
Telomere Shortening and Psychiatric Disorders: A Systematic Review

 , , , and
, , , and
Abstract
1. Introduction
2. Methodology
2.1. Outcome
2.2. Design
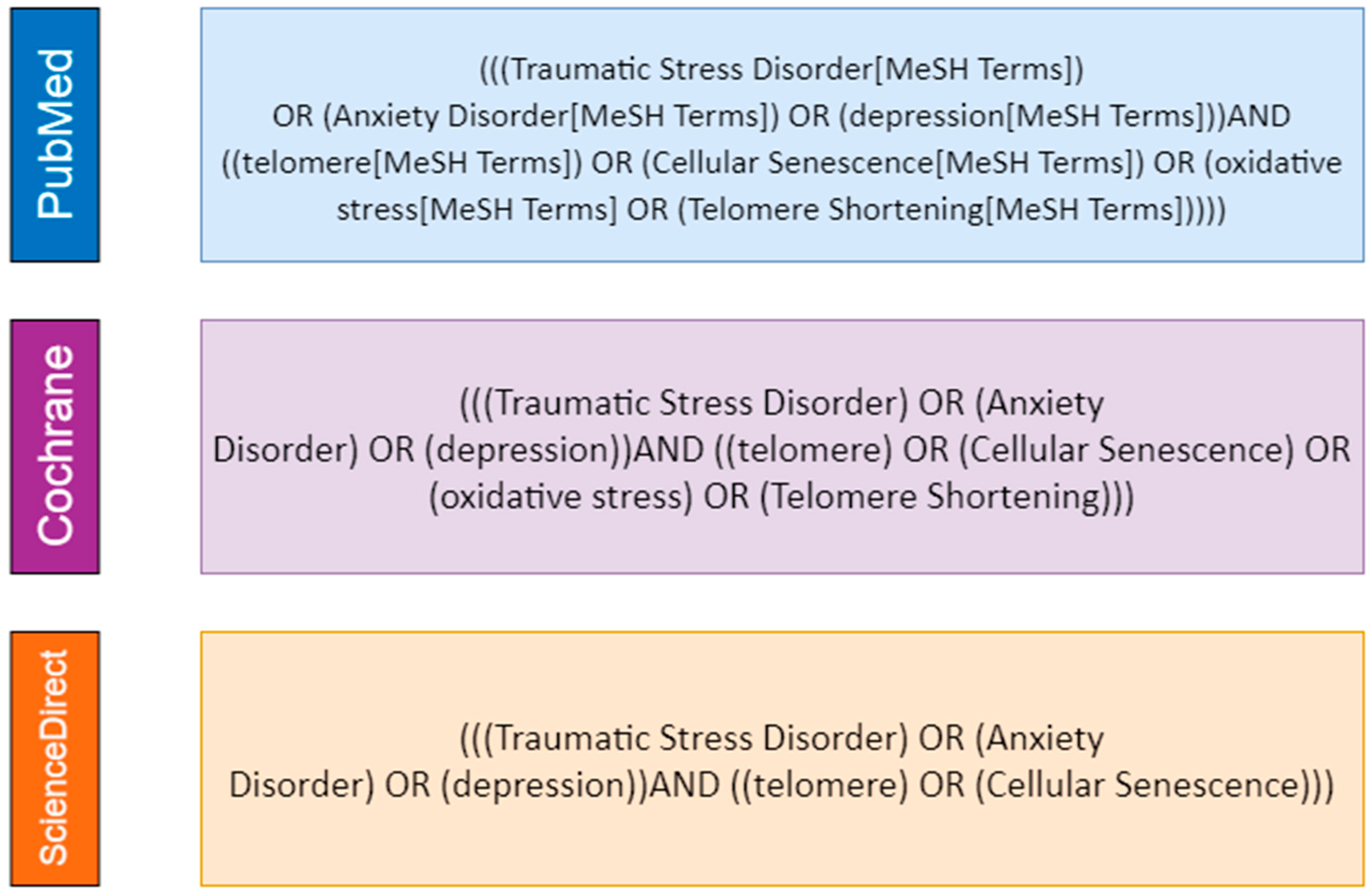
2.3. Search Strategy
2.4. Inclusion and Exclusion Criteria
2.5. Study Selection
2.6. Data Extraction
- Study identification: first author, publication date, country and type of study;
- Participants: age, gender and sample size;
- Variables: main comorbidities and lifestyle factors associated, race/ethnicity, psychiatric diseases (including severity and methods for diagnosis), medication and telomere measurement method.
2.7. Quality Assessment
3. Results
3.1. Telomere Length and Depressive Disorders
3.2. TL and PTSD
3.3. TL and Anxiety Disorders
3.4. Quality Assessment
4. Discussion
4.1. Risk Factors and Telomere Length
4.2. Psychological Distress-Related Diseases and TL
4.2.1. MDD and TL
4.2.2. PTSD and TL
4.2.3. Anxiety and TL
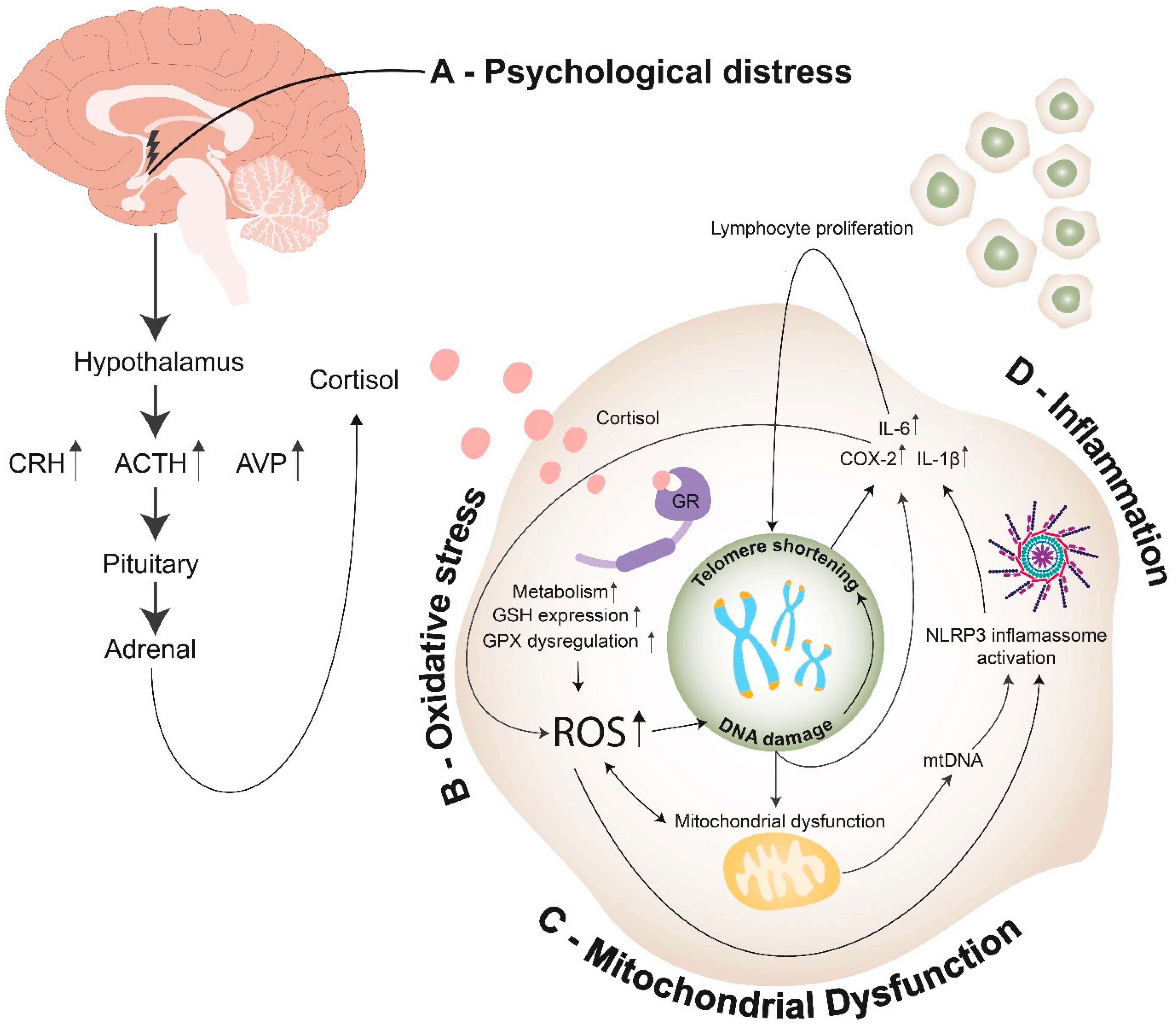
4.3. Mechanisms Underlying TL Shortening and Psychological Distress-Related Diseases
4.3.1. Oxidative Stress
4.3.2. Inflammation
4.3.3. Mitochondrial Dysfunction
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A. Newcastle–Ottawa Quality Assessment Scale Case-Control Studies
- (1)
- Is the case definition adequate?
- (a)
- yes, with independent validation

- (b)
- yes, e.g., record linkage or based on self-reports
- (c)
- no description
- (2)
- Representativeness of the cases
- (a)
- consecutive or obviously representative series of cases

- (b)
- potential for selection biases or not stated
- (3)
- Selection of Controls
- (a)
- community controls

- (b)
- hospital controls
- (c)
- no description
- (4)
- Definition of Controls
- (a)
- no history of disease (endpoint)

- (b)
- no description of source

- (1)
- Comparability of cases and controls on the basis of the design or analysis
- (a)
- study controls for age

- (b)
- study controls for sex, co-morbidities and lifestyle factors

- (1)
- Ascertainment of exposure
- (a)
- secure record (e.g., surgical records)

- (b)
- structured interview where blind to case/control status

- (c)
- interview not blinded to case/control status
- (d)
- written self-report or medical record only
- (e)
- no description
- (2)
- Same method of ascertainment for cases and controls
- (a)
- yes

- (b)
- no
- (3)
- Non-Response rate
- (a)
- same rate for both groups

- (b)
- non-respondents described
- (c)
- rate different and no designation
Appendix B. Newcastle–Ottawa Quality Assessment Scale Cohort Studies
- (1)
- Representativeness of the exposed cohort
- (a)
- truly representative of the average _______________ (describe) in the community

- (b)
- somewhat representative of the average ______________ in the community

- (c)
- selected group of users, e.g., nurses, volunteers
- (d)
- no description of the derivation of the cohort
- (2)
- Selection of the non-exposed cohort
- (a)
- drawn from the same community as the exposed cohort

- (b)
- drawn from a different source
- (c)
- no description of the derivation of the non-exposed cohort
- (3)
- Ascertainment of exposure
- (a)
- secure record (eg surgical records)

- (b)
- structured interview

- (c)
- written self-report
- (d)
- no description
- (4)
- Demonstration that the outcome of interest was not present at the start of the study
- (a)
- yes

- (b)
- no
- (1)
- Comparability of cohorts on the basis of design or analysis
- (a)
- study controls for age

- (b)
- study controls for sex, co-morbidities and lifestyle factors

- (1)
- Assessment of outcome
- (a)
- independent blind assessment

- (b)
- record linkage

- (c)
- self-report
- (d)
- no description
- (2)
- Was follow-up long enough for outcomes to occur
- (a)
- yes (adequate follow-up period: >5 years)

- (b)
- no
- (3)
- Adequacy of follow-up of cohorts
- (a)
- complete follow-up—all subjects accounted for

- (b)
- subjects lost to follow-up unlikely to introduce bias—small number lost—>85% follow-up, or description provided of those lost)

- (c)
- follow-up rate <85% and no description of those lost
- (d)
- no statement
Appendix C. Newcastle–Ottawa Quality Assessment Scale (Adapted for Cross-Sectional Studies)
- (1)
- Representativeness of the sample:
- (a)
- Truly representative of the average in the target population.
 (all subjects or random sampling)
(all subjects or random sampling) - (b)
- Somewhat representative of the average in the target population.
 (non-random sampling)
(non-random sampling) - (c)
- Selected group of users
- (d)
- No description of the sampling strategy
- (2)
- Sample size:
- (a)
- Justified and satisfactory

- (b)
- Not justified
- (3)
- Non-respondents:
- (a)
- Comparability between respondents and non-respondents characteristics is established, and the response rate is satisfactory

- (b)
- The response rate is unsatisfactory, or the comparability between respondents, and non-respondents is unsatisfactory
- (c)
- No description of the response rate or the characteristics of the responders and the non-responders
- (4)
- Ascertainment of the exposure (risk factor):
- (a)
- Validated measurement tool

- (b)
- No description of the measurement tool
- (1)
- The subjects in different outcome groups are comparable, based on the study design or analysis. Confounding factors are controlled
- (a)
- The study controls for the most important factor (select one)

- (b)
- The study controls for any additional factor

- (1)
- Assessment of the outcome:
- (a)
- Independent blind assessment

- (b)
- Record linkage

- (c)
- Self-report
- (d)
- No description
- (2)
- Statistical test:
- (a)
- The statistical test used to analyze the data is clearly described and appropriate, and the measurement of the association is presented, including both confidence intervals and the probability level (p-value)


- (b)
- The statistical test used to analyze the data is clearly described and appropriate, and the measurement of the association is presented, including either confidence intervals or the probability level (p-value)

