
The Origin of the Cyathea delgadii Sternb. Somatic Embryos Is Determined by the Developmental State of Donor Tissue and Mutual Balance of Selected Metabolites


 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material, Culture Conditions, and Evaluation of the SE Efficiency
2.2. Microscopic Examinations
2.2.1. High-Resolution Optical 3D Microscopy
2.2.2. Light Microscopy
2.2.3. Transmission Electron Microscopy (TEM)
2.2.4. Environmental Scanning Electron Microscopy (ESEM)
2.2.5. Fluorescence Microscopy
2.3. Profiling of Plant Metabolites
2.3.1. Preparation of Plant Samples
2.3.2. Soluble Carbohydrates
2.3.3. Starch
2.3.4. Fructooligosaccharides
2.3.5. HPLC Analyses of Neutral Sugars
2.3.6. UHPLC-MS/MS Profiling of Plant Hormones and Related Compounds
2.3.7. Phenolic Acids
2.3.8. Polyamines
2.3.9. Amino Acids
2.3.10. Ethylene
2.4. Statistical Analyses
3. Results
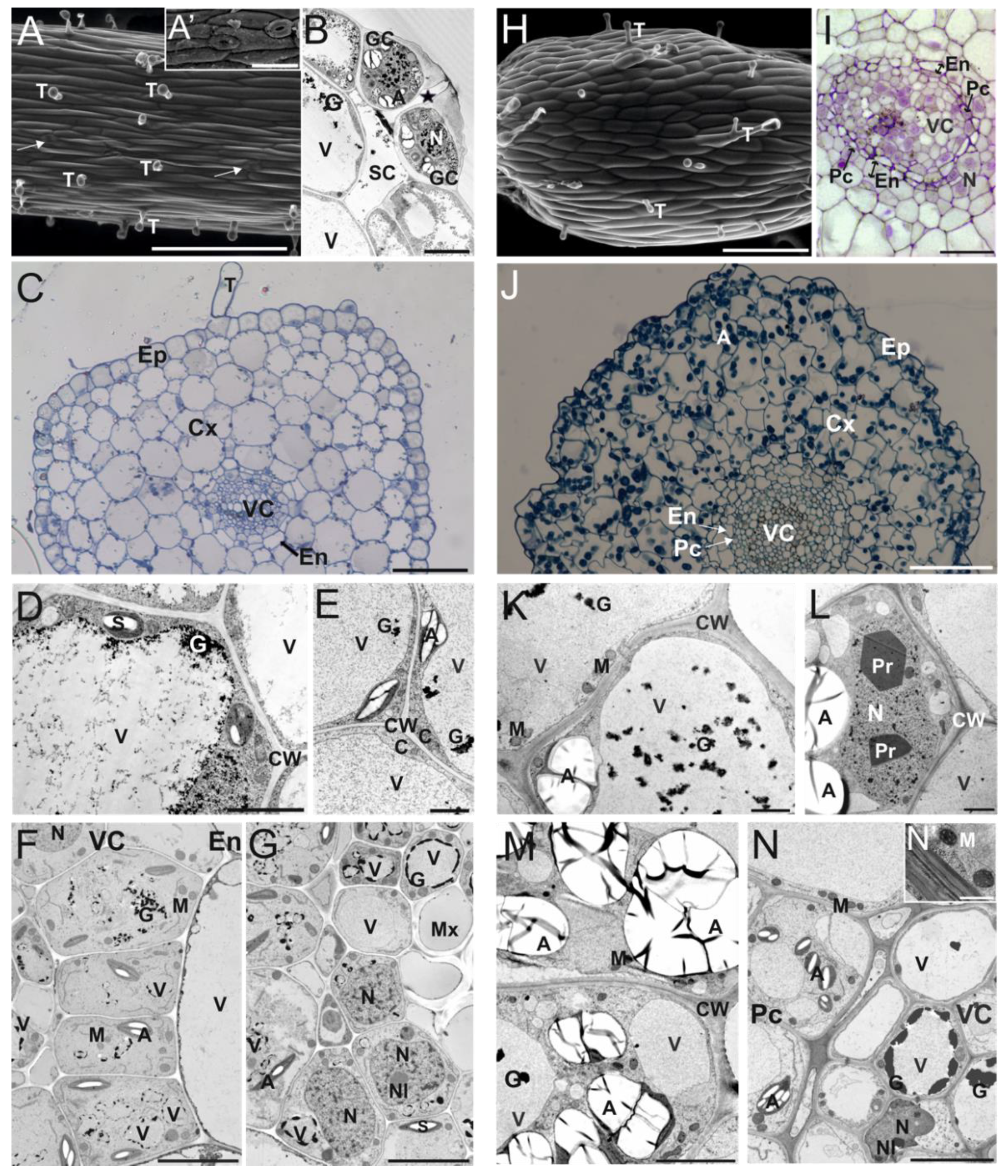
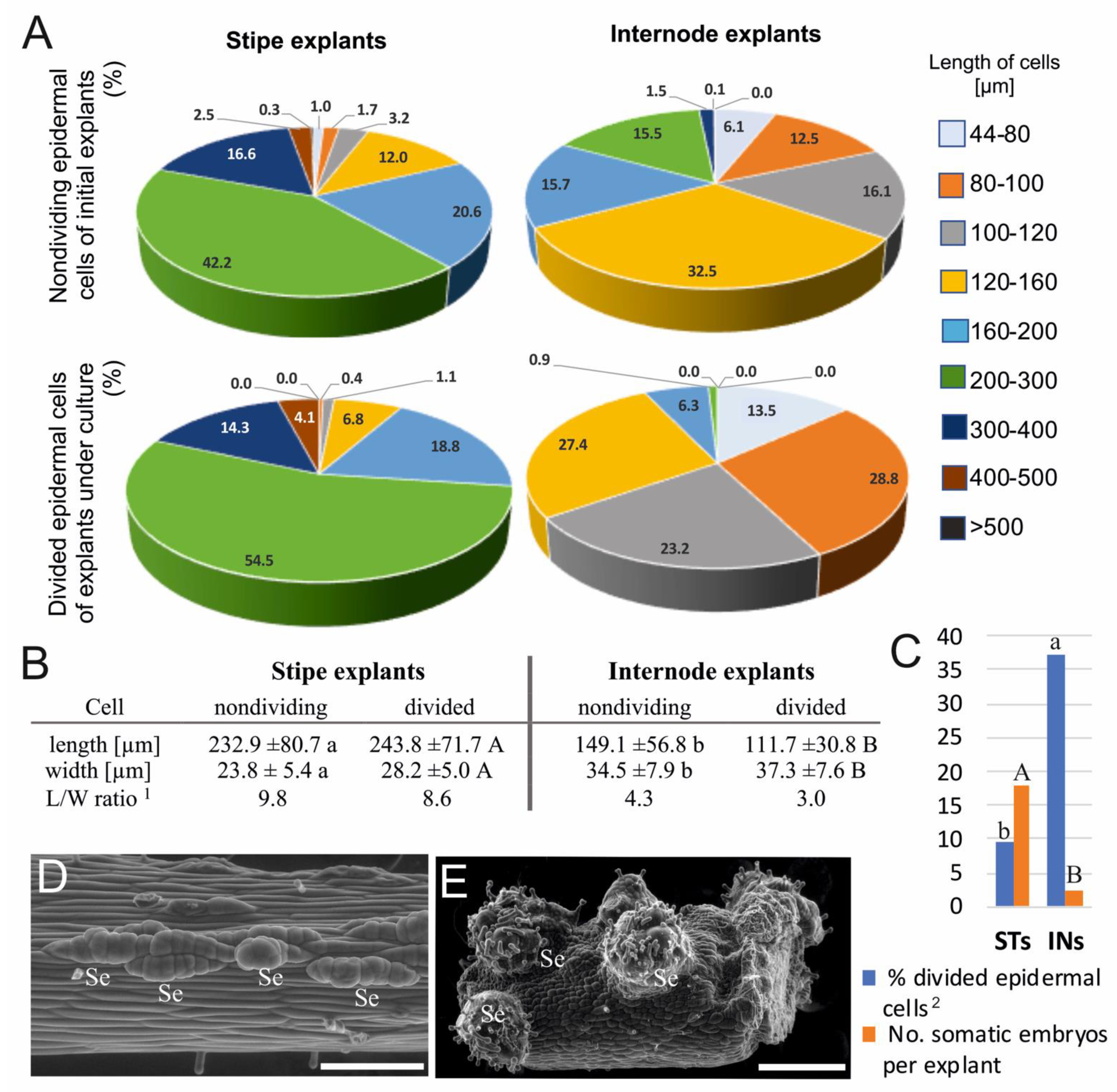
3.1. Cytomorphological Differences between Stipe and Internode Explants
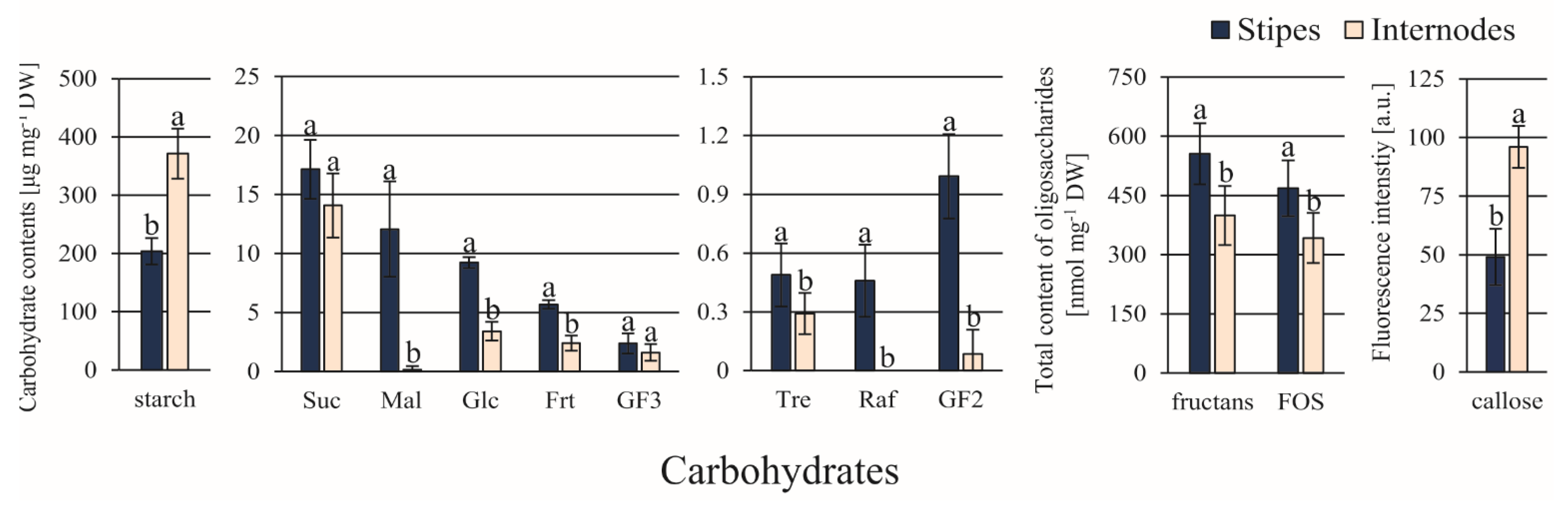
