
Asthmatic Eosinophils Promote Contractility and Migration of Airway Smooth Muscle Cells and Pulmonary Fibroblasts In Vitro

 ,
,
Abstract
1. Introduction
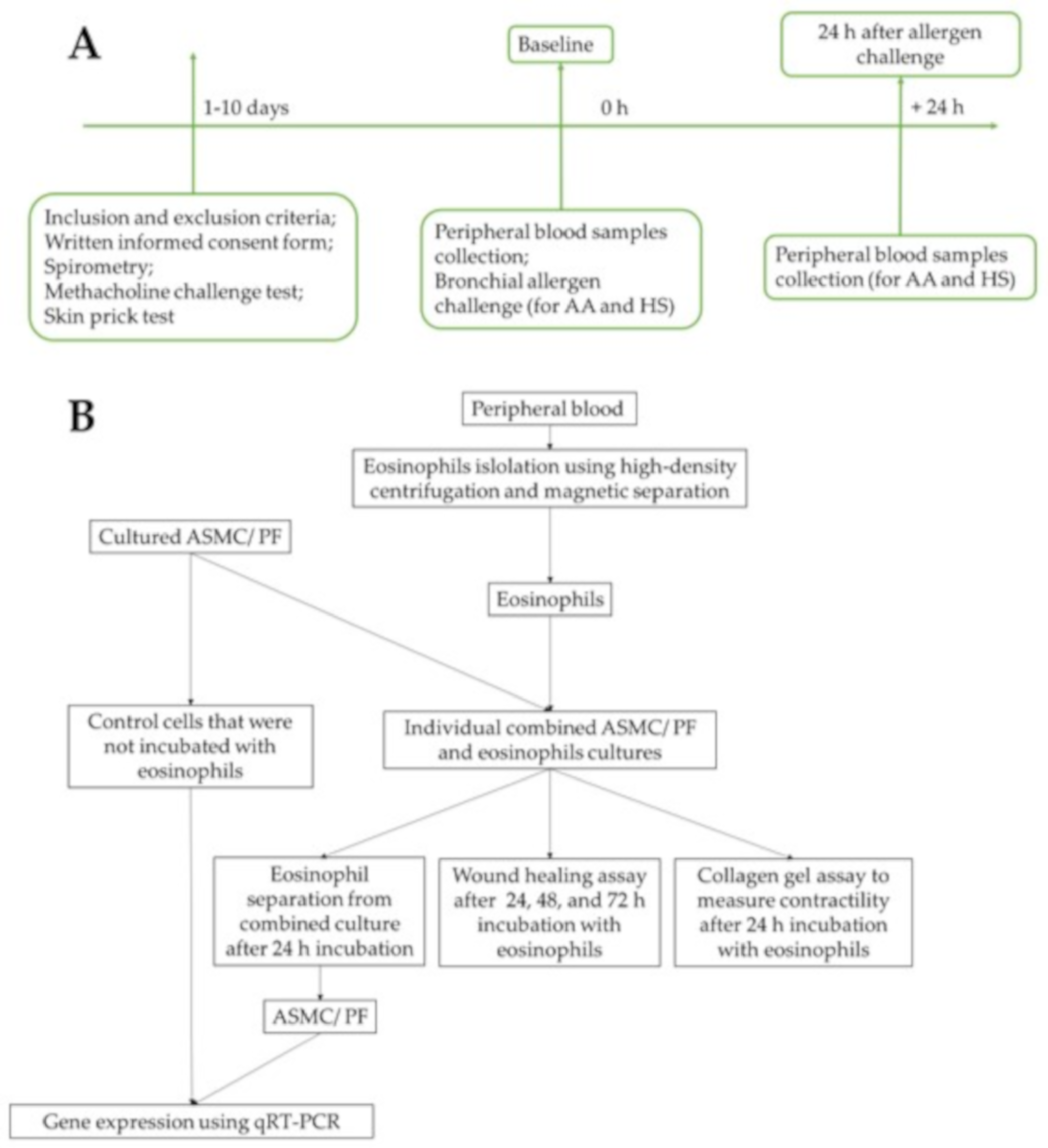
2. Materials and Methods
2.1. Study Subjects
2.2. Lung Function Testing
2.3. Measurement of Airway Responsiveness to Methacholine
2.4. Skin Prick Test
2.5. Bronchial Allergen Challenge
2.6. Isolation of Eosinophils from Peripheral Blood
2.7. Combined Cell Culture of Eosinophils and ASMC or PF
2.8. Culture Medium Treatment with Serum
2.9. Collagen Gel Assay
2.10. Wound Healing Assay
2.11. RNA Isolation and Quantitative Real-Time PCR Analysis
2.12. Statistical Analysis
3. Results
3.1. Demographic and Clinical Characteristics of the Study Population
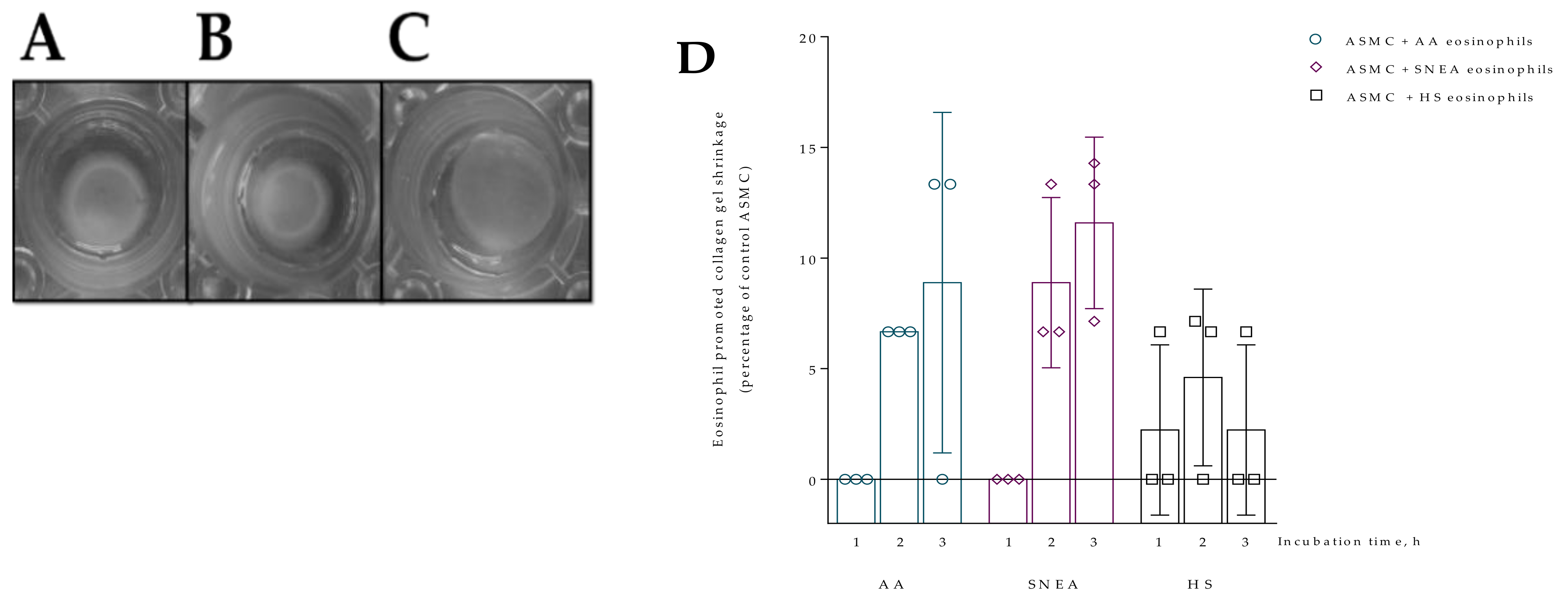
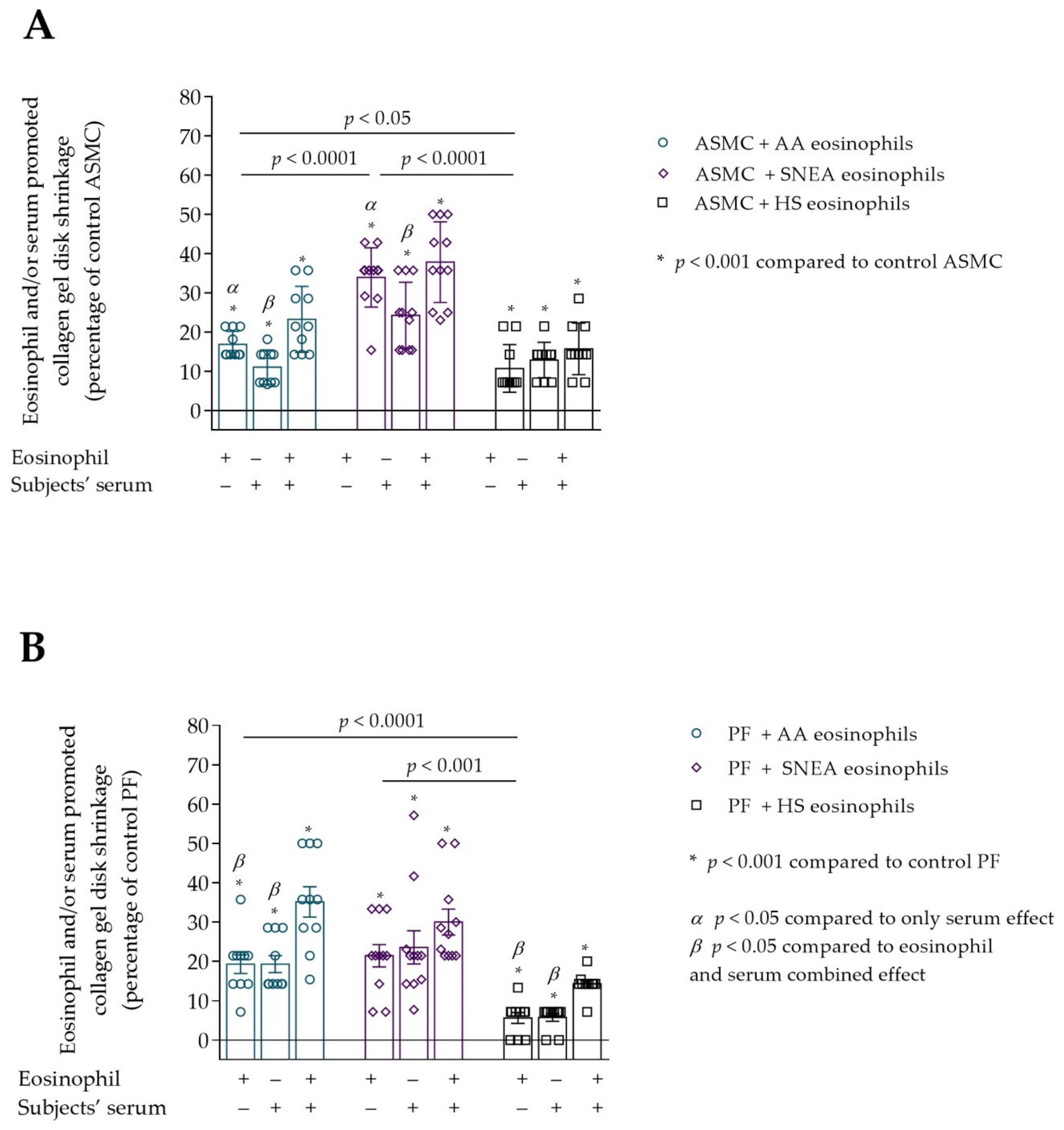
3.2. Contraction of Collagen Gel Disk after Incubation with Eosinophils
3.3. Migration of ASMC and PF after Incubation with Eosinophils
3.4. Gene Expression in ASMC and PF
3.5. The Effect of D. pteronyssinus Allergen Activated Eosinophils In Vivo on the Contractility of ASMC and PF
3.6. The Effect of D. pteronyssinus Allergen Activated Eosinophils In Vivo to the Migration of ASMC and PF
3.7. The Effect of Allergen-Activated Eosinophils on Gene Expression in ASMC and PF
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AA | Allergic asthma |
| ASMC | Airway smooth muscle cells |
| BMI | Body mass index |
| COL1A1 | Collagen I alpha 1 gene |
| COL5A1 | Collagen V alpha 1 gene |
| CTGF | Connective tissue growth factor |
| D. pteronyssinus | Dermatophagoides pteronyssinus |
| ECM | Extracellular matrix |
| FEV1 | Forced expiratory volume in 1 s |
| FN | Fibronectin gene |
| HS | Healthy subject |
| hTERT | Human telomerase reverse transcriptase |
| IgE | Immunoglobin E |
| IL | Interleukin |
| PD20M | Provocative dose of methacholine causing a 20% drop in FEV1 |
| PDGF | Platelet-derived growth factor |
| PF | Pulmonary fibroblasts |
| SD | Standard deviation |
| SEM | Standard error of mean |
| SM22 | Transgelin |
| sm-MHC | Smooth muscle myosin heavy chain |
| sm-MLCK | Smooth muscle myosin light chain kinase |
| SNEA | Severe non-allergic eosinophilic asthma |
| TGF-β | Transforming growth factor β |
| TSLP | Thymic stromal lymphopoietin |
References
- Enilari, O.; Sinha, S. The global impact of asthma in adult populations. Ann. Glob. Health 2019, 85, 2412. [Google Scholar] [CrossRef] [PubMed]
- Chan, V.; Burgess, J.K.; Ratoff, J.C.; O’Connor, B.J.; Greenough, A.; Lee, T.H.; Hirst, S.J. Extracellular matrix regulates enhanced eotaxin expression in asthmatic airway smooth muscle cells. Am. J. Respir. Crit. Care Med. 2006, 174, 379–385. [Google Scholar] [CrossRef]
- Johnson, P.R.; Roth, M.; Tamm, M.; Hughes, M.; Ge, Q.; King, G.; Burgess, J.K.; Black, J.L. Airway smooth muscle cell proliferation is increased in asthma. Am. J. Respir. Crit. Care Med. 2001, 164, 474–477. [Google Scholar] [CrossRef]
- Burgess, J.K.; Johnson, P.R.; Ge, Q.; Au, W.W.; Poniris, M.H.; McParland, B.E.; King, G.; Roth, M.; Black, J.L. Expression of connective tissue growth factor in asthmatic airway smooth muscle cells. Am. J. Respir. Crit. Care Med. 2003, 167, 71–77. [Google Scholar] [CrossRef]