- (c)
- The statistical test is not appropriate, not described or incomplete
References
- Witzany, G. The Viral Origins of Telomeres and Telomerases and Their Important Role in Eukaryogenesis and Genome Maintenance. Biosemiotics 2008, 1, 191–206. [Google Scholar] [CrossRef]
- Chai, W.; Du, Q.; Shay, J.W.; Wright, W.E. Human Telomeres Have Different Overhang Sizes at Leading versus Lagging Strands. Mol. Cell 2006, 21, 427–435. [Google Scholar] [CrossRef] [PubMed]
- Chai, W.; Shay, J.W.; Wright, W.E. Human Telomeres Maintain Their Overhang Length at Senescence. Mol. Cell Biol. 2005, 25, 2158–2168. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.K.; Mattoo, A.R.; Pandita, T.K. Telomeres and Telomerase☆. In Reference Module in Biomedical Sciences; Elsevier: Amsterdam, The Netherlands, 2015; p. B9780128012383988000. ISBN 9780128012383. [Google Scholar]
- Demanelis, K.; Jasmine, F.; Chen, L.S.; Chernoff, M.; Tong, L.; Delgado, D.; Zhang, C.; Shinkle, J.; Sabarinathan, M.; Lin, H.; et al. Determinants of Telomere Length across Human Tissues. Science 2020, 369. [Google Scholar] [CrossRef] [PubMed]
- Saretzki, G. Telomeres, Telomerase and Ageing. In Biochemistry and Cell Biology of Ageing: Part I Biomedical Science; Harris, J.R., Korolchuk, V.I., Eds.; Springer: Singapore, 2018; Volume 90, pp. 221–308. ISBN 9789811328343. [Google Scholar]
- Gobbini, E.; Trovesi, C.; Cassani, C.; Longhese, M.P. Telomere Uncapping at the Crossroad between Cell Cycle Arrest and Carcinogenesis. Mol. Cell. Oncol. 2014, 1, e29901. [Google Scholar] [CrossRef] [PubMed]
- Zee, R.Y.L.; Castonguay, A.J.; Barton, N.S.; Germer, S.; Martin, M. Mean Leukocyte Telomere Length Shortening and Type 2 Diabetes Mellitus: A Case-Control Study. Transl. Res. 2010, 155, 166–169. [Google Scholar] [CrossRef] [PubMed]
- Brouilette, S.W.; Moore, J.S.; McMahon, A.D.; Thompson, J.R.; Ford, I.; Shepherd, J.; Packard, C.J.; Samani, N.J. Telomere Length, Risk of Coronary Heart Disease, and Statin Treatment in the West of Scotland Primary Prevention Study: A Nested Case-Control Study. Lancet 2007, 369, 107–114. [Google Scholar] [CrossRef]
- Malouff, J.M.; Schutte, N.S. A Meta-Analysis of the Relationship between Anxiety and Telomere Length. Anxiety Stress Coping 2017, 30, 264–272. [Google Scholar] [CrossRef]
- Needham, B.L.; Mezuk, B.; Bareis, N.; Lin, J.; Blackburn, E.H.; Epel, E.S. Depression, Anxiety and Telomere Length in Young Adults: Evidence from the National Health and Nutrition Examination Survey. Mol. Psychiatry 2015, 20, 520–528. [Google Scholar] [CrossRef] [PubMed]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G.; Group, T.P. Preferred Reporting Items for Systematic Reviews and Meta-Analyses: The PRISMA Statement. PLoS Med. 2009, 6, e1000097. [Google Scholar] [CrossRef] [PubMed]
- Ouzzani, M.; Hammady, H.; Fedorowicz, Z.; Elmagarmid, A. Rayyan—a Web and Mobile App for Systematic Reviews. Syst. Rev. 2016, 5, 210. [Google Scholar] [CrossRef] [PubMed]
- Wells, G.A.; Shea, B.; O’Connell, D.; Peterson, J.; Welch, V.; Losos, M.; Tugwell, P. The Newcastle-Ottawa Scale (NOS) for Assessing the Quality of Nonrandomised Studies in Meta-Analyses. Available online: http://www.ohri.ca/programs/clinical_epidemiology/oxford.asp (accessed on 25 January 2021).
- Modesti, P.A.; Reboldi, G.; Cappuccio, F.P.; Agyemang, C.; Remuzzi, G.; Rapi, S.; Perruolo, E.; Parati, G.; ESH Working Group on CV Risk in Low Resource Settings. Panethnic Differences in Blood Pressure in Europe: A Systematic Review and Meta-Analysis. PLoS ONE 2016, 11, e0147601. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Rizo, C.; Fernandez-Egea, E.; Miller, B.J.; Oliveira, C.; Justicia, A.; Griffith, J.K.; Heaphy, C.M.; Bernardo, M.; Kirkpatrick, B. Abnormal Glucose Tolerance, White Blood Cell Count, and Telomere Length in Newly Diagnosed, Antidepressant-Naïve Patients with Depression. Brain Behav. Immun. 2013, 28, 49–53. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.H.; Epel, E.S.; Mellon, S.H.; Lin, J.; Reus, V.I.; Rosser, R.; Kupferman, E.; Burke, H.; Mahan, L.; Blackburn, E.H.; et al. Adverse Childhood Experiences and Leukocyte Telomere Maintenance in Depressed and Healthy Adults. J. Affect. Disord. 2014, 169, 86–90. [Google Scholar] [CrossRef] [PubMed]
- Tyrka, A.R.; Parade, S.H.; Price, L.H.; Kao, H.-T.; Porton, B.; Philip, N.S.; Welch, E.S.; Carpenter, L.L. Alterations of Mitochondrial DNA Copy Number and Telomere Length with Early Adversity and Psychopathology. Biol. Psychiatry 2016, 79, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Prabu, P.; Poongothai, S.; Shanthirani, C.S.; Anjana, R.M.; Mohan, V.; Balasubramanyam, M. Altered Circulatory Levels of MiR-128, BDNF, Cortisol and Shortened Telomeres in Patients with Type 2 Diabetes and Depression. Acta Diabetol. 2020, 57, 799–807. [Google Scholar] [CrossRef] [PubMed]
- Vincent, J.; Hovatta, I.; Frissa, S.; Goodwin, L.; Hotopf, M.; Hatch, S.L.; Breen, G.; Powell, T.R. Assessing the Contributions of Childhood Maltreatment Subtypes and Depression Case-Control Status on Telomere Length Reveals a Specific Role of Physical Neglect. J. Affect. Disord. 2017, 213, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Hoen, P.W.; Rosmalen, J.G.M.; Schoevers, R.A.; Huzen, J.; van der Harst, P.; de Jonge, P. Association between Anxiety but Not Depressive Disorders and Leukocyte Telomere Length after 2 Years of Follow-up in a Population-Based Sample. Psychol. Med. 2013, 43, 689–697. [Google Scholar] [CrossRef]
- Bersani, F.S.; Lindqvist, D.; Mellon, S.H.; Epel, E.S.; Yehuda, R.; Flory, J.; Henn-Hasse, C.; Bierer, L.M.; Makotkine, I.; Abu-Amara, D.; et al. Association of Dimensional Psychological Health Measures with Telomere Length in Male War Veterans. J. Affect. Disord. 2016, 190, 537–542. [Google Scholar] [CrossRef] [PubMed]
- Hoen, P.W.; de Jonge, P.; Na, B.Y.; Farzaneh-Far, R.; Epel, E.; Lin, J.; Blackburn, E.; Whooley, M.A. Depression and Leukocyte Telomere Length in Patients with Coronary Heart Disease: Data from The Heart and Soul Study. Psychosom. Med. 2011, 73, 541–547. [Google Scholar] [CrossRef]
- AlAhwal, M.S.; Zaben, F.A.; Sehlo, M.G.; Khalifa, D.A.; Al-Aama, J.Y.; Edris, S.; Ashy, J.A.; Koenig, H.G. Depression and Telomere Length in Colorectal Cancer Patients in Saudi Arabia. Asian J. Psychiatry 2019, 40, 130–131. [Google Scholar] [CrossRef] [PubMed]
- Verhoeven, J.E.; Révész, D.; Picard, M.; Epel, E.E.; Wolkowitz, O.M.; Matthews, K.A.; Penninx, B.W.J.H.; Puterman, E. Depression, Telomeres and Mitochondrial DNA: Between- and within-Person Associations from a 10-Year Longitudinal Study. Mol. Psychiatry 2018, 23, 850–857. [Google Scholar] [CrossRef] [PubMed]
- Verhoeven, J.E.; van Oppen, P.; Révész, D.; Wolkowitz, O.M.; Penninx, B.W.J.H. Depressive and Anxiety Disorders Showing Robust, but Non-Dynamic, 6-Year Longitudinal Association With Short Leukocyte Telomere Length. AJP 2016, 173, 617–624. [Google Scholar] [CrossRef] [PubMed]
- Whisman, M.A.; Richardson, E.D. Depressive Symptoms and Salivary Telomere Length in a Probability Sample of Middle-Aged and Older Adults. Psychosom. Med. 2017, 79, 234–242. [Google Scholar] [CrossRef][Green Version]
- Lin, J.; Blalock, J.A.; Chen, M.; Ye, Y.; Gu, J.; Cohen, L.; Cinciripini, P.M.; Wu, X. Depressive Symptoms and Short Telomere Length Are Associated with Increased Mortality in Bladder Cancer Patients. Cancer Epidemiol. Biomark. Prev. 2015, 24, 336–343. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Zhu, Y.; Yeh, F.; Lin, J.; Lee, E.T.; Cole, S.A.; Calhoun, D.; Zhao, J. Depressive Symptoms Are Associated with Leukocyte Telomere Length in American Indians: Findings from the Strong Heart Family Study. Aging 2016, 8, 2961–2970. [Google Scholar] [CrossRef]
- Shaffer, J.A.; Epel, E.; Kang, M.S.; Ye, S.; Schwartz, J.E.; Davidson, K.W.; Kirkland, S.; Honig, L.S.; Shimbo, D. Depressive Symptoms Are Not Associated with Leukocyte Telomere Length: Findings from the Nova Scotia Health Survey (NSHS95), a Population-Based Study. PLoS ONE 2012, 7, e48318. [Google Scholar] [CrossRef] [PubMed]
- Chae, D.H.; Epel, E.S.; Nuru-Jeter, A.M.; Lincoln, K.D.; Taylor, R.J.; Lin, J.; Blackburn, E.H.; Thomas, S.B. Discrimination, Mental Health, and Leukocyte Telomere Length among African American Men. Psychoneuroendocrinology 2016, 63, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Phillips, A.C.; Robertson, T.; Carroll, D.; Der, G.; Shiels, P.G.; McGlynn, L.; Benzeval, M. Do Symptoms of Depression Predict Telomere Length? Evidence from the West of Scotland Twenty-07 Study. Psychosom. Med. 2013, 75, 288–296. [Google Scholar] [CrossRef]
- Liu, Z.; Zhang, J.; Yan, J.; Wang, Y.; Li, Y. Leucocyte Telomere Shortening in Relation to Newly Diagnosed Type 2 Diabetic Patients with Depression. Oxid. Med. Cell Longev. 2014, 2014, 673959. [Google Scholar] [CrossRef]
- Wang, X.; Sundquist, K.; Hedelius, A.; Palmér, K.; Memon, A.A.; Sundquist, J. Leukocyte Telomere Length and Depression, Anxiety and Stress and Adjustment Disorders in Primary Health Care Patients. BMC Psychiatry 2017, 17, 148. [Google Scholar] [CrossRef]
- Schaakxs, R.; Verhoeven, J.E.; Oude Voshaar, R.C.; Comijs, H.C.; Penninx, B.W.J.H. Leukocyte Telomere Length and Late-Life Depression. Am. J. Geriatr. Psychiatry 2015, 23, 423–432. [Google Scholar] [CrossRef] [PubMed]
- Wolkowitz, O.M.; Mellon, S.H.; Epel, E.S.; Lin, J.; Dhabhar, F.S.; Su, Y.; Reus, V.I.; Rosser, R.; Burke, H.M.; Kupferman, E.; et al. Leukocyte Telomere Length in Major Depression: Correlations with Chronicity, Inflammation and Oxidative Stress—Preliminary Findings. PLoS ONE 2011, 6, e17837. [Google Scholar] [CrossRef]
- Georgin-Lavialle, S.; Moura, D.S.; Bruneau, J.; Chauvet-Gélinier, J.-C.; Damaj, G.; Soucie, E.; Barete, S.; Gacon, A.-L.; Grandpeix-Guyodo, C.; Suarez, F.; et al. Leukocyte Telomere Length in Mastocytosis: Correlations with Depression and Perceived Stress. Brain Behav. Immun. 2014, 35, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Verhoeven, J.E.; Révész, D.; Epel, E.S.; Lin, J.; Wolkowitz, O.M.; Penninx, B.W.J.H. Major Depressive Disorder and Accelerated Cellular Aging: Results from a Large Psychiatric Cohort Study. Mol. Psychiatry 2014, 19, 895–901. [Google Scholar] [CrossRef] [PubMed]
- Puterman, E.; Epel, E.S.; Lin, J.; Blackburn, E.H.; Gross, J.J.; Whooley, M.A.; Cohen, B.E. Multisystem Resiliency Moderates the Major Depression–Telomere Length Association: Findings from the Heart and Soul Study. Brain Behav. Immun. 2013, 33, 65–73. [Google Scholar] [CrossRef]
- Henje Blom, E.; Han, L.K.M.; Connolly, C.G.; Ho, T.C.; Lin, J.; LeWinn, K.Z.; Simmons, A.N.; Sacchet, M.D.; Mobayed, N.; Luna, M.E.; et al. Peripheral Telomere Length and Hippocampal Volume in Adolescents with Major Depressive Disorder. Transl. Psychiatry 2015, 5, e676. [Google Scholar] [CrossRef] [PubMed]
- Vance, M.C.; Bui, E.; Hoeppner, S.S.; Kovachy, B.; Prescott, J.; Mischoulon, D.; Walton, Z.E.; Dong, M.; Nadal, M.F.; Worthington, J.J.; et al. Prospective Association between Major Depressive Disorder and Leukocyte Telomere Length over Two Years. Psychoneuroendocrinology 2018, 90, 157–164. [Google Scholar] [CrossRef]
- Starnino, L.; Busque, L.; Tardif, J.-C.; D’Antono, B. Psychological Profiles in the Prediction of Leukocyte Telomere Length in Healthy Individuals. PLoS ONE 2016, 11, e0165482. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Koenig, H.G.; Al Shohaib, S.; Wang, Z. Religiosity, Depression and Telomere Length in Chinese Older Adults. J. Affect. Disord. 2020, 260, 624–628. [Google Scholar] [CrossRef] [PubMed]
- Wikgren, M.; Maripuu, M.; Karlsson, T.; Nordfjäll, K.; Bergdahl, J.; Hultdin, J.; Del-Favero, J.; Roos, G.; Nilsson, L.-G.; Adolfsson, R.; et al. Short Telomeres in Depression and the General Population Are Associated with a Hypocortisolemic State. Biol. Psychiatry 2012, 71, 294–300. [Google Scholar] [CrossRef] [PubMed]
- Szebeni, A.; Szebeni, K.; DiPeri, T.; Chandley, M.J.; Crawford, J.D.; Stockmeier, C.A.; Ordway, G.A. Shortened Telomere Length in White Matter Oligodendrocytes in Major Depression: Potential Role of Oxidative Stress. Int. J. Neuropsychopharmacol. 2014, 17, 1579–1589. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.J.; Wei, Y.B.; Forsell, Y.; Lavebratt, C. Stress, Depressive Status and Telomere Length: Does Social Interaction and Coping Strategy Play a Mediating Role? J. Affect. Disord. 2017, 222, 138–145. [Google Scholar] [CrossRef]
- Boeck, C.; Salinas-Manrique, J.; Calzia, E.; Radermacher, P.; von Arnim, C.A.F.; Dietrich, D.E.; Kolassa, I.-T.; Karabatsiakis, A. Targeting the Association between Telomere Length and Immuno-Cellular Bioenergetics in Female Patients with Major Depressive Disorder. Sci. Rep. 2018, 8, 9419. [Google Scholar] [CrossRef]
- Jiménez, K.M.; Pereira-Morales, A.J.; Adan, A.; Forero, D.A.; Jiménez, K.M.; Pereira-Morales, A.J.; Adan, A.; Forero, D.A. Telomere Length and Childhood Trauma in Colombians with Depressive Symptoms. Braz. J. Psychiatry 2019, 41, 194–198. [Google Scholar] [CrossRef]
- Wium-Andersen, M.K.; Ørsted, D.D.; Rode, L.; Bojesen, S.E.; Nordestgaard, B.G. Telomere Length and Depression: Prospective Cohort Study and Mendelian Randomisation Study in 67 306 Individuals. Br. J. Psychiatry 2017, 210, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Huzen, J.; van der Harst, P.; de Boer, R.A.; Lesman-Leegte, I.; Voors, A.A.; van Gilst, W.H.; Samani, N.J.; Jaarsma, T.; van Veldhuisen, D.J. Telomere Length and Psychological Well-Being in Patients with Chronic Heart Failure. Age Ageing 2010, 39, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Simon, N.M.; Walton, Z.E.; Bui, E.; Prescott, J.; Hoge, E.; Keshaviah, A.; Schwarz, N.; Dryman, T.; Ojserkis, R.A.; Kovachy, B.; et al. Telomere Length and Telomerase in a Well-Characterized Sample of Individuals with Major Depressive Disorder Compared to Controls. Psychoneuroendocrinology 2015, 58, 9–22. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, N.; Boehner, M.; Groenen, F.; Kalb, R. Telomere Length of Patients with Major Depression Is Shortened but Independent from Therapy and Severity of the Disease. Depress. Anxiety 2010, 27, 1111–1116. [Google Scholar] [CrossRef] [PubMed]
- Karabatsiakis, A.; Kolassa, I.-T.; Kolassa, S.; Rudolph, K.L.; Dietrich, D.E. Telomere Shortening in Leukocyte Subpopulations in Depression. BMC Psychiatry 2014, 14, 192. [Google Scholar] [CrossRef] [PubMed]
- Solomon, Z.; Tsur, N.; Levin, Y.; Uziel, O.; Lahav, M.; Ohry, A. The Implications of War Captivity and Long-Term Psychopathology Trajectories for Telomere Length. Psychoneuroendocrinology 2017, 81, 122–128. [Google Scholar] [CrossRef]
