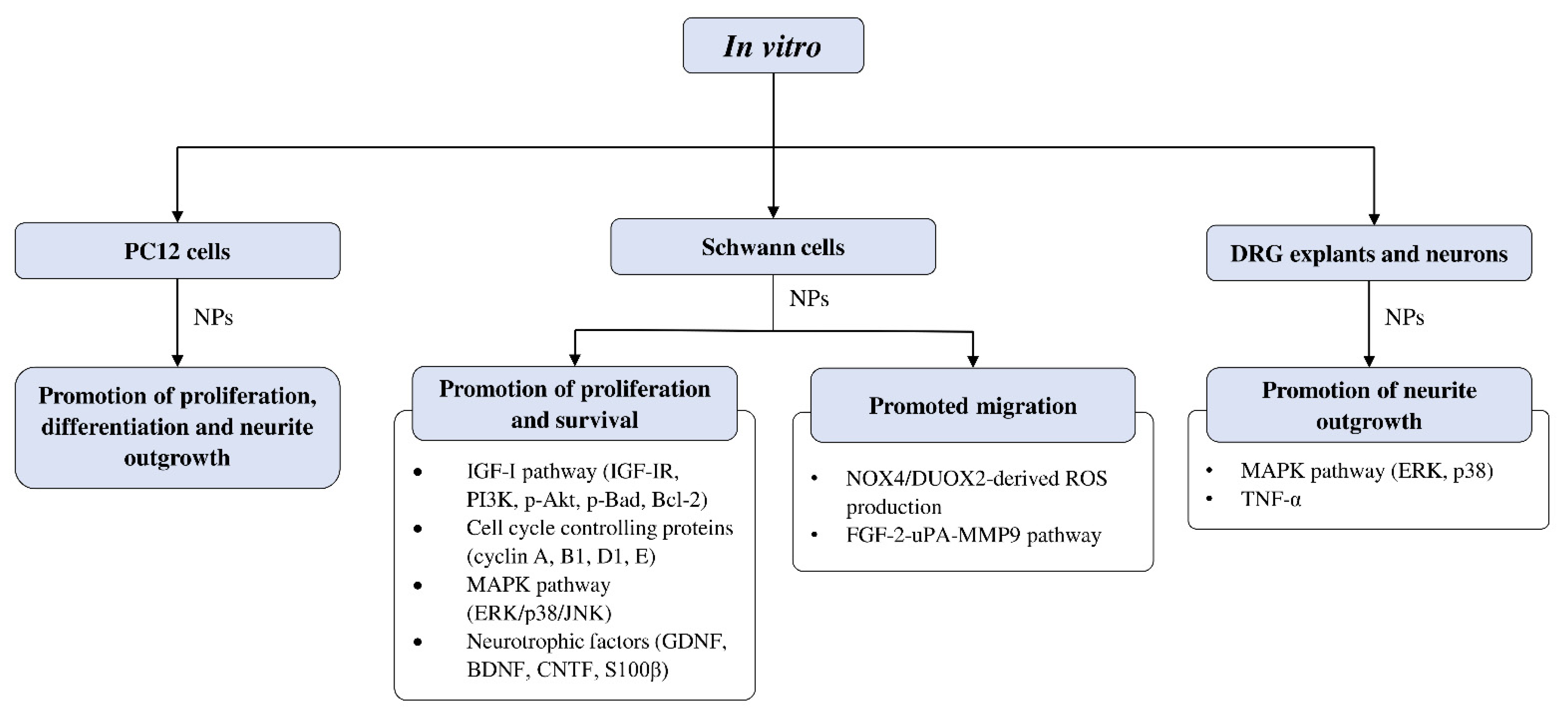
4.4.2. In Vitro Studies on Neuroregenerative Potential of CAMs
Effects of CAMs on Schwann Cell Activity In Vitro
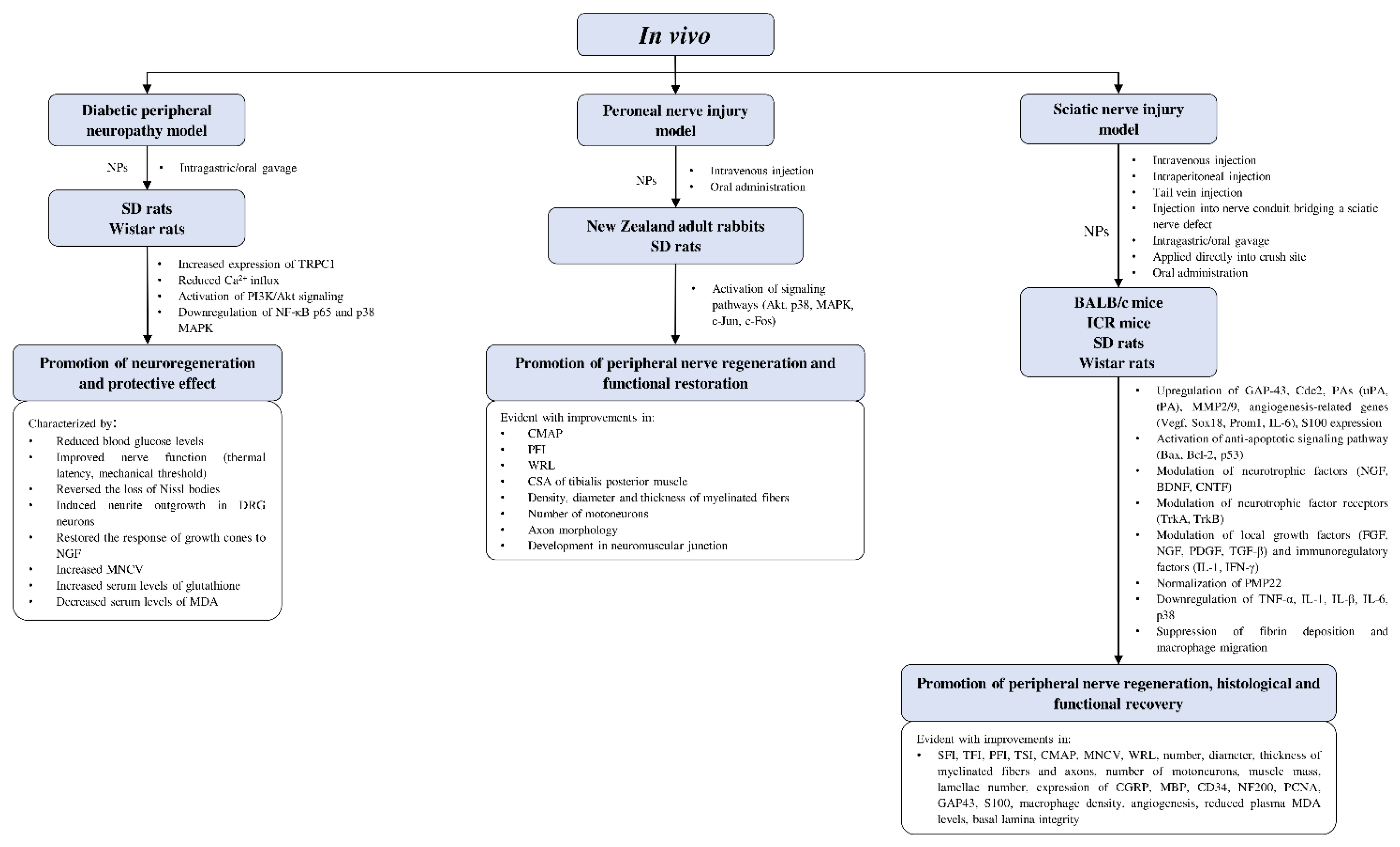
4.4.3. In Vivo Studies on Neuroregenerative Potential of CAMs
Diabetic Peripheral Neuropathy Model
Peroneal Nerve Injury Model
Sciatic Nerve Injury Model
4.4.4. Involvement of CAMs in Combinatorial Approaches for the Treatment of PNI
4.5. Limitations and Future Prospects
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Navarro, X. Functional evaluation of peripheral nerve regeneration and target reinnervation in animal models: A critical overview. Eur. J. Neurosci. 2016, 43, 271–286. [Google Scholar] [CrossRef]
- Wojtkiewicz, D.M.; Saunders, J.; Domeshek, L.; Novak, C.B.; Kaskutas, V.; Mackinnon, S.E. Social impact of peripheral nerve injuries. Hand 2015, 10, 161–167. [Google Scholar] [CrossRef]
- Li, N.Y.; Onor, G.I.; Lemme, N.J.; Gil, J.A. Epidemiology of peripheral nerve injuries in sports, exercise, and recreation in the United States, 2009–2018. Phys. Sportsmed. 2020, 49, 1–8. [Google Scholar] [CrossRef]
- Scholz, T.; Krichevsky, A.; Sumarto, A.; Jaffurs, D.; Wirth, G.A.; Paydar, K.; Evans, G.R.D. Peripheral nerve injuries: An international survey of current treatments and future perspectives. J. Reconstr. Microsurg. 2009, 25, 339–344. [Google Scholar] [CrossRef]
- Antoniadis, G.; Kretschmer, T.; Pedro, M.T.; König, R.W.; Heinen, C.P.G.; Richter, H.P. Iatrogenic nerve injuries - prevalence, diagnosis and treatment. Dtsch. Arztebl. Int. 2014, 111, 273–279. [Google Scholar] [CrossRef]
- Ciaramitaro, P.; Mondelli, M.; Logullo, F.; Grimaldi, S.; Battiston, B.; Sard, A.; Scarinzi, C.; Migliaretti, G.; Faccani, G.; Cocito, D. Traumatic peripheral nerve injuries: Epidemiological findings, neuropathic pain and quality of life in 158 patients. J. Peripher. Nerv. Syst. 2010, 15, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Grinsell, D.; Keating, C.P. Peripheral nerve reconstruction after injury: A review of clinical and experimental therapies. BioMed Res. Int. 2014, 2014, 698256. [Google Scholar] [CrossRef]
- Ray, W.Z.; Mackinnon, S.E. Management of nerve gaps: Autografts, allografts, nerve transfers, and end-to-side neurorrhaphy. Exp. Neurol. 2010, 223, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Wang, E.W.; Zhang, J.; Huang, J.H. Repairing peripheral nerve injury using tissue engineering techniques. Neural Regen. Res. 2015, 10, 1393–1394. [Google Scholar] [CrossRef] [PubMed]
- Houschyar, K.S.; Momeni, A.; Pyles, M.N.; Cha, J.Y.; Maan, Z.N.; Duscher, D.; Jew, O.S.; Siemers, F.; van Schoonhoven, J. The role of current techniques and concepts in peripheral nerve repair. Plast. Surg. Int. 2016, 2016, 4175293. [Google Scholar] [CrossRef]
- Carvalho, C.R.; Oliveira, J.M.; Reis, R.L. Modern trends for peripheral nerve repair and regeneration: Beyond the hollow nerve guidance conduit. Front. Bioeng. Biotechnol. 2019, 7, 337. [Google Scholar] [CrossRef]
- Brunelli, B.; Gorson, K.C. The use of complementary and alternative medicines by patients with peripheral neuropathy. J. Neurol. Sci. 2004, 218, 59–66. [Google Scholar] [CrossRef]
- Muthuraman, A.; Singh, N.; Jaggi, A.S. Effect of hydroalcoholic extract of Acorus calamus on tibial and sural nerve transection-induced painful neuropathy in rats. J. Nat. Med. 2011, 65, 282–292. [Google Scholar] [CrossRef]
- Zhao, X.; Xu, Y.; Zhao, Q.; Chen, C.R.; Liu, A.M.; Huang, Z.L. Curcumin exerts antinociceptive effects in a mouse model of neuropathic pain: Descending monoamine system and opioid receptors are differentially involved. Neuropharmacology 2012, 62, 843–854. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Park, H.J.; Kim, T.K.; Moon, D.E.; Lee, H.J. The effects of Ginkgo biloba extract EGB 761 on mechanical and cold allodynia in a rat model of neuropathic pain. Anesth. Analg. 2009, 108, 1958–1963. [Google Scholar] [CrossRef] [PubMed]
- Calixto, J.B. Efficacy, safety, quality control, marketing and regulatory guidelines for herbal medicines (phytotherapeutic agents). Braz. J. Med. Biol. Res. 2000, 33, 179–189. [Google Scholar] [CrossRef]
- Karimi, A.; Majlesi, M.; Rafieian-Kopaei, M. Herbal versus synthetic drugs; beliefs and facts. J. Nephropharmacology 2015, 4, 27–30. [Google Scholar]
- Lahlou, M. The success of natural products in drug discovery. Pharmacol. Pharm. 2013, 04, 17–31. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the nearly four decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef]
- Weaver, B.A. How taxol/paclitaxel kills cancer cells. Mol. Biol. Cell 2014, 25, 2677–2681. [Google Scholar] [CrossRef]
- Noble, R.L. The discovery of the vinca alkaloids—Chemotherapeutic agents against cancer. Biochem. Cell Biol. 1990, 68, 1344–1351. [Google Scholar] [CrossRef]
- Araújo-Filho, H.G.; Quintans-Júnior, L.J.; Barreto, A.S.; Almeida, J.R.G.S.; Barreto, R.S.S.; Quintans, J.S.S. Neuroprotective effect of natural products on peripheral nerve degeneration: A systematic review. Neurochem. Res. 2016, 41, 647–658. [Google Scholar] [CrossRef]
- Moher, D.; Shamseer, L.; Clarke, M.; Ghersi, D.; Liberati, A.; Petticrew, M.; Shekelle, P.; Stewart, L.A.; Group, P.-P. Preferred reporting items for systematic review and meta-analysis protocols (PRISMA-P) 2015 statement. Syst. Rev. 2015, 4, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Hussain, G.; Wang, J.; Rasul, A.; Anwar, H.; Qasim, M.; Zafar, S.; Aziz, N.; Razzaq, A.; Hussain, R.; de Aguilar, J.L.G.; et al. Current status of therapeutic approaches against peripheral nerve injuries: A detailed story from injury to recovery. Int. J. Biol. Sci. 2020, 16, 116–134. [Google Scholar] [CrossRef] [PubMed]
- Millesi, H. Bridging defects: Autologous nerve grafts. Acta Neurochir. Suppl. 2007, 100, 37–38. [Google Scholar] [CrossRef] [PubMed]
- Griffin, J.W.; Hogan, M.C.V.; Chhabra, A.B.; Deal, D.N. Peripheral nerve repair and reconstruction. J. Bone Joint Surg. Am. 2013, 95, 2144–2151. [Google Scholar] [CrossRef]