- Jiang, H.; Rao, K.; Halayko, A.J.; Kepron, W.; Stephens, N.L. Bronchial smooth muscle mechanics of a canine model of allergic airway hyperresponsiveness. J. Appl. Physiol. 1992, 72, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Cheng, Z.; Kong, H.; Wang, Y.; Unruh, H.; Stephens, N.L.; Laviolette, M. Changes in biophysical and biochemical properties of single bronchial smooth muscle cells from asthmatic subjects. Am. J. Physiol. Lung Cell. Mol. Physiol. 2002, 283, L1181–L1189. [Google Scholar] [CrossRef]
- Ammit, A.J.; Armour, C.L.; Black, J.L. Smooth-muscle myosin light-chain kinase content is increased in human sensitized airways. Am. J. Respir. Crit. Care Med. 2000, 161, 257–263. [Google Scholar] [CrossRef]
- An, S.; Bai, T.; Bates, J.; Black, J.; Brown, R.H.; Brusasco, V.; Chitano, P.; Deng, L.; Dowell, M.; Eidelman, D. Airway smooth muscle dynamics: A common pathway of airway obstruction in asthma. Eur. Respir. J. 2007, 29, 834–860. [Google Scholar] [CrossRef] [PubMed]
- Salter, B.; Pray, C.; Radford, K.; Martin, J.G.; Nair, P. Regulation of human airway smooth muscle cell migration and relevance to asthma. Respir. Res. 2017, 18, 156. [Google Scholar] [CrossRef] [PubMed]
- Baum, J.; Duffy, H.S. Fibroblasts and myofibroblasts: What are we talking about? J. Cardiovasc. Pharm. 2011, 57, 376–379. [Google Scholar] [CrossRef]
- Hinz, B.; Phan, S.H.; Thannickal, V.J.; Galli, A.; Bochaton-Piallat, M.-L.; Gabbiani, G. The myofibroblast: One function, multiple origins. Am. J. Pathol. 2007, 170, 1807–1816. [Google Scholar] [CrossRef]
- Kendall, R.T.; Feghali-Bostwick, C.A. Fibroblasts in fibrosis: Novel roles and mediators. Front. Pharm. 2014, 5, 123. [Google Scholar] [CrossRef] [PubMed]
- Manuyakorn, W. Airway remodelling in asthma: Role for mechanical forces. Asia Pac. Allergy 2014, 4, 19–24. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ozier, A.; Allard, B.; Bara, I.; Girodet, P.-O.; Trian, T.; Marthan, R.; Berger, P. The pivotal role of airway smooth muscle in asthma pathophysiology. J. Allergy 2011, 2011, 742710. [Google Scholar] [CrossRef] [PubMed]
- Carr, T.F.; Berdnikovs, S.; Simon, H.-U.; Bochner, B.S.; Rosenwasser, L.J. Eosinophilic bioactivities in severe asthma. World Allergy Organ. J. 2016, 9, 21. [Google Scholar] [CrossRef]
- Januskevicius, A.; Janulaityte, I.; Kalinauskaite-Zukauske, V.; Gosens, R.; Malakauskas, K. The Enhanced Adhesion of Eosinophils Is Associated with Their Prolonged Viability and Pro-Proliferative Effect in Asthma. J. Clin. Med. 2019, 8, 1274. [Google Scholar] [CrossRef]
- Gosens, R.; Stelmack, G.L.; Dueck, G.; McNeill, K.D.; Yamasaki, A.; Gerthoffer, W.T.; Unruh, H.; Gounni, A.S.; Zaagsma, J.; Halayko, A.J. Role of caveolin-1 in p42/p44 MAP kinase activation and proliferation of human airway smooth muscle. Am. J. Physiol. Lung Cell. Mol. Physiol. 2006, 291, L523–L534. [Google Scholar] [CrossRef] [PubMed]
- Ngo, P.; Ramalingam, P.; Phillips, J.A.; Furuta, G.T. Collagen gel contraction assay. Cell Cell Interact. 2006, 341, 103–110. [Google Scholar]
- Liang, C.-C.; Park, A.Y.; Guan, J.-L. In vitro scratch assay: A convenient and inexpensive method for analysis of cell migration in vitro. Nat. Protoc. 2007, 2, 329–333. [Google Scholar] [CrossRef]
- Walford, H.H.; Doherty, T.A. Diagnosis and management of eosinophilic asthma: A US perspective. J. Asthma Allergy 2014, 7, 53–65. [Google Scholar] [CrossRef]
- Bousquet, J.; Chanez, P.; Lacoste, J.Y.; Barnéon, G.; Ghavanian, N.; Enander, I.; Venge, P.; Ahlstedt, S.; Simony-Lafontaine, J.; Godard, P.; et al. Eosinophilic inflammation in asthma. N. Engl. J. Med. 1990, 323, 1033–1039. [Google Scholar] [CrossRef]