- Verhoeven, J.E.; Penninx, B.W.J.H.; Milaneschi, Y. Unraveling the Association between Depression and Telomere Length Using Genomics. Psychoneuroendocrinology 2019, 102, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Mamdani, F.; Rollins, B.; Morgan, L.; Myers, R.M.; Barchas, J.D.; Schatzberg, A.F.; Watson, S.J.; Akil, H.; Potkin, S.G.; Bunney, W.E.; et al. Variable Telomere Length across Post-Mortem Human Brain Regions and Specific Reduction in the Hippocampus of Major Depressive Disorder. Transl. Psychiatry 2015, 5, e636. [Google Scholar] [CrossRef]
- Zhang, L.; Hu, X.-Z.; Russell, D.W.; Benedek, D.M.; Fullerton, C.S.; Naifeh, J.A.; Li, X.; Chen, Z.; Wu, H.; Ng, T.H.H.; et al. Association between Leukocyte Telomere Length and Hostility in US Army Service Members. Neurosci. Lett. 2019, 706, 24–29. [Google Scholar] [CrossRef] [PubMed]
- O’Donovan, A.; Epel, E.; Lin, J.; Wolkowitz, O.; Cohen, B.; Maguen, S.; Metzler, T.; Lenoci, M.; Blackburn, E.; Neylan, T.C. Childhood Trauma Associated with Short Leukocyte Telomere Length in Post-Traumatic Stress Disorder. Biol. Psychiatry 2011, 70, 465–471. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.I.; Mueller, S.G.; Wu, G.W.Y.; Lin, J.; Ng, P.; Yehuda, R.; Flory, J.D.; Abu-Amara, D.; Reus, V.I.; Gautam, A.; et al. Effect of Combat Exposure and Posttraumatic Stress Disorder on Telomere Length and Amygdala Volume. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2020, 5, 678–687. [Google Scholar] [CrossRef]
- Malan, S.; Hemmings, S.; Kidd, M.; Martin, L.; Seedat, S. Investigation of Telomere Length and Psychological Stress in Rape Victims. Depress. Anxiety 2011, 28, 1081–1085. [Google Scholar] [CrossRef] [PubMed]
- Boks, M.P.; van Mierlo, H.C.; Rutten, B.P.F.; Radstake, T.R.D.J.; De Witte, L.; Geuze, E.; Horvath, S.; Schalkwyk, L.C.; Vinkers, C.H.; Broen, J.C.A.; et al. Longitudinal Changes of Telomere Length and Epigenetic Age Related to Traumatic Stress and Post-Traumatic Stress Disorder. Psychoneuroendocrinology 2015, 51, 506–512. [Google Scholar] [CrossRef]
- Roberts, A.L.; Koenen, K.C.; Chen, Q.; Gilsanz, P.; Mason, S.M.; Prescott, J.; Ratanatharathorn, A.; Rimm, E.B.; Sumner, J.A.; Winning, A.; et al. Posttraumatic Stress Disorder and Accelerated Aging: PTSD and Leukocyte Telomere Length in a Sample of Civilian Women. Depress. Anxiety 2017, 34, 391–400. [Google Scholar] [CrossRef] [PubMed]
- Ladwig, K.-H.; Brockhaus, A.C.; Baumert, J.; Lukaschek, K.; Emeny, R.T.; Kruse, J.; Codd, V.; Häfner, S.; Albrecht, E.; Illig, T.; et al. Posttraumatic Stress Disorder and Not Depression Is Associated with Shorter Leukocyte Telomere Length: Findings from 3000 Participants in the Population-Based KORA F4 Study. PLoS ONE 2013, 8, e64762. [Google Scholar] [CrossRef]
- Jergović, M.; Tomičević, M.; Vidović, A.; Bendelja, K.; Savić, A.; Vojvoda, V.; Rac, D.; Lovrić-Čavar, D.; Rabatić, S.; Jovanovic, T.; et al. Telomere Shortening and Immune Activity in War Veterans with Posttraumatic Stress Disorder. Prog. Neuropsychopharmacol. Biol. Psychiatry 2014, 54, 275–283. [Google Scholar] [CrossRef] [PubMed]
- Avetyan, D.; Zakharyan, R.; Petrek, M.; Arakelyan, A. Telomere Shortening in Blood Leukocytes of Patients with Posttraumatic Stress Disorder. J. Psychiatr. Res. 2019, 111, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.Y.; Kim, S.J.; Choi, J.R.; Lee, S.-T.; Kim, J.; Hwang, I.S.; Chung, H.G.; Choi, J.H.; Kim, H.W.; Kim, S.H.; et al. The Effect of Trauma and PTSD on Telomere Length: An Exploratory Study in People Exposed to Combat Trauma. Sci. Rep. 2017, 7, 4375. [Google Scholar] [CrossRef]
- Zhang, L.; Hu, X.-Z.; Benedek, D.M.; Fullerton, C.S.; Forsten, R.D.; Naifeh, J.A.; Li, X.; Li, H.; Benevides, K.N.; Smerin, S.; et al. The Interaction between Stressful Life Events and Leukocyte Telomere Length Is Associated with PTSD. Mol. Psychiatry 2014, 19, 856–857. [Google Scholar] [CrossRef]
- Verhoeven, J.E.; Révész, D.; van Oppen, P.; Epel, E.S.; Wolkowitz, O.M.; Penninx, B.W.J.H. Anxiety Disorders and Accelerated Cellular Ageing. Br. J. Psychiatry 2015, 206, 371–378. [Google Scholar] [CrossRef] [PubMed]
- Groer, M.; Louis-Jacques, A.; Szalacha, L.; Redwine, L.; Dracxler, R.; Keefe, D. Relationship of Anxiety, Inflammation, and Telomere Length in Postpartum Women: A Pilot Study. Biol. Res. Nurs. 2020, 22, 256–262. [Google Scholar] [CrossRef] [PubMed]
- Prelog, M.; Hilligardt, D.; Schmidt, C.A.; Przybylski, G.K.; Leierer, J.; Almanzar, G.; El Hajj, N.; Lesch, K.-P.; Arolt, V.; Zwanzger, P.; et al. Hypermethylation of FOXP3 Promoter and Premature Aging of the Immune System in Female Patients with Panic Disorder? PLoS ONE 2016, 11, e0157930. [Google Scholar] [CrossRef]
- Shalev, I.; Entringer, S.; Wadhwa, P.D.; Wolkowitz, O.M.; Puterman, E.; Lin, J.; Epel, E.S. Stress and Telomere Biology: A Lifespan Perspective. Psychoneuroendocrinology 2013, 38, 1835–1842. [Google Scholar] [CrossRef] [PubMed]
- Martens, D.S.; Janssen, B.G.; Bijnens, E.M.; Clemente, D.B.P.; Vineis, P.; Plusquin, M.; Nawrot, T.S. Association of Parental Socioeconomic Status and Newborn Telomere Length. JAMA Netw. Open 2020, 3, e204057. [Google Scholar] [CrossRef] [PubMed]
- Welendorf, C.; Nicoletti, C.F.; Pinhel, M.A.D.S.; Noronha, N.Y.; de Paula, B.M.F.; Nonino, C.B. Obesity, Weight Loss, and Influence on Telomere Length: New Insights for Personalized Nutrition. Nutrition 2019, 66, 115–121. [Google Scholar] [CrossRef]
- Astuti, Y.; Wardhana, A.; Watkins, J.; Wulaningsih, W.; PILAR Research Network. Cigarette Smoking and Telomere Length: A Systematic Review of 84 Studies and Meta-Analysis. Environ. Res. 2017, 158, 480–489. [Google Scholar] [CrossRef] [PubMed]
- Entringer, S.; Epel, E.S.; Kumsta, R.; Lin, J.; Hellhammer, D.H.; Blackburn, E.H.; Wüst, S.; Wadhwa, P.D. Stress Exposure in Intrauterine Life Is Associated with Shorter Telomere Length in Young Adulthood. Proc. Natl. Acad. Sci. USA 2011, 108, E513–E518. [Google Scholar] [CrossRef] [PubMed]
- Price, L.H.; Kao, H.-T.; Burgers, D.E.; Carpenter, L.L.; Tyrka, A.R. Telomeres and Early-Life Stress: An Overview. Biol. Psychiatry 2013, 73, 15–23. [Google Scholar] [CrossRef]
- Shalev, I.; Moffitt, T.E.; Sugden, K.; Williams, B.; Houts, R.M.; Danese, A.; Mill, J.; Arseneault, L.; Caspi, A. Exposure to Violence during Childhood Is Associated with Telomere Erosion from 5 to 10 Years of Age: A Longitudinal Study. Mol. Psychiatry 2013, 18, 576–581. [Google Scholar] [CrossRef] [PubMed]
- Ornish, D.; Lin, J.; Chan, J.M.; Epel, E.; Kemp, C.; Weidner, G.; Marlin, R.; Frenda, S.J.; Magbanua, M.J.M.; Daubenmier, J.; et al. Effect of Comprehensive Lifestyle Changes on Telomerase Activity and Telomere Length in Men with Biopsy-Proven Low-Risk Prostate Cancer: 5-Year Follow-up of a Descriptive Pilot Study. Lancet Oncol. 2013, 14, 1112–1120. [Google Scholar] [CrossRef]
- Wolkowitz, O.M.; Reus, V.I.; Mellon, S.H. Of Sound Mind and Body: Depression, Disease, and Accelerated Aging. Dialogues Clin. Neurosci. 2011, 13, 25–39. [Google Scholar]
- Paul, L. Diet, Nutrition and Telomere Length. J. Nutr. Biochem. 2011, 22, 895–901. [Google Scholar] [CrossRef]
- Liang, G.; Schernhammer, E.; Qi, L.; Gao, X.; De Vivo, I.; Han, J. Associations between Rotating Night Shifts, Sleep Duration, and Telomere Length in Women. PLoS ONE 2011, 6, e23462. [Google Scholar] [CrossRef]
- Jacobs, T.L.; Epel, E.S.; Lin, J.; Blackburn, E.H.; Wolkowitz, O.M.; Bridwell, D.A.; Zanesco, A.P.; Aichele, S.R.; Sahdra, B.K.; MacLean, K.A.; et al. Intensive Meditation Training, Immune Cell Telomerase Activity, and Psychological Mediators. Psychoneuroendocrinology 2011, 36, 664–681. [Google Scholar] [CrossRef] [PubMed]
- Pavanello, S.; Hoxha, M.; Dioni, L.; Bertazzi, P.A.; Snenghi, R.; Nalesso, A.; Ferrara, S.D.; Montisci, M.; Baccarelli, A. Shortened Telomeres in Individuals with Abuse in Alcohol Consumption. Int. J. Cancer 2011, 129, 983–992. [Google Scholar] [CrossRef]
- Valdes, A.M.; Andrew, T.; Gardner, J.P.; Kimura, M.; Oelsner, E.; Cherkas, L.F.; Aviv, A.; Spector, T.D. Obesity, Cigarette Smoking, and Telomere Length in Women. Lancet 2005, 366, 662–664. [Google Scholar] [CrossRef]
- Diaz, V.A.; Mainous, A.G.; Everett, C.J.; Schoepf, U.J.; Codd, V.; Samani, N.J.; Samanii, N.J. Effect of Healthy Lifestyle Behaviors on the Association between Leukocyte Telomere Length and Coronary Artery Calcium. Am. J. Cardiol. 2010, 106, 659–663. [Google Scholar] [CrossRef] [PubMed]
- Danese, A.; Pariante, C.M.; Caspi, A.; Taylor, A.; Poulton, R. Childhood Maltreatment Predicts Adult Inflammation in a Life-Course Study. Proc. Natl. Acad. Sci. USA 2007, 104, 1319–1324. [Google Scholar] [CrossRef]
- von Zglinicki, T. Oxidative Stress Shortens Telomeres. Trends Biochem. Sci. 2002, 27, 339–344. [Google Scholar] [CrossRef]
- Behl, C.; Lezoualc’h, F.; Trapp, T.; Widmann, M.; Skutella, T.; Holsboer, F. Glucocorticoids Enhance Oxidative Stress-Induced Cell Death in Hippocampal Neurons in Vitro. Endocrinology 1997, 138, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Epel, E.S.; Blackburn, E.H.; Lin, J.; Dhabhar, F.S.; Adler, N.E.; Morrow, J.D.; Cawthon, R.M. Accelerated Telomere Shortening in Response to Life Stress. Proc. Natl. Acad. Sci. USA 2004, 101, 17312–17315. [Google Scholar] [CrossRef] [PubMed]
- Darrow, S.M.; Verhoeven, J.E.; Révész, D.; Lindqvist, D.; Penninx, B.W.J.H.; Delucchi, K.L.; Wolkowitz, O.M.; Mathews, C.A. The Association Between Psychiatric Disorders and Telomere Length: A Meta-Analysis Involving 14,827 Persons. Psychosom. Med. 2016, 78, 776–787. [Google Scholar] [CrossRef]
- Lin, P.-Y.; Huang, Y.-C.; Hung, C.-F. Shortened Telomere Length in Patients with Depression: A Meta-Analytic Study. J. Psychiatr. Res. 2016, 76, 84–93. [Google Scholar] [CrossRef] [PubMed]
- Ridout, K.K.; Ridout, S.J.; Price, L.H.; Sen, S.; Tyrka, A.R. Depression and Telomere Length: A Meta-Analysis. J. Affect. Disord. 2016, 191, 237–247. [Google Scholar] [CrossRef]
- Schutte, N.S.; Malouff, J.M. The Association between Depression and Leukocyte Telomere Length: A Meta-Analysis: Depression and Leukocyte Telomere Length. Depress. Anxiety 2015, 32, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, J.; Zhou, J.; Huang, P.; Li, J. The Association between Post-Traumatic Stress Disorder and Shorter Telomere Length: A Systematic Review and Meta-Analysis. J. Affect. Disord. 2017, 218, 322–326. [Google Scholar] [CrossRef] [PubMed]
- Puterman, E.; Epel, E. An Intricate Dance: Life Experience, Multisystem Resiliency, and Rate of Telomere Decline Throughout the Lifespan. Soc. Personal. Psychol. Compass 2012, 6, 807–825. [Google Scholar] [CrossRef] [PubMed]
- Depression. Available online: https://www.who.int/westernpacific/health-topics/depression (accessed on 15 January 2021).
- Depression and Other Common Mental Disorders. Available online: https://www.who.int/publications-detail-redirect/depression-global-health-estimates (accessed on 15 January 2021).
- Clark, L.A.; Cuthbert, B.; Lewis-Fernández, R.; Narrow, W.E.; Reed, G.M. Three Approaches to Understanding and Classifying Mental Disorder: ICD-11, DSM-5, and the National Institute of Mental Health’s Research Domain Criteria (RDoC). Psychol. Sci. Public Interest 2017, 18, 72–145. [Google Scholar] [CrossRef]
- Hammen, C. Risk Factors for Depression: An Autobiographical Review. Annu. Rev. Clin. Psychol. 2018, 14, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Czarny, P.; Wigner, P.; Galecki, P.; Sliwinski, T. The Interplay between Inflammation, Oxidative Stress, DNA Damage, DNA Repair and Mitochondrial Dysfunction in Depression. Progress Neuro Psychopharmacol. Biol. Psychiatry 2018, 80, 309–321. [Google Scholar] [CrossRef] [PubMed]
- Dean, J.; Keshavan, M. The Neurobiology of Depression: An Integrated View. Asian J. Psychiatry 2017, 27, 101–111. [Google Scholar] [CrossRef]
- Penninx, B.W.; Milaneschi, Y.; Lamers, F.; Vogelzangs, N. Understanding the Somatic Consequences of Depression: Biological Mechanisms and the Role of Depression Symptom Profile. BMC Med. 2013, 11, 129. [Google Scholar] [CrossRef] [PubMed]
- Fasching, C.L. Telomere Length Measurement as a Clinical Biomarker of Aging and Disease. Crit. Rev. Clin. Lab. Sci. 2018, 55, 443–465. [Google Scholar] [CrossRef] [PubMed]
- Yehuda, R.; Hoge, C.W.; McFarlane, A.C.; Vermetten, E.; Lanius, R.A.; Nievergelt, C.M.; Hobfoll, S.E.; Koenen, K.C.; Neylan, T.C.; Hyman, S.E. Post-Traumatic Stress Disorder. Nat. Rev. Dis. Primers 2015, 1, 1–22. [Google Scholar] [CrossRef] [PubMed]
- van Zuiden, M.; Geuze, E.; Willemen, H.L.D.M.; Vermetten, E.; Maas, M.; Heijnen, C.J.; Kavelaars, A. Pre-Existing High Glucocorticoid Receptor Number Predicting Development of Posttraumatic Stress Symptoms After Military Deployment. AJP 2011, 168, 89–96. [Google Scholar] [CrossRef]
- Galatzer-Levy, I.R.; Steenkamp, M.M.; Brown, A.D.; Qian, M.; Inslicht, S.; Henn-Haase, C.; Otte, C.; Yehuda, R.; Neylan, T.C.; Marmar, C.R. Cortisol Response to an Experimental Stress Paradigm Prospectively Predicts Long-Term Distress and Resilience Trajectories in Response to Active Police Service. J. Psychiatr. Res. 2014, 56, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Lohr, J.B.; Palmer, B.W.; Eidt, C.A.; Aailaboyina, S.; Mausbach, B.T.; Wolkowitz, O.M.; Thorp, S.R.; Jeste, D.V. Is Post-Traumatic Stress Disorder Associated with Premature Senescence? A Review of the Literature. Am. J. Geriatr. Psychiatry 2015, 23, 709–725. [Google Scholar] [CrossRef] [PubMed]
- Miao, X.-R.; Chen, Q.-B.; Wei, K.; Tao, K.-M.; Lu, Z.-J. Posttraumatic Stress Disorder: From Diagnosis to Prevention. Mil. Med. Res. 2018, 5, 32. [Google Scholar] [CrossRef] [PubMed]
- Kok, B.C.; Herrell, R.K.; Thomas, J.L.; Hoge, C.W. Posttraumatic Stress Disorder Associated with Combat Service in Iraq or Afghanistan: Reconciling Prevalence Differences between Studies. J. Nerv. Ment. Dis. 2012, 200, 444–450. [Google Scholar] [CrossRef] [PubMed]
- Sundin, J.; Herrell, R.K.; Hoge, C.W.; Fear, N.T.; Adler, A.B.; Greenberg, N.; Riviere, L.A.; Thomas, J.L.; Wessely, S.; Bliese, P.D. Mental Health Outcomes in US and UK Military Personnel Returning from Iraq. Br. J. Psychiatry 2014, 204, 200–207. [Google Scholar] [CrossRef] [PubMed]
- Babaev, O.; Piletti Chatain, C.; Krueger-Burg, D. Inhibition in the Amygdala Anxiety Circuitry. Exp. Mol. Med. 2018, 50, 18. [Google Scholar] [CrossRef] [PubMed]
- Kolla, B.P.; Mansukhani, M.P.; Bostwick, J.M. The Influence of Antidepressants on Restless Legs Syndrome and Periodic Limb Movements: A Systematic Review. Sleep Med. Rev. 2018, 38, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, R.D.; Cowles, R.A.; Galea, S.; Jacobi, F. Gastritis and Mental Disorders. J. Psychiatr. Res. 2013, 47, 128–132. [Google Scholar] [CrossRef]
- Gong, S.; Miao, Y.-L.; Jiao, G.-Z.; Sun, M.-J.; Li, H.; Lin, J.; Luo, M.-J.; Tan, J.-H. Dynamics and Correlation of Serum Cortisol and Corticosterone under Different Physiological or Stressful Conditions in Mice. PLoS ONE 2015, 10, e0117503. [Google Scholar] [CrossRef]
- Ramos, A.T.; Tufik, S.; Troncone, L.R.P. Control of Stress-Induced ACTH Secretion by Vasopressin and CRH: Additional Evidence. Neuropsychobiology 2016, 73, 184–190. [Google Scholar] [CrossRef]