- Li, R.; Li, D.H.; Zhang, H.Y.; Wang, J.; Li, X.K.; Xiao, J. Growth factors-based therapeutic strategies and their underlying signaling mechanisms for peripheral nerve regeneration. Acta Pharmacol. Sin. 2020, 41, 1289–1300. [Google Scholar] [CrossRef]
- Kubiak, C.A.; Grochmal, J.; Kung, T.A.; Cederna, P.S.; Midha, R.; Kemp, S.W.P. Stem-cell–based therapies to enhance peripheral nerve regeneration. Muscle Nerve 2020, 61, 449–459. [Google Scholar] [CrossRef]
- Gordon, T.; English, A.W. Strategies to promote peripheral nerve regeneration: Electrical stimulation and/or exercise. Eur. J. Neurosci. 2016, 43, 336–350. [Google Scholar] [CrossRef]
- Choudhari, A.S.; Mandave, P.C.; Deshpande, M.; Ranjekar, P.; Prakash, O. Phytochemicals in cancer treatment: From preclinical studies to clinical practice. Front. Pharmacol. 2020, 10, 1614. [Google Scholar] [CrossRef]
- Kumar, G.P.; Khanum, F. Neuroprotective potential of phytochemicals. Pharmacogn. Rev. 2012, 6, 81–90. [Google Scholar] [CrossRef]
- Rotshenker, S. Wallerian degeneration: The innate-immune response to traumatic nerve injury. J. Neuroinflammation 2011, 8, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Menorca, R.M.G.; Fussell, T.S.; Elfar, J.C. Peripheral nerve trauma: Mechanisms of injury and recovery. Hand Clin. 2013, 29, 317–330. [Google Scholar] [CrossRef]
- Perry, V.H.; Brown, M.C.; Gordon, S. The macrophage response to central and peripheral nerve injury: A possible role for macrophages in regeneration. J. Exp. Med. 1987, 165, 1218–1223. [Google Scholar] [CrossRef]
- Tetzlaff, W. Tight junction contact events and temporary gap junctions in the sciatic nerve fibres of the chicken during Wallerian degeneration and subsequent regeneration. J. Neurocytol. 1982, 11, 839–858. [Google Scholar] [CrossRef]
- Sulaiman, W.; Gordon, T. Neurobiology of peripheral nerve injury, regeneration, and functional recovery: From bench top research to bedside application. Ochsner J. 2013, 13, 100–108. [Google Scholar] [PubMed]
- Simons, M.; Trotter, J. Wrapping it up: The cell biology of myelination. Curr. Opin. Neurobiol. 2007, 17, 533–540. [Google Scholar] [CrossRef] [PubMed]
- Arancibia-Carcamo, I.L.; Attwell, D. The node of Ranvier in CNS pathology. Acta Neuropathol. 2014, 128, 161–175. [Google Scholar] [CrossRef]
- Jessen, K.R.; Mirsky, R. The success and failure of the Schwann cell response to nerve injury. Front. Cell. Neurosci. 2019, 13, 1–33. [Google Scholar] [CrossRef]
- Arthur-Farraj, P.J.; Latouche, M.; Wilton, D.K.; Quintes, S.; Chabrol, E.; Banerjee, A.; Woodhoo, A.; Jenkins, B.; Rahman, M.; Turmaine, M.; et al. c-Jun reprograms Schwann cells of injured nerves to generate a repair cell essential for regeneration. Neuron 2012, 75, 633–647. [Google Scholar] [CrossRef]
- Atanasoski, S.; Shumas, S.; Dickson, C.; Scherer, S.S.; Suter, U. Differential cyclin D1 requirements of proliferating Schwann cells during development and after injury. Mol. Cell. Neurosci. 2001, 18, 581–592. [Google Scholar] [CrossRef]
- Kim, H.A.; Pomeroy, S.L.; Whoriskey, W.; Pawlitzky, I.; Benowitz, L.I.; Sicinski, P.; Stiles, C.D.; Roberts, T.M. A developmentally regulated switch regenerative growth of Schwann cells through cyclin D1. Neuron 2000, 26, 405–416. [Google Scholar] [CrossRef]
- Yang, D.P.; Zhang, D.P.; Mak, K.S.; Bonder, D.E.; Scott, L.; Kim, H.A. Schwann cell proliferation during Wallerian degeneration is not necessary for regeneration and remyelination of the peripheral nerves: Axon-dependent removal of newly generated Schwann cells by apoptosis. Mol. Cell. Neurosci. 2008, 38, 80–88. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Zhao, H.; Yang, L.; Li, L.; Zhang, T.; Pan, J.; Meng, Y.; Shen, W.; Yuan, Y. Achyranthes bidentata polypeptides promotes migration of Schwann cells via NOX4/DUOX2-dependent ROS production in rats. Neurosci. Lett. 2019, 696, 99–107. [Google Scholar] [CrossRef]
- Cheng, Q.; Yuan, Y.; Sun, C.; Gu, X.; Cao, Z.; Ding, F. Neurotrophic and neuroprotective actions of Achyranthes bidentata polypeptides on cultured dorsal root ganglia of rats and on crushed common peroneal nerve of rabbits. Neurosci. Lett. 2014, 562, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Q.; Jiang, C.; Wang, C.; Yu, S.; Zhang, Q.; Gu, X.; Ding, F. The Achyranthes bidentata polypeptide k fraction enhances neuronal growth in vitro and promotes peripheral nerve regeneration after crush injury in vivo. Neural Regen. Res. 2014, 9, 2142–2150. [Google Scholar] [CrossRef]
- Wang, Y.; Shen, W.; Yang, L.; Zhao, H.; Gu, W.; Yuan, Y. The protective effects of Achyranthes bidentata polypeptides on rat sciatic nerve crush injury causes modulation of neurotrophic factors. Neurochem. Res. 2012, 38, 538–546. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Shen, H.; Yao, J.; Hu, N.; Ding, F.; Gu, X. The protective effects of Achyranthes bidentata polypeptides in an experimental model of mouse sciatic nerve crush injury. Brain Res. Bull. 2010, 81, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Ding, F.; Cheng, Q.; Gu, X. The repair effects of Achyranthes bidentata extract on the crushed common peroneal nerve of rabbits. Fitoterapia 2008, 79, 161–167. [Google Scholar] [CrossRef]
- Ju, D.T.; Liao, H.E.; Shibu, M.A.; Ho, T.J.; Padma, V.V.; Tsai, F.J.; Chung, L.C.; Day, C.H.; Lin, C.C.; Huang, C.Y. Nerve Regeneration potential of protocatechuic acid in RSC96 Schwann cells by induction of cellular proliferation and migration through IGF-IR-PI3K-Akt signaling. Chin. J. Physiol. 2015, 58, 412–419. [Google Scholar] [CrossRef]