- Busse, W.W.; Sedgwick, J.B. Eosinophils in asthma. Ann. Allergy 1992, 68, 286–290. [Google Scholar] [PubMed]
- Hough, K.P.; Curtiss, M.L.; Blain, T.J.; Liu, R.-M.; Trevor, J.; Deshane, J.S.; Thannickal, V.J. Airway Remodeling in Asthma. Front. Med. 2020, 7, 191. [Google Scholar] [CrossRef] [PubMed]
- Prakash, Y.S. Airway smooth muscle in airway reactivity and remodeling: What have we learned? Am. J. Physiol. Lung Cell. Mol. Physiol. 2013, 305, L912–L933. [Google Scholar] [CrossRef]
- Zuyderduyn, S.; Sukkar, M.B.; Fust, A.; Dhaliwal, S.; Burgess, J.K. Treating asthma means treating airway smooth muscle cells. Eur. Respir. J. 2008, 32, 265–274. [Google Scholar] [CrossRef] [PubMed]
- Kalinauskaite-Zukauske, V.; Januskevicius, A.; Janulaityte, I.; Miliauskas, S.; Malakauskas, K. Expression of eosinophil β chain-signaling cytokines receptors, outer-membrane integrins, and type 2 inflammation biomarkers in severe non-allergic eosinophilic asthma. BMC Pulm. Med. 2019, 19, 158. [Google Scholar] [CrossRef] [PubMed]
- Bakakos, A.; Loukides, S.; Bakakos, P. Severe Eosinophilic Asthma. J. Clin. Med. 2019, 8, 1375. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.K.; Cheung, P.F.Y.; Ip, W.K.; Lam, C.W.K. Intracellular Signaling Mechanisms Regulating Toll-Like Receptor–Mediated Activation of Eosinophils. Am. J. Respir. Cell Mol. Biol. 2007, 37, 85–96. [Google Scholar] [CrossRef]
- Webb, R.C. Smooth muscle contraction and relaxation. Adv. Physiol. Educ. 2003, 27, 201–206. [Google Scholar] [CrossRef]
- Doeing, D.C.; Solway, J. Airway smooth muscle in the pathophysiology and treatment of asthma. J. Appl. Physiol. 2013, 114, 834–843. [Google Scholar] [CrossRef]
- Shaifta, Y.; MacKay, C.E.; Irechukwu, N.; O’Brien, K.A.; Wright, D.B.; Ward, J.P.T. Transforming growth factor-β enhances Rho-kinase activity and contraction in airway smooth muscle via the nucleotide exchange factor ARHGEF1. J. Physiol. 2018, 596, 47–66. [Google Scholar] [CrossRef] [PubMed]
- Michalik, M.; Wójcik-Pszczoła, K.; Paw, M.; Wnuk, D.; Koczurkiewicz, P.; Sanak, M.; Pękala, E.; Madeja, Z. Fibroblast-to-myofibroblast transition in bronchial asthma. Cell Mol. Life Sci. 2018, 75, 3943–3961. [Google Scholar] [CrossRef] [PubMed]
- Hinz, B.; Dugina, V.; Ballestrem, C.; Wehrle-Haller, B.; Chaponnier, C. Alpha-smooth muscle actin is crucial for focal adhesion maturation in myofibroblasts. Mol. Biol. Cell 2003, 14, 2508–2519. [Google Scholar] [CrossRef]
- Kohan, M.; Muro, A.F.; White, E.S.; Berkman, N. EDA-containing cellular fibronectin induces fibroblast differentiation through binding to alpha4beta7 integrin receptor and MAPK/Erk 1/2-dependent signaling. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2010, 24, 4503–4512. [Google Scholar] [CrossRef]
- Altman, L.C.; Hill, J.S.; Hairfield, W.M.; Mullarkey, M.F. Effects of corticosteroids on eosinophil chemotaxis and adherence. J. Clin. Investig. 1981, 67, 28–36. [Google Scholar] [CrossRef]
- Kato, M.; Schleimer, R.P. Antiinflammatory steroids inhibit granulocyte/macrophage colony-stimulating factor production by human lung tissue. Lung 1994, 172, 113–124. [Google Scholar] [CrossRef]
- Tobler, A.; Meier, R.; Seitz, M.; Dewald, B.; Baggiolini, M.; Fey, M.F. Glucocorticoids downregulate gene expression of GM-CSF, NAP-1/IL-8, and IL-6, but not of M-CSF in human fibroblasts. Blood 1992, 79, 45–51. [Google Scholar] [CrossRef]