- Kyrou, I.; Tsigos, C. Stress Hormones: Physiological Stress and Regulation of Metabolism. Curr. Opin. Pharmacol. 2009, 9, 787–793. [Google Scholar] [CrossRef]
- Hariri, A.R.; Holmes, A. Finding Translation in Stress Research. Nat. Neurosci. 2015, 18, 1347–1352. [Google Scholar] [CrossRef] [PubMed]
- Transduction of Psychosocial Stress into the Neurobiology of Recurrent Affective Disorder. AJP 1992, 149, 999–1010. [CrossRef]
- Silva, C.C.; Correa, A.M.B.; Kushmerick, C.; Sharma, N.M.; Patel, K.P.; de Almeida, J.F.Q.; Moreira, F.A.; Ferreira, A.J.; Fontes, M.A.P. Angiotensin-Converting Enzyme 2 Activator, DIZE in the Basolateral Amygdala Attenuates the Tachycardic Response to Acute Stress by Modulating Glutamatergic Tone. Neuropeptides 2020, 83, 102076. [Google Scholar] [CrossRef] [PubMed]
- Loria, A.S.; Osborn, J.L. Maternal Separation Diminishes α-Adrenergic Receptor Density and Function in Renal Vasculature from Male Wistar-Kyoto Rats. Am. J. Physiol. Ren. Physiol. 2017, 313, F47–F54. [Google Scholar] [CrossRef] [PubMed]
- Cohen, B.E.; Edmondson, D.; Kronish, I.M. State of the Art Review: Depression, Stress, Anxiety, and Cardiovascular Disease. AJHYPE 2015, 28, 1295–1302. [Google Scholar] [CrossRef]
- Pêgo, J.M.; Sousa, J.C.; Almeida, O.; Sousa, N. Stress and the Neuroendocrinology of Anxiety Disorders. In Behavioral Neurobiology of Anxiety and Its Treatment; Stein, M.B., Steckler, T., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; Volume 2, pp. 97–118. ISBN 9783642029110. [Google Scholar]
- Rodrigues, R.; Petersen, R.B.; Perry, G. Parallels between Major Depressive Disorder and Alzheimer’s Disease: Role of Oxidative Stress and Genetic Vulnerability. Cell Mol. Neurobiol 2014, 34, 925–949. [Google Scholar] [CrossRef]
- Godoy, L.D.; Rossignoli, M.T.; Delfino-Pereira, P.; Garcia-Cairasco, N.; de Lima Umeoka, E.H. A Comprehensive Overview on Stress Neurobiology: Basic Concepts and Clinical Implications. Front. Behav. Neurosci. 2018, 12, 127. [Google Scholar] [CrossRef] [PubMed]
- Juruena, M.F. Early-Life Stress and HPA Axis Trigger Recurrent Adulthood Depression. Epilepsy Behav. 2014, 38, 148–159. [Google Scholar] [CrossRef]
- Spencer, R.L.; McEwen, B.S. Adaptation of the Hypothalamic-Pituitary-Adrenal Axis to Chronic Ethanol Stress. Neuroendocrinology 1990, 52, 481–489. [Google Scholar] [CrossRef]
- Raffington, L.; Schmiedek, F.; Heim, C.; Shing, Y.L. Cognitive Control Moderates Parenting Stress Effects on Children’s Diurnal Cortisol. PLoS ONE 2018, 13, e0191215. [Google Scholar] [CrossRef] [PubMed]
- Antoni, M.H.; Dhabhar, F.S. The Impact of Psychosocial Stress and Stress Management on Immune Responses in Patients with Cancer. Cancer 2019, 125, 1417–1431. [Google Scholar] [CrossRef]
- Warner-Schmidt, J.L.; Duman, R.S. Hippocampal Neurogenesis: Opposing Effects of Stress and Antidepressant Treatment. Hippocampus 2006, 16, 239–249. [Google Scholar] [CrossRef] [PubMed]
- Cuadrado-Tejedor, M.; Ricobaraza, A.; Frechilla, D.; Franco, R.; Pérez-Mediavilla, A.; Garcia-Osta, A. Chronic Mild Stress Accelerates the Onset and Progression of the Alzheimer’s Disease Phenotype in Tg2576 Mice. JAD 2012, 28, 567–578. [Google Scholar] [CrossRef] [PubMed]
- Kendler, K.S.; Gardner, C.O.; Prescott, C.A. Toward a Comprehensive Developmental Model for Major Depression in Men. AJP 2006, 163, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Kendler, K.S.; Gardner, C.O.; Prescott, C.A. Toward a Comprehensive Developmental Model for Major Depression in Women. AJP 2002, 159, 1133–1145. [Google Scholar] [CrossRef]
- Pisoschi, A.M.; Pop, A. The Role of Antioxidants in the Chemistry of Oxidative Stress: A Review. Eur. J. Med. Chem. 2015, 97, 55–74. [Google Scholar] [CrossRef] [PubMed]
- Burton, G.J.; Jauniaux, E. Oxidative Stress. Best Pract. Res. Clin. Obs. Gynaecol 2011, 25, 287–299. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B.; Gutteridge, J.M.; Cross, C.E. Free Radicals, Antioxidants, and Human Disease: Where Are We Now? J. Lab. Clin. Med. 1992, 119, 598–620. [Google Scholar] [PubMed]
- Gutteridge, J.M.C.; Halliwell, B. Invited Review Free Radicals in Disease Processes: A Compilation of Cause and Consequence. Free Radic. Res. Commun. 1993, 19, 141–158. [Google Scholar] [CrossRef]
- Angelier, F.; Costantini, D.; Blévin, P.; Chastel, O. Do Glucocorticoids Mediate the Link between Environmental Conditions and Telomere Dynamics in Wild Vertebrates? A Review. Gen. Comp. Endocrinol. 2018, 256, 99–111. [Google Scholar] [CrossRef] [PubMed]
- Salim, S. Oxidative Stress and the Central Nervous System. J. Pharmacol. Exp. Ther. 2017, 360, 201–205. [Google Scholar] [CrossRef]
- Hassan, W.; Noreen, H.; Castro-Gomes, V.; Mohammadzai, I.; da Rocha, J.B.T.; Landeira-Fernandez, J. Association of Oxidative Stress with Psychiatric Disorders. Curr. Pharm. Des. 2016, 22, 2960–2974. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, W.; Lingner, J. Impact of Oxidative Stress on Telomere Biology. Differentiation 2018, 99, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Pinak, M. CHAPTER 10—Enzymatic recognition of radiation-produced oxidative DNA lesion. Molecular dynamics approach. In Modern Methods for Theoretical Physical Chemistry of Biopolymers; Starikov, E.B., Lewis, J.P., Tanaka, S., Eds.; Elsevier Science: Amsterdam, The Netherlands, 2006; pp. 191–210. ISBN 9780444522207. [Google Scholar]
- Fouquerel, E.; Barnes, R.P.; Uttam, S.; Watkins, S.C.; Bruchez, M.P.; Opresko, P.L. Targeted and Persistent 8-Oxoguanine Base Damage at Telomeres Promotes Telomere Loss and Crisis. Mol. Cell 2019, 75, 117–130.e6. [Google Scholar] [CrossRef]
- Coluzzi, E.; Leone, S.; Sgura, A. Oxidative Stress Induces Telomere Dysfunction and Senescence by Replication Fork Arrest. Cells 2019, 8, 19. [Google Scholar] [CrossRef]
- Black, C.N.; Bot, M.; Scheffer, P.G.; Cuijpers, P.; Penninx, B.W.J.H. Is Depression Associated with Increased Oxidative Stress? A Systematic Review and Meta-Analysis. Psychoneuroendocrinology 2015, 51, 164–175. [Google Scholar] [CrossRef]
- Vaváková, M.; Ďuračková, Z.; Trebatická, J. Markers of Oxidative Stress and Neuroprogression in Depression Disorder. Oxidative Med. Cell. Longev. 2015, 2015, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Ceylan, D.; Yılmaz, S.; Tuna, G.; Kant, M.; Er, A.; Ildız, A.; Verim, B.; Akış, M.; Akan, P.; İşlekel, H.; et al. Alterations in Levels of 8-Oxo-2′-Deoxyguanosine and 8-Oxoguanine DNA Glycosylase 1 during a Current Episode and after Remission in Unipolar and Bipolar Depression. Psychoneuroendocrinology 2020, 114, 104600. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.W.; Lin, A.P.; Wolf, E.J.; Miller, D.R. Oxidative Stress, Inflammation, and Neuroprogression in Chronic PTSD. Harv. Rev. Psychiatry 2018, 26, 57–69. [Google Scholar] [CrossRef]
- Teyssier, J.-R.; Ragot, S.; Chauvet-Gélinier, J.-C.; Trojak, B.; Bonin, B. Expression of Oxidative Stress-Response Genes Is Not Activated in the Prefrontal Cortex of Patients with Depressive Disorder. Psychiatry Res. 2011, 186, 244–247. [Google Scholar] [CrossRef] [PubMed]
- Bouayed, J.; Rammal, H.; Soulimani, R. Oxidative Stress and Anxiety. Oxidative Med. Cell. Longev. 2009, 2, 63–67. [Google Scholar] [CrossRef] [PubMed]
- Eitan, E.; Hutchison, E.R.; Mattson, M.P. Telomere Shortening in Neurological Disorders: An Abundance of Unanswered Questions. Trends Neurosci. 2014, 37, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Lucassen, P.J.; Pruessner, J.; Sousa, N.; Almeida, O.F.X.; Van Dam, A.M.; Rajkowska, G.; Swaab, D.F.; Czéh, B. Neuropathology of Stress. Acta Neuropathol. 2014, 127, 109–135. [Google Scholar] [CrossRef] [PubMed]
- Powell, T.R.; De Jong, S.; Breen, G.; Lewis, C.M.; Dima, D. Telomere Length as a Predictor of Emotional Processing in the Brain. Hum. Brain Mapp. 2019, 40, 1750–1759. [Google Scholar] [CrossRef] [PubMed]
- Sokol, C.L.; Luster, A.D. The Chemokine System in Innate Immunity. Cold Spring Harb. Perspect. Biol. 2015, 7. [Google Scholar] [CrossRef] [PubMed]
- Anthony, D.C.; Couch, Y.; Losey, P.; Evans, M.C. The Systemic Response to Brain Injury and Disease. Brain Behav. Immun. 2012, 26, 534–540. [Google Scholar] [CrossRef]
- Block, M.L.; Zecca, L.; Hong, J.-S. Microglia-Mediated Neurotoxicity: Uncovering the Molecular Mechanisms. Nat. Rev. Neurosci. 2007, 8, 57–69. [Google Scholar] [CrossRef]
- Howren, M.B.; Lamkin, D.M.; Suls, J. Associations of Depression with C-Reactive Protein, IL-1, and IL-6: A Meta-Analysis. Psychosom. Med. 2009, 71, 171–186. [Google Scholar] [CrossRef]
- Dowlati, Y.; Herrmann, N.; Swardfager, W.; Liu, H.; Sham, L.; Reim, E.K.; Lanctôt, K.L. A Meta-Analysis of Cytokines in Major Depression. Biol. Psychiatry 2010, 67, 446–457. [Google Scholar] [CrossRef]
- Dahl, J.; Ormstad, H.; Aass, H.C.D.; Malt, U.F.; Bendz, L.T.; Sandvik, L.; Brundin, L.; Andreassen, O.A. The Plasma Levels of Various Cytokines Are Increased during Ongoing Depression and Are Reduced to Normal Levels after Recovery. Psychoneuroendocrinology 2014, 45, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Ho, R.C.-M.; Mak, A. Interleukin (IL)-6, Tumour Necrosis Factor Alpha (TNF-α) and Soluble Interleukin-2 Receptors (SIL-2R) Are Elevated in Patients with Major Depressive Disorder: A Meta-Analysis and Meta-Regression. J. Affect. Disord. 2012, 139, 230–239. [Google Scholar] [CrossRef] [PubMed]
- Wium-Andersen, M.K.; Ørsted, D.D.; Nielsen, S.F.; Nordestgaard, B.G. Elevated C-Reactive Protein Levels, Psychological Distress, and Depression in 73, 131 Individuals. JAMA Psychiatry 2013, 70, 176–184. [Google Scholar] [CrossRef]
- Menter, A.; Augustin, M.; Signorovitch, J.; Yu, A.P.; Wu, E.Q.; Gupta, S.R.; Bao, Y.; Mulani, P. The Effect of Adalimumab on Reducing Depression Symptoms in Patients with Moderate to Severe Psoriasis: A Randomized Clinical Trial. J. Am. Acad. Dermatol. 2010, 62, 812–818. [Google Scholar] [CrossRef]
- Langley, R.G.; Feldman, S.R.; Han, C.; Schenkel, B.; Szapary, P.; Hsu, M.-C.; Ortonne, J.-P.; Gordon, K.B.; Kimball, A.B. Ustekinumab Significantly Improves Symptoms of Anxiety, Depression, and Skin-Related Quality of Life in Patients with Moderate-to-Severe Psoriasis: Results from a Randomized, Double-Blind, Placebo-Controlled Phase III Trial. J. Am. Acad. Dermatol. 2010, 63, 457–465. [Google Scholar] [CrossRef] [PubMed]
- Raison, C.L.; Rutherford, R.E.; Woolwine, B.J.; Shuo, C.; Schettler, P.; Drake, D.F.; Haroon, E.; Miller, A.H. A Randomized Controlled Trial of the Tumor Necrosis Factor Antagonist Infliximab for Treatment-Resistant Depression: The Role of Baseline Inflammatory Biomarkers. JAMA Psychiatry 2013, 70, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Aubert, G.; Lansdorp, P.M. Telomeres and Aging. Physiol. Rev. 2008, 88, 557–579. [Google Scholar] [CrossRef] [PubMed]
- Franceschi, C.; Bonafè, M.; Valensin, S.; Olivieri, F.; De Luca, M.; Ottaviani, E.; De Benedictis, G. Inflamm-Aging. An Evolutionary Perspective on Immunosenescence. Ann. N. Y. Acad. Sci. 2000, 908, 244–254. [Google Scholar] [CrossRef]
- O’Donovan, A.; Pantell, M.S.; Puterman, E.; Dhabhar, F.S.; Blackburn, E.H.; Yaffe, K.; Cawthon, R.M.; Opresko, P.L.; Hsueh, W.-C.; Satterfield, S.; et al. Cumulative Inflammatory Load Is Associated with Short Leukocyte Telomere Length in the Health, Aging and Body Composition Study. PLoS ONE 2011, 6, e19687. [Google Scholar] [CrossRef]
- Salminen, A.; Kaarniranta, K.; Kauppinen, A. Inflammaging: Disturbed Interplay between Autophagy and Inflammasomes. Aging 2012, 4, 166–175. [Google Scholar] [CrossRef] [PubMed]
- Chinta, S.J.; Woods, G.; Rane, A.; Demaria, M.; Campisi, J.; Andersen, J.K. Cellular Senescence and the Aging Brain. Exp. Gerontol. 2015, 68, 3–7. [Google Scholar] [CrossRef] [PubMed]
- Iwata, M.; Ota, K.T.; Duman, R.S. The Inflammasome: Pathways Linking Psychological Stress, Depression, and Systemic Illnesses. Brain Behav. Immun. 2013, 31, 105–114. [Google Scholar] [CrossRef]
- Honda, T.; Segi-Nishida, E.; Miyachi, Y.; Narumiya, S. Prostacyclin-IP Signaling and Prostaglandin E2-EP2/EP4 Signaling Both Mediate Joint Inflammation in Mouse Collagen-Induced Arthritis. J. Exp. Med. 2006, 203, 325–335. [Google Scholar] [CrossRef] [PubMed]
- Narumiya, S. Prostanoids and Inflammation: A New Concept Arising from Receptor Knockout Mice. J. Mol. Med. 2009, 87, 1015–1022. [Google Scholar] [CrossRef] [PubMed]
- Aoki, T.; Narumiya, S. Prostaglandins and Chronic Inflammation. Trends Pharmacol. Sci. 2012, 33, 304–311. [Google Scholar] [CrossRef]
- Chung, H.Y.; Sung, B.; Jung, K.J.; Zou, Y.; Yu, B.P. The Molecular Inflammatory Process in Aging. Antioxid. Redox Signal. 2006, 8, 572–581. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Jung, K.J.; Yu, B.P.; Cho, C.G.; Choi, J.S.; Chung, H.Y. Modulation of Redox-Sensitive Transcription Factors by Calorie Restriction during Aging. Mech. Ageing Dev. 2002, 123, 1589–1595. [Google Scholar] [CrossRef]
- Lima, I. Verificação De Comprimento Telomérico Em Pacientes Com Transtorno Afetivo Bipolar E Sua Relação Com Aspectos Clínicos. Master Degree, UFMG, Belo Horizonte, Brazi, 2014. Available online: https://repositorio.ufmg.br/bitstream/1843/BUBD-AW5FUT/1/dissertac_ao.pdf (accessed on 8 August 2014).
- Hardbower, D.M.; de Sablet, T.; Chaturvedi, R.; Wilson, K.T. Chronic Inflammation and Oxidative Stress: The Smoking Gun for Helicobacter Pylori-Induced Gastric Cancer? Gut Microbes 2013, 4, 475–481. [Google Scholar] [CrossRef]
- Giorgi, C.; Marchi, S.; Simoes, I.C.M.; Ren, Z.; Morciano, G.; Perrone, M.; Patalas-Krawczyk, P.; Borchard, S.; Jędrak, P.; Pierzynowska, K.; et al. Mitochondria and Reactive Oxygen Species in Aging and Age-Related Diseases. Int. Rev. Cell Mol. Biol. 2018, 340, 209–344. [Google Scholar] [CrossRef] [PubMed]
- Gonzales-Ebsen, A.C.; Gregersen, N.; Olsen, R.K. Linking Telomere Loss and Mitochondrial Dysfunction in Chronic Disease. Front. Biosci. 2017, 22, 117–127. [Google Scholar] [CrossRef]
- Saretzki, G.; Murphy, M.P.; von Zglinicki, T. MitoQ Counteracts Telomere Shortening and Elongates Lifespan of Fibroblasts under Mild Oxidative Stress. Aging Cell 2003, 2, 141–143. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Trimarchi, J.R.; Smith, P.J.S.; Keefe, D.L. Mitochondrial Dysfunction Leads to Telomere Attrition and Genomic Instability. Aging Cell 2002, 1, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.; Vadodaria, K.C.; Lenkei, Z.; Kato, T.; Gage, F.H.; Marchetto, M.C.; Santos, R. Mitochondria, Metabolism, and Redox Mechanisms in Psychiatric Disorders. Antioxid. Redox Signal. 2019, 31, 275–317. [Google Scholar] [CrossRef]
- Cantó, C.; Auwerx, J. PGC-1alpha, SIRT1 and AMPK, an Energy Sensing Network That Controls Energy Expenditure. Curr. Opin. Lipidol. 2009, 20, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Sahin, E.; Colla, S.; Liesa, M.; Moslehi, J.; Müller, F.L.; Guo, M.; Cooper, M.; Kotton, D.; Fabian, A.J.; Walkey, C.; et al. Telomere Dysfunction Induces Metabolic and Mitochondrial Compromise. Nature 2011, 470, 359–365. [Google Scholar] [CrossRef] [PubMed]