- Chang, Y.M.; Ye, C.X.; Ho, T.J.; Tsai, T.N.; Chiu, P.L.; Tsai, C.C.; Lin, Y.M.; Kuo, C.H.; Tsai, F.J.; Tsai, C.H.; et al. Alpinia oxyphylla Miquel fruit extract activates MAPK-mediated signaling of PAs and MMP2/9 to induce Schwann cell migration and nerve regeneration. Int. J. Artif. Organs 2014, 37, 402–413. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.H.; Chen, J.J. The mechanism of astragaloside IV promoting sciatic nerve regeneration. Neural Regen. Res. 2013, 8, 2256–2265. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.Y.; Yao, C.H.; Liu, B.S.; Liu, C.J.; Chen, G.W.; Chen, Y.S. The role of astragaloside in regeneration of the peripheral nerve system. J. Biomed. Mater. Res. A 2006, 76, 463–469. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.S.; Chen, C.C.; Chang, L.C.; Yao, C.H.; Hsu, Y.M.; Lin, J.H.; Yang, T.Y.; Chen, Y.H. Increased calcitonin gene-related peptide and macrophages are involved in Astragalus membranaceus-mediated peripheral nerve regeneration in rats. Am. J. Chin. Med. 2018, 46, 69–86. [Google Scholar] [CrossRef]
- Fang, W.K.; Ko, F.Y.; Wang, H.L.; Kuo, C.H.; Chen, L.M.; Tsai, F.J.; Tsai, C.H.; Chen, Y.S.; Kuo, W.W.; Huang, C.Y. The proliferation and migration effects of huangqi on RSC96 Schwann cells. Am. J. Chin. Med. 2009, 37, 945–959. [Google Scholar] [CrossRef] [PubMed]
- Hussin, H.M.; Lawi, M.M.; Haflah, N.H.M.; Kassim, A.Y.M.; Idrus, R.B.H.; Lokanathan, Y. Centella asiatica (L.)-neurodifferentiated mesenchymal stem cells promote the regeneration of peripheral nerve. Tissue Eng. Regen. Med. 2020, 17, 237–251. [Google Scholar] [CrossRef]
- Huang, C.Y.; Kuo, W.W.; Shibu, M.A.; Hsueh, M.F.; Chen, Y.S.; Tsai, F.J.; Yao, C.H.; Lin, C.C.; Pan, L.F.; Ju, D.T. Citrus medica var. sarcodactylis (foshou) activates fibroblast growth factor-2 signaling to induce migration of RSC96 Schwann cells. Am. J. Chin. Med. 2014, 42, 443–452. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.T.; Tsai, Y.L.; Chen, Y.S.; Jong, G.P.; Chen, W.K.; Wang, H.L.; Tsai, F.J.; Tsai, C.H.; Lai, T.Y.; Tzang, B.S.; et al. Dangshen (Codonopsis pilosula) activates IGF-I and FGF-2 pathways to induce proliferation and migration effects in RSC96 Schwann cells. Am. J. Chin. Med. 2010, 38, 359–372. [Google Scholar] [CrossRef]
- Tamaddonfard, E.; Farshid, A.A.; Ahmadian, E.; Hamidhoseyni, A. Crocin enhanced functional recovery after sciatic nerve crush injury in rats. Iran. J. Basic Med. Sci. 2013, 16, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Tehranipour, M.; Javaheri, R. Neuroprotective effect of Curcuma longa alcoholic extract on peripheral nerves degeneration after sciatic nerve compression in rats. J. Biol. Sci. 2009, 9, 889–893. [Google Scholar] [CrossRef][Green Version]
- Noorafshan, A.; Omidi, A.; Karbalay-Doust, S. Curcumin protects the dorsal root ganglion and sciatic nerve after crush in rat. Pathol. Res. Pract. 2011, 207, 577–582. [Google Scholar] [CrossRef]
- Noorafshan, A.; Omidi, A.; Karbalay-Doust, S.; Aliabadi, E.; Dehghani, F. Effects of curcumin on the dorsal root ganglion structure and functional recovery after sciatic nerve crush in rat. Micron 2011, 42, 449–455. [Google Scholar] [CrossRef]
- Ma, J.; Liu, J.; Yu, H.; Wang, Q.; Chen, Y.; Xiang, L. Curcumin promotes nerve regeneration and functional recovery in rat model of nerve crush injury. Neurosci. Lett. 2013, 547, 26–31. [Google Scholar] [CrossRef]
- Tello Velasquez, J.; Nazareth, L.; Quinn, R.J.; Ekberg, J.A.K.; St John, J.A. Stimulating the proliferation, migration and lamellipodia of Schwann cells using low-dose curcumin. Neuroscience 2016, 324, 140–150. [Google Scholar] [CrossRef]
- Yüce, S.; Cemal Gökçe, E.; Işkdemir, A.; Koç, E.R.; Cemil, D.B.; Gökçe, A.; Sargon, M.F. An experimental comparison of the effects of propolis, curcumin, and methylprednisolone on crush injuries of the sciatic nerve. Ann. Plast. Surg. 2015, 74, 684–692. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.M.; Namgung, U.K.; Hong, K.E. Growth-promoting activity of sanyak (Dioscoreae rhizoma) extract on injured sciatic nerve in rats. JAMS J. Acupunct. Meridian Stud. 2009, 2, 228–235. [Google Scholar] [CrossRef]
- Chen, B.; Niu, S.P.; Wang, Z.Y.; Wang, Z.W.; Deng, J.X.; Zhang, P.X.; Yin, X.F.; Han, N.; Kou, Y.H.; Jiang, B.G. Local administration of icariin contributes to peripheral nerve regeneration and functional recovery. Neural Regen. Res. 2015, 10, 84–89. [Google Scholar] [CrossRef]
- Kou, Y.; Wang, Z.; Wu, Z.; Zhang, P.; Zhang, Y.; Yin, X.; Wong, X.; Qiu, G.; Jiang, B. Epimedium extract promotes peripheral nerve regeneration in rats. Evid. Based Complement. Altern. Med. 2013, 2013, 954798. [Google Scholar] [CrossRef]
- Liu, B.S.; Yao, C.H.; Hsu, S.H.; Yeh, T.S.; Chen, Y.S.; Kao, S.T. A novel use of genipin-fixed gelatin as extracellular matrix for peripheral nerve regeneration. J. Biomater. Appl. 2004, 19, 21–34. [Google Scholar] [CrossRef]
- Zuo, W.; Xu, F.; Zhang, K.; Zheng, L.; Zhao, J. Proliferation-enhancing effects of gastrodin on RSC96 Schwann cells by regulating ERK1/2 and PI3K signaling pathways. Biomed. Pharmacother. 2016, 84, 747–753. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Zhou, X.; He, B.; Dai, T.; Zheng, C.; Yang, C.; Zhu, S.; Zhu, J.; Zhu, Q.; Liu, X. Ginkgo biloba extract (EGb 761) promotes peripheral nerve regeneration and neovascularization after acellular nerve allografts in a rat model. Cell. Mol. Neurobiol. 2015, 35, 273–282. [Google Scholar] [CrossRef]
- Hsu, S.H.; Chang, C.J.; Tang, C.M.; Lin, F.T. In vitro and in vivo effects of Ginkgo biloba extract EGb 761 on seeded Schwann cells within poly (DL-lactic acid-co-glycolic acid) conduits for peripheral nerve regeneration. J. Biomater. Appl. 2004, 19, 163–182. [Google Scholar] [CrossRef]
- Lu, M.C.; Lai, T.Y.; Hwang, J.M.; Chen, H.T.; Chang, S.H.; Tsai, F.J.; Wang, H.L.; Lin, C.C.; Kuo, W.W.; Huang, C.Y. Proliferation- and migration-enhancing effects of ginseng and ginsenoside Rg1 through IGF-I- and FGF-2-signaling pathways on RSC96 Schwann cells. Cell Biochem. Funct. 2009, 27, 186–192. [Google Scholar] [CrossRef]
- Ma, J.; Li, W.; Tian, R.; Lei, W. Ginsenoside Rg1 promotes peripheral nerve regeneration in rat model of nerve crush injury. Neurosci. Lett. 2010, 478, 66–71. [Google Scholar] [CrossRef]