- Rolfe, F.G.; Hughes, J.M.; Armour, C.L.; Sewell, W.A. Inhibition of interleukin-5 gene expression by dexamethasone. Immunology 1992, 77, 494–499. [Google Scholar]
- Zhang, T.; Day, J.H.; Su, X.; Guadarrama, A.G.; Sandbo, N.K.; Esnault, S.; Denlinger, L.C.; Berthier, E.; Theberge, A.B. Investigating Fibroblast-Induced Collagen Gel Contraction Using a Dynamic Microscale Platform. Front. Bioeng. Biotechnol. 2019, 7, 196. [Google Scholar] [CrossRef] [PubMed]
- Brown, R.H.; Mitzner, W.; Bulut, Y.; Wagner, E.M. Effect of lung inflation in vivo on airways with smooth muscle tone or edema. J. Appl. Physiol. 1997, 82, 491–499. [Google Scholar] [CrossRef][Green Version]
- Dunnill, M.; Massarella, G.; Anderson, J. A comparison of the quantitative anatomy of the bronchi in normal subjects, in status asthmaticus, in chronic bronchitis, and in emphysema. Thorax 1969, 24, 176–179. [Google Scholar] [CrossRef] [PubMed]
- Raeburn, D.; Webber, S. Proinflammatory potential of the airway epithelium in bronchial asthma. Eur. Respir. J. 1994, 7, 2226–2233. [Google Scholar] [CrossRef] [PubMed]
- Jeffery, P.K. Remodeling in asthma and chronic obstructive lung disease. Am. J. Respir. Crit. Care Med. 2001, 164, S28–S38. [Google Scholar] [CrossRef] [PubMed]
- Bousquet, J.; Jeffery, P.K.; Busse, W.W.; Johnson, M.; Vignola, A.M. Asthma. From bronchoconstriction to airways inflammation and remodeling. Am. J. Respir. Crit. Care Med. 2000, 161, 1720–1745. [Google Scholar] [CrossRef]
- Bousquet, J.; Lacoste, J.Y.; Chanez, P.; Vic, P.; Godard, P.; Michel, F.B. Bronchial elastic fibers in normal subjects and asthmatic patients. Am. J. Respir. Crit. Care Med. 1996, 153, 1648–1654. [Google Scholar] [CrossRef]
- Carroll, N.G.; Perry, S.; Karkhanis, A.; Harji, S.; Butt, J.; James, A.L.; Green, F.H.Y. The Airway Longitudinal Elastic Fiber Network and Mucosal Folding in Patients with Asthma. Am. J. Respir. Crit. Care Med. 2000, 161, 244–248. [Google Scholar] [CrossRef] [PubMed]
- Pepe, C.; Foley, S.; Shannon, J.; Lemiere, C.; Olivenstein, R.; Ernst, P.; Ludwig, M.S.; Martin, J.G.; Hamid, Q. Differences in airway remodeling between subjects with severe and moderate asthma. J. Allergy Clin. Immunol. 2005, 116, 544–549. [Google Scholar] [CrossRef]
- Cockcroft, D.W.; Davis, B.E. Mechanisms of airway hyperresponsiveness. J. Allergy Clin. Immunol. 2006, 118, 551–559. [Google Scholar] [CrossRef]
- Sapienza, S.; Du, T.; Eidelman, D.H.; Wang, N.S.; Martin, J.G. Structural Changes in the Airways of Sensitized Brown Norway Rats after Antigen Challenge. Am. Rev. Respir. Dis. 1991, 144, 423–427. [Google Scholar] [CrossRef]
- Aszodi, A.; Legate, K.R.; Nakchbandi, I.; Fässler, R. What mouse mutants teach us about extracellular matrix function. Annu. Rev. Cell Dev. Biol. 2006, 22, 591–621. [Google Scholar] [CrossRef]
- Yamauchi, E.; Shoji, S.; Nishihara, M.; Shimoda, T.; Nishima, S. Contribution of lung fibroblast migration in the fibrotic process of airway remodeling in asthma. Allergol. Int. Off. J. Jpn. Soc. Allergol. 2008, 57, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Muniz, V.S.; Weller, P.F.; Neves, J.S. Eosinophil crystalloid granules: Structure, function, and beyond. J. Leukoc. Biol. 2012, 92, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Zagai, U.; Lundahl, J.; Klominek, J.; Venge, P.; Sköld, C.M. Eosinophil Cationic Protein Stimulates Migration of Human Lung Fibroblasts In Vitro. Scand. J. Immunol. 2009, 69, 381–386. [Google Scholar] [CrossRef] [PubMed]