- Lindqvist, D.; Epel, E.S.; Mellon, S.H.; Penninx, B.W.; Révész, D.; Verhoeven, J.E.; Reus, V.I.; Lin, J.; Mahan, L.; Hough, C.M.; et al. Psychiatric Disorders and Leukocyte Telomere Length: Underlying Mechanisms Linking Mental Illness with Cellular Aging. Neurosci. Biobehav Rev. 2015, 55, 333–364. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Authors | Date of Publication | Country | Study Design | Patients | Age, Years, Mean (SD) | Sex, n (%) Male | Main Comorbidities/ Lifestyle factors Associated | Race/Ethnicity | Other Associated Psychiatric Diseases in This Study | Medication | Telomere Measurement and Tissue | Measurement of Psychiatric Disorder | Level of Depression | Telomere Lenght | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Garcia-Rizo et al. [16] | Feburary 2013 | Spain | Case-control study | Control | 70 (For telomere content, n = 48) | 27.8 (6.8) | (62.2%) | BMI: 23.7 (2.9), Mean number of cigarettes/day: 6.2 (8.3) | NA | - | NA | mTL, Southern blot, Leukocytes | SCID | MDD (n = 15) | Shortened telomere are present early in the course of depression independently of the confounding. Abnormal glucose tolerance and lymphopenia were also related to MDD |
| Case | 15 (For telomere content, n = 9) | 30.7 (10.0) | 60% | BMI: 23.4 (4.1), Mean number of cigarettes/day: 9.9 (12.7) | NA | ||||||||||
| Chen et al. [17] | December 2014 | USA | Case-control study | Control | 20 | Patients were individually matched on age (±3 years), gender and ethnicity Full patients data were not available | - | NA | mTL, PCR, Leukocytes | QIDS, SCID | MDD (n = 20) | In healthy controls, greater ACE exposure was associated with shorter LTL but was unassociated with telomerase activity. In MDD, greater ACE exposure was unrelated to LTL but was associated with increased telomerase activity and with a higher telomerase: | |||
| Case | 20 | ||||||||||||||
| Tyrka et al. [18] | January 2016 | USA | Case-control study | Control | 113 | 28.5 (9.2) | 50 (44.2%) | Smokers (8.3%) | White (82.3%) | Adversities, depression, PTSD and anxiety | NA | mTL, qPCR, Leukocytes | SCID, STAI, PSS, CD-RISC | Significantly higher mtDNA copy numbers and shorter telomeres were seen in individuals with major depression, depressive disorders, and anxiety disorders, as well as those with parental loss and childhood maltreatment. | |
| Case 1—Adverity with no psychiatric disorder | 66 | 31.3 (11.1) | 26 (39.4%) | Smokers (7.8%) | White (80.3%) | ||||||||||
| Case 2—Psichyatric disorder with no adversity | 39 | 30.7 (10.4) | 15 (38.5%) | Smokers (7.7%) | White (92.3%) | MDD (n = 6), depressive (n = 7) | |||||||||
| Case 3—Adversity and psychiatric disorder | 72 | 34.8 (12.0) | 22 (30.6%) | Smokers (17.1%) | White (81.9%) | MDD (n = 7), depressive (n = 18) | |||||||||
| Prabu et al. [19] | July 2020 | India | Case-control study | Control—NGT and no depression | 40 | 48 (10) | 21 (52.5%) | BMI: 26 (4.2) | NA | - | 66 T2DM patients were on anti-diabetic medication alone. 14 T2DM patients were on anti-diabetic plus antihypertensive/statin medication. | RTL, rQ-PCR, blood sample | PHQ-9, PHQ-12 | NA | Patients with type 2 diabetes and depression exhibited increased circulatory levels of miR-128 and serum cortisol and shortened telomeres. |
| Case 1—NGT with depression | 40 | 50 (11) | 20 (50%) | BMI: 26.8 (5.8) | |||||||||||
| Case 2—T2DM and no depression | 40 | 54 (6) | 21 (52.5%) | BMI: 25.4 (4.9) | |||||||||||
| Case 3—T2DM with depression | 40 | 54 (7) | 21 (52.5%) | BMI: 25.5 (3.7) | |||||||||||
| Vincent et al. [20] | February 2017 | United Kingdom | Case-control study | Control | 100 | 50.84 (16.89) | F:51 (51%) M:49 (49%) | BMI: 26.89 (5.39) | White | - | NA | RTL, qPCR, Leukocytes | CIS-R, SCAN | - | Shortened RTL was specifically associated with childhood physical neglect, but not the other subtypes of maltreatment or depression case/control status. |
| Case | 80 | 48.63 (13.9) | F: 52 (65%) M: 28 (35%) | BMI: 28.47 (6.87) | Mild depression (n = 15), moderate or severe depression (55), mixed depression/anxiety (n = 10) | ||||||||||
| Hoen et al. [21] | August 2012 | Netherlands | Case-control study | Control | 980 | 53.7 (11.3) | F: 551 M: 465 | Smoking (n = 225; 77%), Alcohol consumption (n = 788; 80%), Sedentarism (n = 505; 52%) | NA | Anxiety | mTL, PCR, Leukocytes | CIDI | NA | No association was found between depressive disorders and shorter telomeres at follow-up. Anxiety disorders predicted shorter telomere length at follow-up in a general population cohort. | |
| Case | 97 | 51.3 (10.7) | F: 62 M: 36 | Smokers (n = 32; 65%); Alcohol consumption (n = 78; 80%), Sedentarism (n = 50; 52%) | Antidepressant use (n = 14; 15%) | ||||||||||
| Bersani et al. [22] | October 2015 | USA | Cross-sectional study | Control | 76 | 34.64 (9.17) | All male | Years of education (mean ± SD): 14.79 ± 2.44, current smokers (n): 11 | Hispanic (n = 35); Non Hispanic (n = 42)* | PTSD | Statins (n = 2), NSAIDs (n = 5), antidepressants (n = 13), antibiotics (n = 1), hormone drugs for prostate cancer (n = 1), analgesics (n = 1) | RTL, PCR, Granulocytes | CAPS, BDI-II, ETI, SCL-90-GSI, PSS, PANAS | PTSD associated with MDD (n = 17) | Early trauma, severity of perceived stress and general psychopathological symptoms are more closely associated with shorter TL than is the severity of core diagnostic symptoms of PTSD or MDD |
| Hoen et al. [23] | September 2011 | USA | Cohort study | Control | 746 | 68.1 (10.6) | 634 (85%) | Smoking (n = 131, 18%) | White (85%) | - | CDIS-IV, PHQ-9 | Depression is associated with reduced leukocyte telomere length in patients with coronary heart disease but does not predict 5-year change in telomere length | |||
| Case | 206 | 61.7 (10.8) | 142 (69%) | Smoking (n = 58, 28%) | White (60%) | Antidepressant use (n = 99, 48%) | mTL, qPCR, Leukocytes | Major Depression | |||||||
| AlAhwal et al. [24] | February 2019 | Saudi Arabia | Cross-sectional study | Cohort—Patients with colorectal cancer in Saudi Arabia | 50 | 54.5 (11.8) | 26 (52%) | - | NA | - | - | TL measurement not specified, rQ-PCR and tissue not specified | SCID-I, HDRS | MDD (10%), disthymia (10%), minor depression (4.0%) | TL progressively shortened from no depressive disorder to minor depression to dysthymia to major depressive disorder. TL was also strongly and inversely correlated with severity of depressive symptoms on the HDRS. |
| Needham et al. [11] | April 2015 | USA | Cross-sectional study | No depression | 966 | 29.2 (5.9) | 425 (44%) | NA | Non-Hispanic white (50.2%); African american (19.5%); Mexican American (30.3%) | Anxiety | Antidepressant use (n = 32, 3.3%) | RTL, qPCR, Leukocytes | CIDI | Neither depressive nor anxiety disorders were directly associated with telomere length, in young adults. There was suggestive evidence that pharmacologically-treated MD is associated with shorter telomere length, likely reflecting the more severe nature of MD that has come to clinical attention. | |
| MD or depressed affect | 198 | 30.3 (5.8) | 84 (42.4%) | Non-Hispanic white (51.0%); African american (19.7%); Mexican American (29.3%) | Antidepressant use (n = 20, 10.1%) | Depressed affect only (n = 123, 62.1%); Major depression (n = 75, 37.9%) | |||||||||
| Verhoeven et al. [25] | April 2018 | USA, Canada and The Netherlands | Cohort study | Year 15 | 977 | 40.5 (3.6) | 338 (34.6%) | Obese: 32.7%, former smoker: 18.7%, current smoker: 19%; heavy drinker: 9.3%; number of somatic diseases, mean (SD): 0.9 (1.1); physical activity, mean (SD): 336 (274) | White (59.4%) | - | Current antidepressant use (n = 67, 6.9%) | mTL, qPCR, Leukocytes | CES-D | CES-D score: 8.5 (7.3); Depression past year (n = 86, 8.8%); Depression life-time (n = 151, 15.5%) | Evidence for a long-term, between-person relationship of depressive symptoms with LTL, rather than a dynamic and direct within-person relationship |
| Year 20 | 45.4 (3.6) | Obese: 38.5%, former smoker: 21.5%, current smoker: 15.5%; heavy drinker: 10.3%; number of somatic diseases, mean (SD): 1.0 (1.2); physical activity, mean (SD): 334 (273) | Current antidepressant use (n = 101, 10.3%) | CES-D score: 9.0 (7.8); Depression past year (n = 111, 11.4%); Depression life-time (n = 182, 18.6%) | |||||||||||
| Year 25 | 50.4 (3.6) | Obese: 38.5%, former smoker: 21.5%, current smoker: 15.5%; heavy drinker: 10.3%; number of somatic diseases, mean (SD): 1.0 (1.2); physical activity, mean (SD): 335 (272) | Current antidepressant use (n = 109, 11.1%) | CES-D score: 9.0 (7.4); Depression past year (n = 131, 13,4%); Depression life-time (n = 210, 21.5%) | |||||||||||
| Verhoeven et al. [26] | June 2016 | Usa and The Netherlands | Cohort study | Baseline | 2936 | 41.8 (13.1) | 33.6% | Obese: 16.7%, former smoker: 33.2, current smoker: 38.7%; heavy drinker: 12.7% | NA | Current anxiety disorder (n = 535, 32.0%) | Tricyclic antidepressants (n = 79, 2.7%); Selective serotonin reuptake inhibitor (n = 502, 17.1), Other antidepressants (n = 164, 5.6%) | RTL, qPCR, Leukocytes | CIDI | Current Depressive diagnosis disorder (n = 389, 23.3%), Control (n = 644, 21.9%), Remitted (n = 620, 21.1%) | Robust associations of depressive and anxiety disorders with shorter telomeres, but interestingly, it did not demonstrate that depressive and anxiety disorders and LTL change together over time, suggesting the absence of a direct within-person relationship. Short LTL is suggested to be either a long-term consequence or an underlying vulnerability factor for depressive or anxiety disorders |
| Year 6 | 1883 | 48.6 (12.9) | 34.6% | Obese: 19.8%, former smoker: 42.3%, current smoker: 28.2%; heavy drinker: 9.9% | Current anxiety disorder (n = 190, 36.0%) | Tricyclic antidepressants (n = 56, 3.0%); Selective serotonin reuptake inhibitor (n = 224, 11.9), Other antidepressants (n = 104, 5.5%) | Current Depressive diagnosis disorder (n = 159, 30.1%), Control (n = 440, 23.4%), Remitted (n = 915, 48.6%) | ||||||||
| Whisman et al. [27] | February 2017 | USA | Cross-sectional study | 3609 | 67.0 (10.0) | 1404 (43.7) | BMI, mean (SD): 28.4 (6.0), Diabetes (19,5%), Cancer (14.1%), Heart disease (25.0), Stroke (6.0) | White (87.7%), Black (9.7%), Latin (7.4%), Other (2.6%) | - | NA | NA, RT-qPCR, saliva sample | CES-D | Depressive symptoms, mean (SD): 1.5 (2.0) | Higher levels of depressive symptoms were associated with shorter salivary telomeres in men, and this association was incremental to several potential confounds. | |
| Lin et al. [28] | February 2015 | USA | Cross-sectional study | 464 | 64.87 (10.99) | 368 (79.31) | Current smoker (n = 132, 28.45%), former smoker (n = 199, 42.89%), Non-Muscle-Invasive Bladder Cancer (n = 234, 53.67%) | White (93.1%), Hispanic (3.23%), Black (3.23%), Other (0.44%) | - | NA | RTL, rQ-PCR, Lymphocytes | CES-D, SCID | Alive with CESD < 16 (n = 280, 84.08%), Alive with CES-D ≥16 (63, 67.74%), Dead with CES-D < 16 (n = 53, 15.92%), Dead with CES-D ≥ 16 (n = 30, 32.26%) | Short telomere length and depressive symptoms are associated with bladder cancer mortality individually and jointly. | |
| Zhao et al. [29] | November 2016 | USA | Cross-sectional study | 2175 | 40.4 (17.0) | 847 (38.9%) | BMI: 31.3 (7.5), smoker (36.4%), drinker (58.0%, physical activity, steps/d, mean (SD): 5845.3 (3899.9) | American indians | - | Antidepressant use: 5.2% | RTL, qPCR, Leukocytes | CES-D | None (50.2%), mild (21.9%), moderate (15.6%), severe (12.3%) | Results showed that individuals with a higher level of depressive symptoms had shorter LTL. | |
| Shaffer et al. [30] | October 2012 | USA | Cross-sectional study | 2225 | 48.2 (18.9) | 1115 (50.1%) | BMI: 27.0 (5.5), Diabetes mellitus (n = 101, 4.5%), Previous ischemic heart disease (n = 160, 7.2%) | NA | - | NA | mTL, RT-PCR, Leukocytes | CES-D | Probable depressive disorder (CES-D ≥ 16): (n = 269, 12.1%), Elevated depressive symptoms (CES-D ≥ 10): n = 613 (27.6%) | Concurrent depressive symptoms were not associated with leukocyte telomere length in a large, representative, population-based study. | |
| Chae et al. [31] | January 2016 | USA | Cross-sectional study | 92 | 43.86 (5.73) | 92 (100%) | Smorkers (n = 51, 55.4%) | African american | Anxiety, mean (SD): 5.02 (3.86) | Medication use (non-specified): (n = 29, 31.5%) | mTL, qPCR, Leukocytes | CES-D, HADS | Depression, mean (SD): 7.51 (4.86) | Racial discrimination was associated with shorter LTL among those with lower levels of depressive symptoms | |
| Phillips et al. [32] | April 2013 | United Kingdom | Cross-sectional study | Youngest cohort | 337 | 36.6 (0.67) | 47% | Smoker (n = 76, 23%) | NA | - | Antidepressant use (n = 18, 5%) | mTL, qPCR, Leukocytes | HADS | Depression score at wave 5: 3.2 (4.54) | Depressive symptoms are longitudinally associated with shorter telomere length, but only in younger adults. |
| Middle cohort | 441 | 57.1 (111) | 45% | Smoker (n = 111, 25%) | Antidepressant use (n = 52, 12%) | Depression score at wave 5: 4.1 (4.66) | |||||||||
| Older cohort | 285 | 76.1 (0.84) | 45% | Smoker (n = 38, 13%) | Antidepressant use (n = 33, 12%) | Depression score at wave 5: 4.4 (4.50) | |||||||||
| Liu et al. [33] | April 2014 | China | Case-control study | Control 1—NGT and no depression | 46 | 51.22 (8.32) | 19 (41.30) | Current smokers (n = 12, 26.09%); Current drinkers (n = 7, 15.22%) | NA | - | NA | RTL, qPCR, Leukocytes | HADS-D | NA | Oxidative stress contributes to both telomere length shortening and depression development in newly diagnosed type 2 diabetic patients, while in depression status, some other mechanisms besides oxidative stress may also affect the telomere length |
| Control 2—NGT with depression | 6 | 55.33 (6.56) | 3 (50.00) | Current smokers (n = 2, 33.33%); Current drinkers (n = 1, 16.67%) | |||||||||||
| Case 1—T2DM and no depression | 54 | 54.83 (8.58) | 24 (44.44) | Current smokers (n = 16, 29.63%); Current drinkers (n = 11, 20.37%) | |||||||||||
| Case 2—T2DM with depression | 17 | 54.71 (8.10) | 10 (58.82) | Current smokers (n = 5, 29.41%); Current drinkers (n = 2, 11.76%) | |||||||||||
| Wang et al. [34] | April 2017 | Sweden | Case-control study | Control | 320 | 44.6 (12.5) | 197 (62) | Smokers—NA | NA | - | Telomere length, as compared with healthy controls, is shortened in patients with depression, anxiety and stress and adjustment disorders | ||||
| Case | 181 | 41.9 (11.1) | 22 (12) | Smokers (n = 27, 15%) | Antidepressant use (n = 63, 35%) | mTL, rQ-PCR, Leukocyte | PHQ-9, HADS-A/HADS-D, MADRS-S | Baselines—Median score (IQR): MADRS-S—20 (14–25); HAD-D—8 (6–11); HAD-A—12 (9–15); PHQ-9—13 (9–17) | |||||||