- Wang, L.; Yuan, D.; Zhang, D.; Zhang, W.; Liu, C.; Cheng, H.; Song, Y.; Tan, Q. Ginsenoside Re promotes nerve regeneration by facilitating the proliferation, differentiation and migration of Schwann cells via the ERK- and JNK-dependent pathway in rat model of sciatic nerve crush injury. Cell. Mol. Neurobiol. 2015, 35, 827–840. [Google Scholar] [CrossRef]
- Renno, W.M.; Al-Maghrebi, M.; Al-Banaw, A. (-)-Epigallocatechin-3-gallate (EGCG) attenuates functional deficits and morphological alterations by diminishing apoptotic gene overexpression in skeletal muscles after sciatic nerve crush injury. Naunyn. Schmiedebergs. Arch. Pharmacol. 2012, 385, 807–822. [Google Scholar] [CrossRef]
- Renno, W.M.; Al-Maghrebi, M.; Alshammari, A.; George, P. (-)-Epigallocatechin-3-gallate (EGCG) attenuates peripheral nerve degeneration in rat sciatic nerve crush injury. Neurochem. Int. 2013, 62, 221–231. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Jiang, X.; Yin, G.; Lin, H. Syringic acid promotes proliferation and migration of Schwann cells via down-regulating MiR-451-5p. Acta Biochim. Biophys. Sin. (Shanghai) 2019, 51, 1198–1207. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Liu, Y.; Yang, G.; Xu, Z.; Chen, J. Ursolic acid induces neural regeneration after sciatic nerve injury. Neural Regen. Res. 2013, 8, 2510–2519. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Tian, L.; He, L.; Chen, N.; Ramakrishna, S.; So, K.F.; Mo, X. Lycium barbarum polysaccharide encapsulated poly lactic-co-glycolic acid nanofibers: Cost effective herbal medicine for potential application in peripheral nerve tissue engineering. Sci. Rep. 2018, 8, 8669. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Huang, C.Y.; Tsai, F.J.; Tsai, C.C.; Yao, C.H.; Chen, Y.S. Growth-promoting effects of quercetin on peripheral nerves in rats. Int. J. Artif. Organs 2011, 34, 1095–1105. [Google Scholar] [CrossRef]
- Lu, M.; Yi, T.; Xiong, Y.; Wang, Q.; Yin, N. Cortex Mori Radicis extract promotes neurite outgrowth in diabetic rats by activating PI3K/AKT signaling and inhibiting Ca2+ influx associated with the upregulation of transient receptor potential canonical channel 1. Mol. Med. Rep. 2020, 21, 320–328. [Google Scholar] [CrossRef]
- Hsiang, S.W.; Lee, H.C.; Tsai, F.J.; Tsai, C.C.; Yao, C.H.; Chen, Y.S. Puerarin accelerates peripheral nerve regeneration. Am. J. Chin. Med. 2011, 39, 1207–1217. [Google Scholar] [CrossRef]
- Chen, H.-T.; Yao, C.-H.; Chao, P.-D.L.; Hou, Y.-C.; Chiang, H.-M.; Hsieh, C.-C.; Ke, C.-J.; Chen, Y.-S. Effect of serum metabolites of Pueraria lobata in rats on peripheral nerve regeneration: In vitro and in vivo studies. J. Biomed. Mater. Res. Part B Appl. Biomater. 2007, 84B, 256–262. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, P.; Kou, Y.; Yin, X.; Han, N.; Jiang, B. Hedysari extract improves regeneration after peripheral nerve injury by enhancing the amplification effect. PLoS ONE 2013, 8, e67921. [Google Scholar] [CrossRef]
- Wei, S.Y.; Zhang, P.X.; Han, N.; Dang, Y.; Zhang, H.B.; Zhang, D.Y.; Fu, Z.G.; Jiang, B.G. Effects of Hedysari polysaccharides on regeneration and function recovery following peripheral nerve injury in rats. Am. J. Chin. Med. 2009, 37, 57–67. [Google Scholar] [CrossRef]
- Sheng, Q.S.; Wang, Z.J.; Zhang, J.; Zhang, Y.G. Salidroside promotes peripheral nerve regeneration following crush injury to the sciatic nerve in rats. Neuroreport 2013, 24, 217–223. [Google Scholar] [CrossRef]
- Zuo, W.; Wu, H.; Zhang, K.; Lv, P.; Xu, F.; Jiang, W.; Zheng, L.; Zhao, J. Baicalin promotes the viability of Schwann cells in vitro by regulating neurotrophic factors. Exp. Ther. Med. 2017, 14, 507–514. [Google Scholar] [CrossRef] [PubMed]
- Morani, A.S.; Bodhankar, S.L.; Mohan, V.; Thakurdesai, P.A. Ameliorative effects of standardized extract from Trigonella foenum-graecum L. seeds on painful peripheral neuropathy in rats. Asian Pac. J. Trop. Med. 2012, 5, 385–390. [Google Scholar] [CrossRef]
- Zhang, Y.G.; Sheng, Q.S.; Wang, H.K.; Lv, L.; Zhang, J.; Chen, J.M.; Xu, H. Triptolide improves nerve regeneration and functional recovery following crush injury to rat sciatic nerve. Neurosci. Lett. 2014, 561, 198–202. [Google Scholar] [CrossRef] [PubMed]
- Naik, A.K.; Latham, J.R.; Obradovic, A.; Jevtovic-Todorovic, V. Dorsal root ganglion application of muscimol prevents hyperalgesia and stimulates myelin protein expression after sciatic nerve injury in rats. Anesth. Analg. 2012, 114, 674–682. [Google Scholar] [CrossRef]
- Wong, K.H.; Kanagasabapathy, G.; Naidu, M.; David, P.; Sabaratnam, V. Hericium erinaceus (Bull.: Fr.) Pers., a medicinal mushroom, activates peripheral nerve regeneration. Chin. J. Integr. Med. 2016, 22, 759–767. [Google Scholar] [CrossRef]
- Wong, K.H.; Kanagasabapathy, G.; Bakar, R.; Phan, C.W.; Sabaratnam, V. Restoration of sensory dysfunction following peripheral nerve injury by the polysaccharide from culinary and medicinal mushroom, Hericium erinaceus (Bull.: Fr.) Pers. through its neuroregenerative action. Food Sci. Technol. 2015, 35, 712–721. [Google Scholar] [CrossRef]
- Wong, K.H.; Naidu, M.; David, P.; Abdulla, M.A.; Abdullah, N.; Kuppusamy, U.R.; Sabaratnam, V. Peripheral nerve regeneration following crush injury to rat peroneal nerve by aqueous extract of medicinal mushroom Hericium erinaceus (Bull.: Fr) Pers. (Aphyllophoromycetideae). Evid. Based Complement. Altern. Med. 2011, 2011, 580752. [Google Scholar] [CrossRef] [PubMed]
- Farha, M.; Parkianathan, L.; Amir, N.A.I.A.; Sabaratnam, V.; Wong, K.H. Functional recovery enhancement by tiger milk mushroom, Lignosus rhinocerotis in a sciatic nerve crush injury model and morphological study of its neurotoxicity. J. Anim. Plant Sci. 2019, 29, 930–942. [Google Scholar]