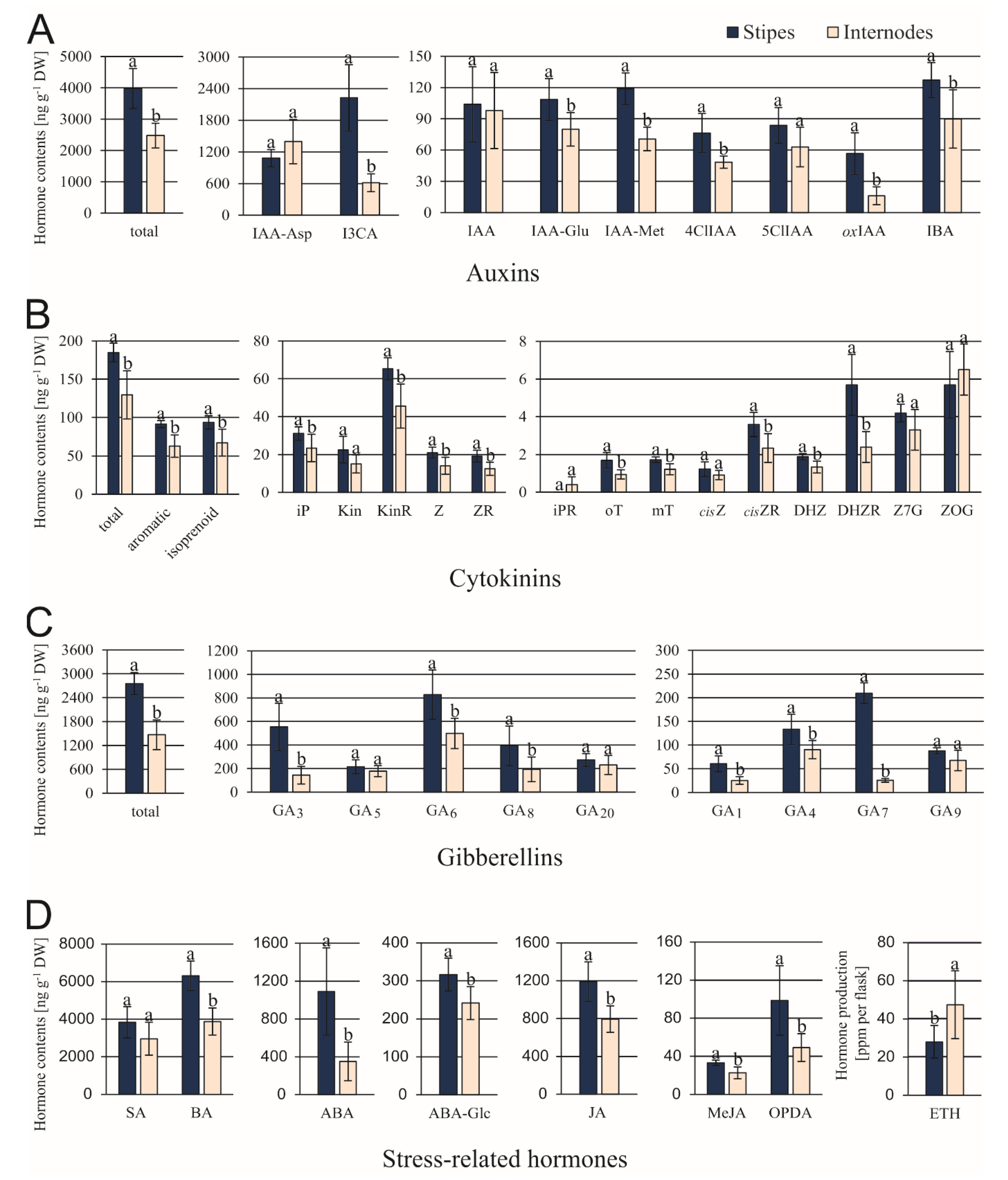
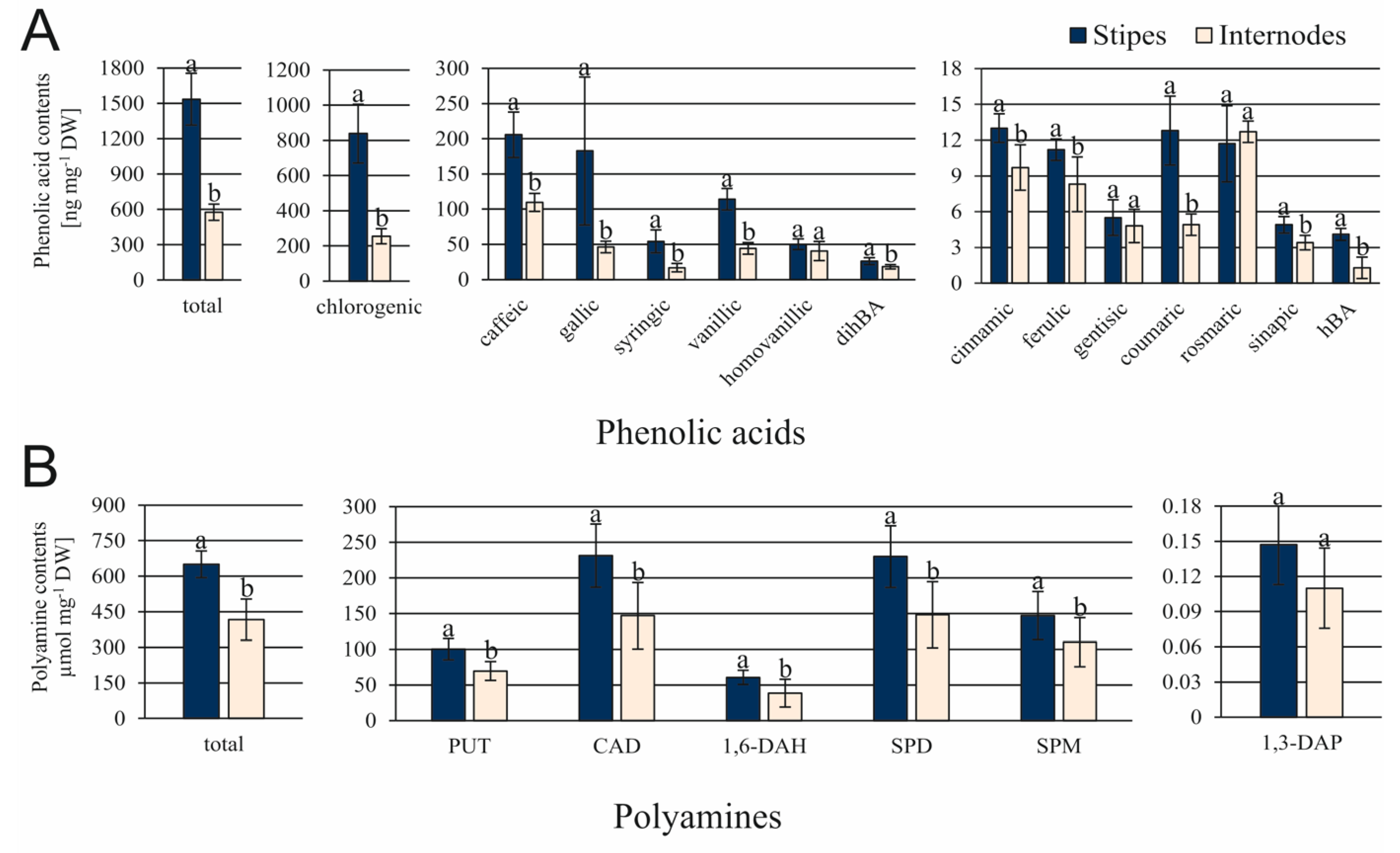
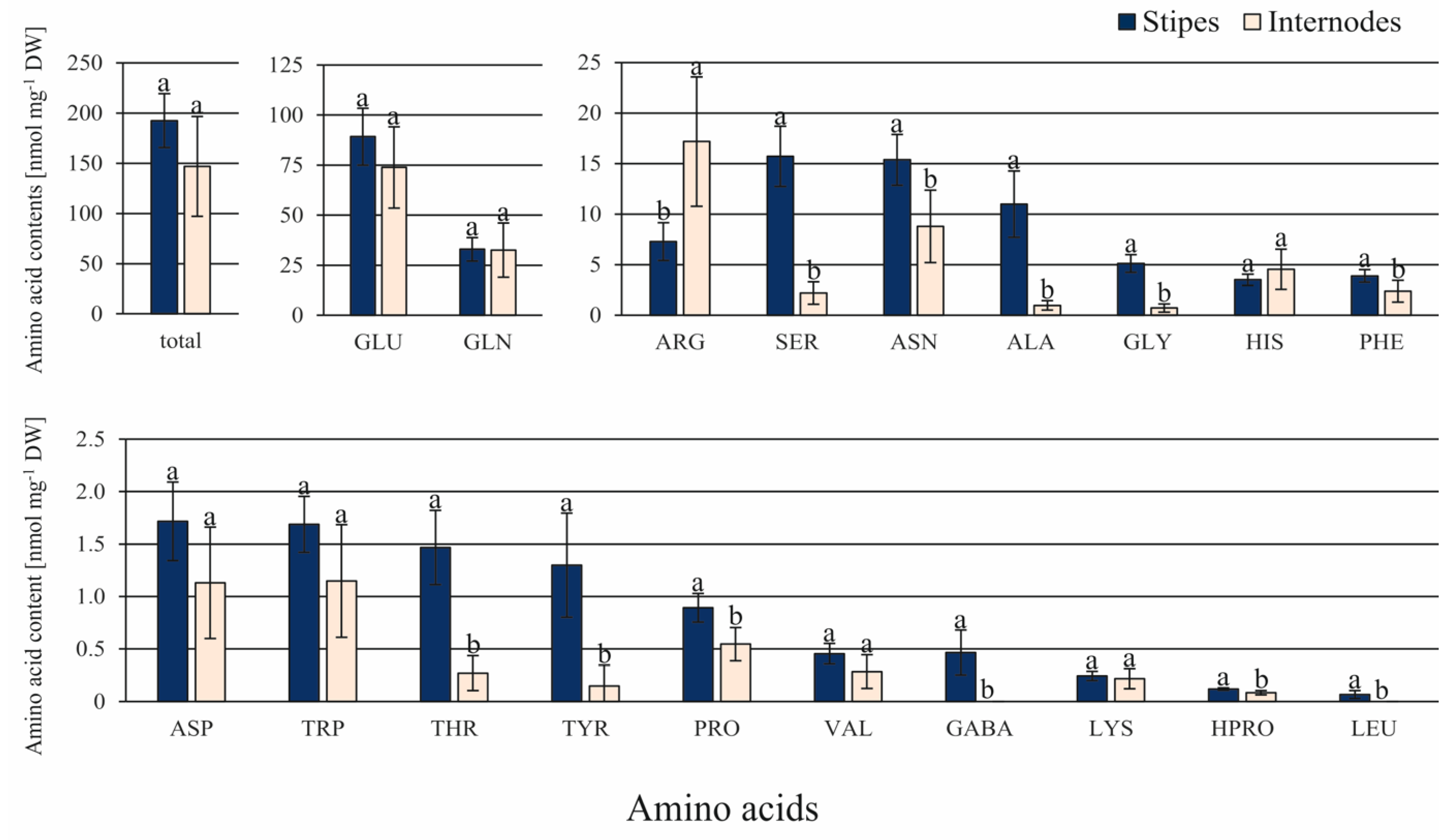
3.2. Differences in the Content of Endogenous Compounds
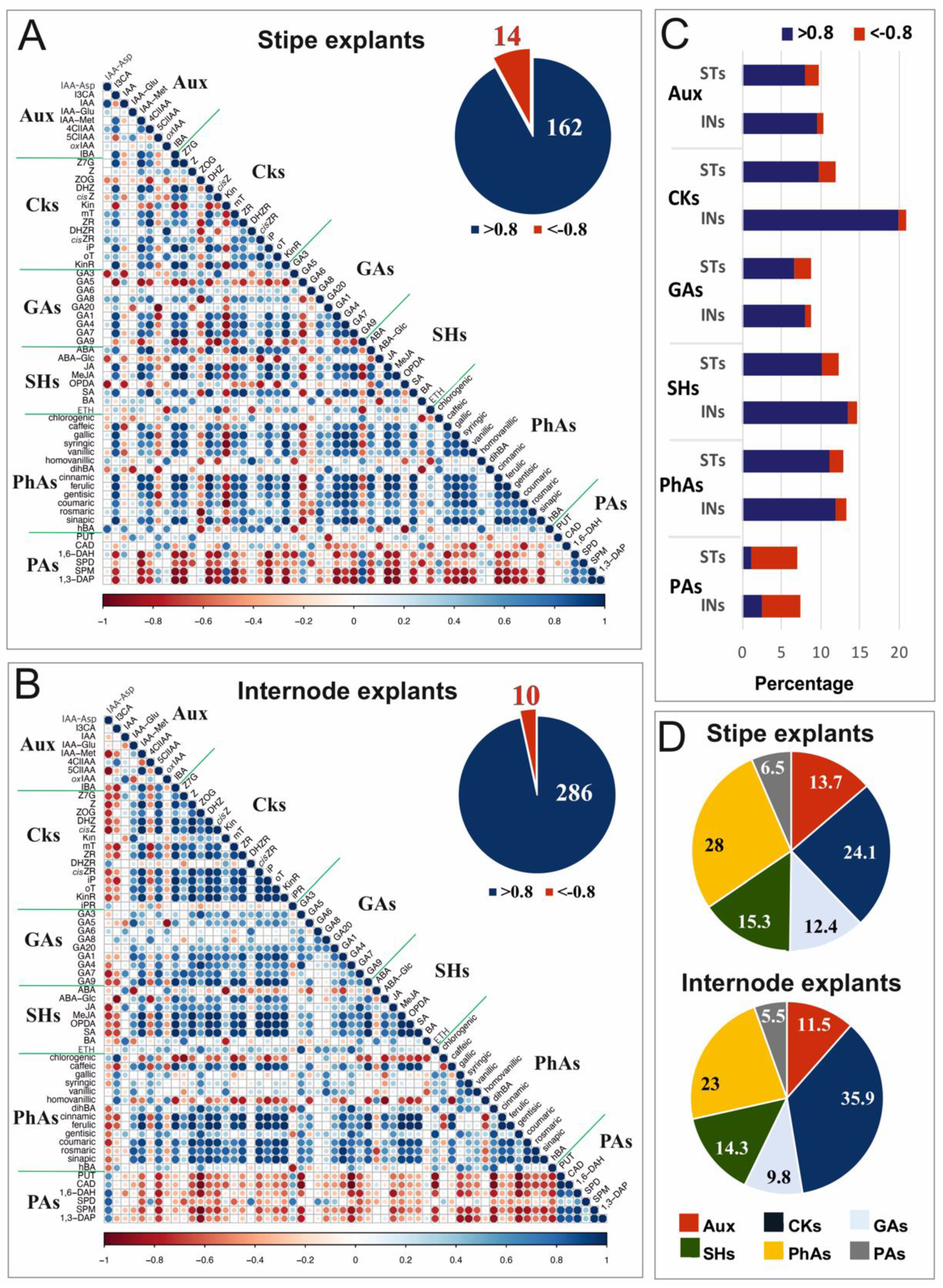
3.3. Analysis of Relationships between Compounds in Stipe and Internode Explants
4. Discussion
4.1. Effect of Cell Size on Embryogenic Competence
4.2. Relationship between Carbohydrates and Somatic Embryo Origin
4.3. Relationship between Phytohormone and Other Compounds Status versus Somatic Embryo Origin
5. Conclusions
- (1)
- High contents of ABA, biologically active gibberellins, and phenolic acids (indicate a more advanced developmental status of explant in which only a limited number of cells is competent to enter embryogenic pathway);
- (2)
- High content of Mal and GF2 (help to maintain the osmotic homeostasis in strongly vacuolated cells);
- (3)
- Numerous positive correlations between ABA and cytokinins (suggest that cell divisions preceding unicellular pathway of somatic embryo formation are controlled by the former).