| Schaakxs et al. [35] | June 2014 | The Netherlands | Case-control study | Control | 128 | 70.1 (7.2) | 49 (38.3) | Cigarette years, median (IQR)—170.00 (598.75); moderate drinker, n (%)—99 (77.3); heavy drinker, n (%)—12 (9.4);years of education, mean (SD): 12.5 (3.5); Obesity, n (%): 26 (20.3%); physical activity, median (IQR): 2.61 (2.96) | NA | - | - | mTL, qPCR, Leukocyte | CIDI, IDS, BAI, AS | - | Mean TL did not differ between depressed persons and never-depressed comparisons |
| Case | 355 | 70.6 (7.4) | 120 (33,8) | Cigarette years, median (IQR)—100.00 (435.00); moderate drinker, n (%)—200 (56.3); heavy drinker, n (%)—15 (4.2); years of education, mean (SD): 10.3 (3.4); Obesity, n (%): 60 (16.9%); physical activity, median (IQR): 1.60 (2.96) | TCA, n (%): 78 (22.0); SSRI, n (%): 94 (26.6); Other antidepressants, n (%): 101 (28.5); Benzodiazepine use, n (%): 141 (39.7) | MDD, n (%): 249 (70.1); dysthymia, n (%): 6 (1.7); Co-morbid MDD and dysthymia, n (%): 92 (25.9%) | |||||||||
| Wolkowitz et al. [36] | May 2011 | USA | Case-control study | Control | 17 | 36.6 (11.8) | 6 (35%) | BMI: 24.8 (3.7); Daily current tobacco use (6%), Years of education, mean (SD): 15.82 (2.28), Yale Physical Activity Survey, mean (SD): 3.11(0.90) | Caucasian (71%), African-American (18%), Asian (6%) Other or Mixed (5%) | NA | mTL, PCR, Leukocyte | HDRS, SCID, ATHF | The depressed group, as a whole, did not differ from the controls in telomere length. However, telomere length was significantly inversely correlated with lifetime depression exposure, even after controlling for age | ||
| Case | 18 | 36.8 (11.0) | 6 (33%) | BMI: 26.3 (5.9); Daily current tobacco use (0%), Years of education, mean (SD): 15.28 (2.06), Yale Physical Activity Survey, mean (SD): 2.10(1.26) | Caucasian (72%), African-American (17%), Asian (6%) Other or Mixed (5%) | - | NA | ||||||||
| Georgin-Lavialle et al. [37] | January 2014 | France | Cross-sectional study | 19 | 43.42 (16.97) | 4 (21%) | NA | Caucasian (100%) | - | - | mTL, rQ-PCR, Leukocytes | DSM-IV, BDI-II, PSS | - | Telomere length was correlated to perceived stress (r = 0.77; p = 0.0001) but not to depression in our population. | |
| Verhoeven et al. [38] | November 2013 | USA and The Netherlands | Cohort study | Control | 510 | 40.5 (14.9) | 203 (39.8%) | Obesity (13.5%), former smoker (35.6%), current smoker (26.1%), heavy drinker (11.8%), Physical activity (in 1000 MET-minutes per week) (mean ± S.D.): 3.8 (3.0) | NA | - | SSRI (0.4%), benzodiazepine use (0.4%) | mTL, qPCR, Leukocyte | CIDI | IDS score, mean (SD): 5.4 (3.6) | Within the current MDD patients, separate analyses showed that both higher depression severity and longer symptom duration in the past 4 years were associated with shorter TL. The study also confirmed the imprint of past exposure to depression, as those with remitted MDD had shorter TL than controls. |
| Case 1: Remitted MDD | 802 | 43.5 (12.5) | 238 (29.7%) | Obesity (16.0%), former smoker (26.9%), current smoker (39.8%), heavy drinker (12.3%), Physical activity (in 1000 MET-minutes per week) (mean ± S.D.): 3.8 (3.0) | Comorbid anxiety disorder: 36.9% | TCA (2.9%), SSRI (16.4%), other antidepressant (3.7%), benzodiazepine use (4.4%) | IDS score, mean (SD): 18.0 (10.2) | ||||||||
| Case 2: Current MDD | 1095 | 40.7 (12.1) | 357 (32.6%) | Obesity (20.4%), former smoker (27.8%), current smoker (45.6%), heavy drinker (13.5%), Physical activity (in 1000 MET-minutes per week) (mean ± S.D.): 3.3 (3.1) | Comorbid anxiety disorder: 65.7% | TCA (4.1%), SSRI (29.6%), other antidepressant (11.0%), benzodiazepine use (14.6%) | IDS score, mean (SD): 32.6 (12.2) | ||||||||
| Putermanet et al. [39] | October 2013 | Cohort study | Control | 743 | 68.2 (10.5) | 84.7% | BMI: 28.29 (5.31), current smoking (17.6%), not at all physical activity (17.4%) | White (60.3%) | Statins (66.8%), Aspirin (72.9%), ARBs and ACEi (52.5%), Antidepressants (10.6%) | RTL, qPCR, Leukocytes | CDIS-IV | MDD was significantly related to LTL at 1 SD below the mean of multisystem resiliency, but not at 1 SD above the mean. This study suggests that MDD associations with biological outcomes should be examined within a psychosocial–behavioral context, because this context shapes the nature of thedirect relationship. | |||
| Case | 205 | 61.7 (10.8) | 143(69.8%) | BMI: 29.01 (5.68), current smoking (28.4%), not at all physical activity (22.9%) | White (60.0%) | Statins (58.6%), Aspirin (73.9%), ARBs and ACEi (50.2%), Antidepressants (48.8%) | NA | ||||||||
| Blom et al. [40] | November 2015 | Case-control study | Control | 63 | 15.8 (0.2) | 22 (35.18%) | NA | NA | NA | STL, qPCR, saliva sample | Adolescents with major depressive disorder exhibited significantly shorter telomere length and significantly smaller right, but not left hippocampal volume. | ||||
| Case | 54 | 15.9 (0.2) | 29 (53.97%) | Generalized anxiety disorder: 16 Social anxiety disorder: 2 Panic disorder: 1 Specific phobia: 4 Posttraumatic stress disorder: 5 Adjustment disorder: 1 Attention deficit hyperactivity disorder: 8 Alcohol/substance dependence: 1 Conduct disorder: 2 Oppositional defiance disorder: 3 Eating disorder (not otherwise specified): 2 | CTQ, BDI-II, CDRS | Beck Depression Inventory II: 26.7 (1.5); Children’s Depression Inventory: 24.1(1.1) | |||||||||
| Vance et al. [41] | April 2018 | USA | Cohort study | Control | 67 | 44.1 (14.0) | 28 (42%) | Highest educational level: Graduate school (33%), College graduate (42%), Partial college (19%), High school graduate or lower (6%). Living with partner/married: 46%. BMI: 24.3 (3.8). Past year exercise level, more than once a week: (91%). Lifetime alcohol or substance use disorder: (10%). Cigarette smoking pack-years, mean (SD): 8.0 (17.3) | White: 73%; Hispanic/Latino: 3% | RTL, qPCR, Leukocyte | MADRS, HAM-A, PSS, ETISR-SF, TEQ | Individuals with MDD at baseline had greater LTL shortening over two years than individuals without MDD (p = 0.03), even after controlling for differences in age, sex, and body mass index (BMI). In the sub-sample of individuals with MDD diagnoses at baseline, no significant associations between LTL change and symptom severity or duration were found. | |||
| Case | 50 | 42.7 (13.2) | 24 (48%) | Highest educational level: Graduate school (18%), College graduate (40%), Partial college (28%), High school graduate or lower (18%). Living with partner/married: 18%. BMI: 26.0 (4.7). Past year exercise level, more than once a week: (63%). Lifetime alcohol or substance use disorder: (22%). Cigarette smoking pack-years, mean (SD): 6.8 (12.5) | White: 82%; Hispanic/Latino: 4% | Antidepressant use >6 months: 52% | MDD, n (%): 15(37%) | ||||||||
| Starnino et al. [42] | October 2016 | Canada | Cohort study | 132 | 45.34 (11.16) | 54 (40,9%) | Glasses of alcohol/week, mean (SD): 3.84 (5.38); Smoker n (%): 19(14%), BMI: 25.35 (5.00) | NA | Anxiety and hostility | - | RTL, qPCR, Leukocyte | BDI-II, BAI | Beck Depression Inventory-II, mean (SD): 7.64 (8.21) | Shorter TL was observed among individuals high in defensiveness and depressive symptoms, as well as in those with less hostility and anxiety. Telomere length is associated with psychological burden though the direction of effect differs depending on the psychological variables under study | |
| Wang et al. [43] | September 2019 | China | Cross-sectional study | 1742 | 63.6 (4.9) | 819 (47.0%) | Education level, n (%): Illiterate: 1020 (58.6), Primary: 432 (24.8), Junior: 196 (11.3), Senior or above: 94 (5.4) | NA | - | NA | RTL, q-PCR, Leukocyte | GDS | Mild depressive, n (%): 155 (8.9) Severe depressive, n (%) 39 (2.2) | Depressive symptoms was negatively correlated with TL in the overall sample. Depressive symptoms significantly mediated the relationship between religiosity and TL (explaining 31.8% of the total variance) in the 65 years and older subgroup | |
| Wikgren et al. [44] | September 2011 | Sweden | Case-control study | Control | 451 | 58.9 (11.6) | 224 (50%) | BMI: 26.1 (3.5), smoking, n (%): 50 (11%) | NA | - | - | NA, qPCR, Leukocytes | BDI, BAI, CES-D, PSQ | CES-D, Median Score (IQR): 6 (3–10) | TL was shorter among patients compared with control subjects (277 base pairs, p = 0.001). Overall, short TL was associated with a hypocortisolemic state (low post-DST cortisol and high percentage of cortisol reduction after the DST) among both patients and control subjects but more pronounced among patients. |
| Case | 91 | 60.4 (13.1) | 36 (40%) | BMI: 26.6 (3.7), smoking, n (%): 14 (15%) | Antidepressant use, n (%): 81 (89%) | CES-D, Median Score (IQR): 11 (6–22) | |||||||||
| Szebeni et al. [45] | June 2014 | USA | Case-control study | Control | 14 | 51 (5) | 13 (93%) | NA | NA | - | NA | RTL, PCR, white matter oligodendrocytes | NA | Relative telomere lengths in white matter oligodendrocytes, but not astrocytes, from both brain regions were significantly shorter for MDD donors as compared to matched control donors | |
| Case | 14 | 51 (5) | 14 (93%) | NA | NA | - | NA | MDD (100%) | |||||||
| Liu et al. [46] | July 2017 | Sweden | Cross-sectional study | 894 | Age, years, median (IQR): 46 (39, 54) | 304 (34%) | Education (%): 0–12 years (51.8%), More than 12 years (48.2%), Obesity (7.4%), Smokers (22.7%), Physical exercise regularity (54.2%) | NA | - | NA | RTL, qPCR, saliva | DSM-IV, AVSI, AVAT | - | In females, depressive status and age had direct negative effects on TL. For males, the effects of stressors and depressive status on TL were mediated by social interaction and the coping strategy worry. In females, no mediation effect of social interaction and coping strategy was detected. | |
| Boeck et al. [47] | June 2018 | Germany | Case-control study | Control | 21 | 57.5 (5.7) | 0% | BMI: 24.5 (3.0); Smoking, yes, n (%): 3(14.3%); Physical activity, yes, n (%):18 (85.7%) | NA | - | Medication, n (%): Antihypertensive drugs: 3 (14.3%), Tyroid hormone: 3 (14.3%), Sedatives: 1 (4.8%) | mTL, qFISH, PBMC | BDI-II | BDI-II sum score (mean ± S.D.): 2.1 ± 2.2 | All of the observed TL changes were signifcantly or marginally signifcantly associated with depressive symptom severity as assessed by the Becks Depression Inventory (BDI-II) sum score. Furthermore, the BDI-II also showed a signifcant negative correlation with TL in memory T helper cells |
| Case | 18 | 59.3 (6.6) | 0% | BMI: 29.2 (7.5); Smoking, yes, n (%): 8(44.4%); Physical activity, yes, n (%):11 (61.1%) | Medication, n (%): Antidepressants: 13 (72.2%) Antipsychotics: 5 (27.8%) Antihypertensive drugs: 7 (38.9%) Tyroid hormone: 5 (27.8%) Sedatives: 5 (27.8%) Analgesics: 3 (16.7%) Laxatives: 2 (11.1%) Vitamins (B1,B6,B12): 1 (5.6%) Statins: 1 (5.6%) | BDI-II sum score (mean ± S.D.): 23.8 ± 10.9 | |||||||||
| Jiménez et al. [48] | October 2018 | Colombia | Case-control study | Control | 52 | 21 (3) | F:36 (69%) M:16 (31%) | NA | latin | - | NA | RTL, MMqPCR, Leukocyte | PHQ-9, CES-D, HADS, CTQ | - | Correlation in the clinically significant depressive symptoms group between TL and sexual abuse |
| Case | 40 | 21 (3) | F:34 (85%) M:6 (15%) | ||||||||||||
| Wium-Andersen et al. [49] | January 2017 | Denmark | Cross-sectional study | Quartile 1 | 16,820 | 52 (42–61) | 6943 (41%) | Never smokers, No. (%): 7029 (42), Drinks/week, median (IQR):7 (3–14), Less than 3 years of education, No. (%): 10,062 (60), Low leisure time physical activity, inactive or less than 2–4 h light/day, No. (%): 8658 (51%), BMI, median (IQR): 25 (23–28), chronic disease, No. (%): 5647 (34%) | - | Prescription antidepressant medication, No (%): 7748 (12%) | mTL, modified MMqPCR, Leukocyte | ICD-8, ICD-10 | NA | Attendance at hospital for depression was associated with short telomere length cross-sectionally, but not prospectively. Further, purchase of antidepressant medication was not associated with short telomere length cross-sectionally or prospectively. The genetic analyses suggested that telomere length was not causally associated with attendance at hospital for depression or with purchase of antidepressant medication. | |
| Quartile 2 | 16,829 | 55 (45–65) | 7391 (44%) | Never smokers, No. (%): 6557 (39), Drinks/week, median (IQR):7 8 (3–15), Less than 3 years of education, No. (%): 10,260 (61), Low leisure time physical activity, inactive or less than 2–4 h light/day, No. (%): 8931 (53), BMI, median (IQR): 25 (23–28), chronic disease, No (%): 6415 (38) | |||||||||||
| Quartile 3 | 16,828 | 59 (49–68) | 7630 (45%) | Never smokers, No. (%): 6096 (36), Drinks/week, median (IQR):8 (4–15), Less than 3 years of education, No. (%): 10,698 (64), Low leisure time physical activity, inactive or less than 2–4 h light/day, No. (%): 9100 (54), BMI, median (IQR): 26 (23–29), chronic disease, No. (%): 7320 (44) | |||||||||||
| Quartile 4 | 16,829 | 64 (54–72) | 8100 (48%) | Never smokers, No. (%): 7029 (42), Drinks/week, median (IQR):7 (3–14), Less than 3 years of education, No. (%): 10,062 (60), Low leisure time physical activity, inactive or less than 2–4 h light/day, No. (%): 8658 (51%), BMI, median (IQR): 25 (23–28), chronic disease, No (%): 5647 (34%) | NA | ||||||||||
| Huzen et al. [50] | January 2010 | The Netherlands and United Kingdom | Cross-sectional study | 890 | 73 (64–79) | 535 (61%) | BMI: 26 (24–30) | NA | - | - | mTL, RT-qPCR, Leukocyte | CES-D, DS14, RAND-36 | Severe depression, n (%): 154 (18%), depressive symptoms only, n (%): 145 (16,3%) | A lower perceived mental health on the RAND-36 score was associated with shorter telomere length. Telomere length was not associated with the CES-D or DS14 score. | |
| Simon et al. [51] | August 2015 | USA | Case-control study | Control | 166 | 41.3 (13.7) | 77 (46%) | Educational level: Graduate school: 35%, College graduate (40%), Partial college (18%), High school graduate or less (7%). Lifetime alcohol or drug abuse/dependence, N (%): 13 (8). BMI: 25.4 (4.2) | White: 68%, Black or African American:17%, Asian:7%, Native American/Alaska Native: 0%, Other: 8%. Not Hispanic/Latino: 90%, Hispanic/Latino: 10% | RTL, RT-qPCR, Southern blot, Leukocytes | DSM-IV, SCID, MADRS, CIRS, TEQ, ETISR-SF, ICG | MADRS Total Score, Mean (SD): 1.9 (2.4) | Our well-characterized, well-powered examination of concurrently assessed telomere length and telomerase activity in individuals with clinically significant, chronic MDD and matched controls failed to provide strong evidence of an association of MDD with shorter LTL, while telomerase activity was lower in men with MDD. | ||
| Case | 166 | 41.3 (13.8) | 77(46%) | Educational level: Graduate school: 20%, College graduate (35%), Partial college (25%), High school graduate or less (17%). Lifetime alcohol or drug abuse/dependence, N (%): 47 (28%) | White: 78%, Black or African American:11%, Asian:4%, Native American/Alaska Native: 1%, Other: 4%. Not Hispanic/Latino: 92%, Hispanic/Latino: 7% | Current anxiety disorder (50%), litime anxiety disorder (55%) | Anti-depressant use > 6 months, N (%): 65 (39), Mood stabilizer use > 6 months, N (%): 6 (4), Benzodiazepine use > 6 months, N (%): 19 (11) Antipsychotic use > 6 months, N (%): 10 (6) | MADRS Total Score, Mean (SD): 28.2 (6.0) | |||||||