- Erjefält, J.S.; Greiff, L.; Andersson, M.; Ädelroth, E.; Jeffery, P.K.; Persson, C.G.A. Degranulation patterns of eosinophil granulocytes as determinants of eosinophil driven disease. Thorax 2001, 56, 341–344. [Google Scholar] [CrossRef] [PubMed]
- McBrien, C.N.; Menzies-Gow, A. The Biology of Eosinophils and Their Role in Asthma. Front. Med. 2017, 4, 93. [Google Scholar] [CrossRef]
- Saito, K.; Nagata, M.; Kikuchi, I.; Sakamoto, Y. Leukotriene D4 and eosinophil transendothelial migration, superoxide generation, and degranulation via β2 integrin. Ann. Allergy Asthma Immunol. 2004, 93, 594–600. [Google Scholar] [CrossRef]
- Sukkar, M.B.; Stanley, A.J.; Blake, A.E.; Hodgkin, P.D.; Johnson, P.R.; Armour, C.L.; Hughes, J.M. ‘Proliferative’ and ‘synthetic’ airway smooth muscle cells are overlapping populations. Immunol. Cell Biol. 2004, 82, 471–478. [Google Scholar] [CrossRef]
- Wright, D.B.; Trian, T.; Siddiqui, S.; Pascoe, C.D.; Johnson, J.R.; Dekkers, B.G.J.; Dakshinamurti, S.; Bagchi, R.; Burgess, J.K.; Kanabar, V.; et al. Phenotype modulation of airway smooth muscle in asthma. Pulm. Pharmacol. Ther. 2013, 26, 42–49. [Google Scholar] [CrossRef]
- Ma, X.; Wang, Y.; Stephens, N.L. Serum deprivation induces a unique hypercontractile phenotype of cultured smooth muscle cells. Am. J. Physiol. 1998, 274, C1206–C1214. [Google Scholar] [CrossRef]
- Halayko, A.J.; Camoretti-Mercado, B.; Forsythe, S.M.; Vieira, J.E.; Mitchell, R.W.; Wylam, M.E.; Hershenson, M.B.; Solway, J. Divergent differentiation paths in airway smooth muscle culture: Induction of functionally contractile myocytes. Am. J. Physiol. 1999, 276, L197–L206. [Google Scholar] [CrossRef]
- Halayko, A.J.; Salari, H.; Ma, X.; Stephens, N.L. Markers of airway smooth muscle cell phenotype. Am. J. Physiol. 1996, 270, L1040–L1051. [Google Scholar] [CrossRef]
- Johnson, P.R.A.; Black, J.L.; Carlin, S.; Ge, Q.; Underwood, P.A. The Production of Extracellular Matrix Proteins by Human Passively Sensitized Airway Smooth-Muscle Cells in Culture. Am. J. Respir. Crit. Care Med. 2000, 162, 2145–2151. [Google Scholar] [CrossRef] [PubMed][Green Version]
- White, E.S. Lung extracellular matrix and fibroblast function. Ann. Am. Thorac. Soc. 2015, 12, S30–S33. [Google Scholar] [CrossRef] [PubMed]
- Al-Alawi, M.; Hassan, T.; Chotirmall, S.H. Transforming growth factor β and severe asthma: A perfect storm. Respir. Med. 2014, 108, 1409–1423. [Google Scholar] [CrossRef] [PubMed]
- Scharenberg, M.A.; Pippenger, B.E.; Sack, R.; Zingg, D.; Ferralli, J.; Schenk, S.; Martin, I.; Chiquet-Ehrismann, R. TGF-β-induced differentiation into myofibroblasts involves specific regulation of two MKL1 isoforms. J. Cell Sci. 2014, 127, 1079–1091. [Google Scholar] [CrossRef]
- Desmouliere, A.; Geinoz, A.; Gabbiani, F.; Gabbiani, G. Transforming growth factor-beta 1 induces alpha-smooth muscle actin expression in granulation tissue myofibroblasts and in quiescent and growing cultured fibroblasts. J. Cell Biol. 1993, 122, 103–111. [Google Scholar] [CrossRef]
- Akamatsu, T.; Arai, Y.; Kosugi, I.; Kawasaki, H.; Meguro, S.; Sakao, M.; Shibata, K.; Suda, T.; Chida, K.; Iwashita, T. Direct isolation of myofibroblasts and fibroblasts from bleomycin-injured lungs reveals their functional similarities and differences. Fibrogen. Tissue Repair 2013, 6, 15. [Google Scholar] [CrossRef]