- (1)
- High contents of storage substances (as starch providing carbon backbones and energy and ARG as a source of organic nitrogen, which facilitate quick and extensive cell divisions);
- (2)
- High content of callose (indicates a more meristematic character of the explant cells);
- (3)
- Increased ethylene production (involved in local induction of auxin biosynthesis).
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 1,3-DAP | 1,3-Diaminopropane |
| 1,6-DAH | 1,6-Diaminohexane |
| 4ClIAA | 4-Chloroindole-3-acetic-acid |
| 5ClIAA | 5-Chloroindole-3-acetic acid |
| ABA | Abscisic acid |
| ABA-Glc | Abscisic acid glucosyl ester |
| ALA | Alanine |
| ARG | Arginine |
| ASN | Asparagine |
| ASP | Aspartic acid |
| AVA | 5-Aminovaleric acid |
| BA | Benzoic acid |
| CAD | Cadaverine |
| cisZ | cis-Zeatin |
| cisZR | cis-Zeatin riboside |
| DHZ | Dihydrozeatin |
| DHZR | Dihydrozeatin riboside |
| dihBA | 3,4-Dihydroxobenzoic acid |
| DW | Dry weight |
| ETH | Ethylene |
| FOS | Fructooligosaccharides |
| Frt | Fructose |
| GABA | Gamma-aminobutyric acid |
| GA | Gibberellin |
| GF2 | 1-Kestose |
| GF3 | Nystose |
| GF4 | Fructosylnystose |
| Glc | Glucose |
| GLN | Glutamine |
| GLU | Glutamic acid |
| GLY | Glycine |
| hBA | p-Hydroxobenzoic acid |
| HIS | Histidine |
| HPRO | trans-4-Hydroxyproline |
| I3CA | Indole-3-carboxylic acid |
| IAA | Indole-3-acetic acid |
| IAA-Asp | Indole-3-acetyl-l-aspartic acid |
| IAA-Glu | Indole-3-acetyl-l-glutamic acid |
| IAA-Met | Indole-3-acetic acid methyl ester |
| IBA | Indole-3-butyric acid |
| ILE | Isoleucine |
| IN | Internode |
| iP | N6-Isopentenyladenine |
| iPR | N6-Isopentenyladenosine |
| JA | Jasmonic acid |
| Kin | Kinetin |
| KinR | Kinetin riboside |
| LEU | Leucine |
| LYS | Lysine |
| Mal | Maltose |
| MeJA | Jasmonic acid methyl ester |
| MET | Methionine |
| MS | Murashige and Skoog’s medium |
| mT | meta-Topolin |
| OPDA | 12-Oxo-phytodienoic acid |
| oT | ortho-Topolin |
| oxIAA | Oxindole-3-acetic acid |
| PHE | Phenylalanine |
| PRO | Proline |
| PUT | Putrescine |
| Raf | Raffinose |
| SA | Salicylic acid |
| SE | Somatic embryogenesis |
| SER | Serine |
| SPD | Spermidine |
| SPM | Spermine |
| ST | Stipe |
| Suc | Sucrose |
| THR | Threonine |
| Tre | Trehalose |
| TRP | Tryptophan |
| TYR | Tyrosine |
| VAL | Valine |
| Z | trans-Zeatin |
| Z7G | trans-Zeatin-7-glucoside |
| ZOG | trans-Zeatin-o-glucoside |
| ZR | trans-Zeatin riboside |
References
- Ikeuchi, M.; Ogawa, Y.; Iwase, A.; Sugimoto, K. Plant regeneration: Cellular origins and molecular mechanisms. Development 2016, 143, 442–451. [Google Scholar] [CrossRef] [PubMed]
- Williams, E.G.; Maheswaran, G. Somatic embryogenesis: Factors influencing coordinated behaviour of cells as an embryogenic group. Ann. Bot. 1986, 57, 443–462. [Google Scholar] [CrossRef]
- Yang, J.L.; Seong, E.S.; Kim, M.J.; Ghimire, B.K.; Kang, W.H.; Yu, C.Y.; Li, C.H. Direct somatic embryogenesis from pericycle cells of broccoli (Brassica oleracea L. var. italica) root explants. Plant Cell Tiss. Organ Cult. 2010, 100, 49–58. [Google Scholar] [CrossRef]
- Grzyb, M.; Mikuła, A. Explant type and stress treatment determine the uni- and multicellular origin of somatic embryos in the tree fern Cyathea delgadii Sternb. Plant Cell Tissue Organ. Cult. 2019, 136, 221–230. [Google Scholar] [CrossRef]
- Bhusare, B.P.; John, C.K.; Bhatt, V.P.; Nikam, T.D. Induction of somatic embryogenesis in leaf and root explants of Digitalis lanata Ehrh.: Direct and indirect method. S. Afr. J. Bot. 2020, 130, 356–365. [Google Scholar] [CrossRef]
- Centeno, M.L.; Rodríguez, R.; Feito, I.; Fernandez, B. Relationship between endogenous auxin and cytokinin levels and morphogenic responses in Actinidia deliciosa tissue cultures. Plant Cell Rep. 1996, 16, 58–62. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Q.; Han, Z.; Kang, X. Adventitious shoot regeneration from leaf, petiole and root explants in triploid (Populus alba × P. glandulosa) × P. tomentosa. Plant Cell Tissue Organ. Cult. 2019, 138, 121–130. [Google Scholar] [CrossRef]
- Mannucci, A.; Mariotti, L.; Castagna, A.; Santin, M.; Trivellini, A.; Reyes, T.H.; Mensuali-Sodi, A.; Ranieri, A.; Quartacci, M.F. Hormone profile changes occur in roots and leaves of Micro-Tom tomato plants when exposing the aerial part to low doses of UV-B radiation. Plant Physiol. Biochem. 2020, 148, 291–301. [Google Scholar] [CrossRef]
- Sharma, P.; Rajam, M.V. Spatial and temporal changes in endogenous polyamine levels associated with somatic embryogenesis from different regions of hypocotyl of eggplant (Solanum melongena L.). J. Plant Physiol. 1995, 146, 658–664. [Google Scholar] [CrossRef]