| Harttman et al. [52] | November 2010 | Germany | Case-control study | Control | 20 | 49.1 (15.2) | 11 (55%) | Smokers (n = 7, 35%) | NA | - | Major depressive disorder is associated with shortened telomeres. However, differences in the applied therapy, the duration of illness, or the severity of depression do not seem to have any influence on telomere length. | ||||
| Case | 54 | 49.1 (14.1) | 21 (39%) | Smokers (n = 16, 29.6%) | NA | - | TAD ≤ 1: n = 20, TAD > 1 n = 16 | mTL, Southern blot, Leukocyte | HAM-D | HAM-D, (SD) [Range]: 29.9 (6.0) [7,8,9,10,11,12,13,14,15,16,17,18,19,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36] | |||||
| Karabatsiakis et al. [53] | July 2014 | Germany | Case-control study | Control | 50 | 51.1 (8) | 0% | Years of education, mean (SD): 15.1 (2.4) | NA | - | mTL, qFISH, Leukocyte | BDI | NA | : A history of depression is associated with shortened telomeres in the main effector populations of the adaptive immune system. Shorter telomeres seem to persist in individuals with lifetime depression independently of the severity of depressive symptoms. | |
| Case 1—Lifetime depressed with irrelevant symptoms | 24 | 53.1 (7.2) | 0% | Years of education, mean (SD): 14.1 (2.1) | SSRI/ SNRI intake, n (%): 10 (41.7%) | ||||||||||
| Case 2—Relevant symptoms of depression | 20 | 53.8 (7.6) | 0% | Years of education, mean (SD): 14.2 (2.9) | SSRI/ SNRI intake, n (%): 9 (45%) | ||||||||||
| Solomon et al. [54] | July 2017 | Israel | Cohort study | 99 | 63.6 (3.7) | NA | Years of education (M, SD): 14, 3.7, Physical exercising regularly: 62 (62.6%) Smoking on a regular basis: 21 (21.2%) | NA | PTSD | NA | mTL, Southern blot, Leukocyte | SCL-90 | NA | Ex-POWs had shorter telomeres compared to controls (Cohen’s d = 0.5 indicating intermediate effect). Ex-POWs with chronic depression had shorter telomeres compared to those with delayed onset of depression (Cohen’s d = 4.89), and resilient ex-POWs (Cohen’s d = 3.87), indicating high effect sizes. | |
| Verhoeven et al. [55] | November 2019 | The Netherlands | Cohort study | 2032 | 42.5 (12.8) | 665 (32.7%) | NA | European ancestry: 2032 (100%) | - | NA | RTL, qPCR, Leukocyte | CIDI, DSM-IV, IDS-SR, NEO-FFI | Lifetime depression diagnosis (% yes, N): 83.1, 1688 Depression severity (average 6-year IDS score) (mean ± S.D.): 19.0 (12.0) | The use of genetic methods in this paper indicated that the established phenotypic association between telomere length and depression is unlikely due to shared underlying genetic vulnerability. These findings suggest that short telomeres in depressed patients may simply represent a generic marker of disease or may originate from non-genetic environmental factors | |
| Mamdani et al. [56] | September 2015 | USA | Case-control study | Control | 10 | 48 (13.0) | 7 (70%) | NA | NA | Schizofrenia and bipolar disorder | NA | RTL, qPCR, brain tissue | UCCIB psychological autopsy protocol | A significant decrease in telomere length was observed specifically in the HIPP of MDD subjects even after controlling for age. In the HIPP of MDD subjects, several genes involved in neuroprotection and in stress response showed altered levels of mRNA. | |
| Case | 10 | 47.3 (11.5) | 3 (30%) | NA | NA | MDD: 10 (100%) | |||||||||
| Authors | Date of Publication | Country | Study Design | Patients | Age, Years, Mean (SD) | Sex, n (%) Male | Main Comorbidities/ Lifestyle Factors Associated | Race/Ethnicity | Other Associated Psychiatric Diseases in this Study | Medication | Telomere Measurement and Tissue | Measurement of Psychiatric Disorder | Telomere Lenght | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Zhang et al. [57] | April 2019 | USA | Cross-sectional | Low hostility | 135 | 31.3 ± 8.7 | 127 (29.8%) | n (%) <12th grade 3 (2.2), High school diploma or G.E.D 33 (24.6), Some college/technical school 57 (42.5), Bachelor’s degree 24 (17.9), Graduate degree 17 (12.7) | 364 Whites, 61 Blacks, 25 Asian or Pacific Islanders, 11 American Indian or Alaskan Natives, and 13 unknown | Depression (28%) and suicide ideation (24%) of the PTSD subjects | NA | RTL, qPCR, Leukocytes | BSI, PCL | Among the participants with PTSD, those with medium or high level of hostility had shorter LTL than those with low level hostility (p < 0.01). Stepwise regression indicated that hostility level and age, but not gender and PTSD, were negatively correlated with LTL. |
| Medium hostility | 267 | 28.5 ± 7.1 | 237 (55.6%) | <12th grade 1 (1.1), High school diploma or G.E.D 73 (27.2), Some college/technical school 133 (49.6), Bachelor’s degree 46 (17.2), Graduate degree 13 (4.8) | ||||||||||
| High hostility | 72 | 26.5 ± 5.9 | 62 (14.6%) | <12th grade 3 (1.4), High school diploma or G.E.D 37 (51.4), Some college/technical school 29 (40.3), Bachelor’s degree 5 (6.9), Graduate degree 0 (0.0) | ||||||||||
| Bersani et al. [22] | October 2015 | USA | Cross-sectional | Control | 41 | 34.64 (9.17) | 41 (100%) | Years of education (mean ± SD): 14.79 ± 2.44, current smokers (n): 11 | 35 hispanic and 42 non-hispanic | Depression | Statins (n = 2), NSAIDs (n = 5), antidepressants (n = 13), antibiotics (n = 1), hormone drugs for prostate cancer (n = 1), analgesics (n = 1) | RTL, PCR, Granulocytes | CAPS, BDI-II, ETI, SCL-90-GSI, PSS, PANAS | TL was negatively correlated with early trauma (p < 0.001),global psycho-pathological severity (p = 0.044) and perceived stress (p = 0.019), positively correlated with positive affect (p = 0.026), not significantly correlated with symptom severity of PTSD, depression or negative affect. |
| Case | 35 (17 also w/ MDD) | 35 (100%) | ||||||||||||
| O’Donovan et al. [58] | September 2011 | USA | Cross-sectional | Control | 47 | 30.68 ± 8.19 | 21 (44.68%) | Female: Education: 15.4 (2.0), BMI: 25.2 (4.2), Current smoker n (%): 5 (20), Alcohol use n (%): 0 (0), Substance use n (%): 1 (4) Past abuse. | 6 African americans, 9 ssian americans, 57 whites, 1 hispanic, 2 hawaiians, 4 pacific islanders and 9 multi-ethnics | 23 past MDD, 8 current MDD | - | qPCR | CAPS, DSM-IV | Participants with PTSD had shorter age-adjusted LTL than controls. Exposure to childhood trauma was also associated with short LTL. In fact, childhood trauma appeared to account for the PTSD group difference in LTL; only participants with PTSD and exposure to multiple categories of childhood trauma had significantly shorter LTL than controls. |
| Male: Education: 15.5 (2.1), BMI: 23.6 (3.1), Current smoker n (%): 3 (14), Alcohol use n (%): 1 (5) Past abuse, Substance use n (%): 0 (0). | ||||||||||||||
| Case | 43 | 30.60 ± 6.63 | 23 (53.48%) | Female: Education: 15.2 (2.1), BMI: 23.9 (2.0), Current smoker n (%): 6 (30), Alcohol use n (%): 4 (20 Past abuse and 4 (20) past dependence, Substance use n (%): 1 (5) past abuse and 2(10) past dependence | ||||||||||
| Male: Education: 14.4 (2.3), BMI: 29.5 (4.3), Current smoker n (%): 4 (18), Alcohol use n (%): 4 (18) Past abuse and 4 (18) past dependence, Substance use n (%): 0 (0) Past abuse and 4 (18) past dependence | ||||||||||||||
| Kang et al. [59] | July 2020 | USA | Cross-sectional | Control 1: low combat exposure | 59 | 33.17 (8.69) | 83 (100%) | Education, Years: 15.01 ± 2.09, Body Mass Index, kg/m2: 28.19 ± 4.14, Waist Circumference, cm: 93.47 ± 11.14, Smoker, Yes/No, n: 17/94, Alcohol Abuse, Yes/No, n: 8/103, Substance Use, Yes/No, n: 2/109, MDD Diagnosis by SCID, Yes/No, n: 0/111, Time Since Severe Combat Event, Months: 65.85 ± 35.15 | 31 Hispanic, 80 others | - | 3 Antidepressant, 6 benzodiazepines and hypnotics and 1 anticonvulsants | RTL, qPCR, Leukocytes | DSM-IV, CAPS, PSS, PANAS | Subjects with PTSD showed shorter telomere length and larger amygdala volume than those without PTSD among veterans exposed to high trauma, while there was no significant group difference in these parameters among those exposed to low trauma. |
| Control 2: high combat exposure | 24 | |||||||||||||
| Case 1: low combat exposure | 12 | 33.66 (8.17) | 65 (100%) | Education, Years: 13.93 ± 1.90, Body Mass Index, kg/m2: 29.79 ± 5.58, Waist Circumference, cm: 98.61 ± 14.72, Smoker, Yes/No, n: 33/69, Alcohol Abuse, Yes/No, n: 13/89, Substance Use Yes/No, n: 6/96, MDD Diagnosis by SCID, Yes/No, n: 50/52, Time Since Severe Combat Event, Months: 77.67 ± 30.95 | 46 Hispanic, 56 others | 50 MDD | 21 Antidepressant, 8 benzodiazepines and hypnotics, 2 antipsychotics and 5 anticonvulsants | |||||||
| Case 2: high combat exposure | 53 | |||||||||||||
| Malan et al. [60] | August 2011 | South Africa | Cohort | Control | 53 | 22.3 | 0 (0) | High school education (Grade 8 and higher) (59 (92%)), primary school education (5 (7.81%)) | 12 (19%) black, 1 (1%) white, and 51 (80%) individuals of mixed ancestry. | 23 (36%) diagnosed with MDD at baseline and 31 (48%) diagnosed with MDD at the 3-month follow-up. | NA | RTL, qPCR, Leukocytes | DSM-IV, CD-RISC, BDI, CES-D, MADRS, ETI | A marginally significant association was evident between relative LTL and PTSD status. |
| Case | 9 | 0 (0) | NA | |||||||||||
| Boks et al. [61] | January 2015 | Netherlands | Cross-sectional | Control: low trauma | 0 | 25.1 (8.1) | 128 (100%) | Smokers = 19, Increase alcohol use = 15, Decrease alcohol use = 10, Unchanged alcohol use = 31 | Dutch ethinicity | - | antibiotics in 4 cases, antihistamines in 3 cases and one case of a benzodiazepine prescription. Stopped medication included: antibiotics in one case, antihistaminic in two cases and one case of oral isotretinoïn. | mTL, qPCR, blood sample | SRIP, ETI | Development of post traumatic stress disorder (PTSD) symptoms was significantly associated with increased telomere length and decreased DNAm ageing. |
| Case: low trauma | 64 | |||||||||||||
| Control: high trauma | 32 | 27.4 (9.3) | ||||||||||||
| Case: high trauma | 32 | |||||||||||||
| Roberts et al. [62] | May 2017 | USA | Cross-sectional | Control | 25 | 45.5 (3.6) | 0 (0) | BMI, blood draw, kg/m2 (Mean (SD)): 25.1 (4.0), Past-month smoking, blood draw, any %(N): 14.3 (4), Past-month alcohol consumption, blood draw, none %(N): 32.1 (9), Diet, least healthy quintile, 1995 %(N): 12.0 (3), Past-month physical activity, blood draw, <1/week %(N): 32.1 (9), High cholesterol, 1995 %(N): 7.1 (1), High blood pressure, 1995 %(N): 3.6 (1) | NA | Depression | Antidepressant 10.7% | RTL, RT-qPCR, Leukocytes | DSM-IV, PHQ-9, PCL-C | Relative to not having PTSD, women with a PTSD diagnosis had shorter log-transformed TL. Adjustment for health behaviors and medical conditions did not attenuate this association. Trauma type was not associated with TL and did not account for the association of PTSD with TL. |
| Case | 66 subclinical PTSD | 46.6 (3.7) | 0 (0) | BMI, blood draw, kg/m2 (Mean (SD)): 26.0 (7.3), Past-month smoking, blood draw, any %(N): 9.0 (6), Past-month alcohol consumption, blood draw, none %(N): 43.9 (29), Diet, least healthy quintile, 1995 %(N): 15.2 (10), Past-month physical activity, blood draw, <1/week %(N): 39.4 (26), High cholesterol, 1995 %(N): 7.6 (5), High blood pressure, 1995 %(N): 3.0 (2) | Antidepressant 9.1% | |||||||||
| 25 PTSD diagnosis | 46.6 (3.9) | 0 (0) | BMI, blood draw, kg/m2 (Mean (SD)): 28.4 (7.6), Past-month smoking, blood draw, any %(N): 16.0 (4), Past-month alcohol consumption, blood draw, none %(N): 56.0 (14), Diet, least healthy quintile, 1995 %(N): 28.0 (7), Past-month physical activity, blood draw, <1/week %(N): 32.0 (8), High cholesterol, 1995 %(N): 12.0 (3), High blood pressure, 1995 %(N): 8.0 (2) | Antidepressant 52% | ||||||||||
| Ladwig et al. [63] | July 2013 | Germany | Cross-sectional | Control | 2687 | 56.5 | 1330 (49.5%) | Means (SD) Low educational level (%) 59.0; Living alone (%) 24.1; BMI (kg/m2) 27.6 (4.8); Current smoking (%) 17.8; Alcohol consumption: No 30.1, Moderate 52.7, High 17.2; Physical inactivity (%) 45.7; Actual hypertension (%) 31.7; TC/HDL-C 4.09 (1.18); History of chronic diseases 16.8 | NA | Depression (PHQ-9) (%)* 3.9, Depressed mood/exhaustion (DEEX) (%)* 18.4 | NA | mTL, qPCR, Leukocytes | PDS, PHQ-9, DEEX | The multiple model revealed a significant association between partial PTSD and TL as well as between full PTSD and shorter TL indicating shorter TL on average for partial and full PTSD. An additional adjustment for depression and depressed mood/exhaustion gave comparable beta estimations. |
| Case | 262 partial PTSD | 52.5 | 100 (38.2%) | Low educational level (%) 57.6; Living alone (%) 29.4; BMI (kg/m2) 27.4 (5.1); Current smoking (%) 21.4; Alcohol consumption: No 30.2, Moderate 49.6, High 20.2; Physical inactivity (%) 42.7; Actual hypertension (%) 24.4; TC/HDL-C 4.04 (1.20); History of chronic diseases 22.9 | Depression (PHQ-9) (%)* 13.0, Depressed mood/exhaustion (DEEX) (%)* 55.0 | |||||||||
| 51 full PTSD | 54.5 | 19 (37.3%) | Low educational level (%) 64.7; Living alone (%) 35.3; BMI (kg/m2) 28.1 (5.7); Current smoking (%) 9.8; Alcohol consumption (%): No 35.3, Moderate 41.2, High 23.5; Physical inactivity (%) 47.1; Actual hypertension (%) 27.5; TC/HDL-C 4.02 (1.21); History of chronic diseases 19.6 | Depression (PHQ-9) (%)* 5.9, Depressed mood/exhaustion (DEEX) (%)* 56.9 | ||||||||||
| Jergović et al. [64] | June 2014 | Croatia | Case-control study | Control | 17 age-matched | 47.2 (1.71) | 17 (100%) | Body mass index 27.13 ± 4; Education: Elementary/high 29 (96.4), University 1 (3.4) Work status: Employed 1 (3.4); Unemployed/retired 29 (96.4); Tobacco use: Yes 17 (58.6); No 12 (41.4); Alcohol use: Yes 5 (20); No 24 (80); Daily physical exercise 1 (3.33) | NA | - | NA | RTL, RT-PCR, PBMCs | CAPS, STAI, BDI | Middle-aged war veterans with current PTSD had shorter PBMC telomere length than their age-matched healthy controls while the elderly had the shortest telomeres. |
| 15 elderly | 80 or older | 2 (13.33%) | NA | NA | - | NA | ||||||||
| Case | 30 | 45.9 (1.12) | 30 (100%) | mean ± SD Body mass index 27.3 ± 2.62; Education: Elementary/high 29 (96.4), University 6 (35.3); Work status: Employed 15 (88.2), Unemployed/retired 2 (11.8); Tobacco use: Yes 9 (52.9), No 8 (47.1); Alcohol use: Yes 11 (64.7), No 6 (35.3); Daily physical exercise 1 (5.88) | NA | 24 (80%) MDD, 13 (43%) panic disorder, 9 (30%) obsessive compulsive disorder, 7 (23%) social phobia | Analgesics (non-steroidal anti-inflammatory drugs, N = 18, 60%; opioid analgesics, N = 3, 10%), hypolipidemics (N = 3, 10%), antihypertensives (N = 3, 10%), proton pump inhibitor (N = 1, 3%), (N = 28, 93%) were treated with psychotropic medication: antidepressants (N = 27, 90%), mood stabilizers (N = 7, 23%), anxiolytics (N = 26, 87%), hypnotics (N = 22, 73%), and antipsychotics (N =14, 47%). | |||||||