- Kim, K.J.; Namgung, U.; Cho, C.S. Protective effects of Bogijetong decoction and its selected formula on neuropathic insults in streptozotocin-induced diabetic animals. Evid. Based Complement. Altern. Med. 2017, 2017, 4296318. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.S.; Yao, C.H.; Chen, T.H.; Hsieh, C.L.; Lao, C.J.; Tsai, C.C. Effect of Buyang Huanwu decoction on peripheral nerve regeneration using silicone rubber chambers. Am. J. Chin. Med. 2001, 29, 423–432. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, Z.; Ye, R.; He, Y.; Li, Y.; Qiu, X. Protective effect of Jiaweibugan decoction against diabetic peripheral neuropathy. Neural Regen. Res. 2013, 8, 1113–1121. [Google Scholar] [CrossRef]
- Wang, Z.Y.; Qin, L.H.; Zhang, W.G.; Zhang, P.X.; Jiang, B.G. Qian-Zheng-San promotes regeneration after sciatic nerve crush injury in rats. Neural Regen. Res. 2019, 14, 683–691. [Google Scholar] [CrossRef]
- Nocera, G.; Jacob, C. Mechanisms of Schwann cell plasticity involved in peripheral nerve repair after injury. Cell Mol. Life Sci. 2020, 77, 3977–3989. [Google Scholar] [CrossRef]
- Srinivasulu, C.; Ramgopal, M.; Ramanjaneyulu, G.; Anuradha, C.M.; Suresh Kumar, C. Syringic acid (SA) ‒ A review of its occurrence, biosynthesis, pharmacological and industrial importance. Biomed. Pharmacother. 2018, 108, 547–557. [Google Scholar] [CrossRef]
- Baliga, M.S.; Shivashankara, A.R.; Venkatesh, S.; Bhat, H.P.; Palatty, P.L.; Bhandari, G.; Rao, S. Phytochemicals in the prevention of ethanol-induced hepatotoxicity: A revisit. In Dietary Interventions in Liver Disease: Foods, Nutrients, and Dietary Supplements; Watson, R.R., Preedy, V.R., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 79–89. ISBN 978-0-12-814466-4. [Google Scholar]
- Li, Y.; Yao, J.; Han, C.; Yang, J.; Chaudhry, M.T.; Wang, S.; Liu, H.; Yin, Y. Quercetin, inflammation and immunity. Nutrients 2016, 8, 167. [Google Scholar] [CrossRef]
- Desbarats, J.; Birge, R.B.; Mimouni-Rongy, M.; Weinstein, D.E.; Palerme, J.S.; Newell, M.K. Fas engagement induces neurite growth through ERK activation and P35 upregulation. Nat. Cell Biol. 2003, 5, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Schneider-Schaulies, J.; Kirchhoff, F.; Archelos, J.; Schachner, M. Down-regulation of myelin-associated glycoprotein on Schwann cells by interferon-γ and tumor necrosis factor-α affects neurite outgrowth. Neuron 1991, 7, 995–1005. [Google Scholar] [CrossRef]
- Larsson, K.; Rydevik, B.; Olmarker, K. Disc related cytokines inhibit axonal outgrowth from dorsal root ganglion cells in vitro. Spine 2005, 30, 621–624. [Google Scholar] [CrossRef] [PubMed]
- Kato, K.; Liu, H.; Kikuchi, S.I.; Myers, R.R.; Shubayev, V.I. Immediate anti-tumor necrosis factor-α (etanercept) therapy enhances axonal regeneration after sciatic nerve crush. J. Neurosci. Res. 2010, 88, 360–368. [Google Scholar] [CrossRef] [PubMed]
- Newbern, J.M.; Li, X.; Shoemaker, S.E.; Zhou, J.; Zhong, J.; Wu, Y.; Bonder, D.; Hollenback, S.; Coppola, G.; Geschwind, D.H.; et al. Specific functions for ERK/MAPK signaling during PNS development. Neuron 2011, 69, 91–105. [Google Scholar] [CrossRef]
- Ishii, A.; Furusho, M.; Bansal, R. Sustained activation of ERK1/2 MAPK in oligodendrocytes and Schwann cells enhances myelin growth and stimulates oligodendrocyte progenitor expansion. J. Neurosci. 2013, 33, 175–186. [Google Scholar] [CrossRef]
- Harrisingh, M.C.; Perez-Nadales, E.; Parkinson, D.B.; Malcolm, D.S.; Mudge, A.W.; Lloyd, A.C. The Ras/Raf/ERK signalling pathway drives Schwann cell dedifferentiation. EMBO J. 2004, 23, 3061–3071. [Google Scholar] [CrossRef] [PubMed]
- Agthong, S.; Kaewsema, A.; Tanomsridejchai, N.; Chentanez, V. Activation of MAPK ERK in peripheral nerve after injury. BMC Neurosci. 2006, 7, 45. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, T.; Sabit, H.; Oya, T.; Ishii, Y.; Hamashima, T.; Tokunaga, A.; Ishizawa, S.; Jie, S.; Kurashige, Y.; Matsushima, T.; et al. Activation of MAP kinases, Akt and PDGF receptors in injured peripheral nerves. J. Peripher. Nerv. Syst. 2009, 14, 165–176. [Google Scholar] [CrossRef]
- Huang, H.; Sun, Z.; Liu, H.; Ma, J.; Hu, M. ERK/MAPK and PI3K/AKT signal channels simultaneously activated in nerve cell and axon after facial nerve injury. Saudi J. Biol. Sci. 2017, 24, 1853–1858. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.P.; Kim, J.; Syed, N.; Tung, Y.J.; Bhaskaran, A.; Mindos, T.; Mirsky, R.; Jessen, K.R.; Maurel, P.; Parkinson, D.B.; et al. P38 MAPK activation promotes denervated Schwann cell phenotype and functions as a negative regulator of Schwann cell differentiation and myelination. J. Neurosci. 2012, 32, 7158–7168. [Google Scholar] [CrossRef]
- Pines, J. Four-dimensional control of the cell cycle. Nat. Cell Biol. 1999, 153, E73. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.; Kaldis, P. Cdks, cyclins and CKIs: Roles beyond cell cycle regulation. Development 2013, 140, 3079–3093. [Google Scholar] [CrossRef]
- Mairet-Coello, G.; Tury, A.; DiCicco-Bloom, E. Insulin-like growth factor-1 promotes G1/S cell cycle progression through bidirectional regulation of cyclins and cyclin-dependent kinase inhibitors via the phosphatidylinositol 3-kinase/Akt pathway in developing rat cerebral cortex. J. Neurosci. 2009, 29, 775–788. [Google Scholar] [CrossRef] [PubMed]
- Syroid, D.E.; Zorick, T.S.; Arbet-Engels, C.; Kilpatrick, T.J.; Eckhart, W.; Lemke, G. A role for insulin-like growth factor-I in the regulation of Schwann cell survival. J. Neurosci. 1999, 19, 2059–2068. [Google Scholar] [CrossRef] [PubMed]
- Hansson, H.A.; Dahlin, L.B.; Danielsen, N.; Fryklund, L.; Nachemson, A.K.; Polleryd, P.; Rozell, B.; Skottner, A.; Stemme, S.; Lundborg, G. Evidence indicating trophic importance of IGF-I in regenerating peripheral nerves. Acta Physiol. Scand. 1986, 126, 609–614. [Google Scholar] [CrossRef]
- Cheng, H.L.; Randolph, A.; Yee, D.; Delafontaine, P.; Tennekoon, G.; Feldman, E.L. Characterization of insulin-like growth factor-I and its receptor and binding proteins in transected nerves and cultured Schwann cells. J. Neurochem. 1996, 66, 525–536. [Google Scholar] [CrossRef]