- Valdés, A.E.; Ordás, R.J.; Fernández, B.; Centeno, M.L. Relationships between hormonal contents and the organogenic response in Pinus pinea cotyledons. Plant Physiol. Biochem. 2001, 39, 377–384. [Google Scholar] [CrossRef]
- Hu, W.; Fagundez, S.; Katin-Grazzini, L.; Li, Y.; Li, W.; Chen, Y.; Wang, X.; Deng, Z.; Xie, S.; McAvoy, R.J.; et al. Endogenous auxin and its manipulation influence in vitro shoot organogenesis of citrus epicotyl explants. Hortic. Res. 2017, 4, 17071. [Google Scholar] [CrossRef]
- Zhang, Y.F.; Zhou, J.H.; Wu, T.; Cao, J.S. Shoot regeneration and the relationship between organogenic capacity and endogenous hormonal contents in pumpkin. Plant Cell Tiss. Organ Cult. 2008, 93, 323–331. [Google Scholar] [CrossRef]
- Grzyb, M.; Kalandyk, A.; Waligórski, P.; Mikuła, A. The content of endogenous hormones and sugars in the process of early somatic embryogenesis in the tree fern Cyathea delgadii Sternb. Plant Cell Tissue Organ. Cult. 2017, 129, 387–397. [Google Scholar] [CrossRef][Green Version]
- Centeno, M.L.; Rodríguez, R.; Berros, B.; Rodríguez, A. Endogenous hormonal content and somatic embryogenic capacity of Corylus avellana L. cotyledons. Plant Cell Rep. 1997, 17, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Dziurka, K.; Dziurka, M.; Warchoł, M.; Czyczyło-Mysza, I.; Marcińska, I.; Noga, A.; Kapłoniak, K.; Skrzypek, E. Endogenous phytohormone profile during oat (Avena sativa L.) haploid embryo development. In Vitro Cell. Dev. Biol. Plant 2019, 55, 221–229. [Google Scholar] [CrossRef]
- Takamura, T.; Tanaka, M. Somatic embryogenesis from the etiolated petiole of Cyclamen (Cyclamen persicum Mill.). Plant Cell Tissue Organ. Cult. 1996, 13, 43–48. [Google Scholar] [CrossRef]
- Grzyb, M.; Kalandyk, A.; Mikuła, A. Effect of TIBA, fluridone and salicylic acid on somatic embryogenesis and endogenous hormone and sugar contents in Cyathea delgadii Sternb. Acta Physiol. Plant 2018, 40, 1. [Google Scholar] [CrossRef]
- Limanton-Gravet, A.; Sotta, B.; Brown, S.; Jullien, M. Analysis of habituated embryogenic lines in Asparagus officinalis L.: Growth characterization, hormone content and ploidy level of calli and regenerated plants. Plant Sci. 2000, 160, 15–26. [Google Scholar] [CrossRef]
- Mikuła, A.; Pożoga, M.; Tomiczak, K.; Rybczyński, J.J. Somatic embryogenesis in ferns: A new experimental system. Plant Cell Rep. 2015, 34, 783–794. [Google Scholar] [CrossRef] [PubMed]
- Mikuła, A.; Pożoga, M.; Grzyb, M.; Rybczyński, J.J. An unique system of somatic embryogenesis in the tree fern Cyathea delgadii Sternb.—The importance of explant type, and physical and chemical factors. Plant Cell Tissue Organ. Cult. 2015, 123, 467–478. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Kaźmierczak, A. Cell number, cell growth, antheridiogenesis, and callose amount is reduced and atrophy induced by deoxyglucose in Anemia phyllitidis gametophytes. Plant Cell Rep. 2008, 27, 813–821. [Google Scholar] [CrossRef]
- Hura, T.; Dziurka, M.; Hura, K.; Ostrowska, A.; Dziurka, K. Different allocation of carbohydrates and phenolics in dehydrated leaves of triticale. J. Plant Physiol. 2016, 202, 1–9. [Google Scholar] [CrossRef]
- Hura, T.; Dziurka, M.; Hura, K.; Ostrowska, A.; Dziurka, K.; Gadzinowska, J. Wheat and rye genome confer specific phytohormone profile features and interplay under water stress in two phenotypes of triticale. Plant Physiol. Biochem. 2017, 118, 494–509. [Google Scholar] [CrossRef] [PubMed]
- Gołębiowska-Pikania, G.; Dziurka, M.; Wąsek, I.; Wajdzik, K.; Dyda, M.; Wędzony, M. Changes in phenolic acid abundance involved in low temperature and Microdochium nivale (Samuels and Hallett) cross tolerance in winter triticale (×Triticosecale Wittmack). Acta Physiol. Plant 2019, 41, 38. [Google Scholar] [CrossRef]
- Hura, T.; Dziurka, M.; Hura, K.; Ostrowska, A.; Dziurka, K. Free and cell wall-bound polyamines under long-term water stress applied at different growth stages of ×Triticosecale Wittm. PLoS ONE 2015, 10, e0135002. [Google Scholar] [CrossRef] [PubMed]
- Schuster, R. Determination of amino acids in biological, pharmaceutical, plant and food samples by automated precolumn derivatization and high-performance liquid chromatography. J. Chromatogr. B 1988, 431, 271–284. [Google Scholar] [CrossRef]
- Woodward, C.; Henderson, J.W.; Wielgos, T. High-Speed Amino Acid Analysis (AAA) on 1.8 µm Reversed-Phase (RP) Columns. 2007. Available online: https://docplayer.net/25325234-High-speed-amino-acid-analysis-aaa-on-1-8-um-reversed-phase-rp-columns-application.html (accessed on 3 June 2021).
- Wei, T.; Simko, V. R Package “Corrplot”: Visualization of a Correlation Matrix. (Version 0.84). 2017. Available online: https://github.com/taiyun/corrplot (accessed on 20 September 2020).
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 20 September 2020).