| Avetyan et al. [65] | April 2019 | Armenia | Case-control study | Control | 49 | 43.5 (9.4) | 49 (100%) | NA | NA | - | NA | RLT, qPCR, Leukocytes | SCID-I, CAPS | Comparison of LTL in diseased and healthy subjects showed that PTSD patients had 1.5 times shorter average LTL than controls. |
| Case | 41 | 46.4 (7.63) | 41 (100%) | NA | ||||||||||
| Kim et al. [66] | June 2017 | South Korea | Cross-sectional | Control —High combat exposure | 11 | 62.82 (5.74) | 11 (100%) | Education (years) 9.45 (4.28) Socioeconomic status: High/Medium/Low, n 2/5/4 AUDIT score 6.73 (7.56) Heavy smoker: Yes/No, n 8/3 | NA | - | 20.8% psychoactive medications | RLT, qPCR, Leukocytes | CAPS, CES, AUDIT | As a whole, no significant difference in telomere length was found between PTSD and non-PTSD groups. In linear regression analysis stratified by trauma levels, among veterans exposed to severe combat, PTSD status, antidepressant use, and education level affected telomere length. |
| light-to-moderate combat exposure | 109 | 62.95 (4.23) | 109 (100%) | Education (years) 10.56 (3.03) Socioeconomic status: High/Medium/Low, n 18/50/41 AUDIT score 6.71 (7.67) Heavy smoker: Yes/No, n 65/44 | ||||||||||
| Case—High combat exposure | 34 | 63.38 (3.13) | 34 (100%) | Education (years) 10.35 (3.28) Socioeconomic status: High/Medium/Low, n 7/14/13 AUDIT score 13.09 (10.77) Heavy smoker: Yes/No, n 20/14 | 71.3% psychoactive medications | |||||||||
| light-to-moderate combat exposure | 88 | 62.84 (3.50) | 88 (100%) | Education (years) 10.38 (2.63) Socioeconomic status: High/Medium/Low, n 18/41/29 AUDIT score 11.01 (10.97) Heavy smoker: Yes/No, n 41/47 | ||||||||||
| Solomon et al. [54] | July 2017 | Israel | Cohort | Control: Resilient | 47 | 63.6 (3.7) | NA | Years of education (M, SD): 14, 3.7, Physical exercising regularly: 62 (62.6%) Smoking on a regular basis: 21 (21.2%) | NA | Depression | NA | mTL, Southern blot, Leukocytes | PTSD-I, DSM-IV | PTSD trajectories were not implicated in telomere length. |
| Case 1: chronic PTSD | 5 | |||||||||||||
| Case 2: delayed PTSD | 46 | |||||||||||||
| Case 3: recovered PTSD | 1 | |||||||||||||
| Zhang et al. [67] | November 2013 | USA | Cross-sectional | Control | 566 | 29.2 ± 7.3 | 412 (72.79%) | NA | 65.3% were White, 13.9% were Black, 7.8% were Asian or Pacific Islander, and 12.9% were American Indian or Alaskan Native | - | NA | RTL, RT-PCR, Leukocytes | PCL, DSMI-IV, SLE | Participants with PTSD had a lower relative T/S ratio than non-PTSD control subjects. This remained true when PTSD subjects were compared with age-matched non-PTSD controls. |
| Case | 84 | 76 (90.47%) | NA | 75.0% were White, 10.2% were Black, 4.5% were Asian or Pacific Islander, and 10.2% were American Indian or Alaskan Native |
| Date of Publication | Country | Study Design | Patients | Age, Years, Mean (SD) | Sex, n (%) Male | Main Comorbidities/ Lifestyle Factors Associated | Race/Ethnicity | Other Associated Psychiatric Diseases in This Study | Medication | Telomere Measurement and Tissue | Measurement of Psychiatric Disorder | Level of Anxiety | Telomere Lenght | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Verhoeven et al. [68] | January 2018 | The Netherlands | Case control study | Control | 582 | 41.7 (14.8) | 230 (39.5%) | Obesity (13.2%), former smoker (35.6%), current smoker (26.8%), moderate drinker (78.2%), heavy drinker (11.5). Years of education, mean (SD): 12.9 (3.2) | NA | Agoraphobia, panic disorder, socialphobia, generalised anxiety disorder | Antidepressant use, % Tricyclic antidepressant (0.2) Selective serotonin reuptake inhibitor (0.5), other antidepressant (0.2). Benzodiazepine use, %: (0.5) | mTL, qPCR, Leukocytes | BAI | BAI, mean (SD): 2.9 (2.9) | Patients with current—but not remitted—anxiety disorder had shorter telomere length, suggesting a process of accelerated cellular ageing, which in part may be reversible after remission. |
| Case 1—Remitted anxiety | 459 | 43.6 (12.7) | 131 (28.5%) | Obesity (17%), former smoker (39.8%), current smoker (33.8%), moderate drinker (73.2%), heavy drinker (11.8%). Years of education, mean (SD): 12.4 (3.4) | Antidepressant use, % Tricyclic antidepressant (2.4) Selective serotonin reuptake inhibitor (16.3), other antidepressant (3.5). Benzodiazepine use, %: (3.9) | BAI, mean (SD): 8.9 (7.3) | |||||||||
| Case 2—Current anxiety group | 1283 | 41.3 (12.4) | 412(32.1%) | Obesity (18.4%), former smoker (29.6%), current smoker (45.6%), moderate drinker (65%), heavy drinker (13.1%). Years of education, mean (SD): 11.6 (3.3) | Antidepressant use, % Tricyclic antidepressant (4.4) Selective serotonin reuptake inhibitor (26.8), other antidepressant (9.1). Benzodiazepine use, %: (13.1) | BAI, mean (SD): 18.5 (10.8) | |||||||||
| Groer M et al. [69] | December 2019 | USA | Cross-sectional | 97 | 29.6 (6.3) | 0 | Smoking (n = 2), more than an hour of exercise per week (n = 5), BMI: 28.8 (5.9). Completed completion or postgraduate education preparation (54%) | The sample was 76% Caucasian (39% of whom were of Hispanic origin) and 15% African American. The remaining 9% were Asian or other racial categories | Depression | NA | RTL, qPCR, DNA was extracted from PBMCs with DNeasy Blood and Tissue Ki | POMS, PSS | NA | There were no statistically significant relationships between TL and demographic factors, stress, depression, or TPO status. There were significant negative correlations between TL and anxiety and a trend for a relationship between TL and IL-6 levels. IL-6 levels were significantly, positively associated with negative moods.Higher anxiety scores and inflammation were associated with shorter TL. Inflammation was related to anxiety and other dysphoric moods and was marginally associated with shorter TLs. | |
| Schaakxs et al. [71] | April 2015 | The Netherlands | cross-sectional study | Control | 128 | 70.1 (7.2) | 49 (38.3) | Cigarette years, median: 170.00 (598.75); moderate drinker, n (%): 99 (77.3); heavy drinker, n (%): 12 (9.4);years of education, mean (SD): 12.5 (3.5); Obesity, n (%): 26 (20.3%); physical activity, median (IQR): 2.61 (2.96) | NA | MDD, n (%): 249 (70.1); dysthymia, n (%): 6 (1.7); Co-morbid MDD and dysthymia, n (%): 92 (25.9%) | - | mTL, qPCR, Leukocyte | CIDI, IDS, BAI, AS, CTI | NA | Mean TL did not differ between depressed persons (bp (SD): 5035 (431)) and never-depressed (bp (SD): 5057 (729)) comparisons. Further, TL was not associated with severity, duration, and age at onset of depression; comorbid anxiety disorders; anxiety symptoms; apathy severity; antidepressant use; benzodiazepine use; cognitive functioning; and childhood trauma |
| Case | 355 | 70.6 (7.4) | 120 (33.8) | Cigarette years, median (IQR): 100.00 (435.00); moderate drinker, n (%): 200 (56.3); heavy drinker, n (%): 15 (4.2); years of education, mean (SD): 10.3 (3.4); Obesity, n (%): 60 (16.9%); physical activity, median (IQR): 1.60 (2.96) | TCA, n (%): 78 (22.0); SSRI, n (%): 94 (26.6); Other antidepressants, n (%): 101 (28.5); Benzodiazepine use, n (%): 141 (39.7) | ||||||||||
| Wang et al. [34] | April 2017 | Sweden | Case-control study | Control | 320 | 44.6 (12.5) | 197 (61.5%) | Smokers—NA | NA | Depression and stress and adjustment disorders | mTL, qRT-PCR, Leukocyte | PHQ-9, HADS-A/HADS-D, MADRS-S | Telomere length, as compared with healthy controls, is shortened in patients with depression, anxiety and stress and adjustment disorders | ||
| Case | 181 | 41.9 (11.1) | 22 (12.15%) | Smokers (n = 27, 15%) | Antidepressant use (n = 63, 35%) | Baselines—Median score (IQR): MADRS-S: 20 (14–25); HAD-D: 8 (6–11); HAD-A: 12 (9–15); PHQ-9: 13 (9–17) | |||||||||
| Tyrka et al. [18] | January 2016 | USA | Case-Control study | Control | 113 | 28.5 (9.2) | 50 (44.2%) | Smokers (8.3%) | White (82.3%) | Adversities, depression, PTSD | NA | mTL, qPCR,, Leukocytes | SCID, STAI, PSS, CD-RISC | Significantly higher mtDNA copy numbers and shorter telomeres were seen in individuals with major depression, depressive disorders, and anxiety disorders, as well as those with parental loss and childhood maltreatment. | |
| Case 1—Adverity with no psychiatric disorder | 66 | 31.3 (11.1) | 26 (39.4%) | Smokers (7.8%) | White (80.3%) | ||||||||||
| Case 2—Psichyatric disorder with no adversity | 39 | 30.7 (10.4) | 15 (38.5%) | Smokers (7.7%) | White (92.3%) | MDD (n = 6), depressive (n = 7) | |||||||||
| Case 3—Adversity and psychiatric disorder | 72 | 34.8 (12.0) | 22 (30.6%) | Smokers (17.1%) | White (81.9%) | MDD (n = 7), depressive (n = 18) | |||||||||
| Prelog M et al. [70] | June 2016 | Germany | Case-control study | Control | 129 | Female (n = 85): 36.8 (10.9) Male (n = 44): 34.1(10.8) | 44 (34.1%) | Not documented | NA | Depression (n = 46) | RTL, RT-qPCR, Leukocytes | SCID-I | Panic disorder (n = 129) | Relative telomere lengths (RTLs) were not different between patients and HC. However, within the patient group, smokers had significantly shorter telomeres (0.91 ± 0.30) compared to non-smokers (1.07 ± 0.37) (p = 0.018) and females (0.96 ± 0.34) had shorter telomeres than males (1.10 ± 0.32) | |
| Case | 129 | Female (n = 85): 36.9 (10.8) Male (n = 44): 34.1(11.7) | 44 (34.1%) | Smokers (n = 39) | Antidepressants, yes, n (%): 52 (40.3%) | ||||||||||
| B L Needham et al. [11] | September 2014 | USA | Cross-sectional study | Control—No anxiety | 952 | 29.3(5.8) | 413 (43.4%) | NA | Non-Hispanic white 485 (51%); African american 184 (19.3%); Mexican American 283 (29.7%) | - | Antidepressant use, n (%): 27 (2.8%) | mTL, qPCR, Leukocytes | CIDI | - | The primary finding from this study is that depressive and anxious symptomatology, overall, have no direct relationship with TL in young adulthood. Although associations did not vary by race/ethnicity, among women (but not men) past-year GAD/PD was associated with shorter TL. There was no direct effect of antidepressant medication use on TL, but among current users of antidepressants, those with past-year MD had shorter TL than those with no depression. To our knowledge, this is the first study to examine relationships between antidepressant medication use and depressive and anxious symptomology, as well as the first to examine variation in these relationships by race/ethnicity, in a nationally representative sample. |
| Case 2—GAD/PD or anxious affect | 212 | 30.0(6.0) | 94 (44.3%) | Non-Hispanic white 101 (47.6%); African american 43 (20.3%); Mexican American 68 (32.1%) | Antidepressant use, n(%): 25 (11.8%) | ||||||||||
| Hoen et al. [21] | August 2012 | Netherlands | Longitudinal study | Control | 980 | 53.7 (11.3) | F: 551 (56.22%) M: 465 (43.78%) | Smoking (n = 225; 77%), Alcohol consumption (n = 788; 80%), Sedentarism (n = 50; 52%) | NA | Depression | mTL, PCR, Leukocytes | CIDI | NA | No association was found between depressive disorders and shorter telomeres at follow-up. Anxiety disorders predicted shorter telomere length at follow-up in a general population cohort. | |
| Case | 97 | 51.3 (10.7) | F: 62 M: 36 | Smokers (n = 32; 65%); Alcohol consumption (n = 78; 80%), Sedentarism (n = 505; 52%) | Antidepressant use (n = 14; 15%) | ||||||||||
| Verhoeven et al. [26] | June 2016 | USA and The Netherlands | Longitudinal study | Baseline | 2936 | 41.8 (13.1) | 986 (33.6%) | Former Smokers (n = 975), Current Smokers (n = 1.136); Mild-moderate drinker (n = 2064), Heavy drinker (n = 373) | NA | Current Depressive diagnosis disorder (n = 389, 23.3%), Control (n = 644, 21.9%), Remitted (n = 620, 21.1%) | Tricyclic antidepressants (n = 79, 2.7%); Selective serotonin reuptake inhibitor (n = 502, 17.1), Other antidepressants (n = 164, 5.6%) | RTL, qPCR, Leukocytes | CIDI | Current anxiety disorder (n = 535, 32.0%) | Robust associations of depressive and anxiety disorders with shorter telomeres, but interestingly, it did not demonstrate that depressive and anxiety disorders and LTL change together over time, suggesting the absence of a direct within-person relationship. Short LTL is suggested to be either a long-term consequence or an underlying vulnerability factor for depressive or anxiety disorders |
| Year 6 | 1883 | 48.6 (12.9) | 641 (34.6%) | Former Smokers (n = 797), Current Smokers (n = 531); Mild-moderate drinker (n = 1367), Heavy drinker (n = 186) | Current Depressive diagnosis disorder (n = 159, 30.1%), Control (n = 440, 23.4%), Remitted (n = 915, 48.6%) | Tricyclic antidepressants (n = 56, 3.0%); Selective serotonin reuptake inhibitor (n = 224, 11.9), Other antidepressants (n = 104, 5.5%) | Current anxiety disorder (n = 190, 36.0%) | ||||||||
| Chae et al. [31] | September 2015 | USA | Cross-sectional study | 92 | 43.86 (5.73) | 92 (100%) | Education, n (%): high school or less 38 (41.3), some college or more 54 (58.7). Work status, n (%): working 42 (45.7), unemployed 50 (54.4). Smoking status, n (%): noncurrent 41 (44.6), current 51 (55.4) Health conditions, mean (SD): 1.73 (1.89) | African American | - | Current doctor-prescribed medication use—Yes n (%): 29 (31.5) | mTL, qPCR, Leukocytes | CES-D, HADS | Anxiety, mean (SD): 5.02 (3.86) | Controlling for sociodemographic factors, greater anxiety symptoms were associated with shorter LTL | |
| Starnino et al. [42] | October 2016 | Canada | Cohort study | 132 | 45.34 (11.16) | 54 (40,9%) | Glasses of alcohol/week, mean (SD): 3.84 (5.38); Smoker n (%): 19(14%), BMI: 25.35 (5.00) | NA | Depression | - | RTL, qPCR, Leukocyte | BDI-II, BAI, CRP, MCSD, CMHo | Beck Depression Inventory-II, mean (SD): 7.64 (8.21) | Shorter TL was observed among individuals high in defensiveness and depressive symptoms, as well as in those with less hostility and anxiety. Telomere length is associated with psychological burden though the direction of effect differs depending on the psychological variables under study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pousa, P.A.; Souza, R.M.; Melo, P.H.M.; Correa, B.H.M.; Mendonça, T.S.C.; Simões-e-Silva, A.C.; Miranda, D.M. Telomere Shortening and Psychiatric Disorders: A Systematic Review. Cells 2021, 10, 1423. https://doi.org/10.3390/cells10061423
Pousa PA, Souza RM, Melo PHM, Correa BHM, Mendonça TSC, Simões-e-Silva AC, Miranda DM. Telomere Shortening and Psychiatric Disorders: A Systematic Review. Cells. 2021; 10(6):1423. https://doi.org/10.3390/cells10061423
Chicago/Turabian StylePousa, Pedro A., Raquel M. Souza, Paulo Henrique M. Melo, Bernardo H. M. Correa, Tamires S. C. Mendonça, Ana Cristina Simões-e-Silva, and Débora M. Miranda. 2021. "Telomere Shortening and Psychiatric Disorders: A Systematic Review" Cells 10, no. 6: 1423. https://doi.org/10.3390/cells10061423
APA StylePousa, P. A., Souza, R. M., Melo, P. H. M., Correa, B. H. M., Mendonça, T. S. C., Simões-e-Silva, A. C., & Miranda, D. M. (2021). Telomere Shortening and Psychiatric Disorders: A Systematic Review. Cells, 10(6), 1423. https://doi.org/10.3390/cells10061423