- Apel, P.J.; Ma, J.; Callahan, M.; Northam, C.N.; Alton, T.B.; Sonntag, W.E.; Li, Z. Effect of locally delivered IGF-1 on nerve regeneration during aging: An experimental study in rats. Muscle Nerve 2010, 41, 335–341. [Google Scholar] [CrossRef]
- Zhu, H.; Xue, C.; Yao, M.; Wang, H.; Zhang, P.; Qian, T.; Zhou, S.; Li, S.; Yu, B.; Wang, Y.; et al. MIR-129 controls axonal regeneration via regulating insulin-like growth factor-1 in peripheral nerve injury. Cell Death Dis. 2018, 9, 720. [Google Scholar] [CrossRef]
- Sowndhararajan, K.; Deepa, P.; Kim, M.; Park, S.J.; Kim, S. Neuroprotective and cognitive enhancement potentials of baicalin: A review. Brain Sci. 2018, 8, 104. [Google Scholar] [CrossRef]
- Stankoff, B.; Aigrot, M.-S.; Noël, F.; Wattilliaux, A.; Zalc, B.; Lubetzki, C. Ciliary neurotrophic factor (CNTF) enhances myelin formation: A novel role for CNTF and CNTF-related molecules. J. Neurosci. 2002, 22, 9221–9227. [Google Scholar] [CrossRef]
- Xiao, J.; Hughes, R.A.; Lim, J.Y.; Wong, A.W.; Ivanusic, J.J.; Ferner, A.H.; Kilpatrick, T.J.; Murray, S.S. A small peptide mimetic of brain-derived neurotrophic factor promotes peripheral myelination. J. Neurochem. 2013, 125, 386–398. [Google Scholar] [CrossRef] [PubMed]
- Wilhelm, J.C.; Xu, M.; Cucoranu, D.; Chmielewski, S.; Holmes, T.; Lau, K.S.; Bassell, G.J.; English, A.W. Cooperative roles of BDNF expression in neurons and Schwann cells are modulated by exercise to facilitate nerve regeneration. J. Neurosci. 2012, 32, 5002–5009. [Google Scholar] [CrossRef]
- Tannemaat, M.R.; Eggers, R.; Hendriks, W.T.; De Ruiter, G.C.W.; Van Heerikhuize, J.J.; Pool, C.W.; Malessy, M.J.A.; Boer, G.J.; Verhaagen, J. Differential effects of lentiviral vector-mediated overexpression of nerve growth factor and glial cell line-derived neurotrophic factor on regenerating sensory and motor axons in the transected peripheral nerve. Eur. J. Neurosci. 2008, 28, 1467–1479. [Google Scholar] [CrossRef] [PubMed]
- Yun, Y.R.; Won, J.E.; Jeon, E.; Lee, S.; Kang, W.; Jo, H.; Jang, J.H.; Shin, U.S.; Kim, H.W. Fibroblast growth factors: Biology, function, and application for tissue regeneration. J. Tissue Eng. 2010, 1, 218142. [Google Scholar] [CrossRef]
- Holland, E.C.; Varmus, H.E. Basic fibroblast growth factor induces cell migration and proliferation after glia-specific gene transfer in mice. Proc. Natl. Acad. Sci. USA 1998, 95, 1218–1223. [Google Scholar] [CrossRef] [PubMed]
- Hossain, W.A.; Morest, D.K. Fibroblast growth factors (FGF-1, FGF-2) promote migration and neurite growth of mouse cochlear ganglion cells in vitro: Immunohistochemistry and antibody perturbation. J. Neurosci. Res. 2000, 62, 40–55. [Google Scholar] [CrossRef]
- Chen, B.; Hu, R.; Min, Q.; Li, Y.; Parkinson, D.B.; Dun, X. FGF5 regulates Schwann cell migration and adhesion. Front. Cell. Neurosci. 2020, 14, 1–12. [Google Scholar] [CrossRef]
- Li, J.; Wuliji, O.; Li, W.; Jiang, Z.G.; Ghanbari, H.A. Oxidative stress and neurodegenerative disorders. Int. J. Mol. Sci. 2013, 14, 24438–24475. [Google Scholar] [CrossRef]
- Areti, A.; Yerra, V.G.; Naidu, V.G.M.; Kumar, A. Oxidative stress and nerve damage: Role in chemotherapy induced peripheral neuropathy. Redox Biol. 2014, 2, 289–295. [Google Scholar] [CrossRef]
- Li, Y.; Shi, Z.; Yu, X.; Feng, P.; Wang, X.-J. The effects of urotensin II on migration and invasion are mediated by NADPH oxidase-derived reactive oxygen species through the c-Jun N-terminal kinase pathway in human hepatoma cells. Peptides 2017, 88, 106–114. [Google Scholar] [CrossRef]
- Ma, S.; Fu, A.; Lim, S.; Chiew, G.G.Y.; Luo, K.Q. MnSOD mediates shear stress-promoted tumor cell migration and adhesion. Free Radic. Biol. Med. 2018, 129, 46–58. [Google Scholar] [CrossRef]
- Yu, J.S.L.; Cui, W. Proliferation, survival and metabolism: The role of PI3K/AKT/MTOR signalling in pluripotency and cell fate determination. Development 2016, 143, 3050–3060. [Google Scholar] [CrossRef]
- Anitha, M.; Gondha, C.; Sutliff, R.; Parsadanian, A.; Mwangi, S.; Sitaraman, S.V.; Srinivasan, S. GDNF rescues hyperglycemia-induced diabetic enteric neuropathy through activation of the PI3K/Akt pathway. J. Clin. Invest. 2006, 116, 344–356. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Gong, H.Y.; Xu, L. PVT1 protects diabetic peripheral neuropathy via PI3K/AKT pathway. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 6905–6911. [Google Scholar] [CrossRef] [PubMed]
- Shim, S.; Ming, G.L. Roles of channels and receptors in the growth cone during PNS axonal regeneration. Exp. Neurol. 2010, 223, 38–44. [Google Scholar] [CrossRef]
- Wu, D.; Huang, W.; Richardson, P.M.; Priestley, J.V.; Liu, M. TRPC4 in rat dorsal root ganglion neurons is increased after nerve injury and is necessary for neurite outgrowth. J. Biol. Chem. 2008, 283, 416–426. [Google Scholar] [CrossRef]
- Heo, D.K.; Chung, W.Y.; Park, H.W.; Yuan, J.P.; Lee, M.G.; Kim, J.Y. Opposite regulatory effects of TRPC1 and TRPC5 on neurite outgrowth in PC12 cells. Cell. Signal. 2012, 24, 899–906. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Tang, J.; Li, G.; Berti-Mattera, L.; Lee, C.A.; Bartkowski, D.; Gale, D.; Monahan, J.; Niesman, M.R.; Alton, G.; et al. Effects of P38 MAPK inhibition on early stages of diabetic retinopathy and sensory nerve function. Investig. Ophthalmol. Vis. Sci. 2010, 51, 2158–2164. [Google Scholar] [CrossRef] [PubMed]
- Suryavanshi, S.V.; Kulkarni, Y.A. NF-Κβ: A potential target in the management of vascular complications of diabetes. Front. Pharmacol. 2017, 8, 798. [Google Scholar] [CrossRef] [PubMed]
- Ruff, C.A.; Staak, N.; Patodia, S.; Kaswich, M.; Rocha-Ferreira, E.; Da Costa, C.; Brecht, S.; Makwana, M.; Fontana, X.; Hristova, M.; et al. Neuronal c-Jun is required for successful axonal regeneration, but the effects of phosphorylation of its N-terminus are moderate. J. Neurochem. 2012, 121, 607–618. [Google Scholar] [CrossRef]
- Kato, N.; Matsumoto, M.; Kogawa, M.; Atkins, G.J.; Findlay, D.M.; Fujikawa, T.; Oda, H.; Ogata, M. Critical role of P38 MAPK for regeneration of the sciatic nerve following crush injury in vivo. J. Neuroinflammation 2013, 10, 757. [Google Scholar] [CrossRef]