- Tomasek, J.J.; Gabbiani, G.; Hinz, B.; Chaponnier, C.; Brown, R.A. Myofibroblasts and mechano-regulation of connective tissue remodelling. Nat. Rev. Mol. Cell Biol. 2002, 3, 349–363. [Google Scholar] [CrossRef]
- Sapudom, J.; Rubner, S.; Martin, S.; Thoenes, S.; Anderegg, U.; Pompe, T. The interplay of fibronectin functionalization and TGF-β1 presence on fibroblast proliferation, differentiation and migration in 3D matrices. Biomater. Sci. 2015, 3, 1291–1301. [Google Scholar] [CrossRef]
- Brenmoehl, J.; Miller, S.-N.; Hofmann, C.; Vogl, D.; Falk, W.; Schölmerich, J.; Rogler, G. Transforming growth factor-beta 1 induces intestinal myofibroblast differentiation and modulates their migration. World J. Gastroenterol. 2009, 15, 1431–1442. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Weitoft, M.; Andersson, C.; Andersson-Sjöland, A.; Tufvesson, E.; Bjermer, L.; Erjefält, J.; Westergren-Thorsson, G. Controlled and uncontrolled asthma display distinct alveolar tissue matrix compositions. Respir. Res. 2014, 15, 67. [Google Scholar] [CrossRef] [PubMed]
- Wenstrup, R.J.; Florer, J.B.; Brunskill, E.W.; Bell, S.M.; Chervoneva, I.; Birk, D.E. Type V collagen controls the initiation of collagen fibril assembly. J. Biol. Chem. 2004, 279, 53331–53337. [Google Scholar] [CrossRef]
- Kalinauskaite-Zukauske, V.; Januskevicius, A.; Janulaityte, I.; Miliauskas, S.; Malakauskas, K. Serum Levels of Epithelial-Derived Cytokines as Interleukin-25 and Thymic Stromal Lymphopoietin after a Single Dose of Mepolizumab in Patients with Severe Non-Allergic Eosinophilic Asthma: A Short Report. Can. Respir. J. 2019, 2019, 8607657. [Google Scholar] [CrossRef]
- Sanz, M.L.; Parra, A.; Prieto, I.; Diéguez, I.; Oehling, A.K. Serum eosinophil peroxidase (EPO) levels in asthmatic patients. Allergy 1997, 52, 417–422. [Google Scholar] [CrossRef]
- Azazi, E.A.; Elshora, A.E.; Tantawy, E.A.; Elsayd, M.A. Serum levels of Interleukin-33 and its soluble receptor ST2 in asthmatic patients. Egypt. J. Chest Dis. Tuberc. 2014, 63, 279–284. [Google Scholar] [CrossRef]
- Hardy, E.; Farahani, M.; Hall, I.P. Regulation of histamine H1 receptor coupling by dexamethasone in human cultured airway smooth muscle. Br. J. Pharmacol. 1996, 118, 1079–1084. [Google Scholar] [CrossRef]
- Tanaka, H.; Watanabe, K.; Tamaru, N.; Yoshida, M. Arachidonic acid metabolites and glucocorticoid regulatory mechanism in cultured porcine tracheal smooth muscle cells. Lung 1995, 173, 347–361. [Google Scholar] [CrossRef]
- Miller, M.; Cho, J.Y.; McElwain, K.; McElwain, S.; Shim, J.Y.; Manni, M.; Baek, J.S.; Broide, D.H. Corticosteroids prevent myofibroblast accumulation and airway remodeling in mice. Am. J. Physiol. Lung Cell. Mol. Physiol. 2006, 290, L162–L169. [Google Scholar] [CrossRef]
- Ihrie, M.D.; Ingram, J.L. Orchestrating Airway Smooth Muscle Cell Migration: GMFγ Phosphorylation Is the Key. Am. J. Respir. Cell Mol. Biol. 2019, 61, 136–138. [Google Scholar] [CrossRef] [PubMed]
- Johnson, P.R.A.; Burgess, J.K. Airway smooth muscle and fibroblasts in the pathogenesis of asthma. Curr. Allergy Asthma Rep. 2004, 4, 102–108. [Google Scholar] [CrossRef]
- Redhu, N.S.; Shan, L.; Movassagh, H.; Gounni, A.S. Thymic stromal lymphopoietin induces migration in human airway smooth muscle cells. Sci. Rep. 2013, 3, 2301. [Google Scholar] [CrossRef]