- Willis, L.; Refahi, Y.; Wightman, R.; Landrein, B.; Teles, J.; Huang, K.C.; Meyerowitz, E.M.; Jönsson, H. Cell size and growth regulation in the Arabidopsis thaliana apical stem cell niche. Proc. Natl. Acad. Sci. USA 2016, 113, E8238–E8246. [Google Scholar] [CrossRef]
- Fransz, P.F.; Schel, J.H.N. Cytodifferentiation during the development of friable embryogenic callus of maize (Zea mays). Can. J. Bot. 1991, 69, 26–33. [Google Scholar] [CrossRef]
- Jones, T.J.; Rost, T.L. The developmental anatomy and ultrastructure of somatic embryos from rice (Oryza sativa L.) scutellum epithelial cells. Bot. Gaz. 1989, 150, 41–49. [Google Scholar] [CrossRef]
- Trigiano, R.N.; Gray, O.J.; Conger, B.V.; McDaniel, J.K. Origin of direct somatic embryos from cultured leaf segments of Dactylis glomerata. Bot. Gaz. 1989, 150, 72–77. [Google Scholar] [CrossRef]
- Canhoto, J.M.; Mesquita, J.F.; Cruz, G.S. Ultrastructural changes in cotyledons of pineapple guava (Myrtaceae) during somatic embryogenesis. Ann. Bot. 1996, 78, 513–521. [Google Scholar] [CrossRef][Green Version]
- Puigderrajols, P.; Mir, G.; Molinas, M. Ultrastructure of early secondary embryogenesis by multicellular and unicellular pathways in cork oak (Quercus suber L.). Ann. Bot. 2001, 87, 179–189. [Google Scholar] [CrossRef]
- Toonen, M.A.J.; De Vries, S.C. Initiation of somatic embryos from single cells. In Embryogenesis—The Generation of a Plant; Wang, T.L., Cuming, A., Eds.; Oxford, Bios Scientific Publishers: Oxford, UK, 1996; pp. 173–189. [Google Scholar]
- Eveland, A.L.; Jackson, D.P. Sugars, signalling, and plant development. J. Exp. Bot. 2011, 63, 3367–3377. [Google Scholar] [CrossRef] [PubMed]
- George, E.F. The components of plant tissue culture media II: Organic additions, osmotic and pH effects, and support systems. In Plant Propagation by Tissue Culture; George, E.F., Hall, M.A., de Klerk, G.J.D., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 115–173. [Google Scholar]
- Mikuła, A.; Tykarska, T.; Zielińska, M.; Kuraś, M.; Rybczyński, J.J. Ultrastructural changes in zygotic embryos of Gentiana punctata (L.) during callus formation and somatic embryogenesis. Acta Biol. Crac. Ser. Bot. 2004, 46, 109–112. [Google Scholar]
- Corredoira, E.; Valladares, S.; Martínez, M.T.; Vieitez, A.M.; San José, M.C. Somatic embryogenesis in Alnus glutinosa (L.) Gaertn. Trees 2013, 27, 1597–1608. [Google Scholar] [CrossRef]
- Mikuła, A.; Tykarska, T.; Kuraś, M.; Rybczyński, J.J. Somatic embryogenesis of Gentiana cruciata (L.): Histological and ultrastructural changes in seedling hypocotyl explant. In Vitro Cell. Dev. Biol. Plant 2005, 41, 686–694. [Google Scholar] [CrossRef]
- Stamp, J.A. Somatic embryogenesis in cassava. The anatomy and morphology of the regeneration process. Ann. Bot. 1987, 57, 451–459. [Google Scholar] [CrossRef]
- Martin, A.B.; Cuadrado, Y.; Guerra, H.; Gallego, P.; Hita, O.; Martin, L.; Dorado, A.; Villalobos, N. Differences in the contents of total sugars, reducing sugars, starch and sucrose in embryogenic and non-embryogenic calli from Medicago arborea L. Plant Sci. 2000, 154, 143–151. [Google Scholar] [CrossRef]
- Endress, V.; Barriuso, J.; Rupérez, P.; Martín, J.P.; Blázquez, A.; Villalobos, N.; Guerra, H.; Martín, L. Differences in cell wall polysaccharide composition between embryogenic and non-embryogenic calli of Medicago arborea L. Plant Cell Tissue Organ. Cult. 2009, 97, 323–329. [Google Scholar] [CrossRef]
- Guo, H.; Guo, H.; Zhang, L.; Fan, Y.; Fan, Y.; Zeng, F. SELTP-assembled battery drives totipotency of somatic plant cell. Plant Biotech. J. 2019, 17, 1188–1190. [Google Scholar] [CrossRef]
- Chen, X.Y.; Kim, J.Y. Callose synthesis in higher plants. Plant Signal. Behav. 2009, 4, 489–492. [Google Scholar] [CrossRef] [PubMed]
- Galatis, B.; Apostolakos, P. A new callose function. Plant Signal. Behav. 2010, 5, 1359–1364. [Google Scholar] [CrossRef] [PubMed]
- Brown, R.C.; Lemmon, B.E. Callose in cell division. In Chemistry, Biochemistry and Biology of (1→3)-β-Glucans and Related Polysaccharides; Basic, A., Fincher, G.B., Stone, B.A., Eds.; Academic Press: Amsterdam, The Netherlands, 2009; pp. 425–437. [Google Scholar]
- Nägele, T.; Heyer, A.G. Mathematical modelling for investigation of plant cold tolerance. In Molecular Approaches in Plant Abiotic Stress; Gaur, R.K., Sharma, P., Eds.; Taylor & Francis Group: Boca Raton, The Netherlands, 2013; pp. 193–208. [Google Scholar]
- Konrádová, H.; Gricová, M.; Lipavská, H. Cold-induced accumulation of raffinose family oligosaccharides in somatic embryos of Norway spruce (Picea abies). In Vitro Cell. Dev. Biol. Plant 2003, 39, 425–427. [Google Scholar] [CrossRef]
- Marszał-Jagacka, J.; Kromer, K. In vitro propagation of rare and endangered serpentine fern species. In Working with Ferns: Issues and Applications; Fernández, H., Kumar, A., Revilla, M.A., Eds.; Springer Science+Business Media: New York, NY, USA; London, UK; Dordrecht, The Netherlands; Heidelberg, Germany, 2011; pp. 149–164. [Google Scholar]
- Blanc, G.; Lardet, L.; Martin, A.; Jacob, J.L.; Carron, M.P. Differential carbohydrate metabolism conducts morphogenesis in embryogenic callus of Hevea brasiliensis (Müll. Arg.). J. Exp. Bot. 2002, 53, 1453–1462. [Google Scholar] [CrossRef]
- Scott, P.; Lyne, R.L. The effect of different carbohydrate sources upon the initiation of embryogenesis from barley microspores. Plant Cell Tissue Organ. Cult. 1994, 36, 129–133. [Google Scholar] [CrossRef]
- Bhandari, K.; Nayyar, H. Low temperature stress in plants: An overview of roles of cryoprotectants in defense. In Physiological Mechanisms and Adaptation Strategies in Plants under Changing Environment: Volume 1; Ahmad, P., Wani, M.R., Eds.; Springer Science + Business Media: New York, NY, USA, 2014; pp. 193–266. [Google Scholar]
- Su, Y.H.; Liu, Y.B.; Zhang, X.S. Auxin-cytokinin interaction regulates meristem development. Mol. Plant 2011, 4, 616–625. [Google Scholar] [CrossRef]
- Stahl, E.; Bellwon, P.; Huber, S.; Schlaeppi, K.; Bernsdorff, F.; Vallat-Michel, A.; Mauch, F.; Zeier, J. Regulatory and functional aspects of indolic metabolism in plant systemic acquired resistance. Mol. Plant 2016, 9, 662–681. [Google Scholar] [CrossRef] [PubMed]