- Sang, Q.; Sun, D.; Chen, Z.; Zhao, W. NGF and PI3K/Akt signaling participate in the ventral motor neuronal protection of curcumin in sciatic nerve injury rat models. Biomed. Pharmacother. 2018, 103, 1146–1153. [Google Scholar] [CrossRef]
- Holahan, M.R. A shift from a pivotal to supporting role for the growth-associated protein (GAP-43) in the coordination of axonal structural and functional plasticity. Front. Cell. Neurosci. 2017, 11, 266. [Google Scholar] [CrossRef] [PubMed]
- Benowitz, L.I.; Routtenberg, A. GAP-43: An intrinsic determinant of neuronal development and plasticity. Trends Neurosci. 1997, 20, 84–91. [Google Scholar] [CrossRef]
- Tang, X.; Chen, Y.; Gu, X.; Ding, F. Achyranthes bidentata Blume extract promotes neuronal growth in cultured embryonic rat hippocampal neurons. Prog. Nat. Sci. 2009, 19, 549–555. [Google Scholar] [CrossRef]
- Fregnan, F.; Muratori, L.; Simões, A.R.; Giacobini-Robecchi, M.G.; Raimondo, S. Role of inflammatory cytokines in peripheral nerve injury. Neural Regen. Res. 2012, 7, 2259–2266. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wei, G.H.; Huang, H.; Lan, Y.P.; Liu, B.; Liu, H.; Zhang, W.; Zuo, Y.X. Nerve injury-related autoimmunity activation leads to chronic inflammation and chronic neuropathic pain. Anesthesiology 2013, 118, 416–429. [Google Scholar] [CrossRef] [PubMed]
- Hobson, M.I.; Gree, C.J.; Terenghi, G. VEGF enhances intraneural angiogenesis and improves nerve regeneration after axotomy. J. Anat. 2000, 197, 591–605. [Google Scholar] [CrossRef]
- Fang, Z.; Ge, X.; Chen, X.; Xu, Y.; Yuan, W.E.; Ouyang, Y. Enhancement of sciatic nerve regeneration with dual delivery of vascular endothelial growth factor and nerve growth factor genes. J. Nanobiotechnol. 2020, 18, 46. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Shin, J.E.; Lee, B.; Kim, H.; Jeon, Y.; Ahn, S.H.; Chi, S.W.; Cho, Y. The stem cell marker Prom1 promotes axon regeneration by down-regulating cholesterol synthesis via Smad signaling. Proc. Natl. Acad. Sci. USA 2020, 117, 15955–15966. [Google Scholar] [CrossRef]
- Jankowski, M.P.; McIlwrath, S.L.; Jing, X.; Cornuet, P.K.; Salerno, K.M.; Koerber, H.R.; Albers, K.M. Sox11 Transcription factor modulates peripheral nerve regeneration in adult mice. Brain Res. 2009, 1256, 43–54. [Google Scholar] [CrossRef]
- Leibinger, M.; Müller, A.; Gobrecht, P.; Diekmann, H.; Andreadaki, A.; Fischer, D. Interleukin-6 contributes to CNS axon regeneration upon inflammatory stimulation. Cell Death Dis. 2013, 4, e609. [Google Scholar] [CrossRef] [PubMed]
- Cox, A.A.; Sagot, Y.; Hedou, G.; Grek, C.; Wilkes, T.; Vinik, A.I.; Ghatnekar, G. Low-dose pulsatile interleukin-6 as a treatment option for diabetic peripheral neuropathy. Front. Endocrinol. (Lausanne) 2017, 8, 89. [Google Scholar] [CrossRef]
- Snipes, G.J.; Suter, U.; Welcher, A.A.; Shooter, E.M. Characterization of a novel peripheral nervous system myelin protein (PMP- 22/SR13). J. Cell Biol. 1992, 117, 225–238. [Google Scholar] [CrossRef]
- Kuhn, G.; Lie, A.; Wilms, S.; Müller, H.W. Coexpression of PMP22 gene with MBP and P0 during de novo myelination and nerve repair. Glia 1993, 8, 256–264. [Google Scholar] [CrossRef]
- Martin, L.J.; Kaiser, A.; Price, A.C. Motor neuron degeneration after sciatic nerve avulsion in adult rat evolves with oxidative stress and is apoptosis. J. Neurobiol. 1999, 40, 185–201. [Google Scholar] [CrossRef]
- Liu, Y.; Wang, H. Peripheral nerve injury induced changes in the spinal cord and strategies to counteract/enhance the changes to promote nerve regeneration. Neural Regen. Res. 2020, 15, 189–198. [Google Scholar] [CrossRef]
- Amiri, S.; Movahedin, M.; Mowla, S.J.; Hajebrahimi, Z.; Tavallaei, M. Differential gene expression and alternative splicing of survivin following mouse sciatic nerve injury. Spinal Cord 2009, 47, 739–744. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Liu, Z.; Zhu, S.; Liu, L.; Ge, J.; Huang, L.; Sun, Z.; Zeng, W.; Huang, J.; Luo, Z. A magnetically responsive nanocomposite scaffold combined with Schwann cells promotes sciatic nerve regeneration upon exposure to magnetic field. Int. J. Nanomedicine 2017, 12, 7815–7832. [Google Scholar] [CrossRef] [PubMed]
- Lundborg, G.; Dahlin, L.B.; Danielsen, N.; Gelberman, R.H.; Longo, F.M.; Powell, H.C.; Varon, S. Nerve regeneration in silicone chambers: Influence of gap length and of distal stump components. Exp. Neurol. 1982, 76, 361–375. [Google Scholar] [CrossRef]
- Chang, Y.C.; Chen, M.H.; Liao, S.Y.; Wu, H.C.; Kuan, C.H.; Sun, J.S.; Wang, T.W. Multichanneled nerve guidance conduit with spatial gradients of neurotrophic factors and oriented nanotopography for repairing the peripheral nervous system. ACS Appl. Mater. Interfaces 2017, 9, 37623–37636. [Google Scholar] [CrossRef] [PubMed]
- Vijayavenkataraman, S. Nerve guide conduits for peripheral nerve injury repair: A review on design, materials and fabrication methods. Acta Biomater. 2020, 106, 54–69. [Google Scholar] [CrossRef]
- Fathi, S.S.; Zaminy, A. Stem cell therapy for nerve injury. World J. Stem Cells 2017, 9, 144–151. [Google Scholar] [CrossRef]
- Willand, M.P.; Nguyen, M.A.; Borschel, G.H.; Gordon, T. Electrical stimulation to promote peripheral nerve regeneration. Neurorehabil. Neural Repair 2016, 30, 490–496. [Google Scholar] [CrossRef]
- Turner, P.V.; Brabb, T.; Pekow, C.; Vasbinder, M.A. Administration of substances to laboratory animals: Routes of administration and factors to consider. J. Am. Assoc. Lab. Anim. Sci. 2011, 50, 600–613. [Google Scholar]
- Katiyar, C.; Kanjilal, S.; Gupta, A.; Katiyar, S. Drug discovery from plant sources: An integrated approach. AYU (An. Int. Q. J. Res. Ayurveda) 2012, 33, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Atanasov, A.G.; Waltenberger, B.; Pferschy-Wenzig, E.M.; Linder, T.; Wawrosch, C.; Uhrin, P.; Temml, V.; Wang, L.; Schwaiger, S.; Heiss, E.H.; et al. Discovery and resupply of pharmacologically active plant-derived natural products: A review. Biotechnol. Adv. 2015, 33, 1582–1614. [Google Scholar] [CrossRef]