- Melzer, C.; von der Ohe, J.; Hass, R.; Ungefroren, H. TGF-β-Dependent Growth Arrest and Cell Migration in Benign and Malignant Breast Epithelial Cells Are Antagonistically Controlled by Rac1 and Rac1b. Int. J. Mol. Sci. 2017, 18, 1574. [Google Scholar] [CrossRef] [PubMed]
- Janulaityte, I.; Januskevicius, A.; Kalinauskaite-Zukauske, V.; Bajoriuniene, I.; Malakauskas, K. In Vivo Allergen-Activated Eosinophils Promote Collagen I and Fibronectin Gene Expression in Airway Smooth Muscle Cells via TGF-β1 Signaling Pathway in Asthma. Int. J. Mol. Sci. 2020, 21, 1837. [Google Scholar] [CrossRef]
- Schaafsma, D.; McNeill, K.D.; Stelmack, G.L.; Gosens, R.; Baarsma, H.A.; Dekkers, B.G.J.; Frohwerk, E.; Penninks, J.-M.; Sharma, P.; Ens, K.M.; et al. Insulin increases the expression of contractile phenotypic markers in airway smooth muscle. Am. J. Physiol. Cell Physiol. 2007, 293, C429–C439. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Bodas, M.; Bhatraju, N.K.; Pattnaik, B.; Gheware, A.; Parameswaran, P.K.; Thompson, M.; Freeman, M.; Mabalirajan, U.; Gosens, R.; et al. Hyperinsulinemia adversely affects lung structure and function. Am. J. Physiol. Lung Cell. Mol. Physiol. 2016, 310, L837–L845. [Google Scholar] [CrossRef] [PubMed]
- McMillan, S.J.; Xanthou, G.; Lloyd, C.M. Manipulation of allergen-induced airway remodeling by treatment with anti-TGF-beta antibody: Effect on the Smad signaling pathway. J. Immunol. 2005, 174, 5774–5780. [Google Scholar] [CrossRef] [PubMed]
- Kalinauskaite-Zukauske, V.; Janulaityte, I.; Januskevicius, A.; Malakauskas, K. Serum levels of epithelial-derived mediators and interleukin-4/interleukin-13 signaling after bronchial challenge with Dermatophagoides pteronyssinus in patients with allergic asthma. Scand. J. Immunol. 2019, 90, e12820. [Google Scholar] [CrossRef] [PubMed]
- Schaafsma, D.; Gosens, R.; Bos, I.S.T.; Meurs, H.; Zaagsma, J.; Nelemans, S.A. Allergic sensitization enhances the contribution of Rho-kinase to airway smooth muscle contraction. Br. J. Pharmacol. 2004, 143, 477–484. [Google Scholar] [CrossRef] [PubMed]
- Gizycki, M.J.; Adelroth, E.; Rogers, A.V.; O’Byrne, P.M.; Jeffery, P.K. Myofibroblast involvement in the allergen-induced late response in mild atopic asthma. Am. J. Respir. Cell Mol. Biol. 1997, 16, 664–673. [Google Scholar] [CrossRef]
- Sakota, Y.; Ozawa, Y.; Yamashita, H.; Tanaka, H.; Inagaki, N. Collagen gel contraction assay using human bronchial smooth muscle cells and its application for evaluation of inhibitory effect of formoterol. Biol. Pharm. Bull. 2014, 37, 1014–1020. [Google Scholar] [CrossRef] [PubMed]
- Bortolozzo, A.S.S.; Rodrigues, A.P.D.; Arantes-Costa, F.M.; Saraiva-Romanholo, B.M.; De Souza, F.C.R.; Brüggemann, T.R.; De Brito, M.V.; Ferreira, R.D.S.; Correia, M.T.D.S.; Paiva, P.M.G.; et al. The Plant Proteinase Inhibitor CrataBL Plays a Role in Controlling Asthma Response in Mice. BioMed Res. Int. 2018, 2018, 9274817. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.G.; Zhu, S.Y.; Heng, B.C.; Dissanayaka, W.L.; Zhang, C.F. TGF-β1-induced differentiation of SHED into functional smooth muscle cells. Stem Cell Res. Ther. 2017, 8, 10. [Google Scholar] [CrossRef] [PubMed]
- Serban, A.I.; Stanca, L.; Geicu, O.I.; Munteanu, M.C.; Dinischiotu, A. RAGE and TGF-β1 cross-talk regulate extracellular matrix turnover and cytokine synthesis in AGEs exposed fibroblast cells. PLoS ONE 2016, 11, e0152376. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AA Patients (n = 13) | SNEA Patients (n = 11) | HS (n = 14) | |