- Sakakibara, H. Cytokinins: Activity, biosynthesis, and translocation. Annu. Rev. Plant Biol. 2006, 57, 431–449. [Google Scholar] [CrossRef]
- Schäfer, M.; Brütting, C.; Meza-Canales, I.D.; Großkinsky, D.K.; Vankova, R.; Baldwin, I.T.; Meldau, S. The role of cis-zeatin-type cytokinins in plant growth regulation and mediating responses to environmental interactions. J. Exp. Bot. 2015, 66, 4873–4884. [Google Scholar] [CrossRef]
- Hedden, P.; Thomas, S.G. Gibberellin biosynthesis and its regulation. Biochem. J. 2012, 444, 11–25. [Google Scholar] [CrossRef]
- Piotrowska, A.; Bajguz, A. Conjugates of abscisic acid, brassinosteroids, ethylene, gibberellins, and jasmonates. Phytochemistry 2011, 72, 2097–2112. [Google Scholar] [CrossRef] [PubMed]
- Prado, M.J.; Largo, A.; Domínguez, C.; González, M.V.; Rey, M.; Centeno, M.L. Determination of abscisic acid and its glucosyl ester in embryogenic callus cultures of Vitis vinifera in relation to the maturation of somatic embryos using a new liquid chromatography-ELISA analysis method. J. Plant Physiol. 2014, 171, 852–859. [Google Scholar] [CrossRef]
- Ivanova, A.; Velcheva, M.; Denchev, P.; Atanassov, A.; Van Onckelen, H.A. Endogenous hormone levels during direct somatic embryogenesis in Medicago falcata. Physiol. Plant 1994, 92, 85–89. [Google Scholar] [CrossRef]
- Jiménez, V.M.; Bangerth, F. Hormonal status of maize initial explants and of the embryogenic and non-embryogenic callus cultures derived from them as related to morphogenesis in vitro. Plant Sci. 2001, 160, 247–257. [Google Scholar] [CrossRef]
- Chater, C.C.C.; Oliver, J.; Casson, S.; Gray, J.E. Putting the brakes on: Abscisic acid as a central environmental regulator of stomatal development. New Phytol. 2014, 202, 376–391. [Google Scholar] [CrossRef]
- Bai, B.; Su, Y.H.; Yuan, J.; Zhang, X.S. Induction of somatic embryos in Arabidopsis requires local YUCCA expression mediated by the down-regulation of ethylene biosynthesis. Mol. Plant 2013, 6, 1247–1260. [Google Scholar] [CrossRef] [PubMed]
- Dempsey, M.A.; Klessig, D.F. How does the multifaceted plant hormone salicylic acid combat disease in plants and are similar mechanisms utilized in humans? BMC Biol. 2017, 15, 23. [Google Scholar] [CrossRef]
- Hosseini, S.S.; Mashayekhi, K.; Alizadeh, M.; Ebrahimi, P. Effect of salicylic acid on somatic embryogenesis and chlorogenic acid levels of carrot (Daucus carota cv. Nantes) explants. J. Ornam. Plant 2011, 1, 105–113. [Google Scholar]
- Reis, E.; Batista, M.T.; Canhoto, J.M. Effect and analysis of phenolic compounds during somatic embryogenesis induction in Feijoa sellowiana Berg. Protoplasma 2008, 232, 193–202. [Google Scholar] [CrossRef]
- Canhoto, J.M.; Cruz, G.S. Histodifferentiation of somatic embryos in cotyledons of pineapple guava (Feijoa sellowiana Berg). Protoplasma 1996, 191, 34–45. [Google Scholar] [CrossRef]
- Delalonde, M.; Barret, Y.; Coumans, M.P. Development of phenolic compounds in maize anthers (Zea mays) during cold pretreatment prior to androgenesis. J. Plant Physiol. 1996, 149, 612–616. [Google Scholar] [CrossRef]
- Martin, K.P.; Madassery, J. Direct and indirect somatic embryogenesis on cotyledon explants of Quassia amara L., an antileukaemic drug plant. In Vitro Cell. Dev. Biol. Plant 2005, 41, 54–57. [Google Scholar] [CrossRef]
- Altamura, M.M.; Torrigiani, P.; Falasca, G.; Rossini, P.; Bagni, N. Morphofunctional gradient in superficial and deep tissues along tobacco stem: Polyamine levels, biosynthesis, oxidation and organogenesis in vitro. J. Plant Physiol. 1993, 142, 543–551. [Google Scholar] [CrossRef]
- Xu, L. The effect of polyamine on flower bud differentiation and bud germination of chrysanthemum. Shandong Agric. Univ. 2015, 2, 31–36. [Google Scholar]
- Winter, G.; Todd, C.D.; Trovato, M.; Forlani, G.; Funck, D. Physiological implications of arginine metabolism in plants. Front. Plant Sci. 2015, 6, 534. [Google Scholar] [CrossRef]
- Kawade, K.; Horiguchi, G.; Hirose, Y.; Oikawa, A.; Hirai, M.Y.; Saito, K.; Fujita, T.; Tsukaya, H. Metabolic control of gametophore shoot formation through arginine in the moss Physcomitrium patens. Cell Rep. 2020, 32, 108–127. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolite | Stipes | Internodes |
|---|---|---|
| Carbohydrates | /-fold greater/ | |
| Starch | 1.9 | |
| Callose | 1.7 | |
| Mal | 75.0 | |
| GF2 | 11.0 | |
| Hormones | ||
| GA3 | 3.9 | |
| GA7 | 4.9 | |
| ABA | 3.1 | |
| ETH | 1.7 | |
| Others | ||
| Total phenolic acids | 2.7 | |
| ARG | 2.4 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mikuła, A.; Tomaszewicz, W.; Dziurka, M.; Kaźmierczak, A.; Grzyb, M.; Sobczak, M.; Zdańkowski, P.; Rybczyński, J. The Origin of the Cyathea delgadii Sternb. Somatic Embryos Is Determined by the Developmental State of Donor Tissue and Mutual Balance of Selected Metabolites. Cells 2021, 10, 1388. https://doi.org/10.3390/cells10061388
Mikuła A, Tomaszewicz W, Dziurka M, Kaźmierczak A, Grzyb M, Sobczak M, Zdańkowski P, Rybczyński J. The Origin of the Cyathea delgadii Sternb. Somatic Embryos Is Determined by the Developmental State of Donor Tissue and Mutual Balance of Selected Metabolites. Cells. 2021; 10(6):1388. https://doi.org/10.3390/cells10061388
Chicago/Turabian StyleMikuła, Anna, Wojciech Tomaszewicz, Michał Dziurka, Andrzej Kaźmierczak, Małgorzata Grzyb, Mirosław Sobczak, Piotr Zdańkowski, and Jan Rybczyński. 2021. "The Origin of the Cyathea delgadii Sternb. Somatic Embryos Is Determined by the Developmental State of Donor Tissue and Mutual Balance of Selected Metabolites" Cells 10, no. 6: 1388. https://doi.org/10.3390/cells10061388
APA StyleMikuła, A., Tomaszewicz, W., Dziurka, M., Kaźmierczak, A., Grzyb, M., Sobczak, M., Zdańkowski, P., & Rybczyński, J. (2021). The Origin of the Cyathea delgadii Sternb. Somatic Embryos Is Determined by the Developmental State of Donor Tissue and Mutual Balance of Selected Metabolites. Cells, 10(6), 1388. https://doi.org/10.3390/cells10061388