- Singh, S.; Sharma, B.; Kanwar, S.S.; Kumar, A. Lead phytochemicals for anticancer drug development. Front. Plant Sci. 2016, 7, 1667. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.C.; Hsu, Y.J.; Chang, C.J.; Lin, C.S.; Martel, J.; Ojcius, D.M.; Ko, Y.F.; Lai, H.C.; Young, J.D. Immunomodulatory properties of medicinal mushrooms: Differential effects of water and ethanol extracts on NK cell-mediated cytotoxicity. Innate Immun. 2016, 22, 522–533. [Google Scholar] [CrossRef]
- Seow, S.L.-S.; Naidu, M.; David, P.; Wong, K.H.; Sabaratnam, V. Potentiation of neuritogenic activity of medicinal mushrooms in rat pheochromocytoma cells. BMC Complement. Altern. Med. 2013, 13, 157. [Google Scholar] [CrossRef]
- Wells, M.L.; Potin, P.; Craigie, J.S.; Raven, J.A.; Merchant, S.S.; Helliwell, K.E.; Smith, A.G.; Camire, M.E.; Brawley, S.H. Algae as nutritional and functional food sources: Revisiting our understanding. J. Appl. Phycol. 2017, 29, 949–982. [Google Scholar] [CrossRef]
- Khan, M.I.; Shin, J.H.; Kim, J.D. The promising future of microalgae: Current status, challenges, and optimization of a sustainable and renewable industry for biofuels, feed, and other products. Microb. Cell Fact. 2018, 17, 1–21. [Google Scholar] [CrossRef]
- Ariede, M.B.; Candido, T.M.; Jacome, A.L.M.; Velasco, M.V.R.; de Carvalho, J.C.M.; Baby, A.R. Cosmetic attributes of algae—A review. Algal Res. 2017, 25, 483–487. [Google Scholar] [CrossRef]
- Borowitzka, M.A. Microalgae as sources of pharmaceuticals and other biologically active compounds. J. Appl. Phycol. 1995, 7, 3–15. [Google Scholar] [CrossRef]
- Shannon, E.; Abu-Ghannam, N. Antibacterial derivatives of marine algae: An overview of pharmacological mechanisms and applications. Mar. Drugs 2016, 14, 81. [Google Scholar] [CrossRef]
- Barbosa, M.; Valentão, P.; Andrade, P.B. Bioactive compounds from macroalgae in the new millennium: Implications for neurodegenerative diseases. Mar. Drugs 2014, 12, 4934–4972. [Google Scholar] [CrossRef] [PubMed]
- Olasehinde, T.A.; Olaniran, A.O.; Okoh, A.I. Macroalgae as a valuable source of naturally occurring bioactive compounds for the treatment of Alzheimer’s disease. Mar. Drugs 2019, 17, 609. [Google Scholar] [CrossRef] [PubMed]
- Hannan, M.A.; Mohibbullah, M.; Hwang, S.Y.; Lee, K.; Kim, Y.C.; Hong, Y.K.; Moon, I.S. Differential neuritogenic activities of two edible brown macroalgae, Undaria pinnatifida and Saccharina japonica. Am. J. Chin. Med. 2014, 42, 1371–1384. [Google Scholar] [CrossRef] [PubMed]
- Mohibbullah, M.; Bhuiyan, M.M.H.; Hannan, M.A.; Getachew, P.; Hong, Y.K.; Choi, J.S.; Choi, I.S.; Moon, I.S. The edible red alga Porphyra yezoensis promotes neuronal survival and cytoarchitecture in primary hippocampal neurons. Cell. Mol. Neurobiol. 2016, 36, 669–682. [Google Scholar] [CrossRef] [PubMed]
- Tirtawijaya, G.; Mohibbullah, M.; Meinita, M.D.N.; Moon, I.S.; Hong, Y.K. The ethanol extract of the rhodophyte Kappaphycus alvarezii promotes neurite outgrowth in hippocampal neurons. J. Appl. Phycol. 2016, 28, 2515–2522. [Google Scholar] [CrossRef]
- Pang, J.R.; Goh, V.M.J.; Tan, C.Y.; Phang, S.M.; Wong, K.H.; Yow, Y.Y. Neuritogenic and in vitro antioxidant activities of Malaysian Gracilaria manilaensis Yamamoto & Trono. J. Appl. Phycol. 2018, 30, 3253–3260. [Google Scholar] [CrossRef]





| Sources | Natural Product | Chemical Structure |
|---|---|---|
| Alpinate Oxyphyllae Fructus (Alpinia oxyphylla Miq) | Protocatechuic acid |  |
| Astragalus membranaceus | Astragaloside IV |  |
| Crocus sativus | Crocin |  |
| Curcuma longa | Curcumin |  |
| Epimedium | Icariin |  |
| Gardenia jasminoides Ellis | Genipin |  |
| Gastrodia elata Blume | Gastrodin |  |
| Ginseng | Ginsenoside Rg1 |  |
| Ginsenoside Re |  | |
| Green tea | (-)-Epigallocatechin-3-gallate (EGCG) |  |
| Isolated from a variety of plants (e.g., grapes, Vitis vinifera; olive, Olea europaea; radish, Raphanus sativus; pumpkin, Cucurbita pepo [101]) | Syringic acid |  |
| Isolated from a variety of plants (e.g., apple, Malus domestica; cranberry, Vaccinium oxycoccus; peppermint, Mentha piperita; and thyme, Thymus vulgaris [102]) | Ursolic acid |  |
| Isolated from a variety of plants (e.g., apple, Malus domestica; caper, Capparis spinosa; onion, Allium cepa; tomato, Solanum lycopersicum; and grapes, Vitis vinifera [103]) | Quercetin |  |
| Pueraria lobata | Puerarin |  |
| Rhodiola rosea L. | Salidroside |  |
| Scutellaria baicalensis Georgi | Baicalin |  |
| Tripterygium wilfordii Hook.F. | Triptolide |  |
| Amanita muscaria | Muscimol |  |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yow, Y.-Y.; Goh, T.-K.; Nyiew, K.-Y.; Lim, L.-W.; Phang, S.-M.; Lim, S.-H.; Ratnayeke, S.; Wong, K.-H. Therapeutic Potential of Complementary and Alternative Medicines in Peripheral Nerve Regeneration: A Systematic Review. Cells 2021, 10, 2194. https://doi.org/10.3390/cells10092194
Yow Y-Y, Goh T-K, Nyiew K-Y, Lim L-W, Phang S-M, Lim S-H, Ratnayeke S, Wong K-H. Therapeutic Potential of Complementary and Alternative Medicines in Peripheral Nerve Regeneration: A Systematic Review. Cells. 2021; 10(9):2194. https://doi.org/10.3390/cells10092194
Chicago/Turabian StyleYow, Yoon-Yen, Tiong-Keat Goh, Ke-Ying Nyiew, Lee-Wei Lim, Siew-Moi Phang, Siew-Huah Lim, Shyamala Ratnayeke, and Kah-Hui Wong. 2021. "Therapeutic Potential of Complementary and Alternative Medicines in Peripheral Nerve Regeneration: A Systematic Review" Cells 10, no. 9: 2194. https://doi.org/10.3390/cells10092194
APA StyleYow, Y.-Y., Goh, T.-K., Nyiew, K.-Y., Lim, L.-W., Phang, S.-M., Lim, S.-H., Ratnayeke, S., & Wong, K.-H. (2021). Therapeutic Potential of Complementary and Alternative Medicines in Peripheral Nerve Regeneration: A Systematic Review. Cells, 10(9), 2194. https://doi.org/10.3390/cells10092194