|---|---|---|---|
| Screening visit (for all groups) |
| ||
| Inclusion criteria |
|
|
|
| Exclusion criteria (for all groups) |
| ||
| Gene | Forward 5′–3′ | Reverse 5′–3′ |
|---|---|---|
| 18S | CGCCGCTAGAGGTGAAATTC | TTGGCAAATGCTTTCGCTC |
| α-sm-actin | TGGGTGACGAAGCAC AGAGC | CTTCAGGGGCAACACGAAGC |
| sm-MHC | CGCCAAGAGACTCGTCTGG | TCTTTCCCAACCGTGACCTTC |
| SM22 | AGGAGCGGCTGGTGGAGTGGAT | CATGTCAGTCTTGATGACCCCATAGT |
| sm-MLCK | GACTGCAAGATTGAAGGATAC | GTTTCCACAATGAGCTCTGC |
| COL1A1 | TCGAGGAGGAAATTCCAATG | ACACACGTGCACCTCATCAT |
| COL5A1 | GGCTCCCGAGAGCAACCT | CGGGACACTCACGAACGAA |
| FN | AGCCAGCAGATCGAGAACAT | TCTTGTCCTTGGGGTTCTTG |
| AA Patients, n = 13 | SNEA Patients, n = 11 | HS, n = 14 | |
|---|---|---|---|
| Age, median (range), years | 25.8 (18.0–40.0) | 54.0 (28.0–80.0) * # | 31.5 (23.0–59.0) |
| Sex, (male/female), n | 7/5 | 6/5 | 4/10 |
| BMI, kg/m2, median (range) | 23.8 (17.3–40.1) | 27.5 (17.5–37.3) | 23.9 (17.0–34.4) |
| Sensitization to D. pteronyssinus/D. farinae/birch/5 grass mixture allergen, n | 13/9/4/3 | NR | NR |
| Wheel diameter by D. pteronyssinus, median (range), mm | 8.4 (4.0–15.0) | 0 | 0 |
| PD20M, geometric mean (range), mg | 0.16 (0.05–0.41) | ND | NR |
| FEV1, % of predicted | 85.2 ± 11.5 | 51.3 ± 26.4 * # | 107.0 ± 11.6 |
| FEV1, L | 3.6 ± 0.8 | 1.7 ± 1.3 * # | 3.8 ± 0.6 |
| Blood eosinophil count, × 109/L | 0.40 ± 0.24 * | 0.63 ± 0.55 * # | 0.19 ± 0.09 |
| Blood eosinophil count, % | 6.17 ± 3.97 * | 10.30 ± 8.55 * # | 2.92 ± 1.10 |
| IgE, IU/mL | 583.0 (88.9–4617.0) * | 196.0 (11.2–795.0) * # | 26.2 (3.0–67.4) |
| AA Patients, n = 13 | HS, n = 14 | |||
|---|---|---|---|---|
| Before the allergen challenge | 24 h after allergen challenge | Before the allergen challenge | 24 h after allergen challenge | |
| Blood eosinophil count, × 109/L | 0.40 ± 0.24 | 0.47 ± 0.21 * | 0.19 ± 0.09 | 0.14 ± 0.05 |
| Blood eosinophil count, % | 6.17 ± 3.97 | 7.08 ± 3.77 * | 2.92 ± 1.10 | 3.07 ± 1.81 |
| IgE, IU/mL | 583 (88.9–4617.0) | 837 (95.1–4325) * | 26.2 (3.0–67.4) | 27.6 (3.0–71.2) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Janulaityte, I.; Januskevicius, A.; Kalinauskaite-Zukauske, V.; Palacionyte, J.; Malakauskas, K. Asthmatic Eosinophils Promote Contractility and Migration of Airway Smooth Muscle Cells and Pulmonary Fibroblasts In Vitro. Cells 2021, 10, 1389. https://doi.org/10.3390/cells10061389
Janulaityte I, Januskevicius A, Kalinauskaite-Zukauske V, Palacionyte J, Malakauskas K. Asthmatic Eosinophils Promote Contractility and Migration of Airway Smooth Muscle Cells and Pulmonary Fibroblasts In Vitro. Cells. 2021; 10(6):1389. https://doi.org/10.3390/cells10061389
Chicago/Turabian StyleJanulaityte, Ieva, Andrius Januskevicius, Virginija Kalinauskaite-Zukauske, Jolita Palacionyte, and Kestutis Malakauskas. 2021. "Asthmatic Eosinophils Promote Contractility and Migration of Airway Smooth Muscle Cells and Pulmonary Fibroblasts In Vitro" Cells 10, no. 6: 1389. https://doi.org/10.3390/cells10061389
APA StyleJanulaityte, I., Januskevicius, A., Kalinauskaite-Zukauske, V., Palacionyte, J., & Malakauskas, K. (2021). Asthmatic Eosinophils Promote Contractility and Migration of Airway Smooth Muscle Cells and Pulmonary Fibroblasts In Vitro. Cells, 10(6), 1389. https://doi.org/10.3390/cells10061389